Responses of Fine Root Functional Traits to Soil Nutrient Limitations in a Karst Ecosystem of Southwest China

1
Key Laboratory of Agro-ecological Processes in Subtropical Region, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, China
2
College of Environmental Science and Engineering, Guilin University of Technology, Guilin 541004, China
3
Karst Dynamics Laboratory, Ministry of Natural Resources, Institute of Karst Geology, Chinese Academy of Geological Sciences, Guilin 541004, China
4
Huanjiang Observation and Research Station for Karst Ecosystems, Huanjiang 547100, China
*
Author to whom correspondence should be addressed.
Forests 2018, 9(12), 743; https://doi.org/10.3390/f9120743
Submission received: 15 October 2018
/
Revised: 10 November 2018
/
Accepted: 21 November 2018
/
Published: 28 November 2018
(This article belongs to the Special Issue The Ecology of Fine Roots and Mycorrhizas in Forests)
Abstract
:Soil nitrogen (N) and phosphorus (P) shortages limit the growth of shrubs, and P shortage limit the growth of trees in karst ecosystems. Changes in fine root functional traits are the important strategies for plants to respond to such nutrient shortages. However, such responses in karst ecosystems are poorly known. To determine the responses of fine root functional traits to soil N and P changes and define their resource-use strategies in the ecosystem, we tested the specific root length (SRL), root tips over the root biomass (RT/RB), and N concentration (Nroot) in the fine roots of four plant species (two shrubs (Alchornea trewioides and Ligustrum sinense) and two trees (Celtis biondii and Pteroceltis tatarinowii)) during the dry (January) and the wet (July) season. The results showed that the SRL, RT/RB, and Nroot in the fine roots of shrub species were lower than those of tree species, and the three parameters were higher in the wet season than in the dry season. Linear regression models revealed that the SRL, RT/RB, and Nroot of overall species increased with increasing soil N and P concentrations and availabilities, and were positively correlated with increasing rhizosphere soil oxalic acid, microbial biomass carbon (C), and the activities of hydrolytic enzymes. In addition, the individual plant species had unique patterns of the three fine root traits that resulted affected by the change of soil nutrients and biochemistry. Thus, the specific root length, root tips over the root biomass, and N concentrations of fine roots were species-specific, affected by seasonal change, and correlated with soil nutrients and biochemistry. Our findings suggests that fine root functional traits increase the ability of plant species to tolerate nutrient shortage in karst ecosystems, and possibly indicated that a P-exploitative strategy in tree species and an N-conservative strategy in shrub species were exhibited.
1. Introduction
The karst landscape of southwestern China encompasses 540,000 km2 and is one of the largest karst regions worldwide [1,2]. Characteristics of this region include peak-cluster depression, caves, sinkholes, rocky desertification, alkaline calcareous soils, and specialized regional vegetation [3,4,5]. Due to the unique topographical factors, the ecological systems in this region are fragile and extremely vulnerable to human activities. During recent decades, the rising human population and the resultant overexploitation of natural resources have accelerated vegetation degradation [6,7]. Thus, the vegetation restoration of degraded landscapes in this region has been a major concern of governments and researchers. However, vegetation restoration is mainly limited by soil nitrogen (N) and phosphorus (P) availabilities. In comparison to ecosystems at the same latitude, karst regions have a higher total soil nutrient content but a lower availability of soil nutrients due to the high calcium and magnesium content [8]. Additionally, available nutrients are easily leached into the underground drainage networks as a result of the shallow soil cover and the highly developed karst fissures [9], which exacerbate the nutrient limitations in the ecosystem.
In our previous study, we found that soil N and P contents and availabilities are increasing with the advancing vegetation successions in karst ecosystems [10], as well as the forest net primary production [11]. Thus, the shrub species are affected by soil N and P limitations, while the tree species are mainly influenced by soil P limitations in this ecosystem [12]. In order to tolerate the nutrient-limited stresses, plants have developed a wide variety of nutrient acquisition strategies, such as the resorption of nutrients from senescent foliage [7], an enhanced genome size [13], symbiosis of roots with arbuscular mycorrhizal fungi, and N-fixing bacteria [14,15,16], or priming by fine roots [17,18]. Besides, the plasticity of fine roots is another effective strategy of plants to respond to soil nutrient limitations [19].
Fine roots, which are generally defined as roots less than two mm in diameter, are essential in plant resource acquisition and are linked to plant growth [20]. With their simple anatomical organization [21,22], fine roots can rapidly respond to environment changes [23]. Particularly, some fine root traits, including specific root length (SRL), number of root tips, N concentrations (Nroot), and so on, can be considered as functions of a plant’s plasticity to acquire nutrient resources [24]. Plants may increase their fine root length, surface area, and number of root tips to better absorb nutrients [25]. The responses of these traits to resources are closely correlated with intrinsic and extrinsic factors. (1) The variations of fine root functional traits are species-specific. In natural mixed forests, the fine roots of the neighboring plants are frequently overlapping, resulting in high interspecific belowground competition [26]. In order to response to resources competition, the root systems of neighboring species would increase their SRL to exploit more soil volumes [27], or increase their root tips to enhance their resource uptake capacity [28]. Some studies have shown that plant species that have high fine root SRL and large amounts of root tips, also have rapid resource acquisition and high nutrients contents [19,29,30,31]. However, some studies have shown that plants with lower fine root SRL are associated with higher Nroot [25,32,33]. This discrepancy between SRL, root tips, and nutrient content may be closely linked to different nutrient strategies between plant species [24]. (2) The variations of these traits are also related to soil environment changes, particularly seasonal changes of soil nutrients availabilities, water, and temperature. In nutrient-limited soils, one study showed that a higher SRL occurs in the more fertile soils [34]. A high SRL can potentially maximize the nutrient uptake rates of plants in the more fertile soils [35]. Generally, nutrient availabilities are higher in the wet season than in the dry season, because it relates to high soil temperature, water, microbes, and enzyme activities [10]. In recent years, Montagnoli et al. [36,37,38] have found that the SRL and other traits are positively correlated with increasing soil temperature and water. Thus, the SRL is related with the increasing interactions of soil nutrients availabilities, water, and temperature. (3) The fine root functional traits will change in different vegetation successions. In natural ecosystems, a strong P limitation frequently occurs in the late stage of vegetation successions. High SRL has been found in this P-limited stage, and it increases with the increasing site age [39].
Approximately a decade ago, the nutrient strategies of plants were widely defined by the leaf economics spectrum [40,41]. Plants with a high specific leaf area, leaf dry matter content, long leaf lifespan, rapid nutrient turnover, and leaf growth were considered exploitative species with nutrient acquisition strategies. In recent years, some researchers have made great efforts to diagnose the nutrient acquisition strategies of some plants by the relationship between fine root traits and nutrient changes as well as the leaf economics spectrum [19,20]. The SRL is the root length/biomass ratio, and can be suggested as an index of root benefit/cost ratio [36]. It have been found that plants increase their fine root growth, mass, and ectomycorrhizal root tips when soil nutrients are decreasing in the relatively fertile soils [42]. However, these traits may increase with the increasing soil nutrients in the nutrient-limited soils [35,43]. The large root system may exploit many more soil volumes and tolerate the stress of nutrient limitations. Thus, plants with a high SRL are suggested as the nutrient exploitative species, and those that have low SRL are the conservative species [36]. Thus, it is important to understand fine root functional traits and their relevance to environmental adaptation in order to be able to assess how plants tolerate nutrient constraints in nutrient-limited ecosystems. In addition, organic acids and hydrolytic enzymes derived from roots and microbes can enhance the release of nutrients from soil organic matter [17], and may be related to the responses of fine root traits to soil nutrient limitations. However, the nutrient acquisition strategies, as well as the responses of fine root traits to soil nutrients availabilities, organic acids and hydrolytic enzymes, and seasonal changes, are poorly understood in the nutrient-limited soils of karst ecosystems.
In this study, our objective was to explore the patterns of fine root functional traits and their response to changes of soil nutrients and seasons in karst ecosystems. Four native plant species, i.e., two shrub species (A. trewioides and L. sinense) and two tree species (C. biondii and P. tatarinowii) [44,45,46,47], were selected as the study species from the ecosystem. The selected shrub species are co-limited by soil N and P shortage, but the tree species are mainly limited by soil P shortage [12]. We measured the specific root length (SRL), root tips over the root biomass (RT/RB), and N concentrations (Nroot) in fine roots, the oxalic acid, microbial biomass carbon (MBC) concentrations, and the activity of β-1,4-N-acetylglucosaminidase (NAG) in rhizosphere soils, and the total N (TN), total P (TP), available N (AN), and available P (AP) in bulk soils of the four study species in the dry (January) and the wet (July) season. Generally, in the nutrient-limited soils, the fine root functional traits would increase with the increasing soil nutrients and advancing successions. We hypothesized that (1) the fine roots of shrubs would have lower SRL, RT/RB, and Nroot than do trees; (2) the three assessed parameters would be higher in the wet season than in the dry season; and (3) these parameters would have positive relationships with increasing soil nutrients.
2. Materials and Methods
2.1. Study Site
This study was conducted at the Huanjiang Observation and Research Station for Karst Ecosystems under the Chinese Academy of Sciences (24°43′58.9″–24°44′48.8″ N, 108°18′56.9″–108°19′58.4″ E) and Mulun National Natural Reserve (25°06′–25°12′ N, 107°53′–108°05′ E) in Huanjiang county, Guangxi Zhuang Autonomous Region, in southwest China. The area of the Huanjiang Observation and Research Station experienced severe deforestation from 1958 to the mid-1980s, and was under natural restoration for almost 30 years. Nowadays, shrubs, including A. trewioides, L. sinense, Cipadessa baccifera, Rhus chinensis, and Indigofera atropurpurea as the common species, dominated ~70% of the hillslopes. The Mulun National Natural Reserve was established in 1991 to protect the remnants of undisturbed mixed evergreen and deciduous broadleaved forests in the karst region. The primary forest, which is dominated by C. biondii, Cleidion bracteosum, Cyclobalanopsis glauca, Cryptocarya chinensis, Loropetalum chinense, Miliusa chunii, and P. tatarinowii, has not been disturbed for over 200 years at this site. Both sites experience a typical subtropical monsoon climate, with a mean annual temperature of approximately 19 °C and a mean annual precipitation of 1400–1500 mm, most of which falls from May to September. Both sites consist of a typical karst landscape in a gentle valley flanked by steeper hills (Figure 1). Soil pH at the study sites ranged from 6.3 to 7.9. Soil depth averaged 50–80 cm in the depressions and 10–30 cm on the hillslopes [18].
2.2. Sampling
In this study, we selected two dominant shrub species (A. trewioides and L. sinense) from the shrublands and two dominant tree species (C. biondii and P. tatarinowii) from the primary forest. Samples of fine roots and associated soils were collected in January (dry season, which is from October each year to March of the next) and July (wet season, which is from April to September) 2014. In order to exclude the effects of slope positioning on fine root functional traits in the same vegetation successive stage, sampling operation was conducted on lower, middle, and upper slope positions [48]. In addition, in order to minimize the effects of interactions that might occur between plants in each slope position, shrubs and trees were separated by approximately 50 cm and 10 m, respectively.
The fine roots and soils of each individual plant of each species were collected from the topsoil at 0–15 cm depth with the excavation method, and the intact segments were extracted with a shovel. The fine roots were then lightly and carefully hand shaken to remove bulk soil after loosening. The remaining soil on the roots, i.e., the rhizosphere soil [49], was also lightly loosened (to minimize the losing of fine roots in the sampled soil) and carefully brushed into a separate polyethylene bag in the field. The rhizosphere soils and fine roots were immediately transported on ice to the laboratory [50]. In the laboratory, fine root samples were washed and then frozen at −20 °C for analysis at a later date [51]. Some broken roots were cleared from the rhizosphere soil samples within four hours of collection, and the soil samples were stored at −20 °C for subsequent analysis with oxalic acid, microbial biomass carbon (MBC), and the activity of β-1,4-N-acetylglucosaminidase (NAG). A total of 87 rhizosphere soil samples, i.e., four plant species × three slope positions × two seasons × four replicates (three replicates if we could not find a fourth individual at a particular site) were collected, as well as fine root samples.
Bulk soil samples were also collected. At ~50 cm away from each sampling point of rhizosphere soils, five soil cores (five-cm diameter, 0–15 cm depth) were collected and then pooled together. This sampling strategy of bulk soil samples ensured that they were representative of the soil environment for each plant. A total 87 bulk soil samples were collected and air-dried.
2.3. Laboratory Analyses
2.3.1. SRL, RT/RB, and Nroot of Fine Roots
Fine root samples were placed in tap water in a transparent plastic tray and scanned at 400 dpi using a flatbed scanner (Epson Expression 10000XL, Epson (China) Co. Ltd, Beijing, China) to obtain images for analysis and comparison. The length and number of root tips were measured using the WinRHIZO image analysis software (Regent Instrument Inc., Sainte-Foy, Quebec, Canada). After scanning, each root sample was oven-dried at 65 °C for at least 48 h to constant weight. The SRL and RT/RB values were calculated using the following equations:
SRL (m g−1) = fine root length/fine root biomass
RT/RB (number mg−1) = fine root tips/fine root biomass
Finally, the fine root samples were ground to pass through a 0.154-mm mesh sieve for further elemental analysis. The N concentrations (mg g−1) of fine roots were determined with an elemental analyzer (vario MAX CN, Elementar Analysensysteme GmbH, Langenselbold, Germany).
2.3.2. Oxalic Acid Concentrations of Rhizosphere Soils
Firstly, 2.5 g of soil were added to five mL of 0.05 mol L−1 phosphate buffer, and then were homogenized for 10 min with a homogenizer to prepare sample suspensions. Secondly, the suspended samples were centrifuged for 10 min at 27,200× g by an ultracentrifuge (CR22 GII; Hitachi Koki Co., Ltd, Kyoto, Japan). Thirdly, the liquid supernatants were purified by a 0.22-μm filtering membrane. Finally, the oxalic acid concentrations of the purified liquid were measured using high-performance liquid chromatography (1260 Infinity LC; Agilent Technologies, Santa Clara, CA, USA) [52].
2.3.3. β-1,4-N-acetylglucosaminidase (NAG) Activities of Rhizosphere Soils
2.3.4. Soil MBC of Rhizosphere Soils
The automatic analysis of chloroform fumigated-K2SO4 extracted carbon (C) was used to measure the rhizosphere soil MBC [55]. Fumigated and unfumigated samples (10 g of soil) were extracted with 40 mL of 0.5 mol L−1 K2SO4, and then the extractable organic C was analyzed using a total organic C analyzer (Shimadzu TOC-Vwp; Shimadzu Corporation, Kyoto, Japan).
2.3.5. Chemical Properties of Rhizosphere and Bulk Soils
Soil total nitrogen (TN) was measured using the Kjeldahl method; available nitrogen (AN) was measured using titration with a diluted solution of H2SO4 after extraction with a mixture of FeSO4 and NaOH; total phosphorus (TP) was digested in a solution of NaOH + H2SO4, and available phosphorus (AP) was extracted with NaHCO3, and the two parameters were analyzed with phosphomolybdate blue [47] (Table 1).
2.3.6. Soil Temperature and Moisture Contents
The database (from 2008 to 2015) of soil temperature and volumetric moisture contents (10 cm depth) was collected from one automatic monitoring system in each site (Figure 2).
2.4. Statistical Analyses
Data treatments and calculations were performed using R x64 3.2.2 statistical software [56]. The homogeneity of variance was tested prior to the statistical analyses. The mixed model (lme4 package in R) was used to compare the TN, TP, AN, and AP in rhizosphere and bulk soils (Table 1) and the SRL, RT/RB, and Nroot in fine roots among four study plant species (Table 2 and Table 3) in both seasons [47]. The probability of significance was defined as p < 0.05. We then built linear regression models to quantify the relationships among SRL, RT/RB, and Nroot in fine roots of the four study plant species and presented them as scatter plots (Figure 3). Linear regression models were then used to evaluate the relationships of three fine root traits to rhizosphere soil parameters (TN, TP, AN, and AP), oxalic acid, MBC, and NAG (Figure 4 and Figure 5).
3. Results
3.1. Soil Nutrient, Temperature, and Water Characteristics
The N and P contents of the rhizosphere and bulk soils were different among the four study species and between the dry and the wet season (Table 1). The four study species had higher TN, AN, and AP in rhizosphere and bulk soils in the wet season than in the dry season. The two tree species had higher TP in rhizosphere and bulk soils in the wet season than in the dry season, but the two shrub species had similar values of these parameters in both seasons. Additionally, the two tree species had higher values of the four measured parameters than the two shrub species in both seasons, except for no differences in TP between the tree and shrub species in the dry season. Soil temperature and water content in the wet season were higher than those in the dry season (Figure 2).
3.2. Patterns of Fine Root Functional Traits
The functional traits of fine roots were species-specific in the dry and the wet season (Table 2). Three of the species (A. trewioides, C. biondii, and P. tatarinowii), had higher values of SRL, RT/RB, and Nroot of fine roots in the wet season than in the dry season. L. sinense had similar values of the three measured parameters in both seasons. The two tree species had higher values of SRL, RT/RB, and Nroot of the fine roots than did the two shrub species in both seasons. Additionally, the specific variation of these fine root traits was affected by the seasons (Table 3).
3.3. Relationships between Fine Foot Functional Fraits and Soil Biochemistry
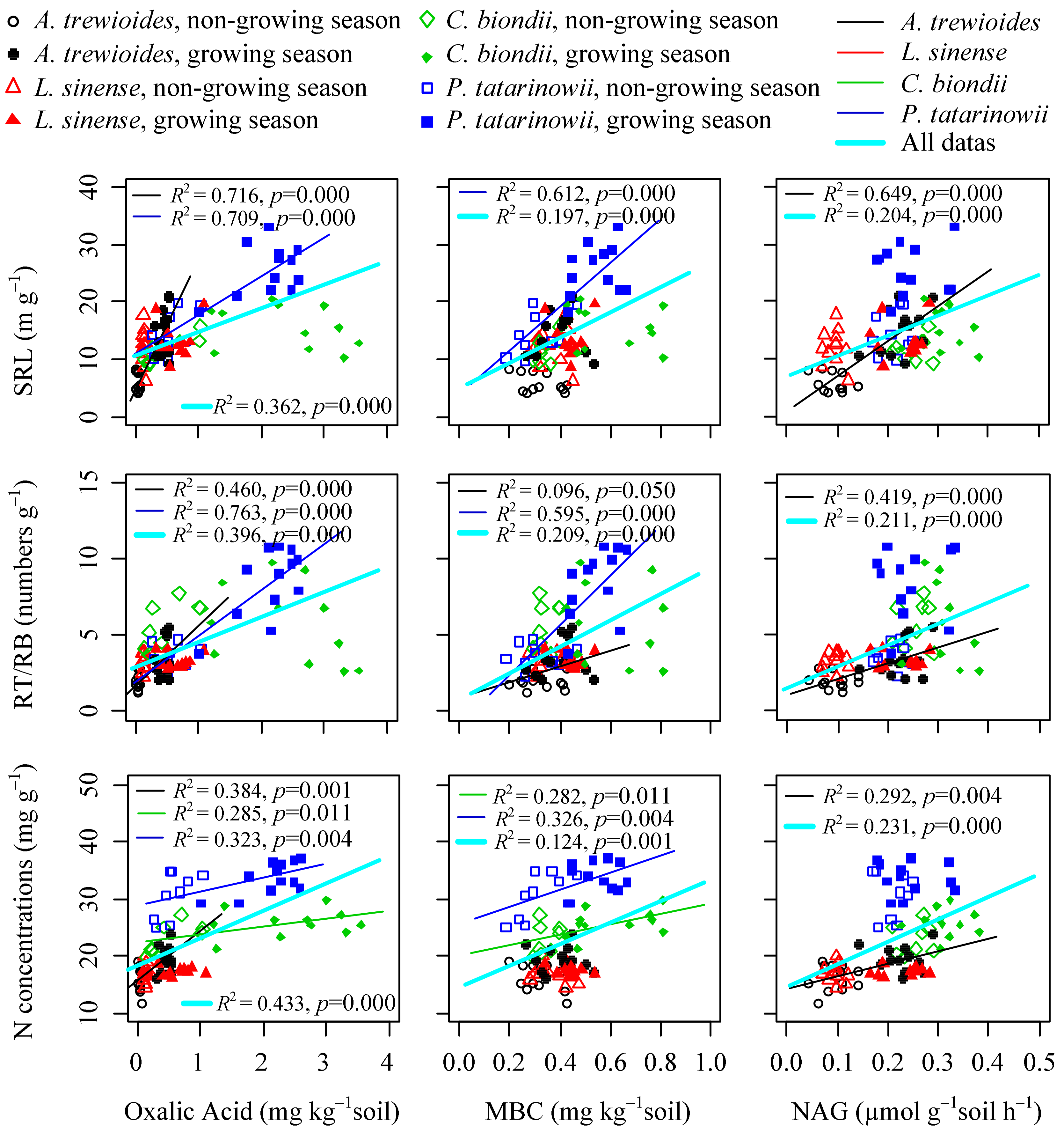
Significantly positive relationships were observed among the SRL, RT/RB, and Nroot in fine roots of the four species or overall species (Figure 3). The SRL of the overall species was positively correlated with soil TN, TP, and AN; the RT/RB and Nroot of the overall species were positively correlated with soil TN, TP, AN, and AP, as well as the Nroot of P. tatarinowii (Figure 4). Additionally, the SRL, RT/RB, and Nroot of overall species were positively correlated with rhizosphere soil oxalic acid, MBC, and NAG; at the species level, different plants had different patterns of three traits related to soil nutrients and biochemistry (Figure 5).
4. Discussion
4.1. Factors Influencing Fine Root Functional Traits
Different plant species can drive specific variations in the functional traits of fine roots [25]. For example, legumes and forbs have higher N concentrations in their fine roots than C3 and C4 plants [57,58]. These studies have shown that physiological activities are associated with nutrient uptake and assimilation, and that the responses are specific to different plant species. Some researchers found that the plant species that have high SRL or nutrient concentrations are associated with faster relative plant growth rates [59]. The trend shows that variations in fine root functional traits occur among plant species based on their taxonomy, which is phylogenetically structured [20]. In the present study, significant differences in fine root functional traits were observed among the four study species, and values of the fine root functional traits were higher in the two tree species than that in the two shrub species (Table 2). Our results were consistent with the findings of Freschet, which is that plant functional types can drive the variations of fine root traits [60]. One potential explanation for our results is that the lower values of fine root functional traits of two shrub species in the early stages are relative to the lower nutrient requirements of the plant species, which have slower growth rates, lower photosynthetic potential, and lower metabolic demand [47]. In our previous study, the plant diversity and complexity of community structure and species composition increased with the progressing vegetation successions [47], as well as the nutrient requirements of the plant species. To enhance nutrient uptake efficiency, plants, especially those with high metabolic demands, may increase their number of root tips for the improved absorption of soil nutrients [61,62]. Thus, tree species in the later successional stage had higher SRL, RT/RB, and Nroot in their fine roots than those in the earlier successional stage, which completely confirmed our first hypothesis.
Season is another important factor that influences fine root functional traits as a result of differing light intensities, soil temperature, water, microbial abundances, and enzymatic activities [36,37,63,64]. Firstly, higher light and temperature conditions can improve the photosynthetic capacity and further increase plant growth in the summer season compared to the winter season [65]. Although strong light or air temperature does not directly increase root length or root tip numbers, it can enhance the photosynthetic efficiency, and therefore increase the nutrient acquisition and uptake efficiency [66]. Thus, plants in high-light environments allocate proportionally more biomass to roots, and therefore have higher SRL compared with those in low-light environments [67]. The length of fine roots increase under air warming [68]. Besides the air temperature, soil temperature can have direct effects on fine root functional traits [36]. One recent article showed that the mean root elongation rates and total root lengths were 47% lower and 83% shorter in cold versus warm soils, respectively [69]. Secondly, the suitable wet conditions in the summer season also promote microorganism activity to further decompose soil nutrients from soil organic matter [18]. Under natural conditions, soil drought limits plant root growth, reduces soil nutrient availabilities, and forces plant roots to forage deeper for nutrients and water [70]. The causes of soil drought include: declining precipitation, large canopy gaps, low plant diversity, strong exposure to sunlight, etc. In general, soil water-holding ability can be improved by high plant diversity and vegetation coverage, especially in the summer [71]. High soil water content relieves the stress of the drought-limited expanding of soil microbial populations, enzymatic activities, and plant root growth. In the studied karst area, soil temperature and water contents were higher in July than in January (Figure 2). Our present study expected that the SRL, RT/RB, and Nroot of fine roots in studied plants (except evergreen shrub L. sinense) in summer (wet reason) were higher than those in winter (dry reason) (Table 2), which completely confirmed our second hypothesis. It might be that high soil water content and temperature could increase the length and root tips of fine roots in a karst ecosystem.
Changes in soil nutrient concentrations and availabilities dramatically shape the varied patterns of fine root functional traits [19,24,25,72]. Generally, root elongation from the higher SRL of fine roots enhances nutrient uptake from soils, resulting in higher nutrient concentrations in plants [73]. One study showed that the SRL of fine roots declined with increasing N addition, i.e., the first and second-order fine roots [33]. Another study demonstrated that the root length and SRL of plants decreases with advancing successional stages [74]. However, Noguchi et al. [72] found that the fine root SRL and Nroot of Sugi are positively related to increasing N input. Our results showed that the SRL, RT/RB, and Nroot in fine roots of overall species were positively correlated with the increasing soil nutrients, oxalic acid, MBC, and NAG (Figure 4 and Figure 5), which completely confirmed our third hypothesis. There are two potential explanations can be proposed for these results. Firstly, plants generally increase their SRL and root tips for enhanced nutrient acquisition in nutrient-limited soils [39,60,75]. In the present study, the patterns of fine SRL and root tips of karst plants may be related to the unique topogrophical characters of karst ecosystems. Plant growth may be limited by P due to high growth rates and N accumulations. In our previous study, leaf N:P ratios increased with the advancing successions, which resulted in P limitation for trees [47]. Soil N, which accumulates from atmospheric N deposition, biological N fixation, and soil organic matter, is rapidly recuperated from grasslands and primary forests [76]. However, the P supply level for plants is lower than soil N, and calcium-rich soils may compete with plants for AP in karst ecosystems [77]. Plants may absorb P (for growth) far away from the plant by elongating their roots. The large root system may exploit many more soil volumes and tolerate the stress of nutrient limitations. Additionally, high soil temperature, water, microbes, and enzymes activities can enhance nutrient availabilities [10]. In nutrient-limited ecosystems, in order to uptake nutrients, these functional traits may increase with the increasing soil nutrients in the more fertile soils [35,43]. Secondly, the exudation of oxalic acid from roots and hyphae, amounts of microbes, and activity of extracellular enzymes have been suggested to play an important role in enhancing nutrient acquisition by plants [17,78]. In the karst ecosystem, P becomes a key limiting factor that constrains plant growth in later vegetation successions [12]. Our results indicated that the changes in fine root functional traits of the tree species were to relieve P stress in karst regions, and were associated with soil biochemistry [18]. Thus, the SRL, RT/RB, and Nroot of fine roots in four study species were significantly related to soil nutrient changes and biochemistry.
4.2. Relationship between Fine Root Functional Traits and Plant Nutrient Acquisition Strategies
Changes in the fine root functional traits present important nutrient acquisition strategies to cope with the fundamental trade-offs between resource exploitation and conservation [24]. Plants with lower SRL, RT/RB, and Nroot can be suggested as the nutrients conservative plants [19]. In the present study, the fine roots of two shrub species had lower SRL, RT/RB, and Nroot than the two tree species (Table 2). In addition, soil N concentrations and availabilities were also lower in the shrubland than in the primary forest (Table 1). According to the suggestions by Fort and Montagnoli [19,36], the shrubs in our study could be considered as the species with N conservative strategies. In contrast, trees could be considered as the species with P exploitation strategies in the karst region. The findings of the present study were consistent with those of the previous study in karst regions, i.e., shrubs experienced N limitation and trees were affected by P limitation [12].
5. Conclusions
In this study, we provide new insights into the responses of the SRL, RT/RB, and Nroot in fine roots of shrub and tree species to the related soil biochemical status and seasons in karst regions. Variations in SRL, RT/RB, and Nroot were observed when plant species and seasons differed. We found that the three observed parameters of fine roots increased in the wet season, and were correlated with soil nutrients and biochemical parameters. Our study highlighted that plants enhanced the elongation and root tips of their fine roots to absorb nutrients in nutrient-limited soils. Moreover, the SRL, RT/RB, and Nroot were positively correlated to rhizosphere oxalic acid, microbial biomass C, and NAG activity. This indicated that the fine roots became associated with organic acid exudation and presumably the symbiosis of microbes (especially arbuscular mycorrhizal fungi) to relieve N and P stress in karst regions. However, literature is scarce on the collaborative response of fine root functional traits and arbuscular mycorrhizal fungi to changes in soil nutrient limitation. Further investigations are needed to reveal the mechanisms and potential applications of these relationships in karst regions.
Author Contributions
P.F., Z.W. and W.K. designed the experiments; P.F. and L.Y. performed the experiments; P.F. and L.Y. conducted the data analyses and prepared the manuscript; P.F. wrote the manuscript; Z.W. and W.K. helped perform the analyses and offered constructive guidance.
Funding
This study was financially supported by the National Key Research and Development Program, grant number 2016YFC0502406, the National Natural Science Foundation of China, grant numbers 31800441, 31741021, and 31670529, and the Natural Science Foundation of Guangxi, grant numbers 2018GXNSFBA138012, 2017GXNSFAA198241, and 2016GXNSFBA380179.
Acknowledgments
We appreciate M.S. Liu, F. Tian, T. Wang, and Q.T. Chen for their assistance in field sampling; the two reviewers and the academic editor for their time and constructive comments and suggestions.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Yuan, D.X. Karstology of China; Geological Publishing House: Beijing, Chinese, 1994. [Google Scholar]
- Qi, X.K.; Wang, K.L.; Zhang, C.H. Effectiveness of ecological restoration projects in a karst region of southwest China assessed using vegetation succession mapping. Ecol. Eng. 2013, 54, 245–253. [Google Scholar] [CrossRef]
- Su, Z.M.; Li, X.K. The types of natural vegetation in karst region of Guangxi and its classified system. Guihaia 2003, 23, 289–293. [Google Scholar]
- Chen, H.S.; Zhang, W.; Wang, K.L.; Fu, W. Soil moisture dynamics under different land uses on karst hillslope in northwest Guangxi, China. Environ. Earth Sci. 2010, 61, 1105–1111. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, K.L.; Chen, H.S.; He, X.Y.; Zhang, J.G. Ancillary information improves kriging on soil organic carbon data for a typical karst peak cluster depression landscape. J. Sci. Food Agr. 2012, 92, 1094–1102. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.J.; Liu, Q.M.; Zhang, D.F. Karst rocky desertification in southwestern China: Geomorphology, landuse, impact and rehabilitation. Land Degrad. Dev. 2004, 15, 115–121. [Google Scholar] [CrossRef]
- Du, Y.X.; Pan, G.X.; Li, L.Q.; Hu, Z.L.; Wang, X.Z. Leaf N/P ratio and nutrient reuse between dominant species and stands: Predicting phosphorus deficiencies in karst ecosystems, southwestern China. Environ. Earth Sci. 2011, 64, 299–309. [Google Scholar] [CrossRef]
- Yang, H.; Cao, J.H.; Sun, L.; Luan, H.N.; Hou, Y.L. Fractions and Distribution of Inorganic Phosphorus in Different Land Use Types of Karst Area. J. Soil Water Conserv. 2010, 2, 135–140. [Google Scholar]
- Zhang, X.B.; Bai, X.Y.; He, X.B. Soil creeping in the weathering crust of carbonate rocks and underground soil losses in the karst mountain areas of southwest China. Carbonates Evaporites 2011, 26, 149–153. [Google Scholar] [CrossRef]
- Pan, F.J.; Zhang, W.; Liang, Y.M.; Liu, S.J.; Wang, K.L. Increased associated effects of topography and litter and soil nutrients on soil enzyme activities and microbial biomass along vegetation successions in karst ecosystem, southwestern China. Environ. Sci. Pollut. R. 2018, 25, 16979–16990. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.P.; Xiang, W.S.; Yao, Y.F.; Huang, F.Z.; Li, X.K. Analyzing variation characteristics of vegetation net primary productivity in Guangxi. Guihaia 2014, 34, 622–628. [Google Scholar]
- Zhang, W.; Zhao, J.; Pan, F.J.; Li, D.J.; Chen, H.S.; Wang, K.L. Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in southwest China. Plant Soil 2015, 391, 77–91. [Google Scholar] [CrossRef]
- Kang, M.; Wang, J.; Huang, H.W. Nitrogen limitation as a driver of genome size evolution in a group of karst plants. Sci. Rep. 2015, 5, 11636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorley, R.M.; Taylor, L.L.; Banwart, S.A.; Leake, J.R.; Beerling, D.J. The role of forest trees and their mycorrhizal fungi in carbonate rock weathering and its significance for global carbon cycling. Plant Cell Environ. 2014, 38, 1947–1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.M.; He, X.Y.; Chen, C.Y.; Feng, S.Z.; Liu, L.; Chen, X.B.; Zhao, Z.W.; Su, Y.R. Influence of plant communities and soil properties during natural vegetation restoration on arbuscular mycorrhizal fungal communities in a karst region. Ecol. Eng. 2015, 82, 57–65. [Google Scholar] [CrossRef]
- Liu, L.; He, X.Y.; Wang, K.L.; Xie, Y.J.; Xie, Q.; O’Donnell, A.G.; Chen, C.Y. The Bradyrhizobium-legume symbiosis is dominant in the shrubby ecosystem of the Karst region, Southwest China. Eur. J. Soil Biol. 2015, 68, 1–8. [Google Scholar] [CrossRef]
- Clarholm, M.; Skyllberg, U.; Rosling, A. Organic acid induced release of nutrients from metal-stabilized soil organic matter—The unbutton model. Soil Biol. Biochem. 2015, 84, 168–176. [Google Scholar] [CrossRef]
- Pan, F.J.; Liang, Y.M.; Zhang, W.; Zhao, J.; Wang, K.L. Enhanced Nitrogen Availability in Karst Ecosystems by Oxalic Acid Release in the Rhizosphere. Front Plant Sci. 2016, 7, 687. [Google Scholar] [CrossRef] [PubMed]
- Fort, F.; Cruz, P.; Catrice, O.; Delbrut, A.; Luzarreta, M.; Stroia, C.; Jouany, C. Root functional trait syndromes and plasticity drive the ability of grassland Fabaceae to tolerate water and phosphorus shortage. Environ. Exp. Bot. 2015, 110, 62–72. [Google Scholar] [CrossRef]
- Valverde-Barrantes, O.J.; Smemo, K.A.; Blackwood, C.B. Fine root morphology is phylogenetically structured, but nitrogen is related to the plant economics spectrum in temperate trees. Funct. Ecol. 2015, 29, 796–807. [Google Scholar] [CrossRef]
- Helmisaari, H.S.; Makkonen, K.; Kellomaki, S.; Valtonen, E.; Malkonen, E. Below- and above-ground biomass, production and nitrogen use in Scots pine stands in eastern Finland. For. Ecol. Manag. 2002, 165, 317–326. [Google Scholar] [CrossRef]
- Montagnoli, A.; Di Iorio, A.; Terzaghi, M.; Trupiano, D.; Scippa, G.S.; Chiatante, D. Influence of soil temperature and water content on fine-root seasonal growth of European beech natural forest in Southern Alps, Italy. Eur. J. For. Res. 2014, 133, 957–968. [Google Scholar] [CrossRef]
- Wurzburger, N.; Wright, S.J. Fine-root responses to fertilization reveal multiple nutrient limitation in a lowland tropical forest. Ecology 2015, 96, 2137–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grassein, F.; Lemauviel-Lavenant, S.; Lavorel, S.; Bahn, M.; Bardgett, R.D.; Desclos-Theveniau, M.; Laine, P. Relationships between functional traits and inorganic nitrogen acquisition among eight contrasting European grass species. Ann. Bot.-London 2015, 115, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Pinno, B.D.; Wilson, S.D. Fine root response to soil resource heterogeneity differs between grassland and forest. Plant Ecol. 2013, 214, 821–829. [Google Scholar] [CrossRef]
- Kalliokoski, T.; Pennanen, T.; Nygren, P.; Sievanen, R.; Helmisaari, H.S. Belowground interspecific competition in mixed boreal forests: Fine root and ectomycorrhiza characteristics along stand developmental stage and soil fertility gradients. Plant Soil 2010, 330, 73–89. [Google Scholar] [CrossRef]
- Beyer, F.; Hertel, D.; Jung, K.; Fender, A.C.; Leuschner, C. Competition effects on fine root survival of Fagus sylvatica and Fraxinus excelsior. For. Ecol. Manag. 2013, 302, 14–22. [Google Scholar] [CrossRef]
- Sun, Y.; Gu, J.C.; Zhuang, H.F.; Wang, Z.Q. Effects of ectomycorrhizal colonization and nitrogen fertilization on morphology of root tips in a Larix gmelinii plantation in northeastern China. Ecol. Res. 2010, 25, 295–302. [Google Scholar] [CrossRef]
- Comas, L.H.; Eissenstat, D.M. Linking fine root traits to maximum potential growth rate among 11 mature temperate tree species. Funct. Ecol. 2004, 18, 388–397. [Google Scholar] [CrossRef] [Green Version]
- McCormack, M.L.; Adams, T.S.; Smithwick, E.A.H.; Eissenstat, D.M. Predicting fine root lifespan from plant functional traits in temperate trees. New Phytol. 2012, 195, 823–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nan, H.; Liu, Q.; Chen, J.; Cheng, X.; Yin, H.; Yin, C.; Zhao, C. Effects of nutrient heterogeneity and competition on root architecture of spruce seedlings: Implications for an essential feature of root foraging. PLoS ONE 2013, 8, e65650. [Google Scholar] [CrossRef] [PubMed]
- Ostonen, I.; Püttsepp, Ü.; Biel, C.; Alberton, O.; Bakker, M.R.; Lõhmus, K.; Majdi, H.; Metcalfe, D.; Olsthoorn, A.F.M.; Pronk, A.; Vanguelova, E.; et al. Specific root length as an indicator of environmental change. Plant Biosyst. 2007, 141, 426–442. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.L.; Fahey, T.J.; Xue, S.; Liu, F. Root morphology and architecture respond to N addition in Pinus tabuliformis, west China. Oecologia 2013, 171, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Kochsiek, A.; Tan, S.; Russo, S.E. Fine root dynamics in relation to nutrients in oligotrophic Bornean rain forest soils. Plant Ecol. 2013, 214, 869–882. [Google Scholar] [CrossRef]
- Espeleta, J.F.; Donovan, L.A. Fine root demography and morphology in response to soil resources availability among xeric and mesic sandhill tree species. Funct. Ecol. 2002, 16, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Montagnoli, A.; Terzaghi, M.; Di Iorio, A.; Scippa, G.S.; Chiatante, D. Fine-root morphological and growth traits in a Turkey-oak stand in relation to seasonal changes in soil moisture in the Southern Apennines, Italy. Ecol. Res. 2012, 27, 1015–1025. [Google Scholar] [CrossRef] [Green Version]
- Montagnoli, A.; Di Iorio, A.; Terzaghi, M.; Trupiano, D.; Scippa, G.S.; Chiatante, D. Influence of soil temperature and water content on fine-root seasonal growth of European beech natural forest in Southern Alps, Italy. Eur. J. For. Res. 2014, 133, 957–968. [Google Scholar] [CrossRef] [Green Version]
- Montagnolia, A.; Dumroeseb, R.K.; Terzaghia, M.; Onellic, E.; Scippad, G.S.; Chiatantea, D. Seasonality of fine root dynamics and activity of root and shoot vascular cambium in a Quercus ilex L. forest (Italy). For. Ecol. Manag. 2018. [Google Scholar] [CrossRef]
- Holdaway, R.J.; Richardson, S.J.; Dickie, I.A.; Peltzer, D.A.; Coomes, D.A. Species- and community-level patterns in fine root traits along a 120,000-year soil chronosequence in temperate rain forest. J. Ecol. 2011, 99, 954–963. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shipley, B.; Lechowicz, M.J.; Wright, I.; Reich, P.B. Fundamental trade-offs generating the worldwide leaf economics spectrum. Ecology 2006, 87, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Ostonen, I.; Helmisaari, H.S.; Borken, W.; Tedersoo, L.; Kukumagi, M.; Bahram, M.; Lindroos, A.J.; Nojd, P.; Uri, V.; Merila, P.; et al. Fine root foraging strategies in Norway spruce forests across a European climate gradient. Glob. Change Biol. 2011, 17, 3620–3632. [Google Scholar] [CrossRef]
- Kou, L.; Guo, D.L.; Yang, H.; Gao, W.L.; Li, S.G. Growth, morphological traits and mycorrhizal colonization of fine roots respond differently to nitrogen addition in a slash pine plantation in subtropical China. Plant Soil. 2015, 391, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.H.; Hu, G.; Zhu, J.D.; Luo, D.H.; Ni, J. Spatial patterns and interspecific associations of dominant tree species in two old-growth karst forests, SW China. Ecol. Res. 2010, 25, 1151–1160. [Google Scholar] [CrossRef]
- Liu, C.C.; Liu, Y.G.; Guo, K.; Li, G.Q.; Zheng, Y.R.; Yu, L.F.; Yang, R. Comparative ecophysiological responses to drought of two shrub and four tree species from karst habitats of southwestern China. Trees-Struct. Funct. 2011, 25, 537–549. [Google Scholar] [CrossRef]
- Wang, Z.H.; Xu, W.Y. Decomposition-rate estimation of leaf litter in karst forests in China based on a mathematical model. Plant Soil 2013, 367, 563–577. [Google Scholar] [CrossRef]
- Pan, F.J.; Zhang, W.; Liu, S.J.; Li, D.J.; Wang, K.L. Leaf N:P stoichiometry across plant functional groups in the karst region of southwestern China. Trees-Struct. Funct. 2015, 29, 883–892. [Google Scholar] [CrossRef]
- Nie, Y.P.; Chen, H.S.; Wang, K.L.; Yang, J. Water source utilization by woody plants growing on dolomite outcrops and nearby soils during dry seasons in karst region of Southwest China. J. Hydrol. 2012, 420, 264–274. [Google Scholar] [CrossRef]
- Macrae, A.; Lucon, C.M.M.; Rimmer, D.L.; O’Donnell, A.G. Sampling DNA from the rhizosphere of Brassica napus to investigate rhizobacterial community structure. Plant Soil 2001, 233, 223–230. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; DeForest, J.L.; Burton, A.J.; Allen, M.F.; Ruess, R.W.; Hendrick, R.L. Fine root architecture of nine North American trees. Ecol. Monogr. 2002, 72, 293–309. [Google Scholar] [CrossRef]
- Guo, D.L.; Mitchell, R.J.; Hendricks, J.J. Fine root branch orders respond differentially to carbon source-sink manipulations in a longleaf pine forest. Oecologia 2004, 140, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Kurosumi, A.; Kaneko, E.; Nakamura, Y. Degradation of reactive dyes by ozonation and oxalic acid-assimilating bacteria isolated from soil. Biodegradation 2008, 19, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Finzi, A.C.; Sinsabaugh, R.L.; Long, T.M.; Osgood, M.P. Microbial community responses to atmospheric carbon dioxide enrichment in a warm-temperate forest. Ecosystems 2006, 9, 215–226. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Wu, J.S.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of Soil Microbial Biomass C by Fumigation Extraction—An Automated Procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: http://www.R-project.org/.
- Tjoelker, M.G.; Craine, J.M.; Wedin, D.; Reich, P.B.; Tilman, D. Linking leaf and root trait syndromes among 39 grassland and savannah species. New Phytol. 2005, 167, 493–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fornara, D.A.; Tilman, D.; Hobbie, S.E. Linkages between plant functional composition, fine root processes and potential soil N mineralization rates. J. Ecol. 2009, 97, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Wahl, S.; Ryser, P. Root tissue structure is linked to ecological strategies of grasses. New Phytol. 2000, 148, 459–471. [Google Scholar] [CrossRef] [Green Version]
- Freschet, G.T.; Valverde-Barrantes, O.J.; Tucker, C.M.; Craine, J.M.; McCormack, M.L.; Violle, C.; Fort, F.; Blackwood, C.B.; Urban-Mead, K.R.; Iversen, C.M.; et al. Climate, soil and plant functional types as drivers of global fine-root trait variation. J. Ecol. 2017, 105, 1182–1196. [Google Scholar] [CrossRef]
- Helmisaari, H.S.; Ostonen, I.; Lohmus, K.; Derome, J.; Lindroos, A.J.; Merila, P.; Nojd, P. Ectomycorrhizal root tips in relation to site and stand characteristics in Norway spruce and Scots pine stands in boreal forests. Tree Physiol. 2009, 29, 445–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, W.H.; Wu, W.; Tong, J.; Deng, X.W.; Tian, D.L.; Zhang, L.; Liu, C.; Peng, C.H. Differences in fine root traits between early and late-successional tree species in a Chinese subtropical forest. Forestry 2013, 86, 343–351. [Google Scholar] [CrossRef]
- Loiola, P.P.; Scherer-Lorenzen, M.; Batalha, M.A. The role of environmental filters and functional traits in predicting the root biomass and productivity in savannas and tropical seasonal forests. For. Ecol. Manag. 2015, 342, 49–55. [Google Scholar] [CrossRef]
- Chen, S.M.; Lin, S.; Loges, R.; Reinsch, T.; Hasler, M.; Taube, F. Independence of seasonal patterns of root functional traits and rooting strategy of a grass-clover sward from sward age and slurry application. Grass Forage Sci. 2016, 71, 607–621. [Google Scholar] [CrossRef]
- Jagodzinski, A.M.; Dyderski, M.K.; Rawlik, K.; Katna, B. Seasonal variability of biomass, total leaf area and specific leaf area of forest understory herbs reflects their life strategies. For. Ecol. Manag. 2016, 374, 71–81. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Tjoelker, M.G.; Vanderklein, D.; Buschena, C. Photosynthesis and respiration rates depend on leaf and root morphology and nitrogen concentration in nine boreal tree species differing in relative growth rate. Funct. Ecol. 1998, 12, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B.; Tjoelker, M.G.; Walters, M.B.; Vanderklein, D.W.; Bushena, C. Close association of RGR, leaf and root morphology, seed mass and shade tolerance in seedlings of nine boreal tree species grown in high and low light. Funct. Ecol. 1998, 12, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Pilon, R.; Picon-Cochard, C.; Bloor, J.M.G.; Revaillot, S.; Kuhn, E.; Falcimagne, R.; Balandier, P.; Soussana, J.F. Grassland root demography responses to multiple climate change drivers depend on root morphology. Plant Soil 2013, 364, 395–408. [Google Scholar] [CrossRef]
- Sebastian, N.; Erika, H.; Christian, K. Critically low soil temperatures for root growth and root morphology in three alpine plant species. Alp. Bot. 2016, 126, 11–21. [Google Scholar] [CrossRef]
- Paz, H.; Pineda-Garcia, F.; Pinzon-Perez, L.F. Root depth and morphology in response to soil drought: Comparing ecological groups along the secondary succession in a tropical dry forest. Oecologia 2015, 179, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.; Tischer, J.; Roscher, C.; Eisenhauer, N.; Ravenek, J.; Gleixner, G.; Attinger, S.; Jensen, B.; de Kroon, H.; Mommer, L.; et al. Plant species diversity affects infiltration capacity in an experimental grassland through changes in soil properties. Plant Soil 2015, 397, 1–16. [Google Scholar] [CrossRef]
- Noguchi, K.; Nagakura, J.; Kaneko, S. Biomass and morphology of fine roots of sugi (Cryptomeria japonica) after 3 years of nitrogen fertilization. Front Plant Sci. 2013, 4, 347. [Google Scholar] [CrossRef] [PubMed]
- Eissenstat, D.M. On the relationship between specific root length and the rate of root proliferation—A field–study using citrus rootstocks. New Phytol. 1991, 118, 63–68. [Google Scholar] [CrossRef]
- Zangaro, W.; Alves, R.A.; Lescano, L.E.; Ansanelo, A.P.; Nogueira, M.A. Investment in fine roots and arbuscular mycorrhizal fungi decrease during succession in three brazilian ecosystems. Biotropica 2012, 44, 141–150. [Google Scholar] [CrossRef]
- Svistoonoff, S.; Creff, A.; Reymond, M.; Sigoillot-Claude, C.; Ricaud, L.; Blanchet, A.; Nussaume, L.; Desnos, T. Root tip contact with low-phosphate media reprograms plant root architecture. Nat. Genet. 2007, 39, 792–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, L.; Li, D.J.; Yang, L.Q.; Luo, P.; Chen, H.; Xiao, K.C.; Song, T.Q.; Zhang, W.; He, X.Y.; Chen, H.S.; et al. Rapid recuperation of soil nitrogen following agricultural abandonment in a karst area, southwest China. Biogeochemistry 2016, 129, 341–354. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Strom, L.; Owen, A.G.; Godbold, D.L.; Jones, D.L. Organic acid behaviour in a calcareous soil implications for rhizosphere nutrient cycling. Soil Biol. Biochem. 2005, 37, 2046–2054. [Google Scholar] [CrossRef]
Figure 1.
Sketch map showing the Huanjiang Observation and Research Station (●) and Mulun National Natural Reserve (▲) and the typical landscape in a karst ecosystem of southwest China.
Figure 1.
Sketch map showing the Huanjiang Observation and Research Station (●) and Mulun National Natural Reserve (▲) and the typical landscape in a karst ecosystem of southwest China.

Figure 2.
Soil temperature (T) and volumetric moisture contents (VMC) at 10 cm depth in Huanjiang Observation and Research Station (T1 and VMC1) and Mulun National Natural Reserve (T2 and VMC2) from 2008 to 2015.
Figure 2.
Soil temperature (T) and volumetric moisture contents (VMC) at 10 cm depth in Huanjiang Observation and Research Station (T1 and VMC1) and Mulun National Natural Reserve (T2 and VMC2) from 2008 to 2015.

Figure 3.
Relationships between specific root length (SRL), root tips over the root biomass (RT/RB), and N concentrations (Nroot) in fine roots of the four study plant species in the dry and the wet season in a karst ecosystem of southwest China.
Figure 3.
Relationships between specific root length (SRL), root tips over the root biomass (RT/RB), and N concentrations (Nroot) in fine roots of the four study plant species in the dry and the wet season in a karst ecosystem of southwest China.

Figure 4.
Relationships between fine root traits (SRL, RT/RB, and N concentration) and soil nutrients concentrations and availabilities in a karst ecosystem of southwest China.
Figure 4.
Relationships between fine root traits (SRL, RT/RB, and N concentration) and soil nutrients concentrations and availabilities in a karst ecosystem of southwest China.

Figure 5.
Relationships between fine root traits (SRL, RT/RB, and N concentration) and rhizosphere soil oxalic acid, microbial biomass carbon (MBC), and activity of a hydrolytic enzyme (NAG) in a karst ecosystem of southwest China.
Figure 5.
Relationships between fine root traits (SRL, RT/RB, and N concentration) and rhizosphere soil oxalic acid, microbial biomass carbon (MBC), and activity of a hydrolytic enzyme (NAG) in a karst ecosystem of southwest China.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Nitrogen and phosphorus concentrations and availabilities in rhizosphere and bulk soils of the four study plant species in a karst ecosystem of southwest China (mean ± SE).
Table 1.
Nitrogen and phosphorus concentrations and availabilities in rhizosphere and bulk soils of the four study plant species in a karst ecosystem of southwest China (mean ± SE).
| Source | Total Nitrogen (g kg−1) | Total Phosphorus (g kg−1) | Available Nitrogen (mg kg−1) | Available Phosphorus (mg kg−1) | |
|---|---|---|---|---|---|
| Dry season | |||||
| Alchornea trewioides | RS | 5.86 ± 0.35 A | 1.45 ± 0.08 A | 346.07 ± 23.59 A | 8.83 ± 0.66 A |
| BS | 4.59 ± 0.33 a | 1.38 ± 0.07 a | 329.18 ± 22.66 a | 5.19 ± 0.67 a | |
| Ligustrum sinense | RS | 6.59 ± 0.39 A | 1.24 ± 0.09 A | 422.58 ± 14.19 B | 12.24 ± 0.92 AB |
| BS | 3.86 ± 0.38 a | 1.34 ± 0.10 a | 308.14 ± 24.41 a | 6.55 ± 0.91 a | |
| Celtis biondii | RS | 10.85 ± 1.23 B | 1.54 ± 0.17 A | 425.86 ± 60.70 B | 15.30 ± 4.11 B |
| BS | 7.58 ± 0.94 b | 1.35 ± 0.12 a | 482.65 ± 37.81 b | 7.54 ± 0.85 b | |
| Pteroceltis tatarinowii | RS | 10.30 ± 1.54 B | 1.52 ± 0.11 A | 425.87 ± 38.30 B | 10.64 ± 1.93 A |
| BS | 7.02 ± 1.06 b | 1.38 ± 0.47 a | 416.99 ± 32.33 ab | 6.49 ± 1.30 a | |
| Wet season | |||||
| Alchornea trewioides | RS | 6.53 ± 0.41 A | 1.48 ± 0.03 A | 440.20 ± 28.59 A | 9.00 ± 0.83 A |
| BS | 4.96 ± 0.26 a | 1.39 ± 0.04 a | 347.25 ± 15.92 a | 5.63 ± 0.32 a | |
| Ligustrum sinense | RS | 8.61 ± 0.72 A | 1.52 ± 0.10 A | 501.32 ± 28.87 A | 12.37 ± 0.70 A |
| BS | 5.66 ± 0.57 a | 1.42 ± 0.11 a | 334.47 ± 21.55 a | 7.44 ± 0.67 a | |
| Celtis biondii | RS | 17.61 ± 1.33 C | 1.86 ± 0.15 B | 847.34 ± 50.52 C | 25.52 ± 4.10 B |
| BS | 11.73 ± 0.62 b | 1.68 ± 0.12 b | 759.88 ± 48.81 c | 26.42 ± 4.19 c | |
| Pteroceltis tatarinowii | RS | 12.21 ± 1.11 B | 1.84 ± 0.15 B | 586.53 ± 36.96 B | 13.31 ± 1.72 A |
| BS | 10.10 ± 1.11 b | 1.68 ± 0.14 b | 547.42 ± 44.14 b | 16.12 ± 2.77 b |
RS, Rhizosphere soil; BS, Bulk soil. Lowercase letters in each column indicate significant differences of bulk soils (p < 0.05); uppercase letters indicate significant differences of rhizosphere soils (p < 0.05) among four species; the bold data indicate significant difference between rhizosphere soil and bulk soil.
Table 2.
Specific root length (SRL), root tips over the root biomass (RT/RB), and N concentrations (Nroot) in fine roots of the four study plant species in the dry (from October to March) and the wet (from April to September) season in a karst ecosystem of southwest China (mean ± SE).
Table 2.
Specific root length (SRL), root tips over the root biomass (RT/RB), and N concentrations (Nroot) in fine roots of the four study plant species in the dry (from October to March) and the wet (from April to September) season in a karst ecosystem of southwest China (mean ± SE).
| SRL (m g−1) | RT/RB (Number mg−1) | Nroot (mg g−1) | |
|---|---|---|---|
| Dry season | |||
| Alchornea trewioides (shrub) | 5.78 ± 0.48 a | 1.95 ± 0.14 a | 16.12 ± 0.67 a |
| Ligustrum sinense (shrub) | 12.08 ± 0.88 b | 3.30 ± 0.17 b | 17.01 ± 0.43 a |
| Celtis biondii (tree) | 11.70 ± 0.79 b | 5.72 ± 0.53 c | 22.97 ± 0.95 b |
| Pteroceltis tatarinowii (tree) | 14.40 ± 1.23 b | 3.60 ± 0.26 b | 30.68 ± 1.35 b |
| Wet season | |||
| Alchornea trewioides (shrub) | 15.19 ± 1.17 a | 3.47 ± 0.35 a | 19.70 ± 0.65 a |
| Ligustrum sinense (shrub) | 13.32 ± 0.98 a | 3.30 ± 0.15 a | 17.30 ± 0.23 a |
| Celtis biondii (tree) | 15.63 ± 1.14 a | 5.76 ± 0.80 b | 25.80 ± 0.72 b |
| Pteroceltis tatarinowii (tree) | 25.64 ± 1.27 b | 8.41 ± 0.66 c | 33.60 ± 0.80 c |
Lowercase letters in each column indicate significant differences (p < 0.05) among the plant species.
Table 3.
Effect of seasons on specific root length (SRL), root tips over the root biomass (RT/RB), and N concentrations (Nroot) in fine roots of the four study plant species in a karst ecosystem of southwest China.
Table 3.
Effect of seasons on specific root length (SRL), root tips over the root biomass (RT/RB), and N concentrations (Nroot) in fine roots of the four study plant species in a karst ecosystem of southwest China.
| Response Variable | Linear Mixed Models | AIC | BIC | T Value | Significant Difference |
|---|---|---|---|---|---|
| SRL | Null model | 558.81 | 566.21 | 4.84 | a |
| Species model | 538.97 | 553.77 | 3.56 | b | |
| Season model | 526.35 | 536.21 | 3.99 | b | |
| Species * Season model | 494.11 | 511.37 | 3.14 | c | |
| RT/RB | Null model | 373.02 | 380.42 | 2.96 | a |
| Species model | 368.20 | 382.99 | 2.09 | b | |
| Season model | 358.95 | 368.81 | 2.50 | b | |
| Species * Season model | 353.26 | 370.52 | 1.60 | c | |
| Nroot | Null model | 494.46 | 501.86 | 4.15 | a |
| Species model | 430.58 | 445.38 | 2.98 | b | |
| Season model | 487.63 | 497.50 | 3.99 | c | |
| Species * Season model | 413.68 | 430.94 | 3.25 | d |
(1) Null model: ƒi = α + bi × Zsite + εi; (2) Species or Season model: ƒi = α + β × X (species or season) + bi × Zsite + εi; (3) Species * Season model: ƒi = α + β1 ×Xspecies + β2 ×Xseason + bi × Zsite + εi. Here, ƒi is the response variable, X is the fixed effect, Z is the random effect, α is the intercept of model, β is the parameter of fixed factor, bi is the parameter of random factor, and εi the unexplained information. The optimal model was defined with minimum AIC (Akaike Information Criterion) and BIC (Bayesian Information Criterion). The random factor was merely captured by the null model. Different letters in the last column indicate instances when the mixed model was better than the null model when mixed factors (species or season) were captured by the model.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pan, F.; Liang, Y.; Wang, K.; Zhang, W. Responses of Fine Root Functional Traits to Soil Nutrient Limitations in a Karst Ecosystem of Southwest China. Forests 2018, 9, 743. https://doi.org/10.3390/f9120743
AMA Style
Pan F, Liang Y, Wang K, Zhang W. Responses of Fine Root Functional Traits to Soil Nutrient Limitations in a Karst Ecosystem of Southwest China. Forests. 2018; 9(12):743. https://doi.org/10.3390/f9120743
Chicago/Turabian StylePan, Fujing, Yueming Liang, Kelin Wang, and Wei Zhang. 2018. "Responses of Fine Root Functional Traits to Soil Nutrient Limitations in a Karst Ecosystem of Southwest China" Forests 9, no. 12: 743. https://doi.org/10.3390/f9120743
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.