A Subcontinental Analysis of Forest Fragmentation Effects on Insect and Disease Invasion


1
Southern Research Station, USDA Forest Service, Research Triangle Park, NC 27709, USA
2
Department of Forestry and Environmental Resources, North Carolina State University, Raleigh, NC 27695, USA
*
Author to whom correspondence should be addressed.
Forests 2018, 9(12), 744; https://doi.org/10.3390/f9120744
Submission received: 24 October 2018
/
Revised: 16 November 2018
/
Accepted: 27 November 2018
/
Published: 29 November 2018
(This article belongs to the Special Issue Impacts of Nonnative Species on the Health of Natural and Planted Forests)
Abstract
:The influences of human and physical factors on species invasions have been extensively examined by ecologists across many regions. However, how habitat fragmentation per se may affect forest insect and disease invasion has not been well studied, especially the related patterns over regional or subcontinental scales. Here, using national survey data on forest pest richness and fragmentation data across United States forest ecosystems, we examine how forest fragmentation and edge types (neighboring land cover) may affect pest richness at the county level. Our results show that habitat fragmentation and edge types both affected pest richness. In general, specialist insects and pathogens were more sensitive to fragmentation and edge types than generalists, while pathogens were much less sensitive to fragmentation and edge types than insect pests. Most importantly, the developed land edge type contributed the most to the richness of nonnative insects and diseases, whether measured by the combination of all pest species or by separate guilds or species groups (i.e., generalists vs. specialists, insects vs. pathogens). This observation may largely reflect anthropogenic effects, including propagule pressure associated with human activities. These results shed new insights into the patterns of forest pest invasions, and it may have significant implications for forest restoration and management.
1. Introduction
Invasion by nonnative pest species causes enormous harm to native ecosystems worldwide [1,2]. Not only does intensive land-use drive the regional-scale homogenization of plant communities, but also that of animal communities [2], especially when both native and nonnative species are considered. To date, the activities of humans have been found to be the major drivers in species invasion [3] but most related studies have been conducted at the same trophic level (i.e., effects of nonnative plants on native plants) and/or over small scales. The net contribution of habitat fragmentation (natural vs. anthropogenic) and cross-trophic effects (i.e., effects of plants on animals) is much less clear.
Previous cross-trophic-level studies on invasion patterns in the forest systems of the United States have examined the effects of propagule pressure, habitat invasibility, and climate conditions on pest richness [4,5,6,7,8]. These studies have investigated the effects of anthropogenic and environmental variables, including human population density, native host, and non-host tree diversity and biomass, forest area, mean annual temperature and precipitation, and elevation [3]. However, mounting evidence shows that landscape mosaics (e.g., those caused by habitat fragmentation) are expected to play a role in the spread of invasive species. Also, different from effects of invasions of same-trophic species, cross-trophic invasions such as insect/disease invasions that cause harm to native plants are much more complex and difficult to study [3].
Indeed, researchers are becoming increasingly more interested in the possible effects of landscape spatial structures. In addition, many natural landscape structures or mosaics have been altered due to human activities that form new edges and cause more fragmentation. Although habitat loss is still regarded as the major cause of species loss, most studies around the world have found positive responses of various related variables (e.g., biodiversity) to habitat fragmentation; such positive responses are not generally related to the habitat area [9].
In addition to the effects of natural fragmentation, conservation practitioners are especially concerned about fragmentation caused by humans. It is expected that anthropogenic fragmentation, which increases overall fragmentation, could have different effects on ecosystem properties than fragmentation caused by natural forces [10]. Furthermore, a recent study found synergism among anthropogenic causes such as roads and adjacent land uses affecting forest plant invasions, suggesting that different pools of invasive plant species originated from different nearby land uses, thereby increasing the potential exposure of forested areas within heterogeneous landscapes [11].
Studies on edge and fragmentation effects are numerous, but are mostly small-scale surveys or experiments [9,12,13]; no continental-scale study has specifically examined the effects of habitat fragmentation per se, especially on species invasions. Yet, human activities have no doubt increased overall habitat fragmentation [9,14], and more importantly, such habitat fragmentation creates more new neighboring land cover types and habitats, and thus increases the diversity of edges. The different types of edge/neighbors may play different roles by either facilitating or hampering pest invasions [12,13,15,16]. Here, we specifically examine the possible effects of habitat fragmentation on forest pest invasions across the United States. Habitat fragmentation considers that caused by both natural and anthropogenic forces [10]. We ask three main questions: (1) Does habitat fragmentation affect nonnative forest pest invasions across the United States? (2) What is the role of neighboring habitat types in forest pest invasions? (3) What are the implications of the fragmentation–invasion relationship in forest management?
2. Materials and Methods
2.1. Forest Pest Data
We used the county-level information for 91 species of non-native forest insect herbivores and forest pathogens known to cause damage across the United States (48 states and 2098 counties for which forest insects and diseases were present, and data were available) from the Alien Forest Pest Explorer database [17]. We then selected and analyzed county occurrence data on 66 pest species that primarily utilize native tree species (Table S1) [3]. These are insects and pathogens that feed on foliage, sap, phloem or wood, and exclude predators, pollinators, and detritivores.
To examine the possibly different responses of different pest groups, the 66 species were first classified into specialists (N = 41) and generalists (N = 25), based on feeding preferences. The latter also included oligophagous species, i.e., those feeding on two or more genera from the same family, or from closely related families (N = 10). For this analysis, the oligophagous agents were combined with the generalists. The 66 species were further regrouped as pathogens (N = 15) and insects (51), and the insects were classified into three feeding guilds (i.e., 26 defoliators, 14 sap feeders, and 11 wood borers) [3]. Pathogens accounted for 24% (6/25) and 21% (9/41) among the generalists and specialists, respectively.
2.2. Habitat Fragmentation Data
We compiled a county-level forest fragmentation dataset using 2011 national land cover data [18,19]. The land cover maps were derived primarily from Landsat satellite images [20] supplemented by ancillary geographic data. They were produced at a spatial resolution of 0.09 ha per pixel with a thematic resolution of 16 land cover classes. We aggregated the 16 land cover classes into five classes called forest, developed, agricultural, grass-shrub, and “other” (Table S2). The classification accuracy of the five aggregated classes [21] was deemed to be adequate for this study. The class “other” was ignored in subsequent analysis (Figure 1).
From the land cover map, we derived metrics that described the total forest area, forest spatial patterns, and forest adjacencies (types of forest edge, or “edge type”). Forest area was expressed as the total county area with forest land cover. Forest spatial patterns were forest area (number of pixels occupied) in a given county in each of three pattern classes (core, islet, and edge) determined by morphological spatial pattern analysis (MSPA; [22]) (Figure 1a). The MSPA class “core” refers to a forest pixel that is more than 30 m (one pixel) away from non-forest. Pixels in the class “islet” comprise contiguous clusters of forest pixels that are too small to contain a core. Pixels in the “edge” class are all of the remaining forest pixels, including interior and exterior edges, and narrow connecting structures. Note that an islet pixel is different from an edge pixel, only because the edge is connected in some way to a core forest cluster, whereas an islet is not connected to the core forest. From those measurements, we also calculated the core/edge area ratio at the county level.
Following procedures used in a previous global assessment [10], we used a moving window algorithm to evaluate the forest adjacencies in the unique 9.92 km2 neighborhood surrounding each forest pixel on the land cover map (Figure 1b). A forest adjacency was defined by the land cover type that was adjacent to a given forest pixel in one of the four cardinal directions. The five possible adjacencies were forest|forest, forest|developed (Fxde), forest|agriculture (Fxag), forest|grass-shrub (Fxgs), and forest|other. In a given window, each adjacency was counted once (i.e., the spatial order of the pixels was ignored) and the total number of forest adjacencies was the sum over all five types of adjacencies. The percentages of total forest adjacencies that involved developed land cover (Fxde), agricultural land cover (Fxag), and grass-shrub land cover (Fxgs) were then calculated in each window. These percentages are thus indicative of both total fragmentation (i.e., the magnitude of forest adjacencies that are not forest|forest adjacencies) and the relative importance of different fragmenting land covers. Since the percentages were calculated for the neighborhood surrounding each forest pixel, we aggregated the percentages to the county level by finding the mean percentage of each forest adjacency type for all forest pixels in a given county. Thus, at the county level, the adjacency metric indicates the type and importance of fragmenting agents for an “average forest edge” in the county.
2.3. Data Analysis
To examine the effects of habitat fragmentation on pest richness, we conducted ordinary-least-squares (OLS) estimation and spatial autoregression analyses (SAR) that took both independent variables and autocorrelation (space) into account. Autocorrelation was included because counties next to each other are likely to be more similar than should be expected at random, and they were measured based on the geodesic distances among the 2098 counties. We then compared the results and corrected Akaike Information Criterion (AICc) values in both analyses (lower AICC values indicate better model fits). For comparison, we also did the same analyses by using the proportional (%) data (e.g., core vs. edge). To meet the requirements (e.g., normality) of regression analysis, all data were log-transformed before analysis.
3. Results
3.1. Effects of Fragmentation
We detected a negative effect of forest edge on nonnative pest richness, but the areas of forest, core, and islet had no significant effects on pest richness (Table 1). Both fragmentation measures, i.e., “core/edge ratio” and “core/(islet + edge) ratio”, exhibited similar positive relationships with pest richness (r2 = 0.031, p < 0.0001 and r2 = 0.046, p < 0.0001, respectively; Figure 2). Using proportional (%) values of core, islet, and edge yielded similar results.
Specialists and generalists exhibited different responses to fragmentation and, in general, specialists were more sensitive to fragmentation than generalists, as indicated by significant responses to both core and edge areas, while generalists only responded significantly to islet areas (Table 2). While forest area had a positive effect on specialist richness, the core area’s effect was negative. For generalists, however, the only negative effect was from the islet area; forest area, core area, and edge area had no effects (Table 2).
We found major differences between nonnative pathogens and insects in terms of their responses to fragmentation. In general, pathogens were negatively affected by edge pixel area. For insect richness, while forest area had a positive effect, all others measures such as core, islet, and edge areas had negative effects (Table 3). The insect guilds also exhibited very different responses to fragmentation. Defoliators were positively influenced by forest area, but negatively by islet area. Sap feeders were positively affected by forest area, while wood borers were positively related to islet area but negatively to edge area (Table 4).
3.2. Edge Effects
There were major differences among forest edge types in the patterns of pest invasions (richness). Forest edges with neighboring developed land had a positive influence on pest richness, while forests with grass/shrub edges had significantly lower nonnative pest richness. Agricultural edge had no significant effect (Table 1).
Specialists and generalists also exhibited some different responses to edge types. Both agricultural and developed edges had positive relationships with generalist richness, but grass/shrub edge had a strong negative effect. For specialists, while the effects of developed and grass/shrub were the same as for generalists, agricultural edge had no significant effects (Table 2).
Both nonnative pathogen richness and insect richness were similarly affected by developed edge (positive) and grass/shrub edge (negative), and there was no significant effect from agricultural edges (Table 3). All three insect guilds also showed similar and significant responses to edge types. Specifically, the three guilds were all positively affected by developed edges, and negatively by grass/shrub edges. However, agricultural edges positively affected defoliators and negatively affected sap feeders, and had no effect on wood borers (Table 4).
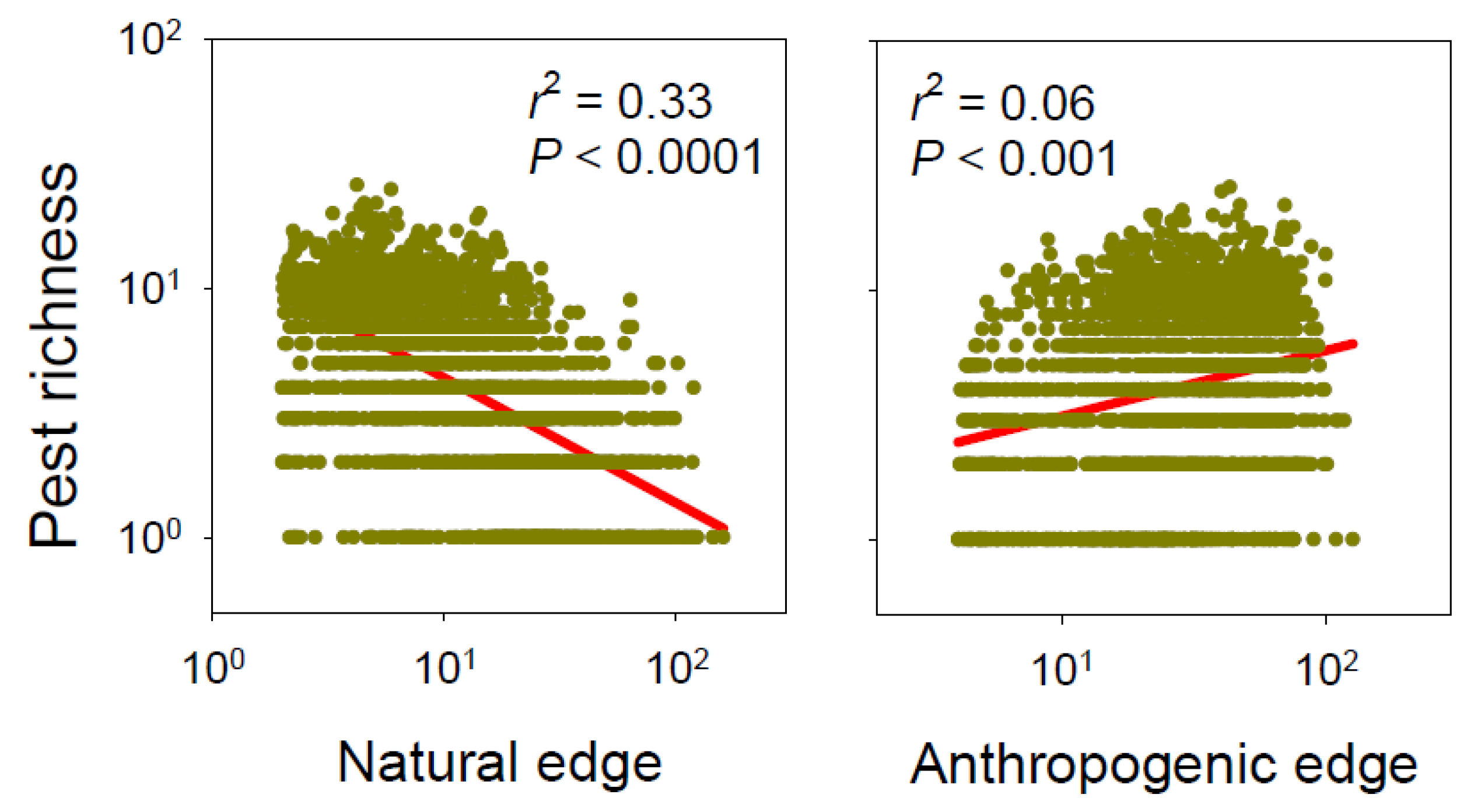
Finally, when forest edges were regrouped as natural (grass/shrub) and anthropogenic (agriculture and developed areas), the former had a negative, and the latter had a positive effect on overall pest richness (Figure 3). Also, in all analyses, when spatial autocorrelation was taken into account, the AICc values were much lower, indicating better fitted models (Table 1, Table 2, Table 3 and Table 4).
4. Discussion
Our results from a subcontinental analysis of forest ecosystems in the United States show that habitat fragmentation clearly influences the likelihood of nonnative pest invasions. The variation of overall pest richness across the forests reflects the net results of all component species showing positive, negative, and neutral responses to forest fragmentation and edge type (e.g., developed land/agriculture vs. grass/shrub) encountered [15]. In our study, the positive correlation of developed land area as edge (Fxde) with pest richness could be due to the propagule pressure related to human activities (e.g., disturbed land and ornamental plants associated with development).
Our study is different from previous studies, based on a single forest type or focusing on smaller-scales, including experimental studies [23]. When all forests across the entire the 48 conterminous US were considered, we found clear effects of habitat fragmentation on forest pest invasions. Most previous studies found positive responses between habitat fragmentation and variables, such as habitat diversity, landscape complementation, overall biodiversity, or individuals/populations of certain species [9]. In our analysis, the classes (core, islet, edge) described the structural role played by a forest pixel, in the context of other forest pixels. This may indicate some underlying mechanisms that have not been examined previously.
Given the strong signal of effects from developed edges, it may seem surprising that the effect of overall anthropogenic edges on insect/disease invasions was not as strong. Additionally, forest borders on a small scale have been found to be preferable habitats for both wood-boring and leaf-feeding insects, possibly because they provide a high variety of microsites and woody plant species [24]. Our results could be explained by the relatively minor impact of agricultural edges alone, and the recognition that most anthropogenic edge was agricultural and not developed. It is worth noting, however, that the positive relationship that we detected between defoliator diversity and agricultural edges, unlike for the other two insect feeding guilds, is consistent with previous work that showed the richness of leaf-feeding insects increased with agricultural land-use intensity, possibly the result of a decrease in natural predators along the intensity gradient [25].
The stronger responses of specialists to fragmentation and edge type than by generalist pests seem easier to understand. They could be due to the possibility that specialists are more responsive to higher host richness [26] and the possibility of the reduced abundance of unique host plants in more fragmented habitats and at forest edges (or ecotones) [15,27]. The positive relationship between specialists (but not generalists) and forest area is probably related to the presence of more hosts in greater amounts of forest, but the negative relationship with core area could reflect their preference for higher fragmentation.
Pathogens were much less sensitive to fragmentation and edge type than insect pests, which could be at least partly due to the fact that they are better dispersers [28]. However, both groups of species have similar proportions of generalists vs. specialists, thus the differences in their responses to fragmentation and edge type cannot be attributed to their species composition in terms of host specificity.
Difference in responses among insect guilds to landscape spatial structure were previously observed in smaller-scale studies [12]. For example, in a fragmentation experiment, Golden and Crist (1999) found that the species richness of sucking herbivores and the abundance of parasitoids declined with fragmentation, but that predators and chewing herbivores were not affected. They attributed the differences to habitat specificity requirements of guild members [23]. Similarly, Novais et al. (2016) found differences in the effect of canopy cover on different insect guilds, with percent canopy cover being positively associated with the richness of leaf-feeding insects, and negatively associated with the richness of sap-feeding insects, and with no significant relation to wood-boring insect richness [29]. These relationships, they concluded, depended in large part on the specific characteristics of the tropical agroforestry system they were studying and of the insect species present.
In general, we could attribute the differences among different groups or guilds to four major factors: (1) feeding preference associated with the nutrient characteristics and abundance of plant species at different edge types [30,31]; (2) chemical and mechanical defense mechanisms of the plants at forest edges (e.g., due to the different plant species compositions and associated herbivores) [32]; (3) predation risk on pests associated with certain predators and their abundance at different edges (including the “natural enemies hypothesis” prediction that higher plant diversity systems support more insect predators that more effectively control herbivore populations [29,33]), and (4) pest ecophysiological and life-history traits that are associated with ecological features and physical positions (i.e., forest layers) at forest edges [34].
We observed some differences in nonnative plant vs. pest invasions, although the related studies are at different scales (plot vs. county). For example, in a recent study of road effects on nonnative plant invasions in the forests of eastern United States, Riitters et al. found that nearby agriculture or developed land significantly increased the possibility of nonnative plant invasion at the plot level. In our study at the county-level, agriculture land as an edge or neighbor had a negative effect on forest insect/disease invasion (measured in richness) (Table 1). However, Riitters et al. concluded that the effect of a road is not as important as the overall landscape context (mosaic) associated with a road network, because when agriculture and developed lands occur together, invasion is increased over each land type alone [11]. For pest invasions, it is also expected that the overall landscape matrix or mosaics will be a major factor affecting invasions, although their importance may be scale-dependent [12].
Finally, the history of land use and fragmentation has not been fully incorporated into our fragmentation and pest invasion analyses, opening an excellent opportunity for further exploration in the future. Another urgent task would be to monitor individual pest species and to assess how they may respond to forest fragmentation and different edge types, especially those made by humans. Furthermore, other critical knowledge gaps exist, especially those that are related to scale or resolution. For example, higher resolution (e.g., plot-level) analyses on forest insect/disease invasion would reveal more detailed information regarding fragmentation and edge effects on invasive species.
Our findings have significant implications for large-scale habitat restoration, land use, and management. For example, how do other human-made edges such as clearcuttings (from logging) and prescribed burned areas affect forest habitat invasibility? Also, future investigations should examine the novel and cross-scale interactions between native and nonnative insects and pathogens, and their interactions with other organisms such as plants, birds, and larger herbivores [35,36]. Under ongoing climate and land use changes, forest ecosystems may be under greater pressure, or even near tipping points, and research on future fragmentation and edge effects is urgently required.
5. Conclusions
Our subcontinental-scale analysis showed that habitat fragmentation and the resulting edges of various types clearly affect patterns of nonnative forest insect and disease invasions. More importantly, in addition to commonly recognized edge effects, different types of edges (i.e., neighboring habitats) play a major role in regulating invasion patterns. Particularly, the developed land edge type contributes the most to the richness of nonnative insects/diseases, either as combined or in separate guilds, or as feeding generalists or specialists. This observation may largely reflect anthropogenic influences, including increased propagule pressure associated with human activities. These findings from regional and continental-scale research will help future efforts in managing forest pest invasions, and in guiding ongoing and planned forest restorations [37].
Supplementary Materials
Figure S1, Table S1, and Table S2 are available online at https://www.mdpi.com/1999-4907/9/12/744/s1.
Author Contributions
Q.G. conceived the research and drafted the manuscript. K.H.R. and K.M.P. provided the forest fragmentation and pest data, respectively. All authors performed data analyses and wrote the paper.
Funding
This research was partly supported by NSF Macrosystems Biology grants (DEB-1241932 and DEB-1638702).
Acknowledgments
We thank reviewers for constructive comments on earlier versions of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McCullough, D.G.; Work, T.T.; Cavey, J.F.; Liebhold, A.M.; Marshall, D. Interceptions of nonindigenous plant pests at US ports of entry and border crossings over a 17-year period. Biol. Invasions 2006, 8, 611–630. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Liebhold, A.M.; McCullough, D.G.; Blackburn, L.M.; Frankel, S.J.; Von Holle, B.; Aukema, J.E. A highly aggregated geographical distribution of forest pest invasions in the USA. Divers. Distrib. 2013, 19, 1208–1216. [Google Scholar] [CrossRef]
- Oswalt, C.; Fei, S.; Guo, Q.; Iannone, B., III; Oswalt, S.; Pijanowski, B.; Potter, K. A subcontinental view of forest plant invasions. NeoBiota 2015, 24, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Rejmanek, M.; Wen, J. Geographical, socioeconomic, and ecological determinants of exotic plant naturalization in the United States: Insights and updates from improved data. NeoBiota 2012, 12, 41. [Google Scholar] [CrossRef]
- Iannone, B.V.; Potter, K.M.; Guo, Q.; Liebhold, A.M.; Pijanowski, B.C.; Oswalt, C.M.; Fei, S. Biological invasion hotspots: A trait-based perspective reveals new sub-continental patterns. Ecography 2016, 39, 961–969. [Google Scholar] [CrossRef]
- Guo, Q. Plant hybridization: The role of human disturbance and biological invasion. Divers. Distrib. 2014, 20, 1345–1354. [Google Scholar] [CrossRef]
- Riitters, R.; Potter, K.M.; Iannone, B.V., III; Oswalt, C.; Guo, Q.; Fei, S. Exposure of protected and unprotected forest to plant invasions in the eastern United States. Forests 2018, 9, 723. [Google Scholar] [CrossRef]
- Fahrig, L. Ecological responses to habitat fragmentation per se. Ann. Rev. Ecol. Evol. Syst. 2017, 48, 1–23. [Google Scholar] [CrossRef]
- Wade, T.G.; Riitters, K.H.; Wickham, J.D.; Jones, K.B. Distribution and causes of global forest fragmentation. Conser. Ecol. 2003, 7, 7. [Google Scholar] [CrossRef]
- Riitters, K.; Potter, K.; Iannone, B.V.; Oswalt, C.; Fei, S.; Guo, Q. Landscape correlates of forest plant invasions: A high-resolution analysis across the eastern United States. Divers. Distrib. 2018, 24, 274–284. [Google Scholar] [CrossRef]
- Pauchard, A.; Alaback, P.B. Edge type defines alien plant species invasions along Pinus contorta burned, highway and clearcut forest edges. For. Ecol. Manag. 2006, 223, 327–335. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E.; Burton, P.J.; Chen, J.; Brosofske, K.D.; Saunders, S.C.; Euskirchen, E.S.; Roberts, D.; Jaiteh, M.S.; Esseen, P.A. Edge influence on forest structure and composition in fragmented landscapes. Conserv. Biol. 2005, 19, 768–782. [Google Scholar] [CrossRef]
- Slancarova, J.; Benes, J.; Kristynek, M.; Kepka, P.; Konvicka, M. Does the surrounding landscape heterogeneity affect the butterflies of insular grassland reserves? A contrast between composition and configuration. J. Insect Conser. 2014, 18, 1–12. [Google Scholar] [CrossRef]
- Ries, L.; Sisk, T.D. A predictive model of edge effects. Ecology 2004, 85, 2917–2926. [Google Scholar] [CrossRef]
- Kappler, R.H.; Knight, K.S.; Koch, J.; Root, K.V. Neighboring Tree Effects and Soil Nutrient Associations with Surviving Green Ash (Fraxinus pennsylvanica) in an Emerald Ash Borer (Agrilus planipennis) Infested Floodplain Forest. Forests 2018, 9, 183. [Google Scholar] [CrossRef]
- USDA Forest Service. Alien Forest Pest Explorer Database. Available online: https://www.nrs.fs.fed.us/tools/afpe (accessed on 25 June 2018).
- USGS. National Geospatial Data Asset (NGDA) Land Use Land Cover (2011 Edition); US Geological Survey: Sioux Falls, SD, USA, 2014.
- Homer, C.; Dewitz, J.; Yang, L.; Jin, S.; Danielson, P.; Xian, G.; Coulston, J.; Herold, N.; Wickham, J.; Megown, K. Completion of the 2011 National Land Cover Database for the conterminous United States–representing a decade of land cover change information. Photogramm. Eng. Remote Sens. 2015, 81, 345–354. [Google Scholar]
- Loveland, T.R.; Dwyer, J.L. Landsat: Building a strong future. Remote Sens. Environ. 2012, 122, 22–29. [Google Scholar] [CrossRef]
- Wickham, J.; Stehman, S.V.; Gass, L.; Dewitz, J.A.; Sorenson, D.G.; Granneman, B.J.; Poss, R.V.; Baer, L.A. Thematic accuracy assessment of the 2011 national land cover database (NLCD). Remote Sens. Environ. 2017, 191, 328–341. [Google Scholar] [CrossRef]
- Soille, P.; Vogt, P. Morphological segmentation of binary patterns. Pattern Recognit. Lett. 2009, 30, 456–459. [Google Scholar] [CrossRef]
- Golden, D.M.; Crist, T.O. Experimental effects of habitat fragmentation on old-field canopy insects: Community, guild and species responses. Oecologia 1999, 118, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Baranchikov, Y.N.; Mattson, W.J.; Hain, F.P.; Payne, T.L. Forest Insect Guilds: Patterns of Interaction with Host Trees; General Technical Report NE-153; U.S. Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Radnor, PA, USA, 1991.
- Klein, A.-M.; Steffan-Dewenter, I.; Tscharntke, T. Predator—Prey ratios on cocoa along a land-use gradient in Indonesia. Biodivers. Conserv. 2002, 11, 683–693. [Google Scholar] [CrossRef]
- Lin, Y.P.; Cook, D.H.; Gullan, P.J.; Cook, L.G. Does host-plant diversity explain species richness in insects? A test using Coccidae (Hemiptera). Ecol. Entomol. 2015, 40, 299–306. [Google Scholar] [CrossRef]
- Valente, J.J.; Betts, M.G. Response to fragmentation by avian communities is mediated by species traits. Divers. Distrib. 2018, in press. [Google Scholar] [CrossRef]
- Brown, J.K.; Hovmoller, M.S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Novais, S.; Macedo-Reis, L.E.; DaRocha, W.D.; Neves, F.S. Effects of habitat management on different feeding guilds of herbivorous insects in cacao agroforestry systems. Rev. Biol. Trop. 2016, 64, 763–777. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, M.R.; Gonzalez, E.; Salvo, A.; Valladares, G. Not all in the same boat: Trends and mechanisms in herbivory responses to forest fragmentation differ among insect guilds. Arthropod-Plant Interact. 2014, 8, 593–603. [Google Scholar] [CrossRef]
- Barbaro, L.; Van Halder, I. Linking bird, carabid beetle and butterfly life-history traits to habitat fragmentation in mosaic landscapes. Ecography 2009, 32, 321–333. [Google Scholar] [CrossRef]
- Yamawo, A.; Tagawa, J.; Hada, Y.; Suzuki, N. Different combinations of multiple defence traits in an extrafloral nectary-bearing plant growing under various habitat conditions. J. Ecol. 2014, 102, 238–247. [Google Scholar] [CrossRef]
- Root, R.B. Organization of a plant-arthropod association in simple and diverse habitats: The fauna of collards (Brassica oleracea). Ecol. Monogr. 1973, 43, 95–124. [Google Scholar] [CrossRef]
- McAvoy, T.J.; Mays, R.; Johnson, N.G.; Salom, S.M. The effects of shade, fertilizer, and pruning on eastern hemlock trees and hemlock woolly adelgid. Forests 2017, 8, 156. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, D.; Guo, Q.; Liu, J.; Wang, L. Interactive effects of large herbivores and plant diversity on insect abundance in a meadow steppe in China. Agric. Ecosyst. Environ. 2015, 212, 245–252. [Google Scholar] [CrossRef]
- Fei, S.; Guo, Q.; Potter, K. Macrosystems ecology: Novel methods and new understanding of multi-scale patterns and processes. Landsc. Ecol. 2016, 31, 217–218. [Google Scholar] [CrossRef]
- Guo, Q.; Brockway, D.G.; Larson, D.L.; Wang, D.; Ren, H. Improving ecological restoration to curb biotic invasion—A practical guide. Invas. Plant Sci. Manag. 2019, in press. [Google Scholar]
Figure 1.
Illustrations of (a) morphological spatial pattern analysis classes, and (b) calculation of forest adjacencies. In (a), each pixel is labeled by the indicated class; in (b), an “edge” refers to an adjacent pair of pixels (cardinal directions), the edges not involving forest are ignored, and each pixel is characterized by the edges in its unique surrounding “window”.
Figure 1.
Illustrations of (a) morphological spatial pattern analysis classes, and (b) calculation of forest adjacencies. In (a), each pixel is labeled by the indicated class; in (b), an “edge” refers to an adjacent pair of pixels (cardinal directions), the edges not involving forest are ignored, and each pixel is characterized by the edges in its unique surrounding “window”.

Figure 2.
Effects of forest core/edge ratio on nonnative pest richness across the forests in the conterminous continental United States (N = 2098 counties).
Figure 2.
Effects of forest core/edge ratio on nonnative pest richness across the forests in the conterminous continental United States (N = 2098 counties).
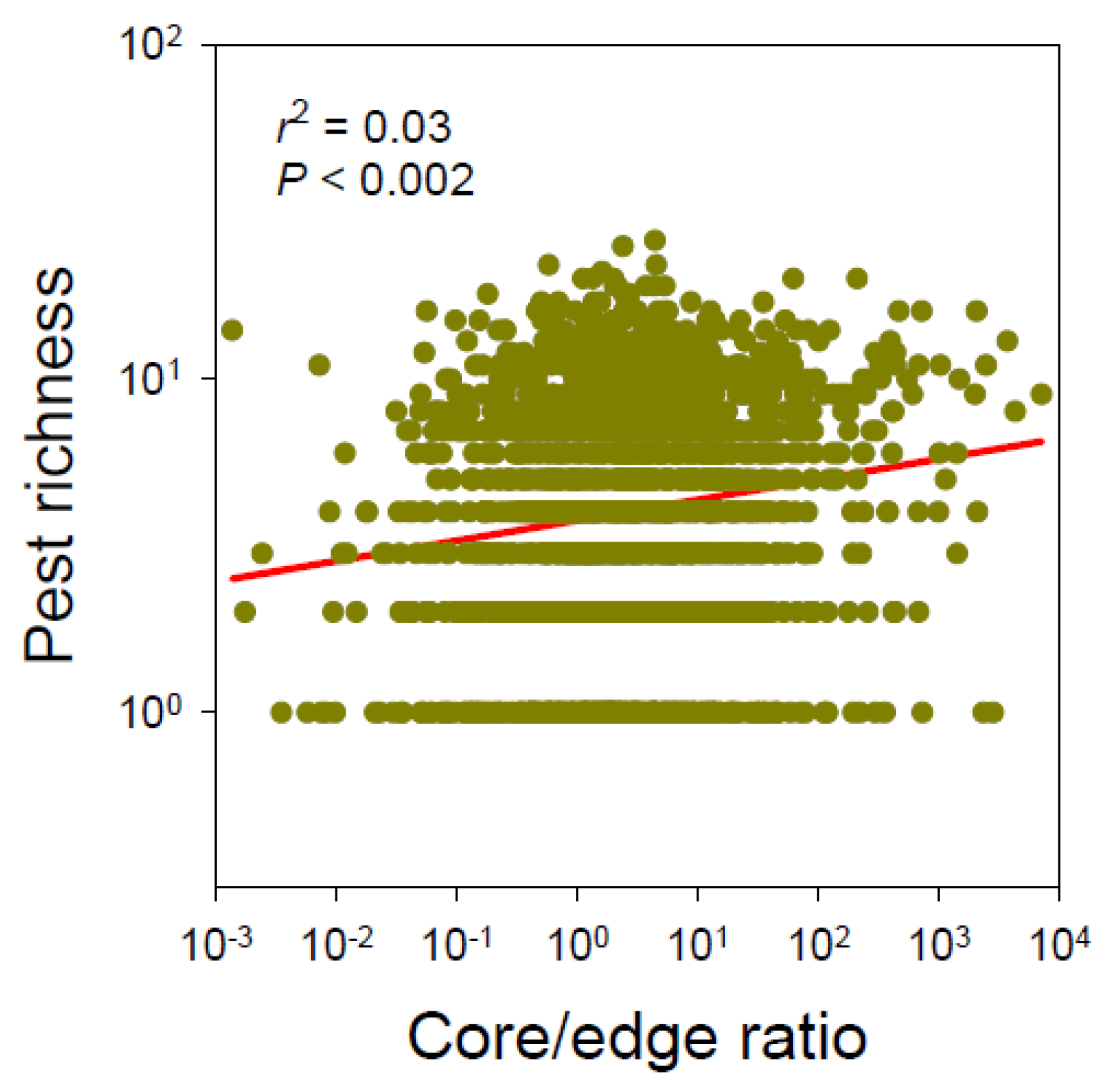
Figure 3.
Effects of natural and anthropogenic forest edges on nonnative pest richness across the forests in the conterminous continental United States (N = 2098 counties).
Figure 3.
Effects of natural and anthropogenic forest edges on nonnative pest richness across the forests in the conterminous continental United States (N = 2098 counties).

{kind=link}
{kind=link}
{kind=link}
Table 1.
Effects of forest fragmentation and edge types on pest richness across the forests in the conterminous United States based on the ordinary-least-squares (OLS) estimation and spatial autoregression analyses (SAR). Data were aggregated to the county level (N = 2098) and were log-transformed before analysis. Forest edge types: Fxag = forest|agriculture, Fxde = forest|developed, and Fxgs = forest|grass-shrub.
Table 1.
Effects of forest fragmentation and edge types on pest richness across the forests in the conterminous United States based on the ordinary-least-squares (OLS) estimation and spatial autoregression analyses (SAR). Data were aggregated to the county level (N = 2098) and were log-transformed before analysis. Forest edge types: Fxag = forest|agriculture, Fxde = forest|developed, and Fxgs = forest|grass-shrub.
| Variable | OLS Coefficient | SAR Coefficient | T | p |
|---|---|---|---|---|
| Constant | 0.726 | 0.374 | 2.309 | 0.021 |
| Forest area | 0.235 | 0.157 | 1.597 | 0.110 |
| Core | −0.143 | −0.051 | −0.654 | 0.513 |
| Islet | −0.014 | −0.028 | −1.206 | 0.228 |
| Edge | −0.047 | −0.037 | −4.119 | <0.001 |
| Fxag | −0.075 | 0.002 | 0.095 | 0.925 |
| Fxde | 0.240 | 0.260 | 9.960 | <0.001 |
| Fxgs | −0.497 | −0.348 | −15.947 | <0.001 |
OLS (r2 = 0.370, AICc: 636.063); SAR (r2 = 0.435, AICc: 406.328).
Table 2.
Effects of forest fragmentation and edge types on pest richness (generalists vs. specialists) across the forests in the conterminous United States, based on the ordinary-least-squares (OLS) estimation and spatial autoregression analyses (SAR). Data were aggregated to the county level (N = 2098) and were log(x + 1)-transformed before analysis.
Table 2.
Effects of forest fragmentation and edge types on pest richness (generalists vs. specialists) across the forests in the conterminous United States, based on the ordinary-least-squares (OLS) estimation and spatial autoregression analyses (SAR). Data were aggregated to the county level (N = 2098) and were log(x + 1)-transformed before analysis.
| Generalists | Specialists | |||||
|---|---|---|---|---|---|---|
| r2 | AICc | r2 | AICc | |||
| OLS | 0.355 | −1596.820 | 0.297 | 218.502 | ||
| SAR | 0.362 | −1617.926 | 0.368 | −5.357 | ||
| Variable | OLS Coefficient | SAR Coefficient | T | OLS Coefficient | SAR Coefficient | T |
| Constant | 0.072 | −0.077 | −0.736 | 0.8 | 0.532 | 3.519 *** |
| Forest area | 0.09 | 0.052 | 0.838 | 0.362 | 0.247 | 2.761 ** |
| Core | −0.02 | 0.017 | 0.336 | −0.278 | −0.15 | −2.113 * |
| Islet | −0.04 | −0.03 | −2.060 * | 0.005 | −0.021 | −1.012 |
| Edge | −0.005 | −0.004 | −0.676 | −0.044 | −0.035 | −4.34 ** |
| Fxag | 0.059 | 0.082 | 4.927 ** | −0.153 | −0.046 | −1.896 |
| Fxde | 0.261 | 0.239 | 12.839 *** | 0.158 | 0.192 | 7.141 ** |
| Fxgs | −0.203 | −0.134 | −8.439 ** | −0.489 | −0.341 | −14.946 *** |
* p < 0.05, ** p < 0.01, *** p < 0.0001.
Table 3.
Effects of forest fragmentation and edge types on pest richness (pathogens vs. insects) across the forests in the conterminous United States based on the ordinary-least-squares (OLS) estimation and spatial autoregression analyses (SAR). Data were aggregated to the county level (N = 2098) and were log(x + 1)-transformed before analysis.
Table 3.
Effects of forest fragmentation and edge types on pest richness (pathogens vs. insects) across the forests in the conterminous United States based on the ordinary-least-squares (OLS) estimation and spatial autoregression analyses (SAR). Data were aggregated to the county level (N = 2098) and were log(x + 1)-transformed before analysis.
| Pathogens | Insects | |||||
|---|---|---|---|---|---|---|
| r2 | AICc | r2 | AICc | |||
| OLS | 0.404 | −1108.708 | 0.251 | 21.133 | ||
| SAR | 0.415 | −1146.721 | 0.360 | −309.589 | ||
| Variable | OLS Coefficient | SAR Coefficient | T | OLS Coefficient | SAR Coefficient | T |
| Constant | 0.375 | 0.006 | 0.056 | 0.383 | 0.353 | 2.482 * |
| Forest area | 0.06 | 0.062 | 0.908 | 0.49 | 0.307 | 3.654 *** |
| Core | 0.037 | 0.035 | 0.646 | −0.392 | −0.202 | −3.027 ** |
| Islet | −0.031 | <0.001 | −0.055 | −0.013 | −0.062 | −3.113 ** |
| Edge | −0.031 | −0.02 | −3.291 ** | −0.019 | −0.018 | −2.40 * |
| Fxag | 0.009 | −0.01 | −0.529 | −0.122 | 0.034 | 1.505 |
| Fxde | 0.182 | 0.157 | 7.659 *** | 0.228 | 0.268 | 10.591 *** |
| Fxgs | −0.331 | −0.248 | −14.180 *** | −0.397 | −0.257 | −11.963 *** |
* p < 0.05, ** p < 0.01, *** p < 0.0001.
Table 4.
Effects of forest fragmentation and edge types on insect pest richness (three guilds) across the forests in the conterminous United States based on the ordinary-least-squares (OLS) estimation and spatial autoregression analyses (SAR). Data were aggregated to the county level (N = 2098) and were log (x+1)-transformed before analysis.
Table 4.
Effects of forest fragmentation and edge types on insect pest richness (three guilds) across the forests in the conterminous United States based on the ordinary-least-squares (OLS) estimation and spatial autoregression analyses (SAR). Data were aggregated to the county level (N = 2098) and were log (x+1)-transformed before analysis.
| Defoliator | Sap Feeder | Wood Borer | |||||||
|---|---|---|---|---|---|---|---|---|---|
| r2 | AICc | r2 | AICc | r2 | AICc | ||||
| OLS | 0.350 | −968.647 | 0.233 | −1524.781 | 0.174 | −398.459 | |||
| SAR | 0.381 | −1069.443 | 0.314 | −1757.464 | 0.25 | −601.435 | |||
| Variable | OLS Coefficient | SAR Coefficient | T | OLS Coefficient | SAR Coefficient | T | OLS Coefficient | SAR Coefficient | T |
| Constant | −0.244 | −0.287 | −2.407 * | 0.083 | 0.197 | 1.952 | 0.566 | 0.565 | 0.267 |
| Forest area | 0.377 | 0.269 | 3.809 *** | 0.231 | 0.141 | 2.358* | 0.229 | 0.05 | 0.644 |
| Core | −0.196 | −0.106 | −1.903 | −0.178 | −0.084 | −1.774 | −0.305 | −0.109 | −1.755 |
| Islet | −0.128 | −0.109 | −6.541 *** | 0.022 | −0.021 | −1.505 | 0.123 | 0.06 | 3.228 *** |
| Edge | −0.005 | −0.004 | −0.672 | 0.001 | −0.006 | −1.033 | −0.021 | −0.019 | −2.651 ** |
| Fxag | 0.027 | 0.076 | 4.010 *** | −0.232 | −0.086 | −5.389 *** | −0.092 | 0.017 | 0.815 |
| Fxde | 0.264 | 0.233 | 10.970 *** | 0.108 | 0.125 | 6.990 *** | 0.101 | 0.163 | 6.921 *** |
| Fxgs | −0.236 | −0.155 | −8.616 *** | −0.2 | −0.148 | −9.689 *** | −0.296 | −0.174 | −8.722 *** |
* p < 0.05, ** p < 0.01, *** p < 0.0001.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Guo, Q.; Riitters, K.H.; Potter, K.M. A Subcontinental Analysis of Forest Fragmentation Effects on Insect and Disease Invasion. Forests 2018, 9, 744. https://doi.org/10.3390/f9120744
AMA Style
Guo Q, Riitters KH, Potter KM. A Subcontinental Analysis of Forest Fragmentation Effects on Insect and Disease Invasion. Forests. 2018; 9(12):744. https://doi.org/10.3390/f9120744
Chicago/Turabian StyleGuo, Qinfeng, Kurt H. Riitters, and Kevin M. Potter. 2018. "A Subcontinental Analysis of Forest Fragmentation Effects on Insect and Disease Invasion" Forests 9, no. 12: 744. https://doi.org/10.3390/f9120744
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.