
Branch Wood Decomposition of Tree Species in a Deciduous Temperate Forest in Korea


Department of Life Science, Chung-Ang University, Seoul 06974, Korea
*
Author to whom correspondence should be addressed.
Forests 2017, 8(5), 176; https://doi.org/10.3390/f8050176
Submission received: 11 April 2017
/
Revised: 11 May 2017
/
Accepted: 12 May 2017
/
Published: 19 May 2017
(This article belongs to the Special Issue Coarse Woody Debris of Forests in a Changing World)
Abstract
:Woody debris, which is supplied by branch litter, is an important component of forest ecosystems as it contains large quantities of organic matter and nutrients. We evaluated changes in branch wood dry weight and nutrient content of six common species (Fraxinus rhynchophylla, Pinus densiflora, Prunus sargentii, Quercus mongolica, Acer pseudosieboldianum, and Symplocos chinensis for. pilosa) in a deciduous temperate forest in Korea for 40 months. Branch wood disk samples 1.4–1.6 cm thick were cut, and mass loss was measured over time using the litterbag method. No significant differences in mass loss were recorded among the six tree species. Further, mass loss was negatively correlated with initial lignin concentration and positively correlated with both initial cellulose concentration and wood density for each species. Species with high wood cellulose content had high wood density while the lignin content in wood was relatively low. Accordingly, cellulose contributed to wood density, creating a relatively lower lignin content, and the decreased lignin concentration increased the wood decomposition rate.
1. Introduction
The balance between primary production and decomposition in the forest ecosystem is important for maintaining sustainable ecosystems. Woody debris on the forest floor is an important component of forest ecosystems [1,2]. Woody materials are a major part of net primary production and biomass and, therefore, are considered nutrient reservoirs, particularly for carbon (C) [1,3,4]. Bray and Gorham [5] conducted a general survey of global litter production, including non-leaf litter, and they found that litter production by deciduous trees in the northern hemisphere totaled 3.2 metric tons ha−1 year−1 and that non-leaf litter production in cool temperate and warm temperate zones equaled 0.9 metric tons ha−1 year−1 and 1.9 metric tons ha−1 year−1, respectively. Further, they found that stem litter in Pinus and Quercus forests comprised 12% and 15% of the total litter, respectively [5]. Similar measurements have been determined for litter production in P. densiflora, Q. mongolica, and Abies holophylla cool temperate forests in Korea (4.33 metric tons ha−1 year−1, 3.76 metric tons ha−1 year−1, and 4.72 metric tons ha−1 year−1, respectively) [6], and stem litter has been found to comprise 16%, 12%, and 13% of the total litter, respectively [6]. Coarse woody debris (CWD) plays an important role in long-term carbon stocks in the terrestrial carbon cycle because of its slow rate of decomposition and long residence time [1,7,8,9]; it additionally provides habitat for a variety of organisms and promotes the survival and growth of seedlings [7,10,11,12]. The type, size, age, and chemical composition of wood litter are considered important and decisive factors in determining the rate of decomposition [7,13,14,15], as wood litter is generally richer in lignin and poorer in nitrogen (N) than leaf litter and is therefore expected to be more recalcitrant at decomposition and nutrient mobilization [7,9,16,17,18,19,20,21]. Therefore, total N, the C/N ratio, and the lignin/N ratio are considered good predictors of litter decomposition rates [3,22,23,24]. Plant materials with more labile compounds are susceptible to decomposition and therefore are considered higher quality, while materials with more recalcitrant compounds are considered lower quality [25,26]. Complex carbon molecules, such as lignin, reduce decomposition rates and slow degradation, while nitrogen increases quality and can lead to increased decomposition [27].
Forest nutrient cycling with respect to woody debris has been widely studied [2,21,28]. Nutrient concentrations in wood depend largely on species and site factors [29]. However, decomposition and nutrient dynamics differ greatly depending on species, initial nutrient concentrations, climate, soil type, topographic conditions, and the decomposer community [30,31,32,33,34,35]. As CWD decays, it becomes fragmented, nutrients are leached, organic matter is broken down by various decomposers, and the original material eventually becomes incorporated into the forest floor and soil [1]. Therefore, more information on woody debris decomposition is needed to better understand the basic functions of forest ecosystems [36]. In particular, few data are currently available on CWD in Korea, although some studies have been conducted on this subject [37,38,39]. However, research on decomposition has not stimulated much interest relative to that on the physiological and ecological characteristics of these systems even though there has been progress made by the Ecological Long-term Research project sponsored by the Ministry of Environment in Korea.
We hypothesized that there are different decomposition rates among tree species because each tree species has different physical properties such as density and chemical properties such as lignin, cellulose and nitrogen content in branch wood, and that the chemical composition affects the physical properties of branch wood among tree species. To test this hypothesis, in the present study, we aimed to determine the chemical components and decomposition rate of woody debris by species in a temperate forest by periodically measuring mass loss. In addition, we attempted to clarify the relationship between the decomposition rate and initial chemical composition of branch woods among tree species.
2. Materials and Methods
2.1. Study Site
We performed the decomposition experiments in a temperate deciduous forest on Mt. Baekwoon (37°21′40″ N, 127°0′14″ E; elevation 567 m above sea level) in the southern suburbs of the Seoul metropolitan area. Average annual temperature and annual precipitation from 1980 to 2010 at the Seoul meteorological station, the nearest station to the experimental site, were 12.5 °C and 1451 mm, respectively. Maximum temperature occurred in the summer (June through August) and was 22.2–25.7 °C, and precipitation was concentrated in this season with monthly precipitation in the range of 133–394 mm. During winter (December through February), average monthly temperature was −2.4–0.4 °C, and monthly precipitation was 21–25 mm. The geological matrix of the Mt. Baekwoon area consists of biotite gneiss [40].
Experimental sites were dominated by Q. mongolica and were usually mixed with P. densiflora in low densities; A. pseudosieboldianum and F. rhynchophylla dominated the subtree layer, and Rhododendron schlippenbachii, S. chinensis for. pilosa, Stephanandra incisa, and Lespedeza bicolor frequently appeared in the shrub layer. Carex lanceolata, Disporum smilacinum, and Pteridium aquilinum var. latiusculum were frequent in the herb layer. Tree coverage was 75–90%, shrub coverage 30–45%, and herb coverage 30–40%. For this study, we chose to investigate the decomposition of F. rhynchophylla, P. densiflora, Prunus sargentii, Q. mongolica, A. pseudosieboldianum and S. chinensis for. pilosa, because these species are widely distributed in deciduous temperate forests in Korea.
2.2. Experimental Design
Disk samples of branch wood were collected from each of the experimental species. For the purpose of this experiment, we considered woody branch material 30–52 mm in diameter following Harmon [1], although several previous studies have evaluated larger material. In September 2007, we obtained samples, except the S. chinensis, immediately after a summer windstorm from newly downed trees with no signs of decay [15]. Branches of S. chinensis were cut from live healthy trees in the same time as other species sampling. Five branch samples per each tree species were collected. Branch wood disk samples for each species were cut to a thickness of 1.4–1.6 cm and dried at 60 °C to a constant mass. Morphological data for the disk samples used in the experiments are shown in Table 1.
Dried woody samples were placed in 10 cm × 10 cm litterbags made of polyvinyl chloride-coated fiberglass (mesh size 2 mm) along with an aluminum identification tag. Five forest floor plots were selected randomly to place litterbags. We selected plots with similar slope (south- or southwest-facing slope) and elevation (190–210 m above sea level) to minimize the impact of other environmental factors. In each plot, 42 litterbags (7 litterbags per tree species) were placed on the forest floor surface in January 2008 for a total of 210 litterbags placed. Litterbags were retrieved seven times at approximately 4–7 month intervals during a decomposition period of 40 months (at 4, 9, 16, 21, 28, 33, and 40 months). At each sampling point, 30 litterbags were randomly collected from the five plots (6 litterbags per plot).
2.3. Mass Loss Measurements
Litterbags collected from the experimental plots were transported to the laboratory using vinyl bags. Soil or roots that penetrated into the litterbags were carefully removed, and wood samples were dried at 60 °C to a constant mass. Mass loss was calculated as the ratio of dry mass (final mass) to initial mass, and the decomposition rate constant (k) was estimated with an exponential model that characterized weight loss [41]:
where X0 was the weight (g) of litter at time 0 and Xt was the weight (g) of litter at time t (years).
k = -[ln (Xt/X0)]/t,
Litter half-life (t0.5), which is the time necessary to reach 50% mass loss, was estimated as follows [41]:
t0.5 = -ln (0.5)/(k),
2.4. Chemical Analysis
Dried wood samples were milled using a Wiley mill (Thomas Scientific, Swedesboro, NJ, USA) with a 0.1-mm mesh and were used for determining organic C, total N, lignin, and cellulose concentrations. The concentration of organic C was measured using the dichromate wet oxidation method as described by Nelson and Sommers [42]. Total N was determined by the Kjeldahl method [43] using a Tecator digestion system (FOSS, Hillerod, Denmark) and a Kjeltec 8100 distillation apparatus (FOSS, Hillerod, Denmark). Lignin and cellulose were determined using the acid detergent fiber method as described by Rowland and Roberts [44]. The concentrations of P, K, Ca, Mg, and Na were determined by the wet digestion method [45] using inductively coupled plasma optical emission spectroscopy (JY-ULTIMA-2, Horiba Jobin Yvon, Longjumeau, France). Soluble carbohydrates (SC) were determined by the anthrone method [46] after hot water extraction.
2.5. Statistical Analysis
Species differences in measured chemical, physical properties and mass loss were analyzed with one-way analyses of variance (ANOVAs) followed by the Shapiro–Wilk test for normality, Levene’s test for homogeneity and Duncan’s new multiple range test (p < 0.05). Similarly, to determine the decomposition differences between tree species, we tested using two-way ANOVAs (time × mass loss). Linear regression analyses were performed to identify relationships among initial chemical components (C, N, lignin, cellulose, C/N, and lignin/N) and mass loss of branch wood samples. All statistical analyses were conducted using SPSS 23.0 (IBM Corp., Armonk, NY, USA). Additionally, we used nonmetric multidimensional scaling (NMDS) to examine the relationships between decomposing branch wood disk samples and initial chemical concentrations for each species using the meta-multidimensional scaling and envfit functions in the Vegan library of R 2.15.3 [47].
3. Results
3.1. Chemical Composition of Branch Wood
The chemical composition of branch wood disk samples for each species is presented in Table 2. The C concentration did not significantly differ among wood species (one-way ANOVA, F = 1.144, p = 0.365), whereas total N was significantly lower in F. rhynchophylla, S. chinensis, and P. sargentii than it was in Q. mongolica, P. densiflora, and A. pseudosieboldianum (one-way ANOVA, F = 3.25, p = 0.022). Therefore, F. rhynchophylla, S. chinensis, and P. sargentii exhibited a significantly higher C/N ratio than did the other tree species (one-way ANOVA, F = 2.923, p = 0.034). Lignin concentration was highest in P. densiflora branches, while F. rhynchophylla had the lowest concentration (one-way ANOVA, F = 57, p < 0.001). Therefore, P. densiflora had the highest lignin/N ratio, and Q. mongolica, F. rhynchophylla, and A. pseudosieboldianum had the lowest values (one-way ANOVA, F = 3.94, p = 0.014). Cellulose concentration among the six tree species ranged from 48.3% in P. densiflora to 63.5% in F. rhynchophylla, and P. sargentii, Q. mongolica, S. chinensis, and A. pseudosieboldianum contained intermediate cellulose concentrations in that order (one way-ANOVA, F = 8.1, p < 0.001). The concentration of nutrient minerals in branch wood samples for each species was species specific. P, K and Na were not significantly different among tree species (one-way ANOVA, F = 1.911, 0.641 and 0.273, p = 0.130, 0.670 and 0.923, respectively); however, Ca was highest in S. chinensis and lowest in P. densiflora and F. rhynchophylla (one-way ANOVA, F = 41.98, p < 0.001), and Mg was highest in F. rhynchophylla and lowest in Q. mongolica and A. pseudosieboldianum (one-way ANOVA, F = 9.187, p < 0.001).
3.2. Mass Loss and Decomposition Rate
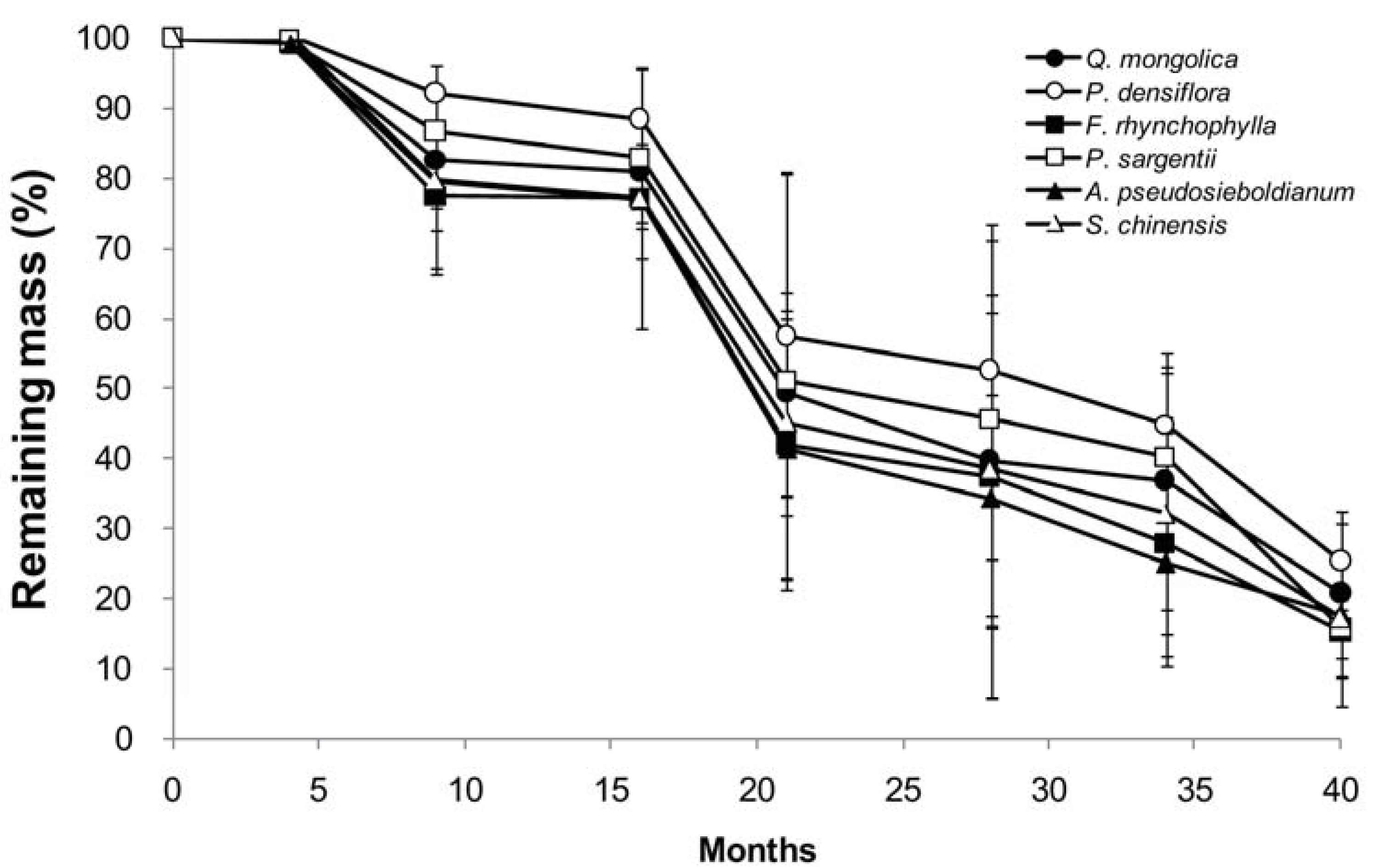
Branch wood disk mass decreased with time in a species-specific manner, and it decreased sizably during the summer seasons (4–9 months, 16–21 months and 28–34 months). Mass loss of the branch wood disk samples over 40 months is shown in Figure 1. In the early stage of decomposition (the first 9 months), branch wood mass decreased by 22.6%, 20.5%, 20.2%, 17.4%, 13.3%, and 7.8% of the initial mass for F. rhynchophylla, A. pseudosieboldianum, S. chinensis, Q. mongolica, P. sargentii, and P. densiflora, respectively (one-way ANOVA, F = 1.594, p = 0.205). After 40 months of decomposition, branch wood mass of all species decreased by more than 75% of their initial mass, with 25.4% of initial mass remaining for P. densiflora, 20.9% for Q. mongolica, 20.1% for P. sargentii, 17.7% for A pseudosieboldianum, 17.2% for S. chinensis, and 15.3% for F. rhynchophylla, respectively (one-way ANOVA, F = 1.043, p = 0.418). The difference in remaining branch wood among species was not statistically significant in overall decomposition stages (two-way ANOVA, F = 0.181, p = 1.000). Decomposition coefficients (k) of branch wood samples ranged from 0.420 in P. densiflora to 0.586 in F. rhynchophylla, and branch wood half-life ranged from 1.18 years in F. rhynchophylla to 1.65 years in P. densiflora (Table 3).
3.3. Changes in Chemical Composition during Decomposition
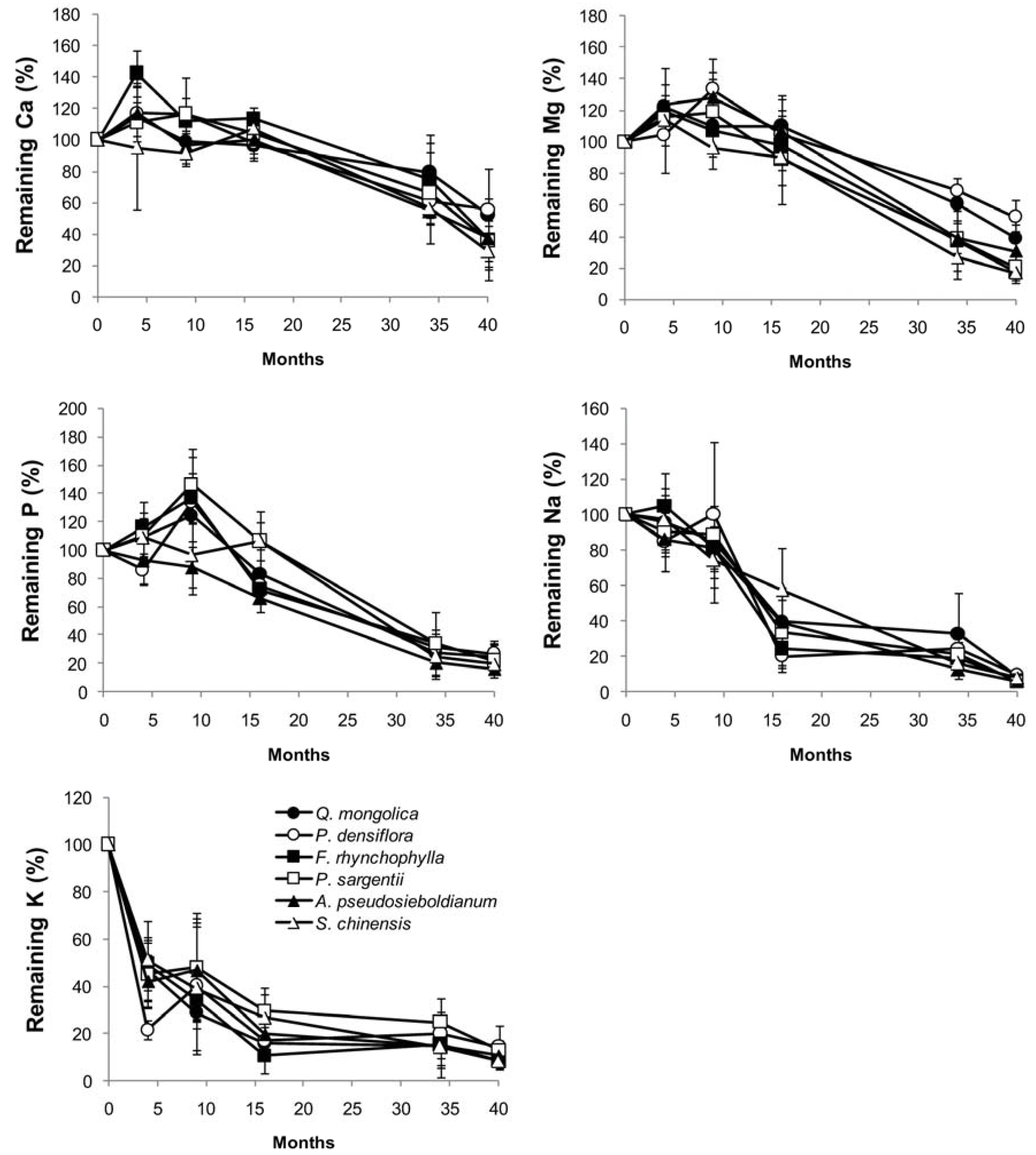
In decomposing wood samples, Ca, Mg, and P contents increased through immobilization during the early stages of decomposition in a species-specific manner (Figure 2), whereas Na and K contents exhibited a rapid decline from the beginning (Figure 2). P content tended to peak 9 months into the experiment and then decreased, whereas the P content of P. densiflora, F. rhynchophylla, and P. sargentii increased 1.3–1.4 times after 9 months. P contents in A. pseudosieboldianum and S. chinensis decreased from the beginning of decomposition. Ca and Mg contents increased in some species during the initial decay period and then gradually decreased during the rest of the experimental period. P exhibited immobilization in all species except for A. pseudosieboldianum, and Ca and Mg exhibited immobilization in all species. After 9 months, the Ca content increased 1.4-fold in F. rhynchophylla and the Mg content by 1.3-fold in P. densiflora. In the case of K and Na, nutrient contents in the branch wood samples exhibited remarkable decreases during the initial 15 months and then gradually decreased after that.
3.4. Factors Affecting Branch Wood Decomposition
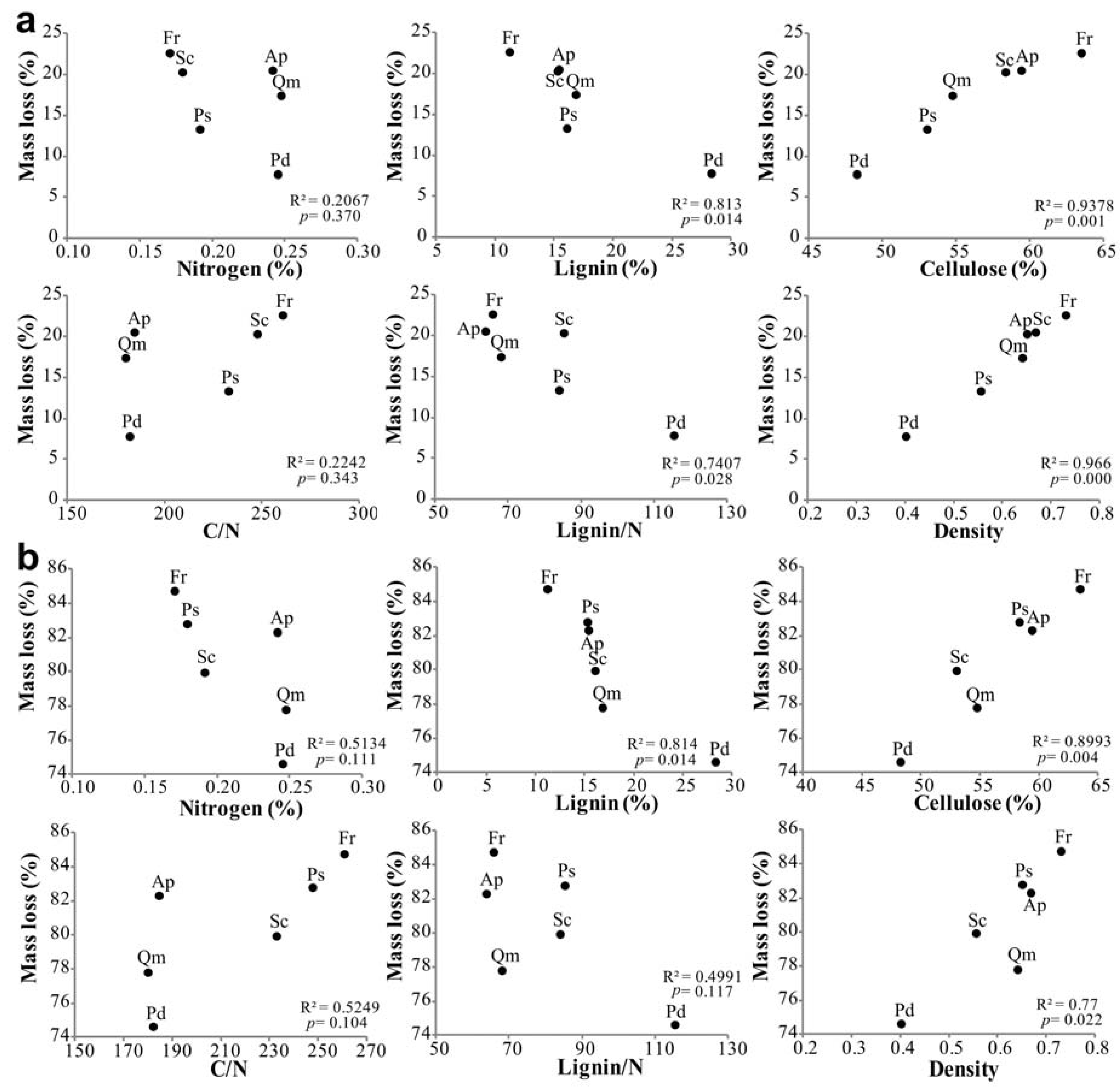
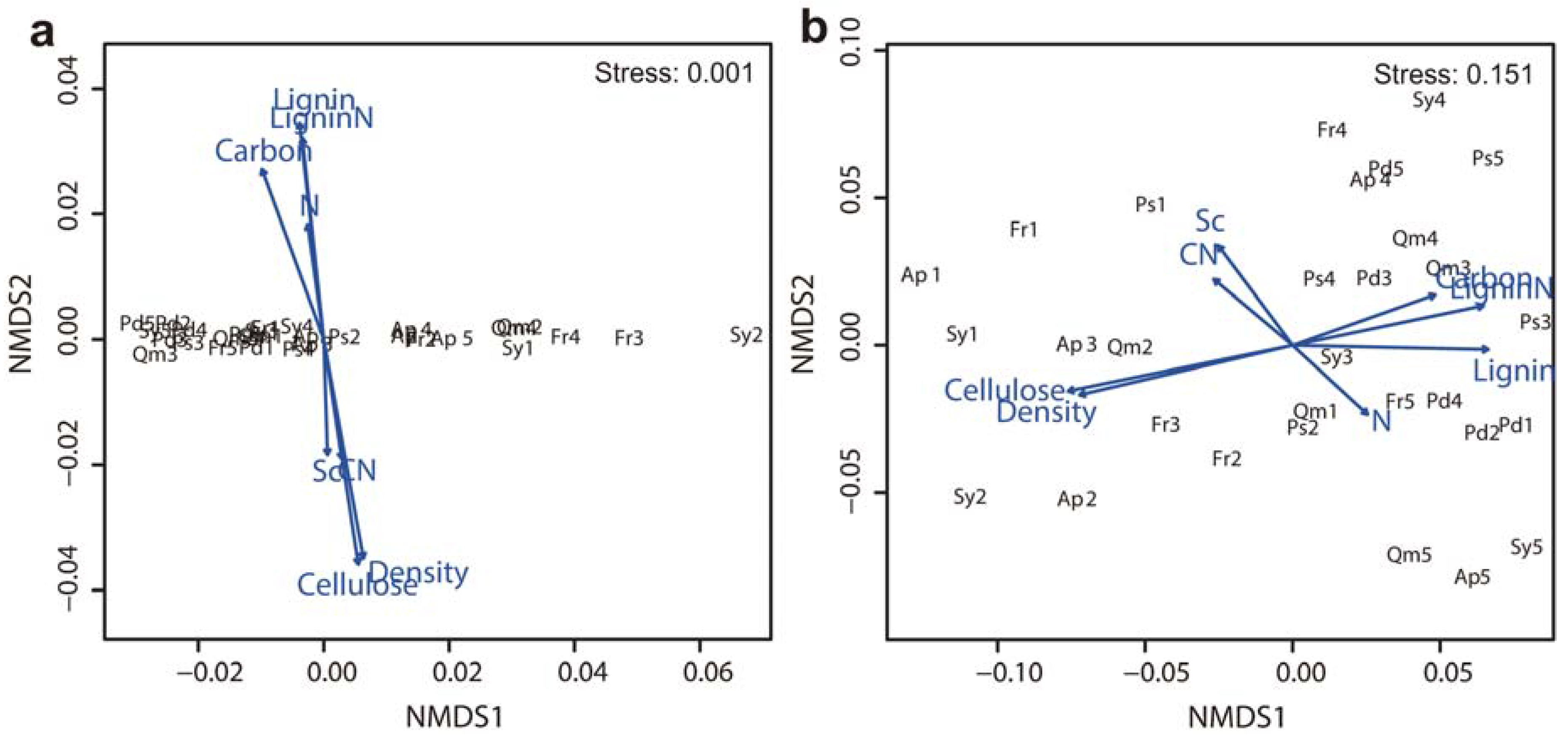
Lignin and cellulose concentrations, lignin/N ratio, and wood density of branch wood samples were the critical factors affecting branch wood decomposition among tree species. In the early decomposition stage (the first 9 months), branch wood mass loss was negatively correlated with initial lignin concentration and the lignin/N ratio. However, cellulose concentration and wood density were positively correlated with the rate of branch wood decomposition among tree species in the early decomposition stage (Figure 3a). These four parameters and index—lignin, cellulose, wood density and lignin/N—exhibited the same trend in the late decomposition stage, although the lignin/N ratio was not statistically significant (Figure 3b). However, the concentrations of nitrogen and soluble carbohydrates as well as branch wood C/N were not significantly related to the decomposition rate. Non-metric multidimensional scaling (NMDS) analysis of the relationships between branch wood decay and the chemical and physical properties of the branch wood samples produced similar results to the linear regression analyses, indicating that branch wood decomposition among tree species was significantly correlated with carbon, cellulose, and the lignin/N ratio in the early decomposition stage (Figure 4a) and that they were significantly related to lignin and cellulose concentrations in the late decomposition stage (Figure 4b).
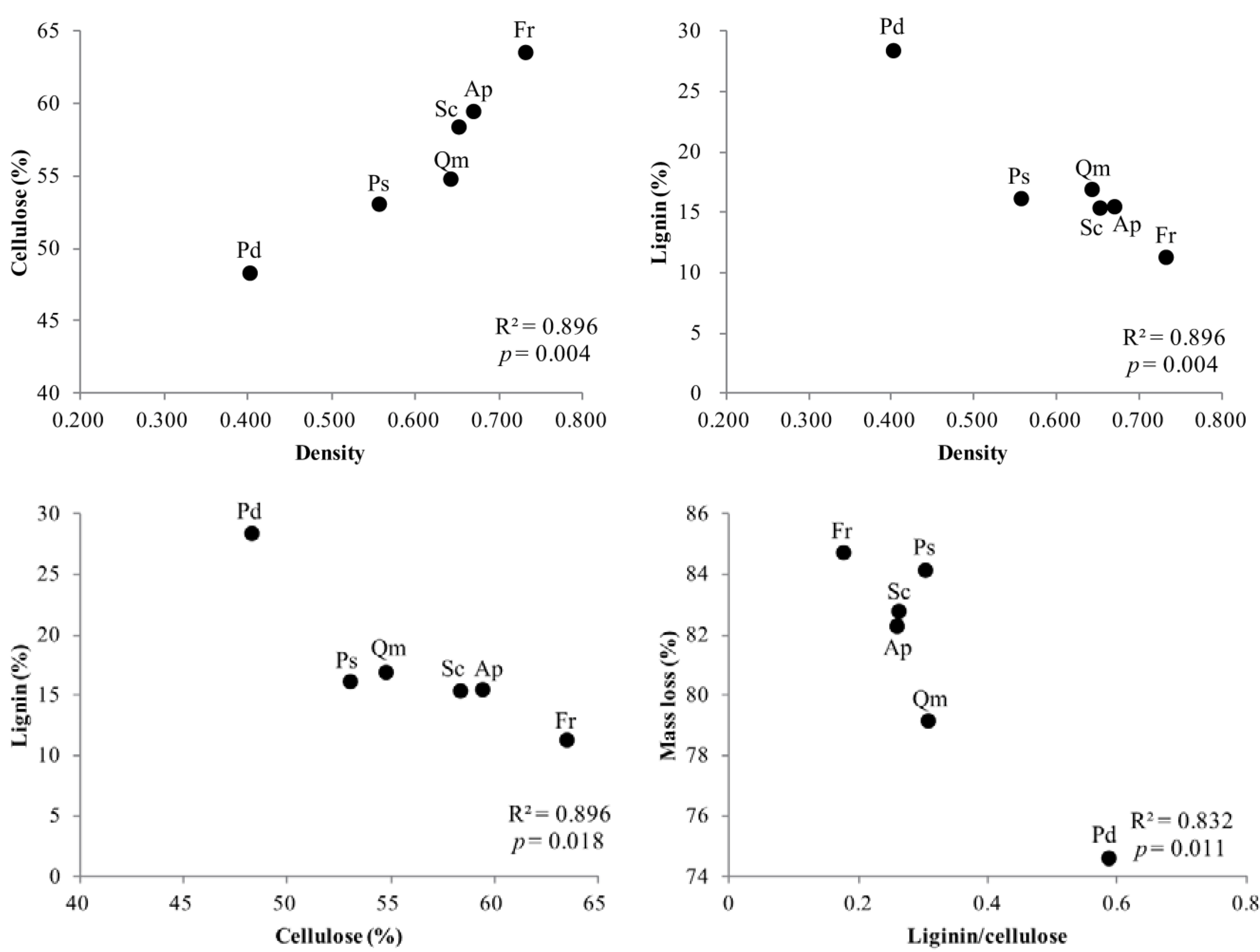
Cellulose concentration was positively correlated with wood density of branch wood among tree species. However, lignin concentration was negatively correlated with wood density as opposed to cellulose. Comparisons between cellulose and lignin concentration, and between lignin/cellulose and mass loss, were negatively correlated, respectively (Figure 5).
4. Discussion
We investigated the decomposition rates and nutrient dynamics of major tree species in a cool temperate deciduous Korean forest to compare among tree species. The six major tree species we investigated differed in decomposition rates in situ even though these rates were not statistically significantly different among species due to large variation between replicates. Ca and Mg dynamics showed similar behaviors with immobilization in the decomposing of all tree species in the early decomposition stage (Figure 2). Most Ca in plants is a component of the cell wall in the form of calcium pectate [48,49,50], which requires microbial degradation for its release. The immobilization of P, also needed for growth of microbial populations, is frequently reported during the early phases of the decomposition process [25,51,52]. The rapid K release is generally attributed to the fact that K is not bound to complex organic molecules as is the case for Ca and Mg; high solubility of K may have led to the higher rates of K losses [52]. These results of nutrient dynamics in tree species are consistent with other studies using leaf litter [50,51,52,53,54].
The differences in the decomposition rates among tree species can be attributed to differences in the initial chemical composition of the branches, since litter quality is an important factor that determines the rates of C and nutrient turnover [7,55,56]. Our results also indicated that each tree species differed in its initial chemical composition and that the decomposition rate was significantly correlated with each parameter of litter quality (Figure 3). Specifically, the concentration of lignin and the lignin/N ratio had negative effects on the decay of branch wood for each species, even more than nitrogen, which is generally considered the limiting factor in organic matter decomposition. This result was in agreement with previous studies, which reported that a relatively high lignin concentration reduces the rate of decomposition in both early- and late-stage decay [9,14,22].
In general, the mass-loss rate may be related to the concentrations of major nutrients, such as N, P, and S, which often are limiting factors for decomposition, not only within a species [57] but also among species, in the early decomposition stage [23]. Nitrogen content and the C/N ratio in litter are important factors controlling the decomposition rate [56,58] because nitrogen is an essential component of microbial functions [59], and high nitrogen content and a low C/N ratio increase the activity of microorganisms [60]. However, P. densiflora, which had relatively high N and lignin concentrations, decomposed slower than did F. rhynchophylla, which contained low N and lignin. This is because lignin more strongly affected decomposition than nitrogen did in this study (Figure 3 and Figure 4). Berg [61] indicated that free, unshielded holocellulose is decomposed in the early decomposition stage and that the late decomposition stage would thus be controlled by the lignin degradation rate, as the cellulose in the remaining parts would be shielded by lignin. This suppressing effect of lignin on litter mass-loss rates in the later decomposition stage can be described as a linear relationship [62,63].
Moreover, our results showed that branch wood that contained high cellulose content and higher density of wood, decomposed more quickly. Low-density wood is mechanically weaker [64,65,66,67,68,69,70] because a volume extension of wood requires fewer resources to construct [71], so it has been reported that wood density has a negative relationship with decomposition rate [72]. However, our results disagreed with previous studies regarding the relationship between mass loss and wood density, and indicated a positive relationship between the decomposition rate and wood density in both early and late stages of decomposition (Figure 3). Interestingly, wood density of the six tree species tested here was strongly negatively related to lignin concentration and strongly positively correlated with cellulose concentration in our experiments (Figure 5). Additionally, the cellulose concentration of branch wood was significantly negatively correlated with lignin concentration among tree species (Figure 5). As shown by these results, the cellulose content contributes to a change in the wood density, and when cellulose content of branch wood is increased, relative concentration of lignin is lower. The structure of wood can be theoretically simplified into a solid material with cellulose and air spaces (cell lumens) in its cell walls. Wood density is mainly driven by fiber in the walls and the lumen fraction [73]. Lignin is covalently linked to wall polysaccharides, especially hemicellulose and cellulose, conferring mechanical strength to the cell wall. These three major components determine the density of the cell wall and the wood tissue. Wood density is more strongly related to cellulose than to lignin (Figure 5), and species with high densities were shown to have low concentrations of lignin, which is the structural component inhibiting decomposition (Table 2, Figure 3). In our results, the lignin/cellulose ratio was negatively correlated with decomposition (Figure 5). As the relative proportion of lignin decreases due to a large amount of cellulose and to a high wood density, branches decompose faster because of the rapid degradation of cellulose that is in an unshielded state not linked to lignin [61].
We anticipated that different tree species have characteristic chemical and physical properties, and these properties affect the decomposition rate. In our study, cellulose contributed to wood density, creating a relatively lower lignin content, and the decreased lignin concentration increased the wood decomposition rate. This may be because the ratio of the recalcitrant substance was reduced, and the degradation of cellulose was largely reflected in the mass loss. This branch wood mass loss was more strongly related to initial cellulose content, lignin concentration, and the lignin/N ratio than to N or the C/N ratio in some species, as was indicated by linear regression and NMDS ordination, which is typical in the late stage of litter decomposition [23,57,73]. Therefore, we concluded that the decomposition rate was affected by the different initial chemical and physical properties of tree species, and found that difference of decomposition rate was controlled by cellulose in our study.
5. Conclusions
The present study demonstrated that in temperate deciduous forests, wood decomposition was correlated to the initial chemical composition and physical properties of the wood. Decomposition of six woody species was highly correlated with wood density and cellulose and lignin concentrations from the early stage to the late stage of decomposition. Additionally, three factors—wood density, cellulose, and lignin—had an interesting relationship. As the amount of cellulose increased, density of the wood increased, the relative concentration of lignin decreased, and, as a result, decomposition rate increased. In particular, the decrease in relative lignin concentration may be an important factor in determining the decomposition rate. Although this study focused on six species, we suggest that cellulose, lignin, and wood density are key factors in the decomposition of wood and that they are controlled by the cellulose content. Further studies are needed to evaluate the decomposition rate of more woody species and the role of cellulose in the regulation of decomposition rates.
Author Contributions
Sangsub Cha and Jae-Kuk Shim conceived and designed the experiments; Sangsub Cha, Hee-Myung Chae, and Sang-Hoon Lee performed the experiments; Sangsub Cha and Jae-Kuk Shim analyzed the data; Sangsub Cha and Jae-Kuk Shim contributed reagents/materials/analysis tools; Sangsub Cha and Jae-Kuk Shim wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.; Lattin, J.; Anderson, N.; Cline, S.; Aumen, N.G.; Sedell, J. Ecology of coarse woody debris in temperate ecosystems. In Advances in Ecological Research; MacFayden, A., Ford, E., Eds.; Academic Press: San Diego, CA, USA, 1986; Volume 15, pp. 133–302. [Google Scholar]
- Laiho, R.; Prescott, C.E. Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: A synthesis. Can. J. For. Res. 2004, 34, 763–777. [Google Scholar] [CrossRef]
- Bargali, S.S. Weight loss and n release in decomposing wood litter in a eucalypt plantation age series. Soil Biol. Biochem. 1996, 28, 699–702. [Google Scholar] [CrossRef]
- Chen, X.; Wei, X.; Scherer, R. Influence of wildfire and harvest on biomass, carbon pool, and decomposition of large woody debris in forested streams of southern interior British Columbia. For. Ecol. Manag. 2005, 208, 101–114. [Google Scholar] [CrossRef]
- Bray, J.R.; Gorham, E. Litter production in forests of the world. In Advances in Ecological Research; Cragg, J.B., Ed.; Academic Press: New York, NY, USA, 1964; Volume 2, pp. 101–157. [Google Scholar]
- Kim, T.G.; Lee, J.C.; Kim, M.J.; Joo, G.J.; Yoo, Y.H.; Min, B.M.; Lee, J.H.; Kim, E.S. The Third Stage Report (2010–2013) of KNLTER (Korea National Long Term Ecological Research); National Institute of Environmental Research: Incheon, Korea, 2013; pp. 225–236.
- Ganjegunte, G.K.; Condron, L.M.; Clinton, P.W.; Davis, M.R.; Mahieu, N. Decomposition and nutrient release from radiata pine (Pinus radiata) coarse woody debris. For. Ecol. Manag. 2004, 187, 197–211. [Google Scholar] [CrossRef]
- Zhou, L.; Dai, L.-m.; Gu, H.-y.; Zhong, L. Review on the decomposition and influence factors of coarse woody debris in forest ecosystem. J. For. Res. 2007, 18, 48–54. [Google Scholar] [CrossRef]
- Garrett, L.; Oliver, G.; Pearce, S.; Davis, M. Decomposition of Pinus radiata coarse woody debris in New Zealand. For. Ecol. Manag. 2008, 255, 3839–3845. [Google Scholar] [CrossRef]
- Abbott, D.; Crossley, D. Woody litter decomposition following clear-cutting. Ecology 1982, 63, 35–42. [Google Scholar] [CrossRef]
- Freedman, B.; Zelazny, V.; Beaudette, D.; Fleming, T.; Johnson, G.; Flemming, S.; Gerrow, J.; Forbes, G.; Woodley, S. Biodiversity implications of changes in the quantity of dead organic matter in managed forests. Environ. Rev. 1996, 4, 238–265. [Google Scholar] [CrossRef]
- Bunnell, F.L.; Houde, I. Down wood and biodiversity—Implications to forest practices. Environ. Rev. 2010, 18, 397–421. [Google Scholar] [CrossRef]
- Gallardo, A.; Merino, J. Leaf decomposition in two Mediterranean ecosystems of southwest Spain: Influence of substrate quality. Ecology 1993, 74, 152–161. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Plant Litter. Decomposition Humus Formation. Carbon Sequestration; Springer: Berlin, Germany, 2008. [Google Scholar]
- Chae, H.M.; Cha, S.; Lee, S.H.; Choi, M.J.; Shim, J.K. Age-related decomposition of Quercus mongolica. Plant Ecol. 2016, 217, 945–957. [Google Scholar] [CrossRef]
- Berg, B.; Staaf, H. Decomposition rate and chemical changes of scots pine needle litter. II. Influence of chemical composition. Ecol. Bull. 1980, 373–390. [Google Scholar]
- Upadhyay, V.; Singh, J. Decomposition of woody branch litter on an altitudinal transect in the Himalaya. Plant Ecol. 1986, 64, 49–53. [Google Scholar] [CrossRef]
- Fioretto, A.; Di Nardo, C.; Papa, S.; Fuggi, A. Lignin and cellulose degradation and nitrogen dynamics during decomposition of three leaf litter species in a Mediterranean ecosystem. Soil Biol. Biochem. 2005, 37, 1083–1091. [Google Scholar] [CrossRef]
- Alban, D.H.; Pastor, J. Decomposition of aspen, spruce, and pine boles on two sites in Minnesota. Can. J. For. Res. 1993, 23, 1744–1749. [Google Scholar] [CrossRef]
- Herrmann, S.; Prescott, C.E. Mass loss and nutrient dynamics of coarse woody debris in three rocky mountain coniferous forests: 21 year results. Can. J. For. Res. 2008, 38, 125–132. [Google Scholar] [CrossRef]
- Saunders, M.R.; Fraver, S.; Wagner, R.G. Nutrient concentration of down woody debris in mixedwood forests in central Maine, USA. Silva Fennica 2011, 45, 197–210. [Google Scholar] [CrossRef]
- Melillo, J.M.; Aber, J.D.; Muratore, J.F. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 1982, 63, 621–626. [Google Scholar] [CrossRef]
- Berg, B.; Ekbohm, G. Litter mass-loss rates and decomposition patterns in some needle and leaf litter types. Long-term decomposition in a scots pine forest. VII. Can. J. Bot. 1991, 69, 1449–1456. [Google Scholar] [CrossRef]
- Cha, S.; Chae, H.M.; Lee, S.H.; Shim, J.K. Effect of elevated atmospheric CO2 concentration on growth and leaf litter decomposition of Quercus acutissima and Fraxinus rhynchphylla. PLoS ONE 2017, 12, e0171197. [Google Scholar] [CrossRef] [PubMed]
- Aber, J.; Melillo, J.M. Terrestrial Ecosystems, 2nd ed.; Brooks/Cole Publishing: Pacific Grove, CA, USA, 2001. [Google Scholar]
- Chapin, F., III; Matson, P.; Mooney, H. Principles of Terrestrial Ecosystem Ecology; Springer: New York, NY, USA, 2002. [Google Scholar]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; Univ of California Press: Berkeley, CA, USA, 1979; Volume 5. [Google Scholar]
- Pastor, J.; Bockheim, J. Distribution and cycling of nutrients in an aspen-mixed-hardwood-spodosol ecosystem in northern Wisconsin. Ecology 1984, 65, 339–353. [Google Scholar] [CrossRef]
- Pallardy, S.G. Physiology of Woody Plants; Academic Press: San Diego, CA, USA, 2010. [Google Scholar]
- Maser, C.; Trappe, J.M. The Seen and Unseen World of the Fallen Tree; USDA Forest Service, Pacific Northwest Forest Experiment Station: Portland, OR, USA, 1984. [Google Scholar]
- Brais, S.; Paré, D.; Lierman, C. Tree bole mineralization rates of four species of the Canadian eastern boreal forest: Implications for nutrient dynamics following stand-replacing disturbances. Can. J. For. Res. 2006, 36, 2331–2340. [Google Scholar] [CrossRef]
- Kuehne, C.; Donath, C.; Müller-Using, S.; Bartsch, N. Nutrient fluxes via leaching from coarse woody debris in a Fagus sylvatica forest in the Solling mountains, Germany. Can. J. For. Res. 2008, 38, 2405–2413. [Google Scholar] [CrossRef]
- Herrmann, S.; Bauhus, J. Effects of moisture, temperature and decomposition stage on respirational carbon loss from coarse woody debris (CWD) of important European tree species. Scand. J. For. Res. 2013, 28, 346–357. [Google Scholar] [CrossRef]
- Gonzalez-Polo, M.; Fernández-Souto, A.; Austin, A.T. Coarse woody debris stimulates soil enzymatic activity and litter decomposition in an old-growth temperate forest of Patagonia, Argentina. Ecosystems 2013, 16, 1025–1038. [Google Scholar] [CrossRef]
- Lombardi, F.; Cherubini, P.; Tognetti, R.; Cocozza, C.; Lasserre, B.; Marchetti, M. Investigating biochemical processes to assess deadwood decay of beech and silver fir in Mediterranean mountain forests. Ann. For. Sci. 2013, 70, 101–111. [Google Scholar] [CrossRef]
- Tuomi, M.; Laiho, R.; Repo, A.; Liski, J. Wood decomposition model for boreal forests. Ecol. Model. 2011, 222, 709–718. [Google Scholar] [CrossRef]
- Jang, D.; Youn, Y. A basic survey about dead tree of old Korean fir stands in Mt. Sorak. Kor. J. Environ. Biol. 2003, 21, 251–256. [Google Scholar]
- Kim, R.H.; Son, Y.; Lim, J.H.; Lee, I.K.; Seo, K.W.; Koo, J.W.; Noh, N.J.; Ryu, S.-R.; Hong, S.K.; Ihm, B.S. Coarse woody debris mass and nutrients in forest ecosystems of Korea. Ecol. Res. 2006, 21, 819–827. [Google Scholar] [CrossRef]
- Noh, N.J.; Son, Y.; Lee, S.K.; Seo, K.W.; Heo, S.J.; Yi, M.J.; Park, P.S.; Kim, R.H.; Son, Y.M.; Lee, K.H. Carbon and nitrogen storage in an age-sequence of Pinus densiflora stands in Korea. Sci. China Life Sci. 2010, 53, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Korea Institute of Geoscience and Mineral Resources. Available online: http://mgeo.kigam.re.kr/map/geology.jsp (accessed on 4 April 2017).
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis Part 3—Chemical Methods; Bartels, J.M., Ed.; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Bremner, J. Nitrogen-total. In Methods of Soil Analysis Part 3—Chemical Methods; Bartels, J.M., Ed.; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Rowland, A.; Roberts, J. Lignin and cellulose fractionation in decomposition studies using acid-detergent fibre methods. Commun. Soil Sci. Plant Anal. 1994, 25, 269–277. [Google Scholar] [CrossRef]
- Helrick, K. Official Methods of Analysis; Association of Official Analytical Chemists, Inc.: Arlington, VA, USA, 1990. [Google Scholar]
- Allen, S.E.; Grimshaw, H.M.; Parkinson, J.A.; Quarmby, C. Chemical Analysis of Ecological Materials; Blackwell Scientific Publications: Oxford, UK, 1974. [Google Scholar]
- R Development Core Team. Available online: http://cran.r-project.org (accessed on 7 February 2017).
- Salisbury, F.B.; Ross, C.W. Plant Physiology, 3rd ed.; Wadsworth Publishing Co.: Belmont, CA, USA, 1985; p. 540. [Google Scholar]
- Palviainen, M.; Finér, L.; Kurka, A.M.; Mannerkoski, H.; Piirainen, S.; Starr, M. Release of potassium, calcium, iron and aluminium from Norway spruce, Scots pine and silver birch logging residues. Plant Soil 2004, 259, 123–136. [Google Scholar] [CrossRef]
- Bonanomi, G.; Incerti, G.; Antignani, V.; Capodilupo, M.; Mazzoleni, S. Decomposition and nutrient dynamics in mixed litter of Mediterranean species. Plant Soil 2010, 331, 481–496. [Google Scholar] [CrossRef]
- Ventura, M; Scandellari, F.; Bonora, E.; Tagliavini, M. Nutrient release during decomposition of leaf litter in a peach (Prunus persica L.) orchard. Nutr. Cycl. Agroecosyst. 2009, 87, 115–125. [Google Scholar]
- Brunetto, G; Ventura, M.; Scandellari, F.; Ceretta, C.A.; Kaminski, J.; Melo, G.W.B.; Taglavini, M. Nutrients release during the decomposition of mowed perennial ryegrass and white clover and its contribution to nitrogen nutrition of grapevine. Nutr. Cycl. Agroecosyst. 2011, 90, 299–308. [Google Scholar]
- Tagliavini, M.; Tonon, G.; Scandellari, F.; Quiñones, A.; Palmieri, S.; Menarbin, G.; Gioacchini, P.; Masia, A. Nutrient recycling during the decomposition of apple leaves (Malus domestica) and mowed grasses in an orchard. Agric. Ecosyst. Environ. 2007, 118, 191–200. [Google Scholar] [CrossRef]
- Rodríguez-Lizana, A.; Carbonell, R.; González, P.; Ordóñez, R. N, P and K released by the field decomposition of residues of a pea-wheat-sunflower rotation. Nutr. Cycl. Agroecosyst. 2010, 87, 199–208. [Google Scholar] [CrossRef]
- Swift, M.; Healey, I.; Hibberd, J.; Sykes, J.; Bampoe, V.; Nesbitt, M. The decomposition of branch-wood in the canopy and floor of a mixed deciduous woodland. Oecologia 1976, 26, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Heal, O.; Anderson, J.; Swift, M. Plant litter quality and decomposition: An historical overview. In Driven by Nature: Plant Litter Quality and Decomposition; Cadisch, G., Ed.; CAB International: Walling Ford, UK, 1997. [Google Scholar]
- Berg, B.; Laskowski, R. Decomposers: Soil microorganisms and animals. In Advances in Ecological Research; Berg, B., Laskowski, R., Eds.; Academic Press: San Diego, CA, USA, 2005; Volume 38, pp. 73–100. [Google Scholar]
- Singh, J.; Gupta, S. Plant decomposition and soil respiration in terrestrial ecosystems. Bot. Rev. 1977, 43, 449–528. [Google Scholar] [CrossRef]
- Zhang, Q.; Zak, J.C. Effects of gap size on litter decomposition and microbial activity in a subtropical forest. Ecology 1995, 76, 2196–2204. [Google Scholar] [CrossRef]
- Berg, B. Litter decomposition and organic matter turnover in northern forest soils. For. Ecol. Manag. 2000, 133, 13–22. [Google Scholar] [CrossRef]
- Fogel, R.; Cromack, K., Jr. Effect of habitat and substrate quality on douglas fir litter decomposition in western Oregon. Can. J. Bot. 1977, 55, 1632–1640. [Google Scholar] [CrossRef]
- Berg, B.; Lundmark, J.E. Decomposition of needle litter in Pinus contorta and Pinus sylvestris monocultures—A comparison. Scand. J. For. Res. 1987, 2, 3–12. [Google Scholar] [CrossRef]
- Niklas, K.J. Mechanical properties of black locust (Robinia pseudoacacia) wood: Correlations among elastic and rupture moduli, proportional limit, and tissue density and specific gravity. Ann. Bot. 1997, 79, 479–485. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S. Functional and ecological xylem anatomy. Perspect. Plant Ecol. Evol. Syst. 2001, 4, 97–115. [Google Scholar] [CrossRef]
- Jacobsen, A.L.; Ewers, F.W.; Pratt, R.B.; Paddock, W.A.; Davis, S.D. Do xylem fibers affect vessel cavitation resistance? Plant Physiol. 2005, 139, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Van Gelder, H.; Poorter, L.; Sterck, F. Wood mechanics, allometry, and life-history variation in a tropical rain forest tree community. New Phytol. 2006, 171, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, A.L.; Agenbag, L.; Esler, K.J.; Pratt, R.B.; Ewers, F.W.; Davis, S.D. Xylem density, biomechanics and anatomical traits correlate with water stress in 17 evergreen shrub species of the Mediterranean-type climate region of south Africa. J. Ecol. 2007, 95, 171–183. [Google Scholar] [CrossRef]
- Osunkoya, O.O.; Sheng, T.K.; Mahmud, N.A.; Damit, N. Variation in wood density, wood water content, stem growth and mortality among twenty-seven tree species in a tropical rainforest on Borneo island. Austral Ecol. 2007, 32, 191–201. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Poorter, L. The relationships of wood-, gas-and water fractions of tree stems to performance and life history variation in tropical trees. Ann. Bot. 2008, 102, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Chambers, J.Q.; Higuchi, N; Schimel, J.P.; Ferreira, L.V.; Melack, J.M. Decomposition and carbon cycling of dead trees in tropical forests of the central Amazon. Oecologia 2000, 122, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Ziemińska, K.; Butler, D.W.; Gleason, S.M.; Wright, I.J.; Westoby, M. Fibre wall and lumen fractions drive wood density variation across 24 Australian angiosperms. AoB Plants 2013, 5, plt046. [Google Scholar] [CrossRef]
- Berg, B.; Matzner, E. The effect of n deposition on the mineralization of c from plant litter and humus. Environ. Rev. 1997, 5, 1–25. [Google Scholar] [CrossRef]
Figure 1.
Change in branch wood disk mass of six tree species during a decomposition period of 40 months. Bars indicate SD (n = 5).
Figure 1.
Change in branch wood disk mass of six tree species during a decomposition period of 40 months. Bars indicate SD (n = 5).
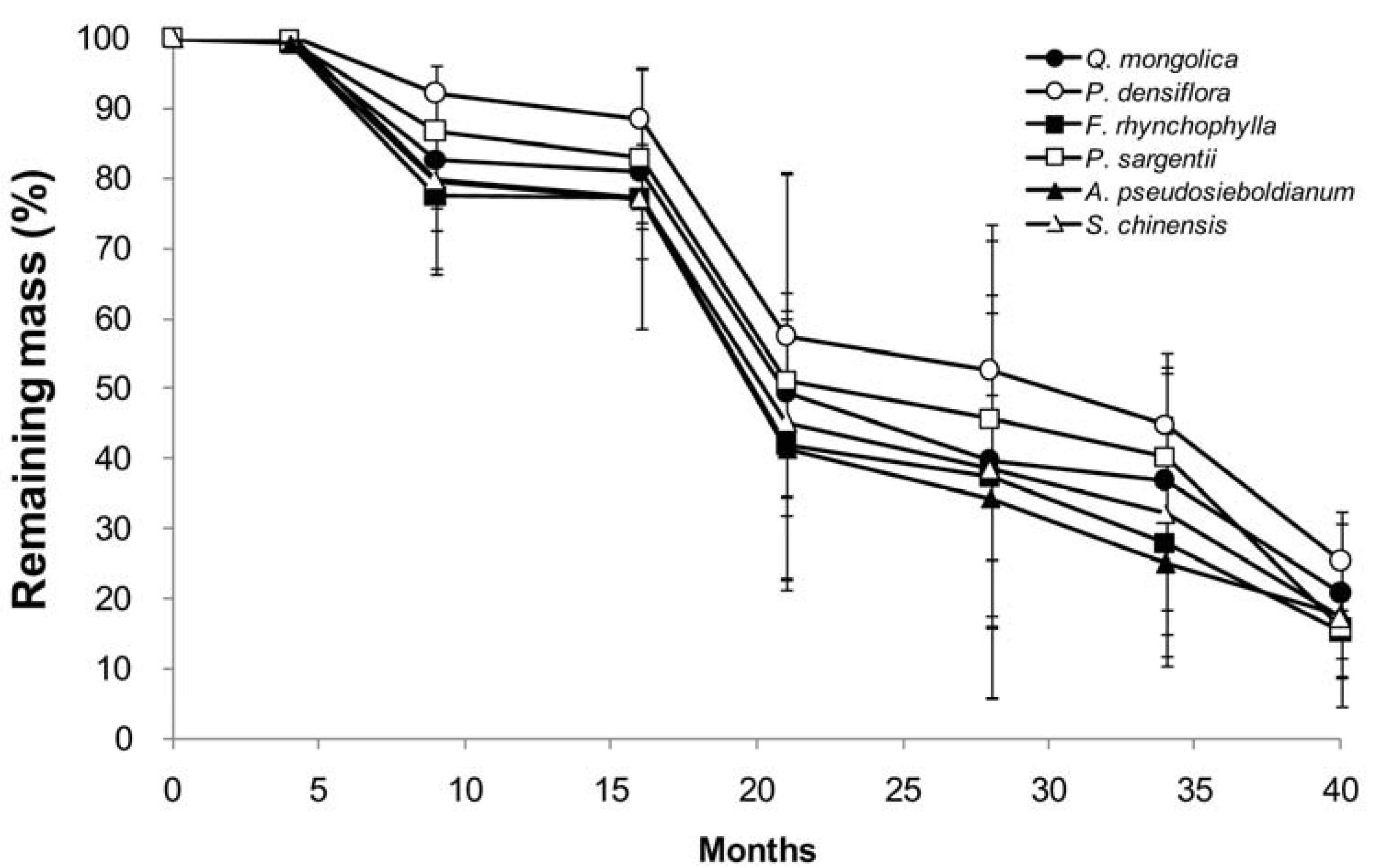
Figure 2.
Change in calcium, phosphorus, potassium, magnesium, and sodium content in branch wood disk samples of six tree species during a decomposition period of 40 months (n = 5).
Figure 2.
Change in calcium, phosphorus, potassium, magnesium, and sodium content in branch wood disk samples of six tree species during a decomposition period of 40 months (n = 5).
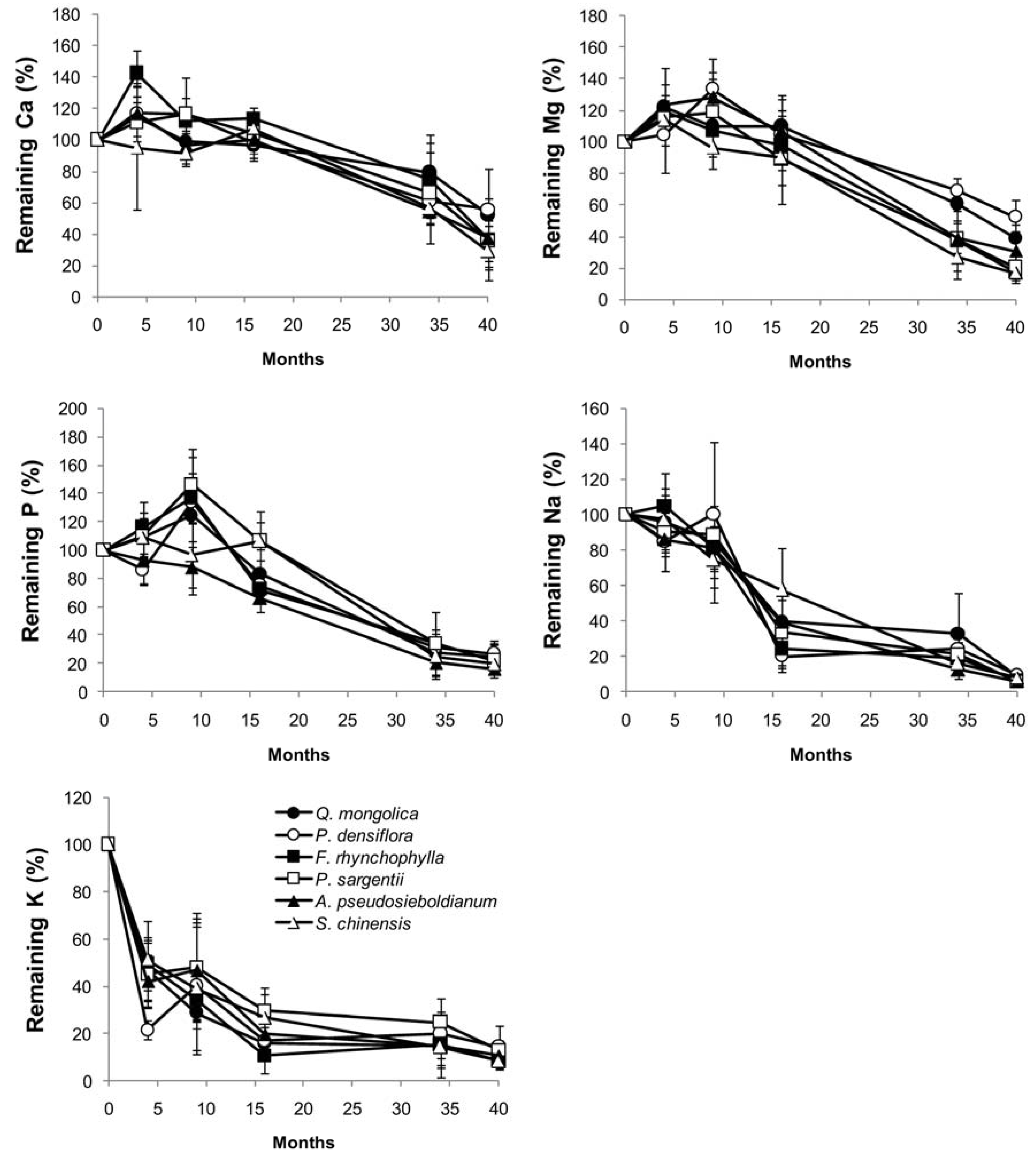
Figure 3.
Linear regression of branch wood mass loss on initial chemical and physical factors for six tree species (a) during the early stage of decomposition (from the beginning of decomposition to 9 months) and (b) during the entire decomposition period of 40 months (n = 5). Ap (A. pseudosieboldianum), Fr (F. rhynchophylla), Pd (P. densiflora), Ps (P. sargentii), Qm (Q. mongolica) and Sc (S. chinensis).
Figure 3.
Linear regression of branch wood mass loss on initial chemical and physical factors for six tree species (a) during the early stage of decomposition (from the beginning of decomposition to 9 months) and (b) during the entire decomposition period of 40 months (n = 5). Ap (A. pseudosieboldianum), Fr (F. rhynchophylla), Pd (P. densiflora), Ps (P. sargentii), Qm (Q. mongolica) and Sc (S. chinensis).

Figure 4.
NMDS ordination biplot based on the mass loss of branch wood for six tree species (a) during the early stage of decomposition (from the beginning of decomposition to 9 months) and (b) during the entire decomposition period of 40 months (n = 5). Angles and lengths of radiating arrows indicate the direction and strength, respectively, of the relationships between litter quality parameters. See abbreviation in Figure 3.
Figure 4.
NMDS ordination biplot based on the mass loss of branch wood for six tree species (a) during the early stage of decomposition (from the beginning of decomposition to 9 months) and (b) during the entire decomposition period of 40 months (n = 5). Angles and lengths of radiating arrows indicate the direction and strength, respectively, of the relationships between litter quality parameters. See abbreviation in Figure 3.
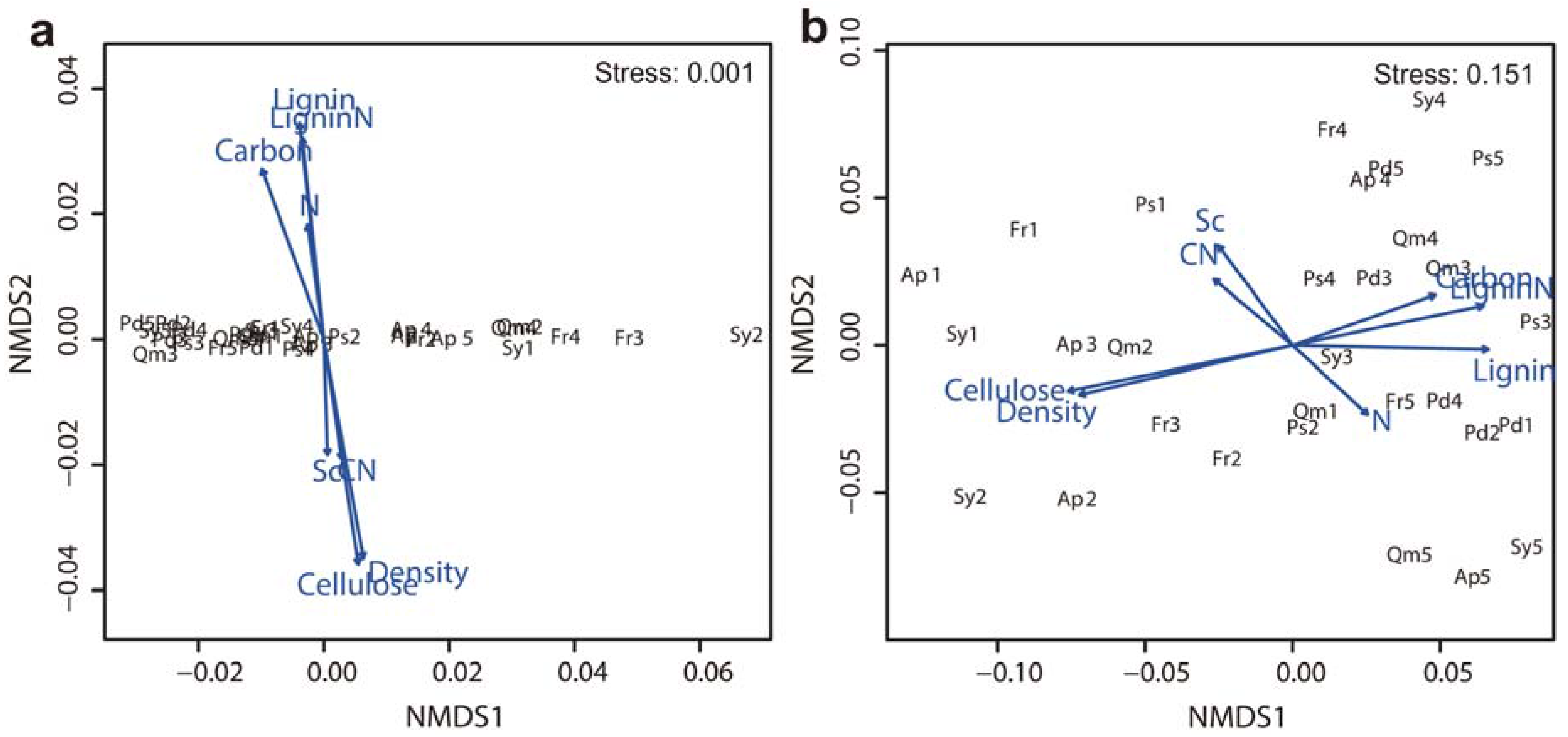
Figure 5.
Linear regression between density, cellulose, lignin concentration, and the lignin/cellulose ratio of branch wood disk samples for a decomposition period of 40 months (n = 5). See abbreviation in Figure 3.
Figure 5.
Linear regression between density, cellulose, lignin concentration, and the lignin/cellulose ratio of branch wood disk samples for a decomposition period of 40 months (n = 5). See abbreviation in Figure 3.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Morphological properties of branch wood disk samples for six tree species. Values are means (n = 5) ± SD.
Table 1.
Morphological properties of branch wood disk samples for six tree species. Values are means (n = 5) ± SD.
| Species | Diameter (mm) | Thickness (mm) | Density (g/cm3) |
|---|---|---|---|
| Quercus mongolica | 51.97 ± 2.57 | 15.74 ± 2.77 | 0.642 ± 0.015 |
| Pinus densiflora | 36.73 ± 5.10 | 14.92 ± 1.22 | 0.403 ± 0.038 |
| Fraxinus rhynchophylla | 29.93 ± 2.72 | 15.57 ± 1.27 | 0.731 ± 0.026 |
| Prunus sargentii | 46.92 ± 3.21 | 14.53 ± 1.24 | 0.557 ± 0.023 |
| Acer pseudosieboldianum | 33.31 ± 2.19 | 14.73 ± 1.44 | 0.669 ± 0.020 |
| Symplocos chinensis | 30.15 ± 2.54 | 16.06 ± 1.83 | 0.652 ± 0.028 |
Table 2.
Morphological properties of branch wood disk samples for six tree species. Values are means (n = 5) ± SD.
Table 2.
Morphological properties of branch wood disk samples for six tree species. Values are means (n = 5) ± SD.
| Q. mongolica | P. densiflora | F. rhynchophylla | P. sargentii | A. pseudosieboldianum | S. chinensis | |
|---|---|---|---|---|---|---|
| Carbon (%) | 44.6 ± 0.09 a 1 | 44.7 ± 0.16 a | 44.6 ± 0.10 a | 44.7 ± 0.04 a | 44.7 ± 0.22 a | 44.5 ± 0.23 a |
| Nitrogen (%) | 0.25 ± 0.03 a | 0.25 ± 0.07 a | 0.17 ± 0.02 b | 0.19 ± 0.03 ab | 0.24 ± 0.05 a | 0.18 ± 0.05 b |
| Lignin (%) | 16.2 ± 1.75 b | 28.4 ± 1.46 a | 10.6 ± 1.85 c | 16.1 ± 1.89 b | 15.5 ± 0.58 b | 16.3 ± 1.88 b |
| Cellulose (%) | 54.8 ± 6.34 bc | 48.3 ± 1.85 d | 63.5 ± 3.18 a | 53.0 ± 4.58 cd | 59.4 ± 2.43 ab | 58.4 ± 3.40 abc |
| SC 2 (%) | 1.54 ± 0.69 a | 1.42 ± 0.87 a | 2.22 ± 0.71 a | 2.58 ± 1.10 a | 2.08 ± 0.65 a | 1.76 ± 0.56 a |
| C/N | 180.1 ± 24.6 c | 182.3 ± 65.6 c | 261.0 ± 29.1 a | 233.1 ± 33.4 ab | 184.7 ± 33.1 bc | 247.9 ± 74.9 ab |
| Lignin/N | 65.6 ± 9.3 b | 115.6 ± 46.7 a | 61.8 ± 9.7 b | 84.1 ± 15.9 b | 63.9 ± 14.2 b | 90.6 ± 26.2 b |
| Ca (μg /g) | 3722 ± 235 b | 1913 ± 257 e | 2678 ± 442 d | 3098 ± 604 cd | 3485 ± 298 bc | 5371 ± 448 a |
| P (μg /g) | 193 ± 46 b | 280 ± 138 ab | 212 ± 56 ab | 201 ± 46 b | 312 ± 45 a | 236 ± 83 ab |
| K (μg /g) | 815± 246 a | 865 ± 776 a | 921 ± 361 a | 562 ± 72 a | 680 ± 58 a | 821 ± 134 a |
| Mg (μg /g) | 219 ± 21 cd | 281 ± 68 cd | 443 ± 105 a | 301 ± 54 bc | 190 ± 27 d | 374 ± 99 ab |
| Na (μg /g) | 461 ± 126 a | 495 ± 145 a | 452 ± 57 a | 501 ± 98 a | 513 ± 64 a | 506 ± 118 a |
1 The same lowercase letter in the same horizontal row denotes non-significant differences at p < 0.05 among the groups. Different lowercase letters in the same row indicate significant differences at p < 0.05 (Duncan’s new multiple range test); 2 SC: Soluble carbohydrate.
Table 3.
Morphological properties of branch wood disk samples for six tree species. Values are means (n = 5) ± SD.
Table 3.
Morphological properties of branch wood disk samples for six tree species. Values are means (n = 5) ± SD.
| Q. mongolica | P. densiflora | F. rhynchophylla | P. sargentii | A. pseudosieboldianum | S. chinensis | |
|---|---|---|---|---|---|---|
| k | 0.478 ± 0.077 | 0.420 ± 0.079 | 0.586 ± 0.138 | 0.560 ± 0.078 | 0.575 ± 0.193 | 0.561 ± 0.166 |
| Half-life (year) | 1.479 ± 0.214 | 1.705 ± 0.355 | 1.233 ± 0.289 | 1.257 ± 0.179 | 1.384 ± 0.675 | 1.321 ± 0.365 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cha, S.; Chae, H.-M.; Lee, S.-H.; Shim, J.-K. Branch Wood Decomposition of Tree Species in a Deciduous Temperate Forest in Korea. Forests 2017, 8, 176. https://doi.org/10.3390/f8050176
AMA Style
Cha S, Chae H-M, Lee S-H, Shim J-K. Branch Wood Decomposition of Tree Species in a Deciduous Temperate Forest in Korea. Forests. 2017; 8(5):176. https://doi.org/10.3390/f8050176
Chicago/Turabian StyleCha, Sangsub, Hee-Myung Chae, Sang-Hoon Lee, and Jae-Kuk Shim. 2017. "Branch Wood Decomposition of Tree Species in a Deciduous Temperate Forest in Korea" Forests 8, no. 5: 176. https://doi.org/10.3390/f8050176
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.