Mapping Tree Density in Forests of the Southwestern USA Using Landsat 8 Data


Abstract
:1. Introduction
2. Materials and Methods
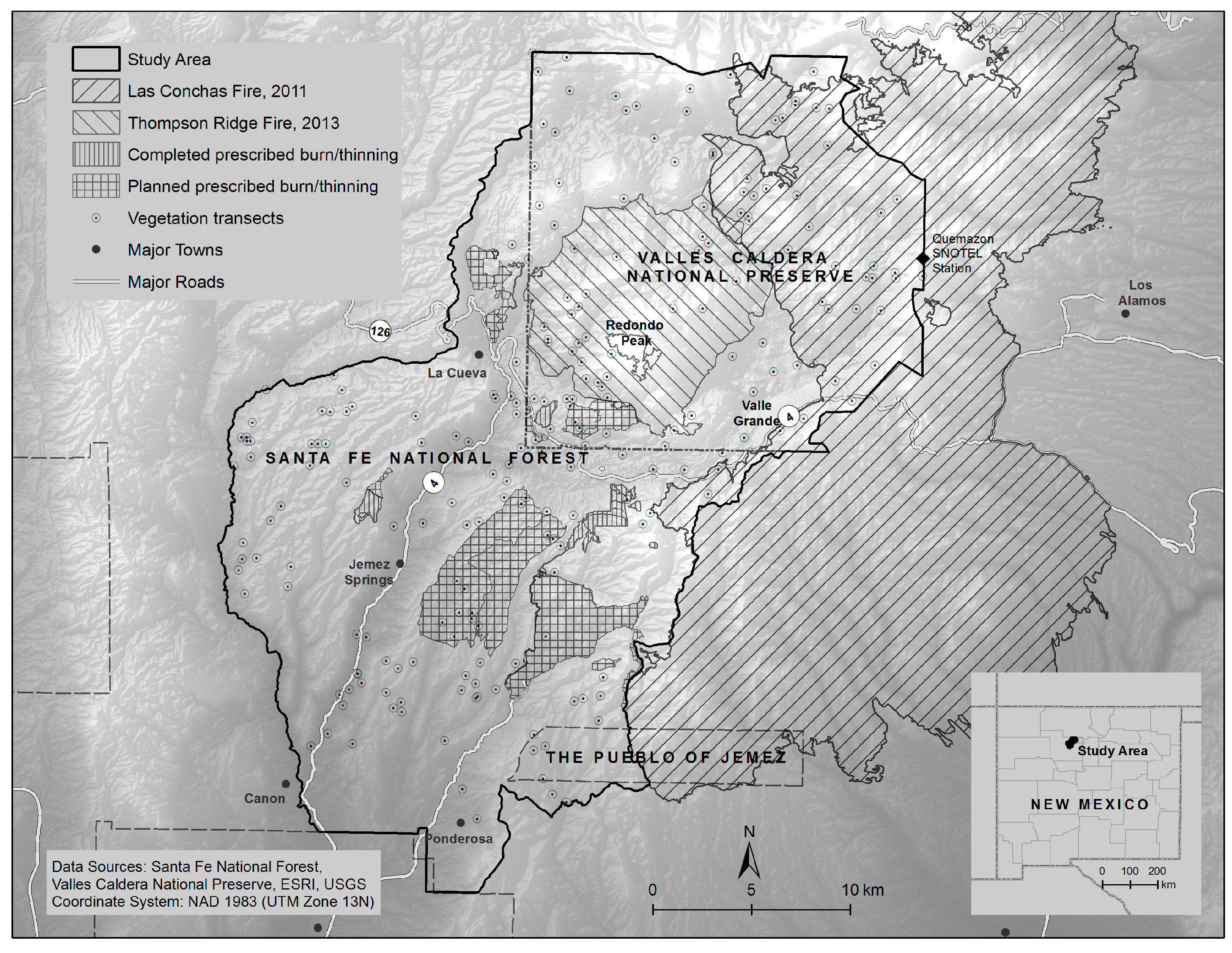
2.1. Study Area
2.2. Field Data
2.3. Remote Sensing Data Acquisition and Processing
2.4. Data Analysis
3. Results
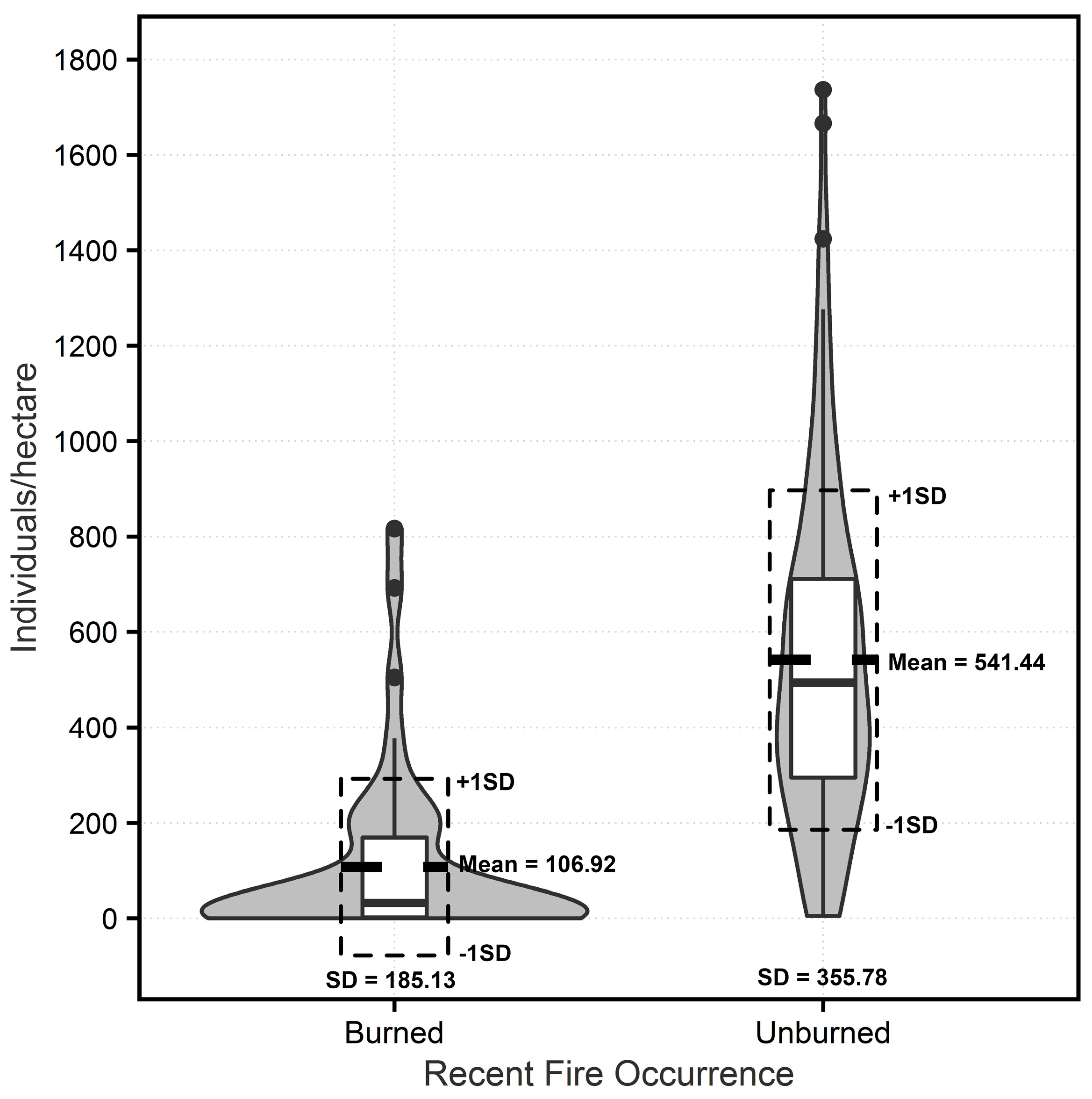
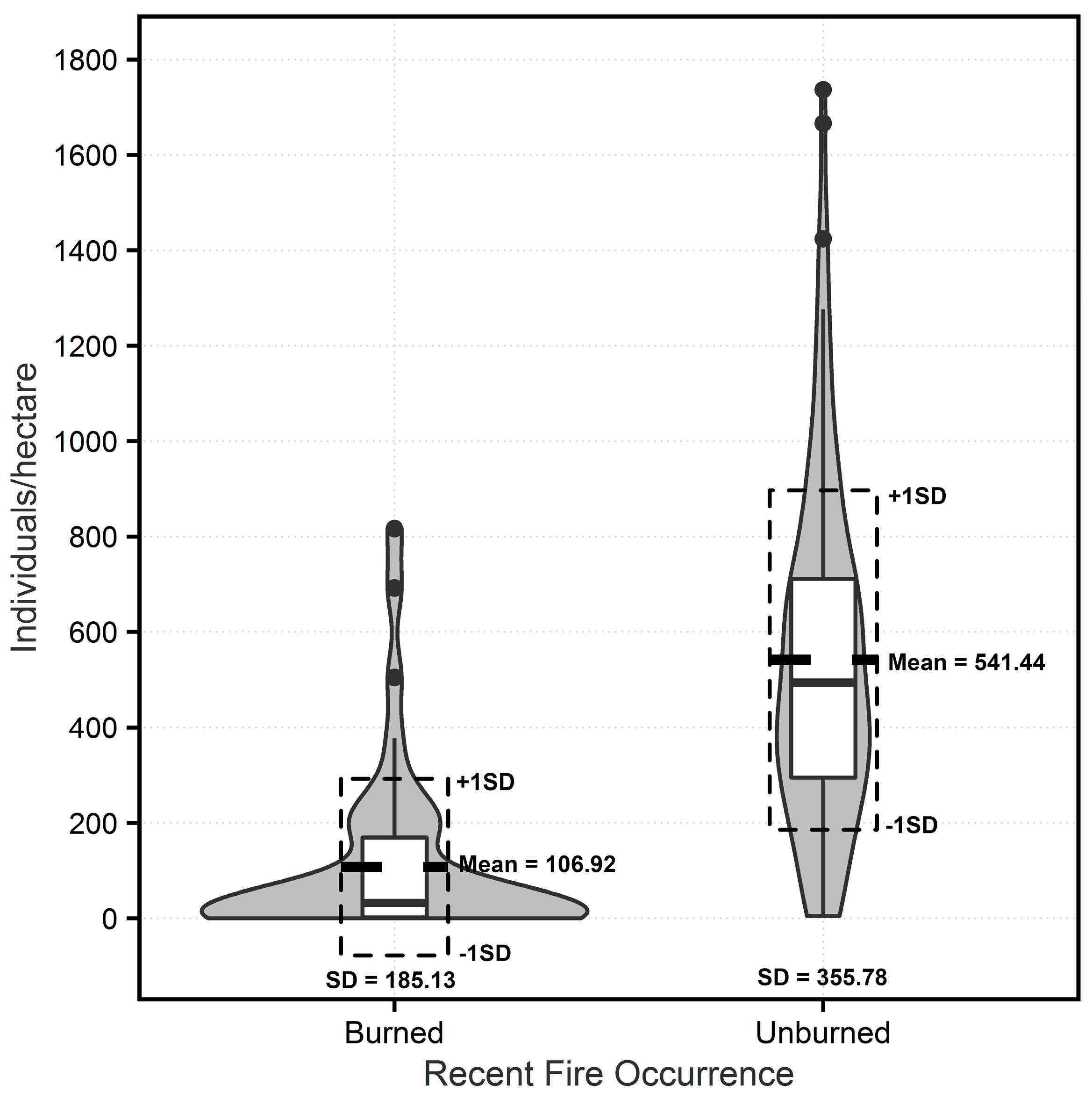
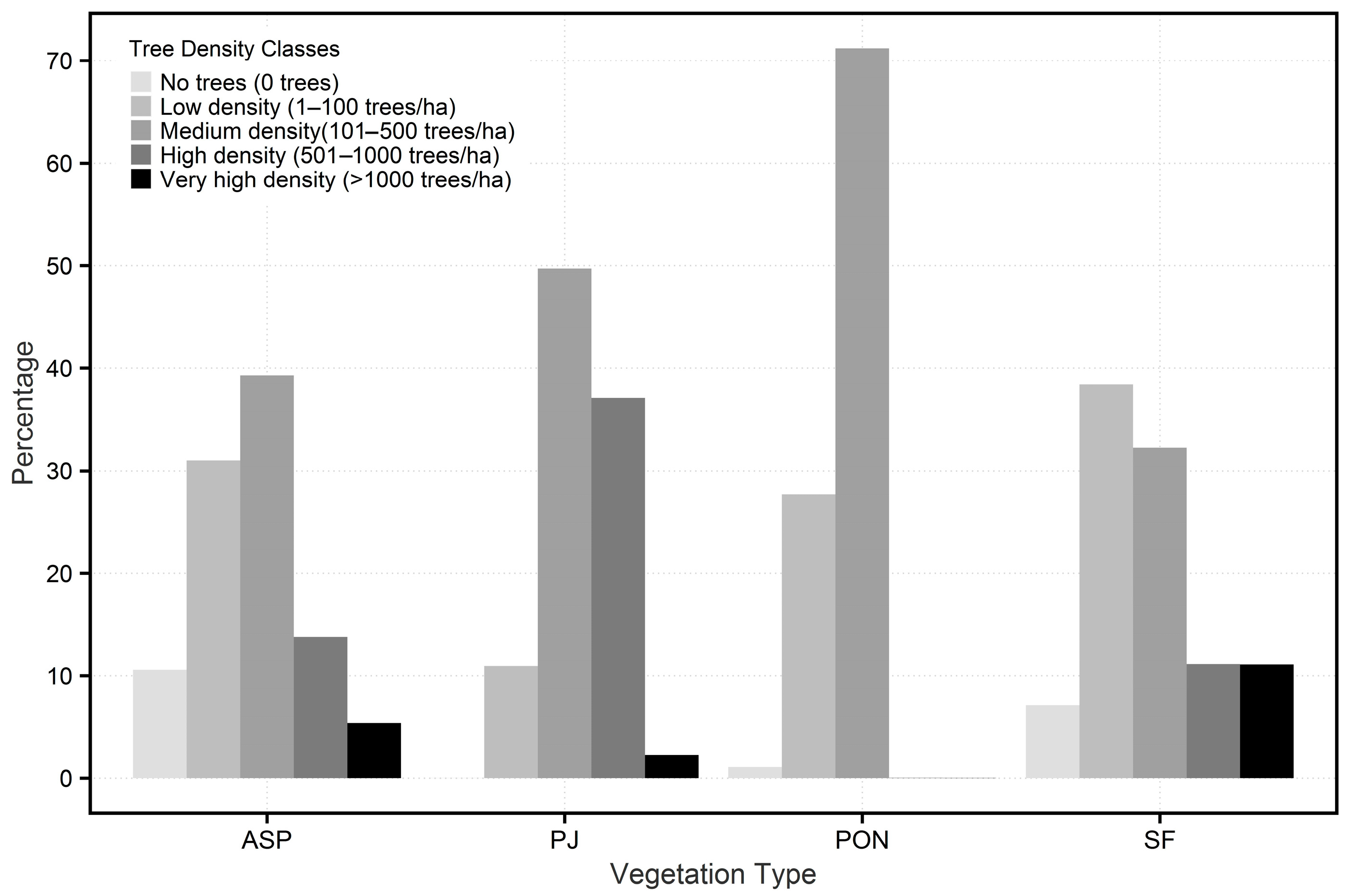
3.1. Measured Tree Density
3.2. Model Selection and Cross-Validation
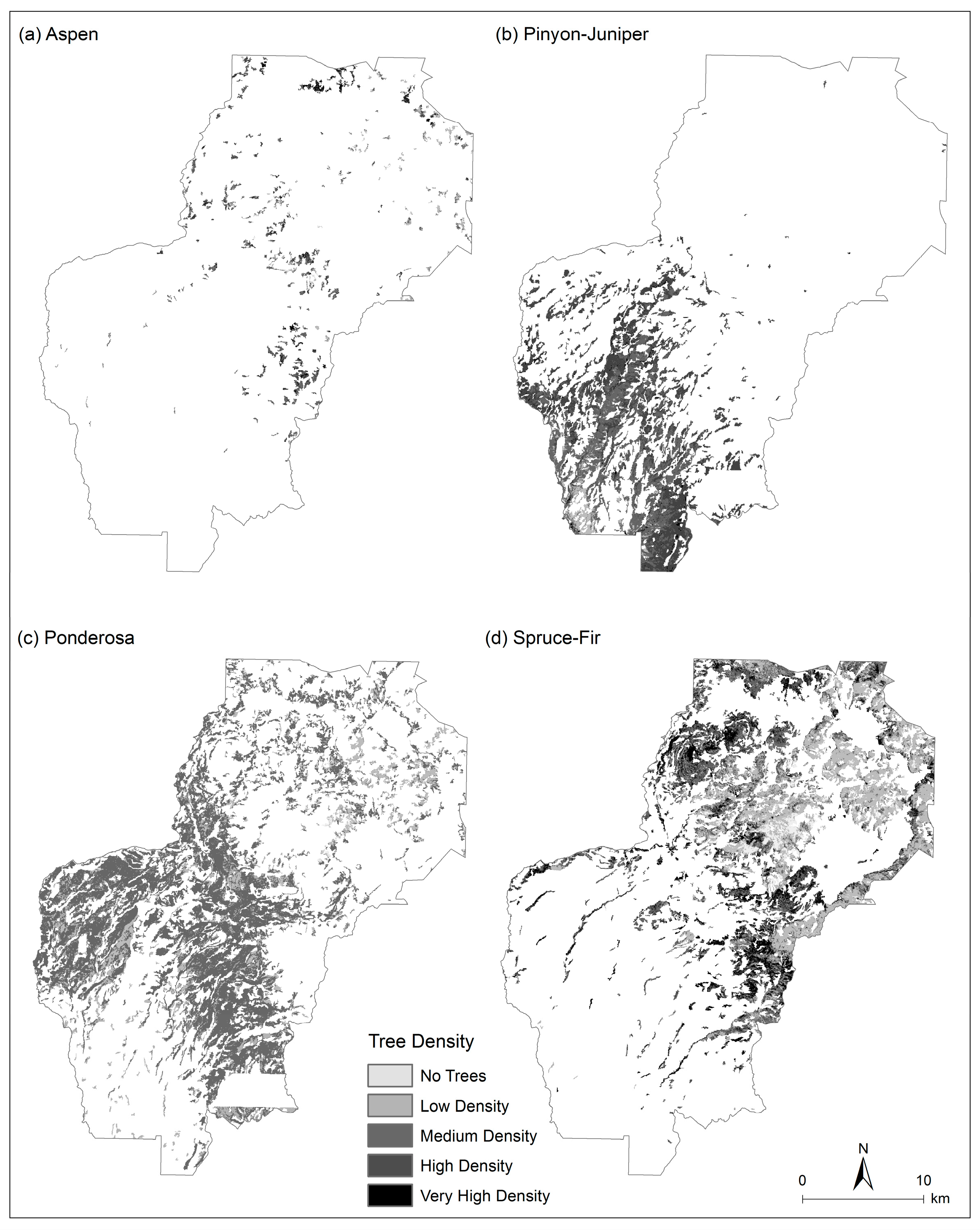
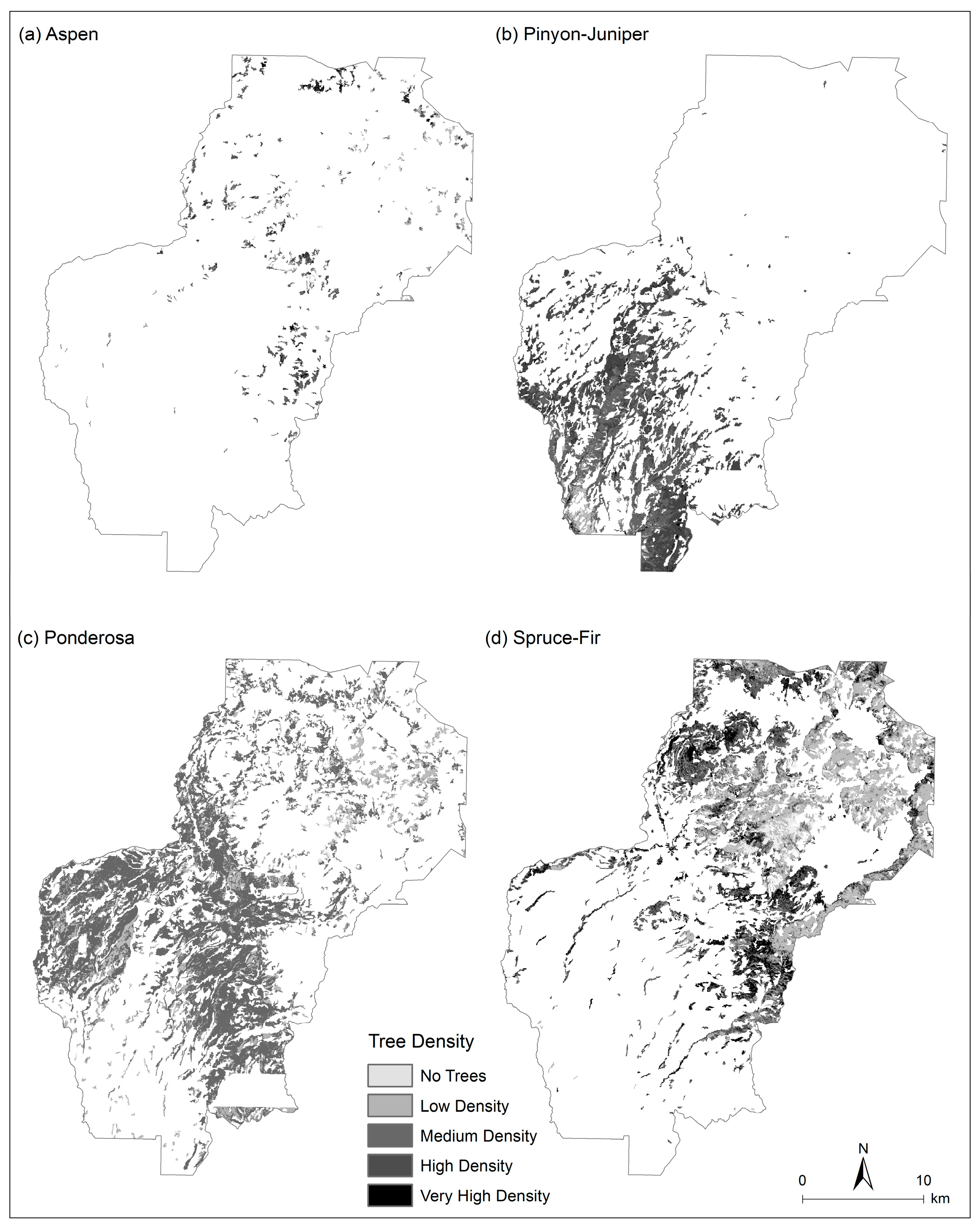
3.3. Tree Density Estimation
4. Discussion
4.1. Current Forest Conditions
4.2. Predicting Tree Density
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Roberts, M.R.; Gilliam, F.S. Patterns and mechanisms of plant diversity in forested ecosystems: Implications for forest management. Ecol. Appl. 1995, 5, 969–977. [Google Scholar] [CrossRef]
- Franklin, J.F.; Spies, T.A.; Van Pelt, R.; Carey, A.B.; Thornburgh, D.A.; Berg, D.R.; Lindenmayer, D.B.; Harmon, M.E.; Keeton, W.S.; Shaw, D.C.; et al. Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. For. Ecol. Manag. 2002, 155, 399–423. [Google Scholar] [CrossRef]
- Propastin, P. Relations between Landsat ETM+ imagery and forest structure parameters in tropical rainforests: A case study from Lore-Lindu National Park in Sulawesi, Indonesia. EARSeL eProc. 2009, 8, 93–106. [Google Scholar]
- Mohammadi, J.; Joibary, S.S.; Yaghmaee, F.; Mahiny, A.S. Modelling forest stand volume and tree density using Landsat ETM+ data. Int. J. Remote Sens. 2010, 31, 2959–2975. [Google Scholar] [CrossRef]
- Rocchini, D. Seeing the unseen by remote sensing: Satellite imagery applied to species distribution modelling. J. Veg. Sci. 2013, 24, 209–210. [Google Scholar] [CrossRef]
- Varga, P.; Chen, H.Y.; Klinka, K. Tree-Size diversity between single-and mixed-species stands in three forest types in western Canada. Can. J. For. Res. 2005, 35, 593–601. [Google Scholar] [CrossRef]
- McRoberts, R.E.; Winter, S.; Chirici, G.; Hauk, E.; Pelz, D.R.; Moser, W.K.; Hatfield, M.A. Large-Scale spatial patterns of forest structural diversity. Can. J. For. Res. 2008, 38, 429–438. [Google Scholar] [CrossRef]
- Legendre, P.; Dale, M.R.; Fortin, M.J.; Gurevitch, J.; Hohn, M.; Myers, D. The consequences of spatial structure for the design and analysis of ecological field surveys. Ecography 2002, 25, 601–615. [Google Scholar] [CrossRef]
- Edwards, T.C.; Cutler, D.R.; Zimmermann, N.E.; Geiser, L.; Moisen, G.G. Effects of sample survey design on the accuracy of classification tree models in species distribution models. Ecol. Model. 2006, 199, 132–141. [Google Scholar] [CrossRef]
- Heywood, V. Global Biodiversity Assessment; Cambridge University Press: Cambridge, UK, 1995; ISBN 9780521564816. [Google Scholar]
- Palmer, M.W. Species–Area curves and the geometry of nature. In Scaling Biodiversity, 1st ed.; Storch, D., Marquet, P.L., Brown, J.H., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 15–31. ISBN 9780521876025. [Google Scholar]
- Ozdemir, I.; Karnieli, A. Predicting forest structural parameters using the image texture derived from WorldView-2 multispectral imagery in a dryland forest, Israel. Int. J. Appl. Earth Obs. Geoinform. 2011, 13, 701–710. [Google Scholar] [CrossRef]
- Hernández-Stefanoni, J.L.; Gallardo-Cruz, J.A.; Meave, J.A.; Rocchini, D.; Bello-Pineda, J.; López-Martínez, J.O. Modeling α-and β-diversity in a tropical forest from remotely sensed and spatial data. Int. J. Appl. Earth Obs. Geoinform. 2012, 19, 359–368. [Google Scholar] [CrossRef]
- Kerr, J.T.; Ostrovsky, M. From space to species: Ecological applications for remote sensing. Trends Ecol. Evol. 2003, 18, 299–305. [Google Scholar] [CrossRef]
- Hudak, A.T.; Crookston, N.L.; Evans, J.S.; Falkowski, M.J.; Smith, A.M.; Gessler, P.E.; Morgan, P. Regression modeling and mapping of coniferous forest basal area and tree density from discrete-return lidar and multispectral satellite data. Can. J. Remote Sens. 2006, 32, 126–138. [Google Scholar] [CrossRef]
- Nagendra, H. Using remote sensing to assess biodiversity. Int. J. Remote Sens. 2001, 22, 2377–2400. [Google Scholar] [CrossRef]
- Palmer, M.W.; Earls, P.; Hoagland, B.W.; White, P.S.; Wohlgemuth, T. Quantitative tools for perfecting species lists. Environmetrics 2002, 13, 121–137. [Google Scholar] [CrossRef]
- Hernández-Stefanoni, J.; Dupuy, J. Mapping species density of trees, shrubs and vines in a tropical forest, using field measurements, satellite multispectral imagery and spatial interpolation. Biodivers. Conserv. 2007, 16, 3817–3833. [Google Scholar] [CrossRef]
- Feilhauer, H.; Schmidtlein, S. Mapping continuous fields of forest α- and β-diversity. Appl. Veg. Sci. 2009, 12, 429–439. [Google Scholar] [CrossRef]
- Jakubauskas, M.E.; Price, K.P. Empirical relationships between structural and spectral factors of Yellowstone lodgepole pine forests. Photogramm. Eng. Remote Sens. 1997, 63, 1375–1380. [Google Scholar]
- Smith, A.M.; Falkowski, M.J.; Hudak, A.T.; Evans, J.S.; Robinson, A.P.; Steele, C.M. A cross-comparison of field, spectral, and lidar estimates of forest canopy cover. Can. J. Remote Sens. 2009, 35, 447–459. [Google Scholar] [CrossRef]
- Kahriman, A.; Günlü, A.; Karahalil, U. Estimation of crown closure and tree density using Landsat TM satellite images in mixed forest stands. J. Indian Soc. Remote Sens. 2014, 42, 559–567. [Google Scholar] [CrossRef]
- Sivanpillai, R.; Smith, C.T.; Srinivasan, R.; Messina, M.G.; Wu, X.B. Estimation of managed loblolly pine stand age and density with Landsat ETM+ data. For. Ecol. Manag. 2006, 223, 247–254. [Google Scholar] [CrossRef]
- Tucker, C.J.; Fung, Y.; Keeling, C.D.; Gammon, R.H. Relationship between atmospheric CO2 variations and a satellite-derived vegetation index. Nature 1986, 319, 195–199. [Google Scholar] [CrossRef]
- Song, C.; Zhang, Y. Forest Cover in China from 1949 to 2006. In Reforesting Landscapes: Linking Pattern and Process, Landscape Series; Nagendra, H., Southworth, J., Eds.; Springer: Dordrecht, The Netherlands, 2010; Volume 10, pp. 341–356. ISBN 978-1-4020-9656-3. [Google Scholar]
- Gould, W. Remote sensing of vegetation, plant species richness, and regional biodiversity hotspots. Ecol. Appl. 2000, 10, 1861–1870. [Google Scholar] [CrossRef]
- Fairbanks, D.H.; McGwire, K.C. Patterns of floristic richness in vegetation communities of California: Regional scale analysis with multi-temporal NDVI. Glob. Ecol. Biogeogr. 2004, 13, 221–235. [Google Scholar] [CrossRef]
- Gillespie, T.W. Predicting woody-plant species richness in tropical dry forests: A case study from south Florida, USA. Ecol. Appl. 2005, 15, 27–37. [Google Scholar] [CrossRef]
- Rocchini, D.; Hernández-Stefanoni, J.L.; He, K.S. Advancing species diversity estimate by remotely sensed proxies: A conceptual review. Ecol. Inform. 2015, 25, 22–28. [Google Scholar] [CrossRef]
- Lu, D.; Mausel, P.; Brondızio, E.; Moran, E. Relationships between forests stand parameters and Landsat TM spectral responses in the Brazilian Amazon Basin. For. Ecol. Manag. 2004, 198, 149–167. [Google Scholar] [CrossRef]
- He, K.S.; Zhang, J.; Zhang, Q. Linking variability in species composition and MODIS NDVI based on beta diversity measurements. Acta Oecol. 2009, 35, 14–21. [Google Scholar] [CrossRef]
- Meng, R.; Dennison, P.E.; Huang, C.; Moritz, M.A.; D’Antonio, C. Effects of fire severity and post-fire climate on short-term vegetation recovery of mixed-conifer and red fir forests in the Sierra Nevada Mountains of California. Remote Sens. Environ. 2015, 171, 311–325. [Google Scholar] [CrossRef]
- Feilhauer, H.; He, K.S.; Rocchini, D. Modelling species distribution using niche-based proxies derived from composite bioclimatic variables and MODIS NDVI. Remote Sens. 2012, 4, 2057–2075. [Google Scholar] [CrossRef]
- Vacchiano, G.; Motta, R. An improved species distribution model for Scots pine and downy oak under future climate change in the NW Italian Alps. Ann. For. Sci. 2015, 72, 321–334. [Google Scholar] [CrossRef]
- Irisarri, J.G.N.; Oesterheld, M.; Paruelo, J.M.; Texeira, M.A. Patterns and controls of above-ground net primary production in meadows of Patagonia: A remote sensing approach. J. Veg. Sci. 2012, 23, 114–126. [Google Scholar] [CrossRef]
- Borowik, T.; Pettorelli, N.; Sönnichsen, L.; Jędrzejewska, B. Normalized difference vegetation index (NDVI) as a predictor of forage availability for ungulates in forest and field habitats. Eur. J. Wildl. Res. 2013, 59, 675–682. [Google Scholar] [CrossRef]
- Vogelmann, J.E.; Tolk, B.; Zhu, Z. Monitoring forest changes in the southwestern United States using multitemporal Landsat data. Remote Sens. Environ. 2009, 113, 1739–1748. [Google Scholar] [CrossRef]
- Madurapperuma, B.D.; Kuruppuarachchi, K.A.J.M. Detecting land-cover change using mappable vegetation related indices: A case study from Sinharaja Man and the Biosphere Reserve. J. Trop. For. Environ. 2014, 4, 50–58. [Google Scholar]
- Franklin, S.E.; Hall, R.J.; Smith, L.; Gerylo, G.R. Discrimination of conifer height, age and crown closure classes using Landsat-5 TM imagery in the Canadian Northwest Territories. Int. J. Remote Sens. 2003, 24, 1823–1834. [Google Scholar] [CrossRef]
- Freitas, S.R.; Mello, M.C.; Cruz, C.B. Relationships between forest structure and vegetation indices in Atlantic Rainforest. For. Ecol. Manag. 2005, 218, 353–362. [Google Scholar] [CrossRef]
- Fulé, P.Z.; Covington, W.W.; Moore, M.M. Determining reference conditions for ecosystem management of southwestern ponderosa pine forests. Ecol. Appl. 1997, 7, 895–908. [Google Scholar] [CrossRef]
- Moore, M.M.; Huffman, D.W.; Fulé, P.Z.; Covington, W.W.; Crouse, J.E. Comparison of historical and contemporary forest structure and composition on permanent plots in southwestern ponderosa pine forests. For. Sci. 2004, 50, 162–176. [Google Scholar]
- Covington, W.W.; Moore, M.M. Postsettlement changes in natural fire regimes and forest structure. J. Sustain. For. 1994, 2, 153–181. [Google Scholar] [CrossRef]
- Kenneth, L.; Dye, I.I.; Ueckert, D.N.; Whisenant, S.G. Redberry juniper-herbaceous understory interactions. J. Range Manag. 1995, 48, 100–107. [Google Scholar] [CrossRef]
- Touchan, R.; Allen, C.D.; Swetnam, T.W. Fire history and climatic patterns in ponderosa pine and mixed-conifer forests of the Jemez Mountains, northern New Mexico. In Fire Effects in Southwestern Forests, Proceedings of the Second La Mesa Fire Symposium RM-GTR-286, Los Alamos, New Mexico, USA, 29–31 March 1996; USDA-Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1996; pp. 33–46. [Google Scholar]
- Miller, C.; Urban, D.L. Modeling the effects of fire management alternatives on Sierra Nevada mixed-conifer forests. Ecol. Appl. 2000, 10, 85–94. [Google Scholar] [CrossRef]
- Lydersen, J.M.; Collins, B.M.; Miller, J.D.; Fry, D.L.; Stephens, S.L. Relating fire-caused change in forest structure to remotely sensed estimates of fire severity. Fire Ecol. 2016, 12, 99–116. [Google Scholar] [CrossRef]
- Mast, J.N.; Fulé, P.J.; Moore, M.M.; Covington, W.W.; Waltz, A.E.M. Restoration of presettlement age structure of an Arizona ponderosa pine forest. Ecol. Appl. 1999, 9, 228–239. [Google Scholar] [CrossRef]
- Kaye, J.P.; Hart, S.C.; Fulé, P.Z.; Covington, W.W.; Moore, M.M.; Kaye, M.W. Initial carbon, nitrogen, and phosphorus fluxes following ponderosa pine restoration treatments. Ecol. Appl. 2005, 15, 1581–1593. [Google Scholar] [CrossRef]
- Finkral, A.J.; Evans, A.M. The effects of a thinning treatment on carbon stocks in a northern Arizona ponderosa pine forest. For. Ecol. Manag. 2008, 255, 2743–2750. [Google Scholar] [CrossRef]
- Fulé, P.Z.; Crouse, J.E.; Cocke, A.E.; Moore, M.M.; Covington, W.W. Changes in canopy fuels and potential fire behavior 1880–2040: Grand Canyon, Arizona. Ecol. Model. 2004, 175, 231–248. [Google Scholar] [CrossRef]
- Gass, T.M.; Robinson, A.P. A hierarchical analysis of stand structure, composition, and burn patterns as indicators of stand age in an Engelmann spruce–subalpine fir forest. Can. J. For. Res. 2007, 37, 884–894. [Google Scholar] [CrossRef]
- Hayes, J.J.; Robeson, S.M. Spatial variability of landscape pattern change following a ponderosa pine wildfire in northeastern New Mexico, USA. Phys. Geogr. 2009, 30, 410–429. [Google Scholar] [CrossRef]
- Lydersen, J.M.; North, M.P.; Collins, B.M. Severity of an uncharacteristically large wildfire, the Rim Fire, in forests with relatively restored frequent fire regimes. For. Ecol. Manag. 2014, 328, 326–334. [Google Scholar] [CrossRef]
- Stevens, J.T.; Safford, H.D.; North, M.P.; Fried, J.S.; Gray, A.N.; Brown, P.M.; Dolanc, C.R.; Dobrowski, S.Z.; Falk, D.A.; Farris, C.A.; et al. Average stand age from forest inventory plots does not describe historical fire regimes in ponderosa pine and mixed-conifer forests of western North America. PLoS ONE 2016, 11, e0147688. [Google Scholar] [CrossRef] [PubMed]
- Butera, M.K. A correlation and regression analysis of percent canopy closure versus TMS spectral response for selected forest sites in the San Juan National Forest, Colorado. IEEE Trans. Geosci. Remote Sens. 1986, 1, 122–129. [Google Scholar] [CrossRef]
- Cohen, W.B.; Spies, T.A. Estimating structural attributes of Douglas Fir-western hemlock forest stands from Landsat and SPOT imagery. Remote Sens. Environ. 1992, 41, 1–17. [Google Scholar] [CrossRef]
- Fiorella, M.; Ripple, W.J. Analysis of conifer forest regeneration using Landsat Thematic Mapper data. Photogramm. Eng. Remote Sens. 1993, 59, 1383–1388. [Google Scholar]
- Peterson, D.L.; Westman, W.E.; Stephenson, N.J.; Ambrosia, V.G.; Brass, J.A.; Spanner, M.A. Analysis of forest structure using Thematic Mapper Simulator data. IEEE Trans. Geosci. Remote Sens. 1986, 24, 113–121. [Google Scholar] [CrossRef]
- Oladi, J. Developing diameter at breast height (DBH) and a height estimation model from remotely sensed data. J. Agric. Sci. 2005, 7, 95–102. [Google Scholar]
- Coop, J.D.; Givnish, T.J. Spatial and temporal patterns of recent forest encroachment in montane grasslands of the Valles Caldera, New Mexico, USA. J. Biogeogr. 2007, 34, 914–927. [Google Scholar] [CrossRef]
- Zapata-Rios, X.; McIntosh, J.; Rademacher, L.; Troch, P.A.; Brooks, P.D.; Rasmussen, C.; Chorover, J. Climatic and landscape controls on water transit times and silicate mineral weathering in the critical zone. Water Resour. Res. 2015, 51, 6036–6051. [Google Scholar] [CrossRef]
- Dick-Peddie, W.A.; Moir, W.H.; Spellenberg, R. New Mexico Vegetation: Past, Present, and Future; University of New Mexico Press: Albuquerque, NM, USA, 1999; ISBN 978-0-8263-2164-0. [Google Scholar]
- SWJ-CFLRP. Southwest Jemez Mountains Collaborative Forest Landscape Restoration Proposal for Funding; Santa Fe National Forest and Valles Caldera National Preserve: Jemez Springs, NM, USA, 2010. [Google Scholar]
- Reynolds, R.T.; Sanchez Meador, A.J.; Youtz, J.A.; Nicolet, T.; Mantonis, M.S.; Jackson, P.L.; DeLorenzo, D.G.; Graves, A.D. Restoring Composition and Structure in Southwestern Frequent-Fire Forests: A Science-Based Framework for Improving Ecosystem Resiliency; Gen. Tech. Rep. RMRSGTR-310; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2013; 76 p. [Google Scholar]
- Mitchell, K. Quantitative Analysis by the Point-Centered Quarter Method. Hobart and William Smith Colleges: Geneva, NY, USA. arXiv. 2015. Available online: https://arxiv.org/abs/1010.3303 (accessed on 25 August 2015).
- Bryant, D.M.; Ducey, M.J.; Innes, J.C.; Lee, T.D.; Eckert, R.T.; Zarin, D.L. Forest community analysis and the point-centered quarter method. Plant Ecol. 2005, 175, 193–203. [Google Scholar] [CrossRef]
- Bhardwaj, A.; Joshi, P.K.; Sam, L.; Singh, M.K.; Singh, S.; Kumar, R. Applicability of Landsat 8 data for characterizing glacier facies and supraglacial debris. Int. J. Appl. Earth Obs. Geoinf. 2015, 38, 51–56. [Google Scholar] [CrossRef]
- USGS. Landsat 8 (L8) Data Users Handbook, version 1.0 (LSDS-1574); United States Geological Survey, Earth Resources Observation System (EROS): Sioux Falls, SD, USA, 2015. [Google Scholar]
- Ding, Y.; Zhao, K.; Zheng, X.; Jiang, T. Temporal dynamics of spatial heterogeneity over cropland quantified by time-series NDVI, near infrared and red reflectance of Landsat 8 OLI imagery. Int. J. Appl. Earth Obs. Geoinf. 2014, 30, 139–145. [Google Scholar] [CrossRef]
- Dube, T.; Mutanga, O. Evaluating the utility of the medium-spatial resolution Landsat 8 multispectral sensor in quantifying aboveground biomass in uMgeni catchment, South Africa. ISPRS J. Photogramm. Remote Sens. 2015, 101, 36–46. [Google Scholar] [CrossRef]
- Scheiner, S.; Gurevitch, J. Design and Analysis of Ecological Experiments; Chapman and Hall: New York, NY, USA, 1998; ISBN 10:0412035618. [Google Scholar]
- Venables, W.N.; Smith, D.M. An introduction to R; Network Theory Limited: East Sussex, UK, 2009; ISBN 0954612086. [Google Scholar]
- United States Department of Agriculture Forest Service, Santa Fe National Forest GIS Data. Available online: https://www.fs.usda.gov/detail/r3/landmanagement/gis/?cid=stelprdb5203736 (accessed on 10 September 2015).
- Stauffer, H.B. Contemporary Bayesian and Frequentist Statistical Research Methods for Natural Resource Scientists; John Wiley & Sons: Hoboken, NJ, USA, 2007; ISBN 978-0-470-16504-1. [Google Scholar]
- Dobrowski, S.Z.; Safford, H.D.; Cheng, Y.B.; Ustin, S.L. Mapping mountain vegetation using species distribution modeling, image-based texture analysis, and object-based classification. Appl. Veg. Sci. 2008, 11, 499–508. [Google Scholar] [CrossRef]
- Yang, L.; Huang, C.; Homer, C.G.; Wylie, B.K.; Coan, M.J. An approach for mapping large-area impervious surfaces: Synergistic use of Landsat-7 ETM+ and high spatial resolution imagery. Can. J. For. Res. 2003, 29, 230–240. [Google Scholar] [CrossRef]
- Allen, C.D. A ponderosa pine natural area reveals its secrets. In Status and Trends of the Nation’s Biological Resources; Opler, P.A., Haecker, C.E., Eds.; US Geological Survey: Reston, VA, USA, 1998; Volume 2, pp. 551–552. [Google Scholar]
- Sisk, T.D.; Prather, J.W.; Hampton, H.M.; Aumack, E.N.; Xu, Y.; Dickson, B.G. Participatory landscape analysis to guide restoration of ponderosa pine ecosystems in the American Southwest. Landsc. Urban Plan. 2006, 78, 300–310. [Google Scholar] [CrossRef]
- Rodman, K.C.; Sánchez Meador, A.J.; Huffman, D.W.; Waring, K.M. Reference conditions and historical fine-scale spatial dynamics in a dry mixed-conifer forest, Arizona, USA. For. Sci. 2016, 62, 268–280. [Google Scholar] [CrossRef]
- Darvishzadeh, R.; Skidmore, A.; Atzberger, C.; van Wieren, S. Estimation of vegetation LAI from hyperspectral reflectance data: Effects of soil type and plant architecture. Int. J. Appl. Earth Obs. Geoinform. 2008, 10, 358–373. [Google Scholar] [CrossRef]
- Verhoef, W.; Bach, H. Coupled soil–leaf-canopy and atmosphere radiative transfer modeling to simulate hyperspectral multi-angular surface reflectance and TOA radiance data. Remote Sens. Environ. 2007, 109, 166–182. [Google Scholar] [CrossRef]
- Jensen, J.R. Remote Sensing of Environment: An Earth Resource Perspective; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2007; ISBN 0131889508. [Google Scholar]
- Song, C.; Woodcock, C.E. Monitoring forest succession with multitemporal Landsat images: Factors of uncertainty. IEEE Trans. Geosci. Remote Sens. 2003, 41, 2557–2567. [Google Scholar] [CrossRef]
- North, M.; Innes, J.; Zald, H. Comparison of thinning and prescribed fire restoration treatments to Sierran mixed-conifer historic conditions. Can. J. For. Res. 2007, 37, 331–342. [Google Scholar] [CrossRef]
- Korb, J.E.; Fulé, P.Z.; Stoddard, M.T. Forest restoration in a surface fire-dependent ecosystem: An example from a mixed conifer forest, southwestern Colorado, USA. For. Ecol. Manag. 2012, 269, 10–18. [Google Scholar] [CrossRef]
- Erickson, C.C.; Waring, K.M. Old Pinus ponderosa growth responses to restoration treatments, climate and drought in a southwestern US landscape. Appl. Veg. Sci. 2014, 17, 97–108. [Google Scholar] [CrossRef]
- Cottam, G.; Curtis, J.T. The use of distance measures in phytosociological sampling. Ecology 1956, 37, 451–460. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Types * | AICc | R2 | Adj R2 | MS ** | Models *** |
|---|---|---|---|---|---|
| All | 681.90 | 0.24 | 0.23 | 3.82 | d^ = 4.03 + 5.89 ND57 |
| (n = 178) | 629.70 | 0.44 | 0.43 | 3.05 | d^ = 7.96 + 17.72 ND57 − 2.50SR1 |
| 623.80 | 0.46 | 0.45 | 3.04 | d^ = 7.54 + 18.60 ND57 − 3.17 SR1 + 17.64 DVI | |
| ASP | 41.90 | 0.92 | 0.91 | 0.59 | d^ = 3.50 + 19.94 ND57 − 2.26 SR3 |
| (n = 16) | 41.10 | 0.94 | 0.92 | 0.57 | d^ = 1.19 + 24.76 ND57 − 2.70 SR3 + 26.86 band4 |
| 33.40 | 0.97 | 0.96 | 0.36 | d^ = 21.06 + 21.16 ND57 − 1.99 SR3 + 284.01 band4 − 478.56 band2 | |
| P-J | 105.30 | 0.34 | 0.32 | 1.1 | d^ = 8.63 − 22.73 band4 |
| (n = 36) | 96.80 | 0.52 | 0.49 | 0.86 | d^ = 3.25 − 50.03 band4 + 84.74 band2 |
| PON | 207.00 | 0.35 | 0.32 | 1.26 | d^ = 7.88 − 47.02 band7 + 66.15 band3 − 0.87 SR3 |
| (n = 67) | 200.00 | 0.43 | 0.40 | 1.15 | d^ = 10.59 − 11.50 band7 + 9.50 band3 − 5.71 SR3 + 23.32 ND57 |
| 199.00 | 0.46 | 0.42 | 1.10 | d^ = 13.51 + 4.93 band7 − 24.93 band3 − 6.39 SR3 + 28.23 ND57 − 0.95 SR2 | |
| S-F | 226.80 | 0.60 | 0.57 | 3.07 | d^ = 3.87 + 2.14 ND57 − 6.12 SR1 + 38.20 NDVI − 1.78 SR2 + 2.57 SR3 |
| (n = 59) | 225.80 | 0.63 | 0.59 | 2.81 | d^ = 2.85 − 1.32 ND57 − 7.87 SR1 + 41.36 NDVI − 1.82 SR2 + 4.24 SR3 + 21.87 DVI |
| 223.80 | 0.66 | 0.62 | 4.42 | d^ = 6.67 − 11.46 ND57 − 8.68 SR1 + 37.32 NDVI − 1.81 SR2 + 5.58 SR3 + 55.51 DVI − 29.87 band7 |
| Tree Density Class ↓ | Area (%) * | ||||
|---|---|---|---|---|---|
| Area (ha) → | Fire 2011 (9962.46) | Fire 2013 (8402.94) | Treated (670.23) | Planned (5793.48) | Not Treated (45361.71) |
| No trees | 7.59 | 16.45 | 0.26 | 0.36 | 0.43 |
| Low density (1–100 trees/ha) | 62.90 | 44.54 | 45.29 | 18.35 | 16.08 |
| Medium density (101–500 trees/ha) | 23.69 | 32.40 | 42.22 | 64.08 | 56.82 |
| High density (501–1000 tress/ha) | 3.14 | 4.02 | 5.48 | 14.45 | 19.86 |
| Very high density (>1000 trees/ha) | 2.68 | 2.59 | 6.75 | 2.77 | 6.81 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Humagain, K.; Portillo-Quintero, C.; Cox, R.D.; Cain, J.W., III. Mapping Tree Density in Forests of the Southwestern USA Using Landsat 8 Data. Forests 2017, 8, 287. https://doi.org/10.3390/f8080287
Humagain K, Portillo-Quintero C, Cox RD, Cain JW III. Mapping Tree Density in Forests of the Southwestern USA Using Landsat 8 Data. Forests. 2017; 8(8):287. https://doi.org/10.3390/f8080287
Chicago/Turabian StyleHumagain, Kamal, Carlos Portillo-Quintero, Robert D. Cox, and James W. Cain, III. 2017. "Mapping Tree Density in Forests of the Southwestern USA Using Landsat 8 Data" Forests 8, no. 8: 287. https://doi.org/10.3390/f8080287
APA StyleHumagain, K., Portillo-Quintero, C., Cox, R. D., & Cain, J. W., III. (2017). Mapping Tree Density in Forests of the Southwestern USA Using Landsat 8 Data. Forests, 8(8), 287. https://doi.org/10.3390/f8080287