Evaluation of the Susceptibility of Several Czech Conifer Provenances to Fusarium circinatum
by
 , , and
, , and
Jorge Martín-García
1,2,* ,
,
Aneta Lukačevičová
3,
Juan Asdrúbal Flores-Pacheco
2,4,5,
Julio Javier Diez
2,4 and
Miloň Dvořák
3 1
Department of Biology, CESAM (Centre for Environmental and Marine Studies), University of Aveiro, Campus Universitario de Santiago, 3810-193 Aveiro, Portugal
2
Sustainable Forest Management Research Institute, University of Valladolid—INIA, Avenida de Madrid 44, 34071 Palencia, Spain
3
Department of Forest Protection and Wildlife Management, Faculty of Forestry and Wood Technology, Mendel University in Brno, Zemědělská 3, Brno 61300, Czech Republic
4
Department of Plant Production and Forest Resources, University of Valladolid, Avenida de Madrid 44, 34071 Palencia, Spain
5
Facultad de Recursos Naturales y Medio Ambiente, Bluefields Indian & Caribbean University—BICU, Avenida Universitaria, Apartado Postal N 88, Bluefields, Nicaragua
*
Author to whom correspondence should be addressed.
Forests 2018, 9(2), 72; https://doi.org/10.3390/f9020072
Submission received: 29 December 2017
/
Revised: 18 January 2018
/
Accepted: 24 January 2018
/
Published: 1 February 2018
(This article belongs to the Special Issue Pine Pitch Canker—Strategies for Management of Gibberella Circinata in Greenhouses and Forests (PINESTRENGTH))
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Pine pitch canker (PPC), caused by Fusarium circinatum, is considered among the most important diseases affecting pines in many locations throughout the world. In Europe, F. circinatum is currently present in the Iberian Peninsula, posing a high risk of its spread into currently disease-free countries in Europe. In the present study, the susceptibility of Pinus sylvestris, Picea abies, and Larix decidua originating in the Czech Republic to F. circinatum was tested. Furthermore, the presence of asymptomatic yet infected seedlings was also checked. This study demonstrated the pathogenicity of F. circinatum to the Czech provenance of P. sylvestris, whereas Picea abies and Larix decidua proved to be tolerant. The reisolation of F. circinatum beyond the inoculation point demonstrated that this quarantine pathogen is able to infect the three conifers tested, giving rise to asymptomatic seedlings for at least eight and a half months. To our knowledge, this is the first study in which the presence of symptomless seedlings has been recorded in the genera Picea and Larix. This finding points out that the European legislation would fail to avoid the risk of new introductions via symptomless seedlings, since this legislation is only restricted to plants of the genus Pinus and the species Pseudotsuga menziesii.
1. Introduction
Forests are important ecosystems from both ecological and economic perspectives. Approximately 33.85% (2,669,850 hectares) of the Czech Republic is aforested, of which 72.1% are conifer forests [1]. Among the conifers, Norway spruce (Picea abies L.), Scots pine (Pinus sylvestris L.), and to a lesser extent, European larch (Larix decidua Mill.) are the most important species. In fact, P. abies and P. sylvestris are native species covering 1,312,204 and 425,687 ha, respectively (i.e., 50.5% and 16.4% of the total forest area in the Czech Republic) [1].
Nowadays, invasive alien species are a major threat to forest ecosystems. In particular, forest pathogens introduced into countries as a result of globalization of trade and free market practices are a major challenge for Europe [2,3]. This, together with climatic change, can increase the risk of spreading and establishing new pathogens in currently disease-free countries [4].
European and Mediterranean Plant Protection Organization (EPPO) has recorded 35 fungi in the List A1 of pests recommended for regulation as quarantine pest (absent from the EPPO region) and 32 in the List A2 (present in the EPPO region but not widely distributed). Fusarium circinatum Nirenberg & O’Donnell (teleomorph = Gibberella circinata), which causes pine pitch canker (PPC), is included in the List A2 [5]. Fusarium circinatum was first detected in 1945 in the south-eastern United States [6], and since then has spread widely to Haiti [7], South Africa [8], Mexico [9], Chile [10], South Korea [11], Japan [12], Uruguay [13], Colombia [14], and Brazil [15], among other regions. Spain was the first European country where the disease was detected [16,17]. The pathogen is currently established in forests in Spain, mainly in the commercial Pinus radiata D. Don and, to a lesser extent, in Pinus pinaster Ait. plantations. The pathogen has also been reported in France [18], Italy [19], and Portugal [20], although in France and Italy it is now considered to have been eradicated.
Fusarium circinatum, the causal agent of PPC, is now considered among the most important pathogens of Pinaceae in the world, devastating Pinus seedlings and mature trees in many countries globally. In fact, the European Food Safety Authority (EFSA) has established that under the current host distribution and climatic conditions, the potentially at risk areas cover over 10 million hectares of pine forests in Europe [21]. Furthermore, several studies have already highlighted that climate change will lead to increased suitability of currently disease-free areas for F. circinatum in the near to medium term future [22,23].
At least 60 species of Pinus along with Pseudotsuga menziesii (Mirb.) Franco are known to be susceptible to PCC [16,21,24,25]. Variation in susceptibility occurs not only among species [26,27,28,29,30], but also among provenances [31,32,33,34,35]. Thereby, taking into account the high genetic variation among the European Scots pine [36,37,38], Norway spruce [39], and European larch populations [40,41], testing the susceptibility of European provenances (e.g., from the Czech Republic) should be a priority.
It is well known that the occurrence of symptoms also depends on the host, as well as biotic and abiotic conditions [25]. Furthermore, the presence of asymptomatic yet infected seedlings is not unusual [42,43,44,45,46,47], which implies a high risk of PPC spread from nurseries to the field due to the latent phase of the infection. Thereby, finding those species or provenances that, not showing symptoms, might trigger the spread of the pathogen into currently disease-free countries in Europe is also a priority.
The work reported here has a twofold objective: (1) to test the susceptibility of P. sylvestris, P. abies, and L. decidua of Czech provenances; and (2) to check whether the presence of asymptomatic yet infected seedlings is common among these species.
2. Material and Methods
2.1. Fungal Isolates and Plant Material
The Fusarium circinatum isolate (FcCa6) used in this work belongs to mating type 2 (MAT-2) and was isolated from an infected Pinus radiata tree located in Comillas (Cantabria, Northern Spain; GPS: 4°17′17.706″ W; 43°20′5.033″ N; 265 m above sea level) [28,48,49,50]. Plant material consisted of seedlings of three Czech conifers: P. sylvestris, P. abies, and L. decidua. Scots pine seedlings were 1.5-year-old plants from South Bohemian Basins, 3.5-year-old Norway spruce seedlings from Brdy Highland (Middle Bohemia), and 1.5-year-old European larch from Bohemian-Moravian Highlands. The seedlings were obtained from a mix of seeds of cones from forest stands designated by the Czech forest legislation (Act No. 149/2003 Coll. and Decree No. 29/2004 Coll.) as “selected” (i.e., forest stand of at least 1 ha, with more than 40 trees of at least 60 years, having the phenotypic classification “A” or “B”, with at least 100 m distance from any forest stand with the worst classification “D”) and phenotypic classification “B”. These forest stands were located at middle altitudes (500–600 m above sea level), with mean annual temperature 6.5–7.5 °C, and annual precipitations 690–800 mm [51].
2.2. Pathogenicity Tests
The spore suspension of F. circinatum was cultured on potato dextrose broth (PDB medium). For that, an Erlenmeyer flask, containing 1 L of PDB and five mycelial agar plugs (diameter 4–5 mm) obtained from the margin of an actively growing colony, was placed on an orbital shaker at 180 cycles per minute for 24 h at 25 °C. Finally, the spore suspension was obtained by filtering twice through sterile cheesecloth to remove hyphae. The spore suspension was adjusted with a hemocytometer at 106 spores ml−1.
Pathogenicity tests were carried out by the stem inoculation technique [52]. Thus, a wound was made with a sterile scalpel 5–7 cm above the collar of each plant after removal of needles from that area. Then 25 seedlings of each species in full growth were inoculated with 100 μL of the spore suspension (106 spore mL−1) and another 25 control seedlings were mock-inoculated in the same way with sterilized distilled water instead of spore suspensions. The inoculated wound was immediately sealed with Parafilm® to prevent drying, and the seedlings were placed in a growth chamber at 21.5 °C with a 16/8 h light/dark photoperiod, following a completely randomized design. Watering and other procedures were conducted as per routine nursery practice, except that no fertilizers or fungicides were applied.
Symptoms were estimated three times per week according to the scale (slightly modified) published by Correll et al. [53] for over three months (102 days), where 0 = healthy plant, 1 = resin and/or necrosis at the point of inoculation and healthy foliage, 2 = resin and/or necrosis beyond the point of inoculation, 3 = accentuated wilting and appreciable dieback, 4 = dead plant.
The presence of asymptomatic yet infected seedlings was checked at three stages: 2, 5, and 8.5 months after inoculation. Five seedlings per species were checked at 2 months after inoculation and ten seedlings per species at 5 and 8.5 months after inoculation (except for P. sylvestris, because all inoculated seedlings had died until 8.5 months after inoculation). Reisolation of the pathogen was carried out from stem tissues >2.5 cm beyond the point of inoculation (upward and downward). For the reisolation, the fragments were sterilized by: dipping in sterile distilled water for 3 min, followed by shaking in 3% sodium hypochlorite (v/v) for 2 min, thereafter shaking in 70% ethanol (v/v) for 2 min, and finally dipping for 5 min in sterile distilled water to remove any remaining traces of disinfectants. The samples were then dried for 1–5 min in a sterile laminar flow cabinet on sterile filter paper, before being cut in small pieces for plating on potato dextrose agar (PDA) with 0.5 g/L of streptomycin sulfate (to prevent bacterial growth) [54]. The plates were placed in growth chambers in the dark at 25 °C and once the colonies started growing, they were subcultured on Spezieller Nährstoffarmer Agar (SNA) media, in which it is feasible to identify the distinguishing features of F. circinatum using a stereomicroscope (e.g., the coiled sterile hyphae) [55].
2.3. Statistical Analyses
Analyses of variance (ANOVAs) and multiple comparison procedures were performed to test the effects of F. circinatum on inoculated seedlings. The scale of symptoms was previously transformed to a quantitative variable using the area under the disease progress curve (AUDPC), calculated as the sum of the area of the corresponding trapezoids for the trial duration [56,57]. As the data violated two of the ANOVA assumptions (normality and homogeneity of variances), robust statistical methods were applied [58]. In particular, heteroscedastic one-way ANOVAs were performed using the generalized Welch procedure and a 0.1 trimmed mean transformation. The ANOVAs were carried out using the “Wilcox’ Robust Statistics (WRS)” package. Survival analysis based on the nonparametric estimator Kaplan–Meier [59] was performed with the “Survival” package [60] to test the mortality up to the end of the experiment (102 days). Survival curves were created with the “Survfit” function and the differences between the curves were tested with the “Survdiff” function. All analyses were performed using R software environment (R Foundation for Statistical Computing, Vienna, Austria).
3. Results
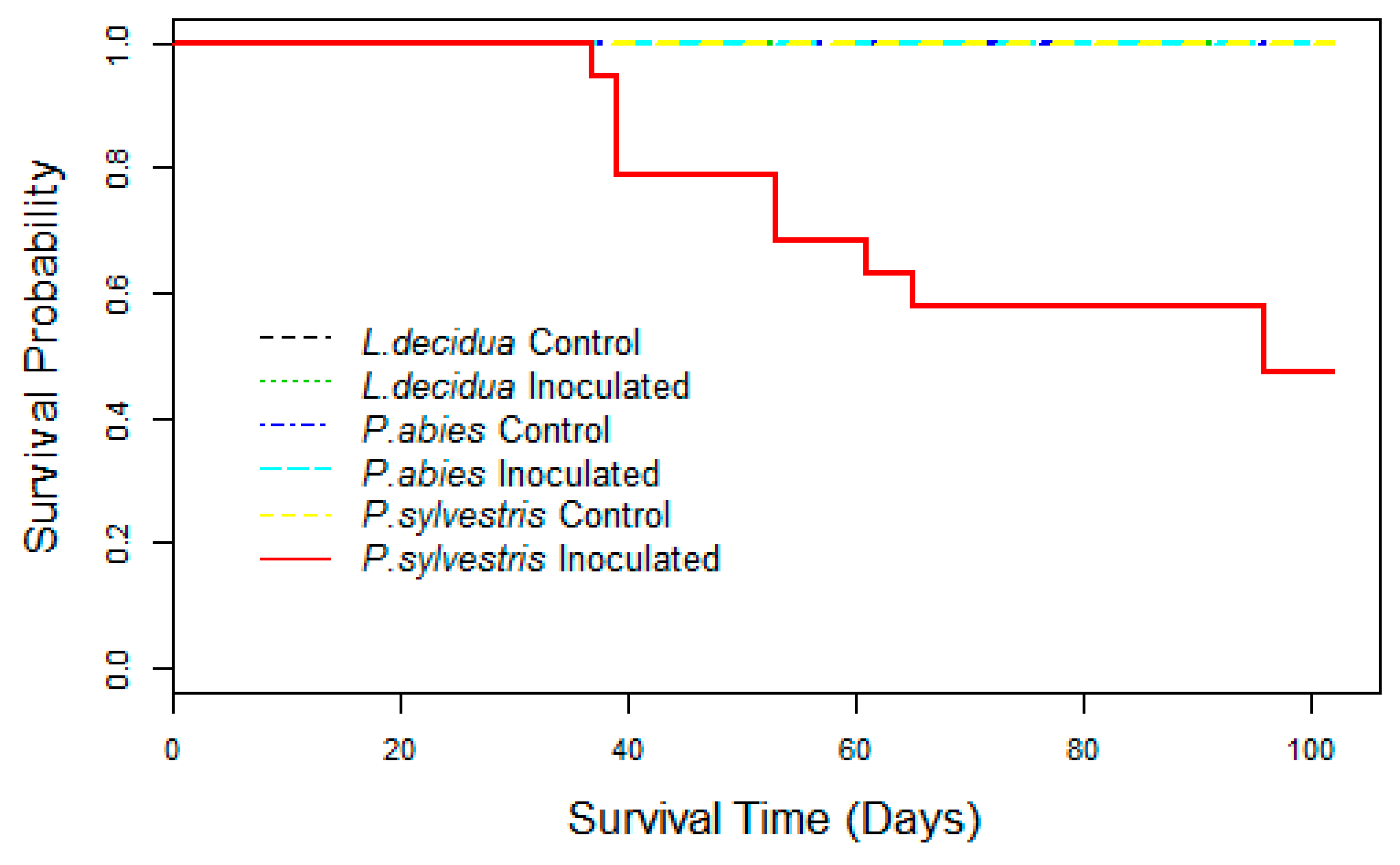
Pinus sylvestris seedlings began to die 37 days after inoculation (days after inoculation), whereas no mortality was recorded in the control seedlings. The survival analyses revealed significant differences between treatments (χ2 = 17.5, p < 0.001) (Figure 1). Nearly 50 percent of P. sylvestris inoculated seedlings died within three months following the inoculation and no survival was recorded eight months after inoculation. Mortality was not recorded in inoculated P. abies and L. decidua seedlings during the trial. Control seedlings did not die during the experimental assay.
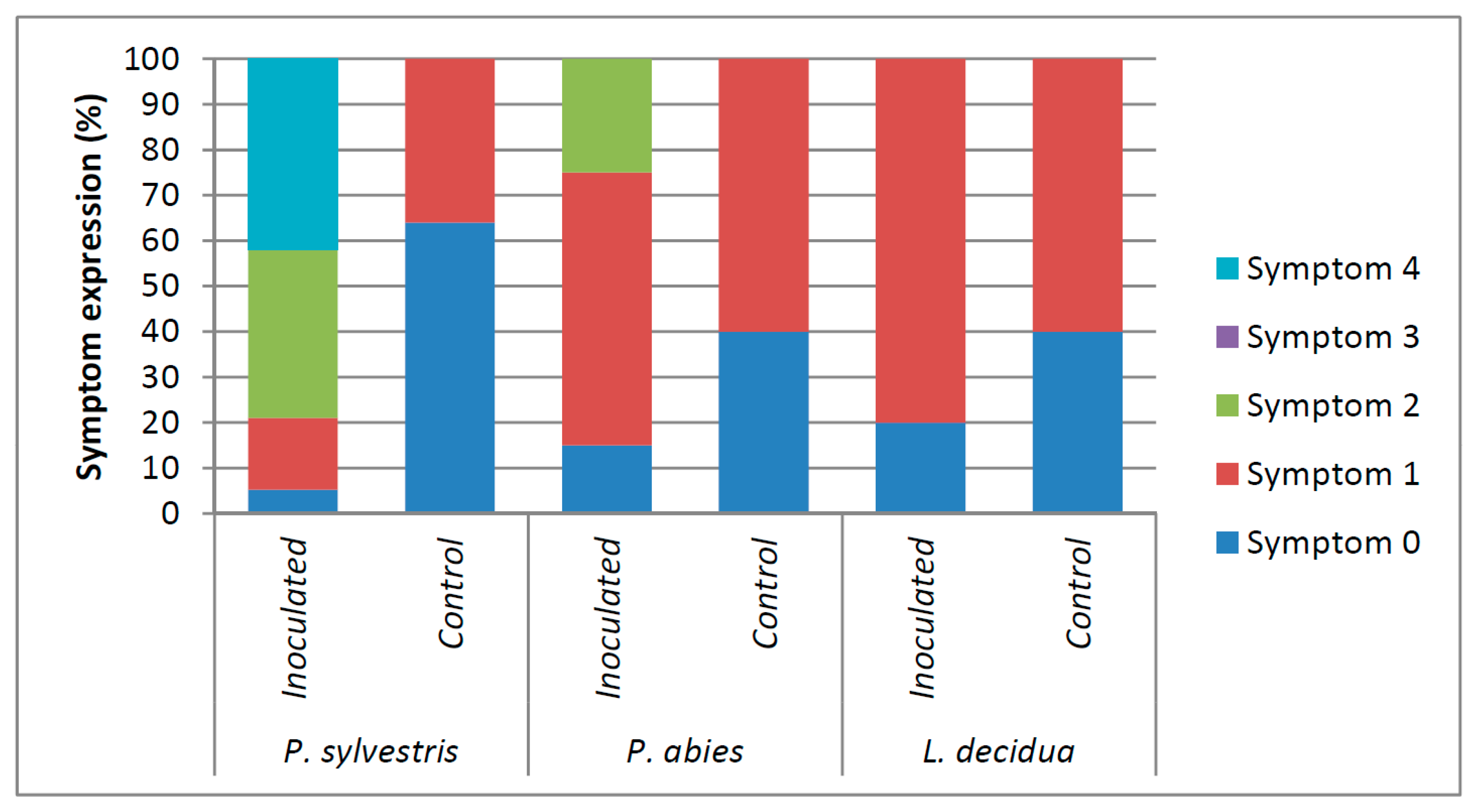
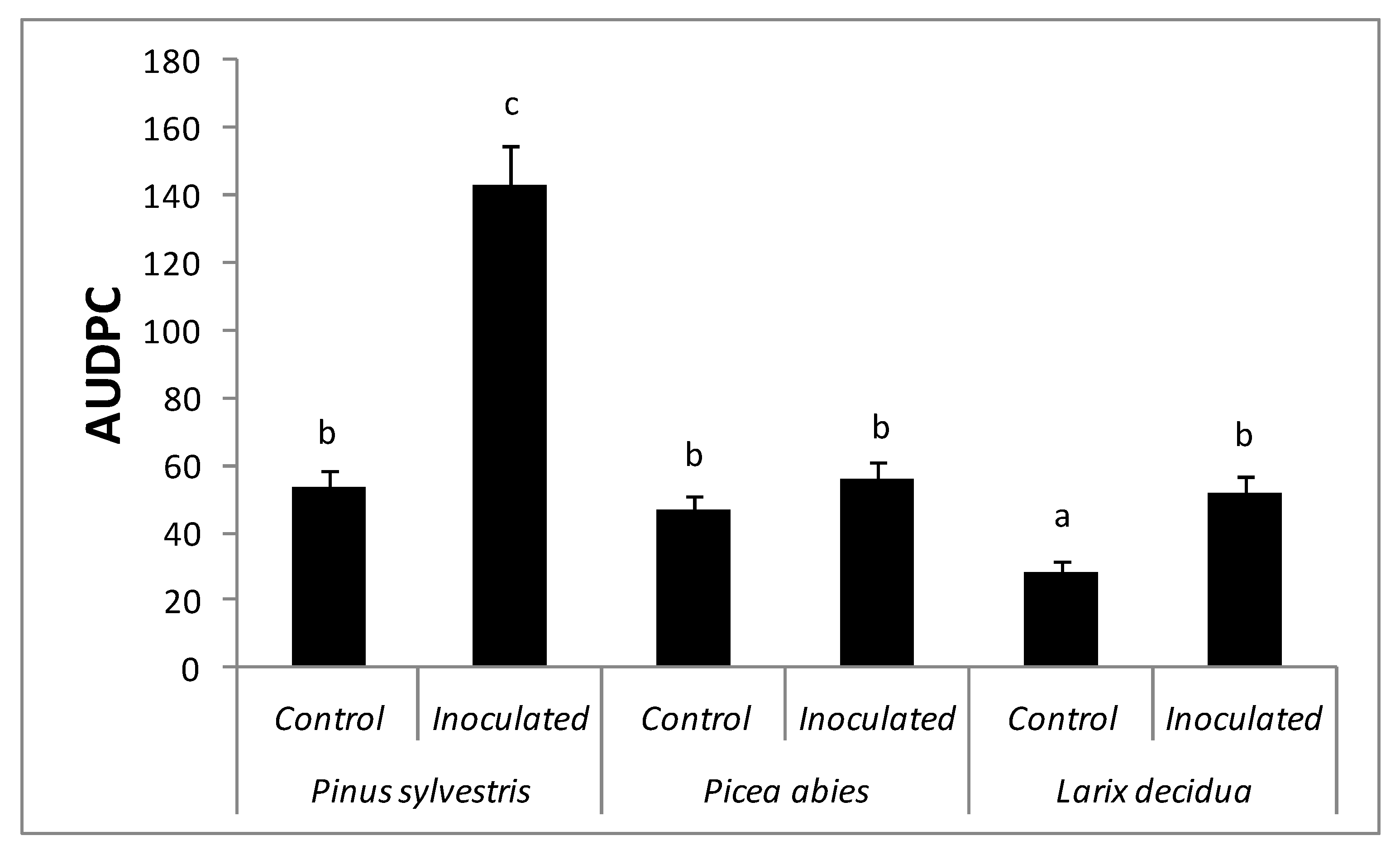
Symptomatology varied according to the treatment (Figure 2). The ANOVA revealed significant differences in the values of AUDPC caused by the F. circinatum inoculations. In fact, a significant interaction between species (P. sylvestris, P. abies, and L. decidua) and treatment (inoculated vs. control) was observed (F = 29.08, p < 0.001) (Figure 3). Inoculated P. sylvestris seedlings showed the highest AUDPC values, confirming its susceptibility to F. circinatum. The values of AUDPC also increased in Larix decidua as a result of the F. circinatum inoculation (Figure 3). This difference seems to be only due to the presence of a higher number of seedlings showing resinosis at the point of inoculation in the inoculated seedlings than the controls (Figure 2). Inoculated P. abies seedlings showed resinosis even beyond the inoculation point (Figure 2), but no significant differences were found in the AUDPC values (Figure 3).
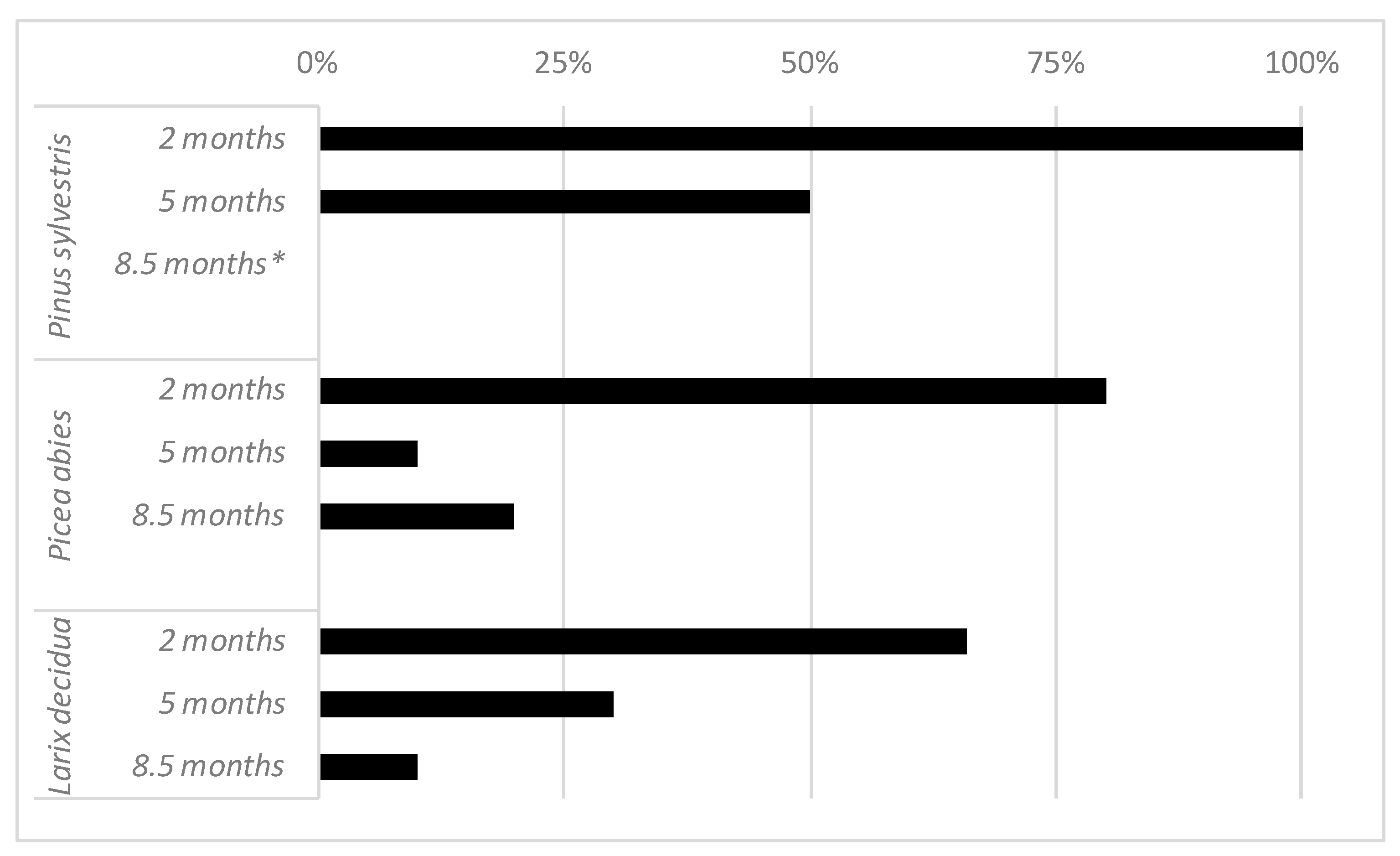
Although the ratio of reisolation of F. circinatum declined over time, the presence of asymptomatic yet infected seedlings was confirmed in the three species up to 8.5 months after inoculation (Figure 4). Fusarium circinatum was reisolated beyond the inoculation point, both upward and downward.
4. Discussion
To our knowledge, this is the first study testing the susceptibility of conifers from the Czech Republic to F. circinatum. The present study demonstrated the pathogenicity of F. circinatum to the Czech provenance of P. sylvestris, which is in concordance with previous studies carried out with Spanish provenances [28,29,30,61]. Inoculated seedlings started dying 37 days after inoculation, although approximately half of the seedlings survived up to 5 months after inoculation. Timing of mortality as a result of F. circinatum inoculations varies according to the host. In fact, the mortality of P. radiata seedlings, the most susceptible species to F. circinatum [25], is commonly 7 days after inoculation [62]. However, the study findings are in contrast to the results obtained by Pérez-Sierra et al. [61] and Mullett et al. [63], who found that all or the majority (>98%) of inoculated P. sylvestris seedlings died 56 and 40 days after inoculation, respectively. This discrepancy could be due to the age of the seedlings. The present study used 1.5-year-old seedlings, whereas the seedlings used by Pérez-Sierra et al. [61] and Mullett et al. [63] were 7–8-month-old and one-year-old seedlings, respectively. So, this seems to point out that P. sylvestris might acquire age-related resistance, which had already been noted by Martín-García et al. [50].
Nevertheless, taking into account that intraspecific genotypic variation has been recognized as a source of resistance to F. circinatum in other pine species [33,35,45,64,65,66], the genetic effect should not be dismissed. In fact, Martín-García et al. [50] found that Romanian provenance of P. sylvestris was not susceptible to F. circinatum and this resistance was related to the high genetic variability of European Scots pine populations [67,68].
In forests, F. circinatum has been traditionally restricted to Pinus species and P. menziesii [21,25,69]. Nevertheless, damping off caused by F. circinatum has been also reported in other conifers. Post-emergence damping-off, but not pre-emergence, has been reported in P. abies [28,50]. On the other hand, Martínez-Álvarez et al. [28], in a field trial, did not find differences in the AUDPC values for 3-year-old P. abies seedlings between inoculated and control plots, which is consistent with the present study. The discrepancy between experiments testing the susceptibility of P. abies might be also due to age-related resistances. In fact, Martínez-Álvarez et al. [28] already pointed out that the differences between the results obtained from laboratory and field trials (including other conifers, not only P. abies) may be a result of the age of the seedlings. The resinosis beyond the point of inoculation found in ca. 25% of the inoculated seedlings confirms that F. circinatum is able to infect P. abies seedlings. However, the fact that no mortality was recorded 8.5 months after inoculation seems to indicate that P. abies seedlings with lignified vascular tissues are able to tolerate the F. circinatum infection.
The AUDPC values for L. decidua were significantly different between inoculated and control seedlings (Figure 3) because resinosis at the point of inoculation was more frequent in inoculated seedlings than controls (Figure 2). However, resinosis beyond the inoculation point or wilting was not observed. This, together with the fact that pre-emergence and post-emergence damping off in L. decidua has been demonstrated [28], seems to also indicate age-related resistances in this conifer.
The reisolation of F. circinatum beyond the inoculation point demonstrated that this quarantine pathogen is able to infect the three conifers tested (P. sylvestris, P. abies, and L. decidua), giving rise to asymptomatic yet infected seedlings for at least eight and a half months. This finding is consistent with results obtained by Elvira-Recuenco et al. [46] and Swett et al. [47] who found symptomless P. radiata seedlings for 475 days after inoculation or at least 52 weeks, respectively. Other studies also demonstrated the presence of asymptomatic seedlings, but all of them were conducted on Pinus species [42,43,44]. To our knowledge, this is the first study in which the presence of asymptomatic yet infected seedlings has been recorded in the Picea and Larix genera.
5. Conclusions
The present study demonstrated the susceptibility of the Czech provenance of P. sylvestris to F. circinatum, which confirms the importance of testing the susceptibility of European conifers both at species and provenance level. Further studies are needed to confirm the susceptibility of mature trees of P. sylvestris in the Czech Republic. However, the fact that P. sylvestris covers ca. 16% of the total forest area in the Czech Republic and that almost 80% of its regeneration is artificial (i.e., by seedlings produced in nurseries) [1] underscores the importance of carrying out surveys to monitor the eventual presence of F. circinatum in nurseries. Europe, and in particular the Czech authorities, should be aware of the high threat posed by this quarantine pathogen not only for the potential serious losses in forests, but also for the forest nurseries sector and the management of public greenery.
This study has also confirmed, for the first time, the presence of asymptomatic yet infected seedlings of P. abies and L. decidua. In 2007, the European Commission adopted emergency measures to prevent the introduction into and the spread within the Community of F. circinatum (Commission Decision 2007/433/EC of 18 June 2007). However, the measures provided within this decision are applied only for plants of the genus Pinus and the species Pseudotsuga menziesii. Therefore, the European legislation fails to control the movement of symptomless seedlings of other genera, such as Picea and Larix. This, together with the fact that these seedlings could remain symptomless for at least eight and a half months, drastically increases the risk of new introductions into disease-free European countries.
Acknowledgments
This work has been supported by grant of the Ministry of education, youth and sports of the Czech Republic LD15046 “Detection and biology of Gibberella circinata—essentials for early warning and management strategies in the Czech Republic”. This article is also based upon work carried out during COST Action FP1406 PINESTRENGTH (Pine pitch canker-strategies for management of Gibberella circinata in greenhouses and forests), supported by COST (European Cooperation in Science and Technology). The Portuguese Foundation for Science and Technology (FCT) supported Jorge Martín-García (Post doc grant-SFRH/BPD/122928/2016). Aneta Lukačevičová was awarded with two Short Term Scientific Missions within the PINESTRENGTH framework to perform the experiment in the Laboratory of Forest Pathology at the University of Valladolid (Spain).
Author Contributions
J.M.-G., A.L., J.A.F.-P., J.J.D., and M.D. conceived and designed the experiment. A.L., J.M.-G., and J.A.F.-P. performed the experiments. J.M.-G. analyzed the data and wrote the paper. All authors reviewed the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ministry of Agriculture of the Czech Republic. Information on Forests and Forestry in the Czech Republic by 2016; Ministry of Agriculture of the Czech Republic: Prague, Czech Republic, 2017.
- Stenlid, J.; Oliva, J.; Boberg, J.B.; Hopkins, A.J.M. Emerging diseases in European forest ecosystems and responses in society. Forests 2011, 2, 486–504. [Google Scholar] [CrossRef]
- Santini, A.; Ghelardini, L.; De Pace, C.; Desprez-Loustau, M.L.; Capretti, P.; Chandelier, A.; Cech, T.; Chira, D.; Diamandis, S.; Gaitniekis, T.; et al. Biogeographical patterns and determinants of invasion by forest pathogens in Europe. New Phytol. 2013, 197, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Ramsfield, T.D.; Bentz, B.J.; Faccoli, M.; Jactel, H.; Brockerhoff, E.G. Forest health in a changing world: Effects of globalization and climate change on forest insect and pathogen impacts. Forestry 2016, 89, 245–252. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization (EPPO). Data sheets on quarantine pests: Gibberella circinata. EPPO Bull. 2005, 35, 383–386. [Google Scholar] [CrossRef]
- Hepting, G.H.; Roth, E.R. Pitch canker, a new disease of some Southern pines. J. For. 1946, 44, 742–744. [Google Scholar]
- Hepting, G.H.; Roth, E.R. Host relations and spread of the pine pitch canker disease. Phytopathology 1953, 43, 475. [Google Scholar]
- Viljoen, A.; Wingfield, M.J.; Marasas, W.F.O. First report of Fusarium subglutinans f. sp. pini on pine seedlings in South Africa. Plant Dis. 1994, 78, 309–312. [Google Scholar] [CrossRef]
- Guerra-Santos, J.J. Pitch canker on Monterey pine in Mexico. In Current and Potential Impacts of Pitch Canker in Radiata Pine, Proceedings of the IMPACT Monterey Workshop, Monterey, CA, USA, 30 November–3 December 1998; Devey, M., Matheson, C., Gordon, T., Eds.; CSIRO Forestry & Forest Products: Canberra, Australia, 1999; Volume 112, pp. 58–61. [Google Scholar]
- Wingfield, M.J.; Jacobs, A.; Coutinho, T.A.; Ahumada, R.; Wingfield, B.D. First report of the pitch canker fungus, Fusarium circinatum, on pines in Chile. Plant Pathol. 2002, 51, 397. [Google Scholar] [CrossRef]
- Lee, J.K.; Lee, S.-H.; Yang, S.-I.; Lee, Y.-W. First report of Pitch Canker disease on Pinus rigida in Korea. Plant Pathol. J. 2000, 16, 52–54. [Google Scholar]
- Kobayashi, T.; Muramoto, M. Pitch canker of Pinus luchuensis, a new disease of Japanese forests. For. Pests 1989, 40, 169–173. [Google Scholar]
- Alonso, R.; Bettucci, L. First report of the pitch canker fungus Fusarium circinatum affecting Pinus taeda seedlings in Uruguay. Australas. Plant Dis. Notes 2009, 4, 91–92. [Google Scholar] [CrossRef]
- Steenkamp, E.T.; Rodas, C.A.; Kvas, M.; Wingfield, M.J. Fusarium circinatum and pitch canker of Pinus in Colombia. Australas. Plant Pathol. 2012, 41, 483–491. [Google Scholar] [CrossRef]
- Pfenning, L.H.; da Silva Costa, S.; de Melo, M.P.; Costa, H.; Ventura, J.A.; Auer, C.G.; dos Santos, Á.F. First report and characterization of Fusarium circinatum, the causal agent of pitch canker in Brazil. Trop. Plant Pathol. 2014, 39, 210–216. [Google Scholar] [CrossRef]
- Dwinell, D. Global Distribution of the Pitch Canker Fungus. In Current and Potential Impacts of Pitch Canker in Radiata Pine, Proceedings of the IMPACT Monterey Workshop, Monterey, CA, USA, 30 November–3 December 1998; CSIRO Forestry & Forest Products: Canberra, Australia, 1999; pp. 54–57. [Google Scholar]
- Landeras, E.; García, P.; Fernández, M.; Braña, M.; Pérez-Sierra, A.; León, M.; Abad-Campos, P.; Armengol, J. Outbreak of Pitch Canker Caused by Fusarium circinatum on Pinus spp. in Northern Spain. Plant Dis. 2005, 89, 1015. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization (EPPO). First Report of Gibberella circinata in France; EPPO: Paris, France, 2006; Volume 5. [Google Scholar]
- Carlucci, A.; Colatruglio, L.; Frisullo, S. First Report of Pitch Canker Caused by Fusarium circinatum on Pinus halepensis and P. pinea in Apulia (Southern Italy). Plant Dis. 2007, 91, 1683. [Google Scholar] [CrossRef]
- Bragança, H.; Diogo, E.; Moniz, F.; Amaro, P. First Report of Pitch Canker on Pines Caused by Fusarium circinatum. Plant Dis. 2009, 93, 1079. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Risk assessment of Gibberella circinata for the EU territory and identification and evaluation of risk management options. Eur. Food Saf. Auth. 2010, 8, 1620. [Google Scholar] [CrossRef]
- Watt, M.S.; Ganley, R.J.; Kriticos, D.J.; Manning, L.K. Dothistroma needle blight and pitch canker: The current and future potential distribution of two important diseases of Pinus species. Can. J. For. Res. 2011, 41, 412–424. [Google Scholar] [CrossRef]
- Ganley, R.J.; Watt, M.S.; Manning, L.; Iturritxa, E. A global climatic risk assessment of pitch canker disease. Can. J. For. Res. 2009, 39, 2246–2256. [Google Scholar] [CrossRef]
- Bezos, D.; Martínez-Álvarez, P.; Fernández, M.; Diez, J.J. Epidemiology and Management of Pine Pitch Canker Disease in Europe—A Review. Balt. For. 2017, 23, 279–293. [Google Scholar]
- Wingfield, M.J.; Hammerbacher, A.; Ganley, R.J.; Steenkamp, E.T.; Gordon, T.R.; Wingfield, B.D.; Coutinho, T.A. Pitch canker caused by Fusarium circinatum—A growing threat to pine plantations and forests worldwide. Australas. Plant Pathol. 2008, 37, 319–334. [Google Scholar] [CrossRef]
- Gordon, T.R.; Okamoto, D.; Storer, A.J.; Wood, D.L. Susceptibility of five landscape pines to pitch canker disease, caused by Fusarium subglutinans f. sp. pini. Hortscience 1998, 33, 868–871. [Google Scholar]
- Enebak, B.S.A.; Stanosz, G.R. Responses of conifer species of the Great Lakes region of North America to inoculation with the pitch canker pathogen Fusarium circinatum. For. Pathol. 2003, 33, 333–338. [Google Scholar] [CrossRef]
- Martínez-Álvarez, P.; Pando, V.; Diez, J.J. Alternative species to replace Monterey pine plantations affected by pitch canker caused by Fusarium circinatum in northern Spain. Plant Pathol. 2014, 63, 1086–1094. [Google Scholar] [CrossRef]
- Iturritxa, E.; Mesanza, N.; Elvira-Recuenco, M.; Serrano, Y.; Quintana, E.; Raposo, R. Evaluation of genetic resistance in Pinus to pitch canker in Spain. Australas. Plant Pathol. 2012, 41, 601–607. [Google Scholar] [CrossRef]
- Iturritxa, E.; Ganley, R.J.; Raposo, R.; García-Serna, I.; Mesanza, N.; Kirkpatrick, S.C.; Gordon, T.R. Resistance levels of Spanish conifers against Fusarium circinatum and Diplodia pinea. For. Pathol. 2013, 43, 488–495. [Google Scholar] [CrossRef]
- Gordon, T.R. Pitch canker disease of pines. Phytopathology 2006, 96, 657–659. [Google Scholar] [CrossRef] [PubMed]
- Matheson, A.C.; Devey, M.E.; Gordon, T.L.; Werner, W.; Vogler, D.R.; Balocchi, C.; Carson, M.J. Heritability of response to inoculation by pine pitch canker of seedlings of radiata pine. Aust. For. 2006, 69, 101–106. [Google Scholar] [CrossRef]
- Hodge, G.R.; Dvorak, W.S. Variation in pitch canker resistance among provenances of Pinus patula and Pinus tecunumanii from Mexico and Central America. New For. 2007, 33, 193–206. [Google Scholar] [CrossRef]
- Mitchell, R.; Wingfield, M.; Steenkamp, E.; Coutinho, T. Tolerance of Pinus patula full-sib families to Fusarium circinatum in a greenhouse study. South. For. J. For. Sci. 2012, 74, 247–252. [Google Scholar] [CrossRef]
- Elvira-Recuenco, M.; Iturritxa, E.; Majada, J.; Alia, R.; Raposo, R. Adaptive potential of maritime pine (Pinus pinaster) populations to the emerging pitch canker pathogen, Fusarium circinatum. PLoS ONE 2014, 9, e114971. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, K.; Cavers, S.; Cottrell, J.E.; Ennos, R.A. Genetic variation for needle traits in Scots pine (Pinus sylvestris L.). Tree Genet. Genomes 2016, 12, 40. [Google Scholar] [CrossRef]
- Belletti, P.; Ferrazzini, D.; Piotti, A.; Monteleone, I.; Ducci, F. Genetic variation and divergence in Scots pine (Pinus sylvestris L.) within its natural range in Italy. Eur. J. For. Res. 2012, 131, 1127–1138. [Google Scholar] [CrossRef]
- Wójkiewicz, B.; Cavers, S.; Wachowiak, W. Current Approaches and Perspectives in Population Genetics of Scots Pine (Pinus sylvestris L.). For. Sci. 2016, 62, 343–354. [Google Scholar] [CrossRef]
- Lagercrantz, U.; Ryman, N. Genetic Structure of Norway Spruce (Picea abies): Concordance of Morphological and Allozymic Variation. Evolution 1990, 44, 38–53. [Google Scholar] [PubMed]
- Maier, J. Genetic variation in European larch (Larix decidua Mill). Ann. For. Sci. 1992, 49, 39–47. [Google Scholar] [CrossRef]
- Lewandowski, A.; Mejnartowicz, L. Levels and patterns of allozyme variation in some European larch (Larix decidua) populations. Hereditas 1991, 115, 221–226. [Google Scholar] [CrossRef]
- Storer, A.J.; Gordon, T.R.; Clark, S.L. Association of the pitch canker fungus, Fusarium subglutinans f. sp. pini, with Monterey pine seeds and seedlings in California. Plant Pathol. 1998, 47, 649–656. [Google Scholar]
- Mitchell, R.G.; Zwolinski, J.; Jones, N.; Coutinho, T. The effect of applying prophylactic measures on the post-planting survival of Pinus patula in South Africa. S. Afr. For. J. 2004, 200, 51–58. [Google Scholar]
- Kim, Y.-S.; Woo, K.-S.; Koo, Y.-B.; Yeo, J.-K. Variation in susceptibility of six pine species and hybrids to pitch canker caused by Fusarium circinatum. For. Pathol. 2008, 38, 419–428. [Google Scholar] [CrossRef]
- Vivas, M.; Zas, R.; Solla, A. Screening of Maritime pine (Pinus pinaster) for resistance to Fusarium circinatum, the causal agent of Pitch Canker disease. Forestry 2012, 85, 185–192. [Google Scholar] [CrossRef]
- Elvira-Recuenco, M.; Iturritxa, E.; Raposo, R. Impact of seed transmission on the infection and development of pitch canker disease in Pinus radiata. Forests 2015, 6, 3353–3368. [Google Scholar] [CrossRef]
- Swett, C.L.; Kirkpatrick, S.C.; Gordon, T.R. Evidence for a Hemibiotrophic Association of the Pitch Canker Pathogen Fusarium circinatum with Pinus radiata. Plant Dis. 2016, 100, 79–84. [Google Scholar] [CrossRef]
- Martínez-Alvarez, P.; Alves-Santos, F.M.; Diez, J.J. In Vitro and In Vivo Interactions between Trichoderma viride and Fusarium circinatum. Silva Fenn. 2012, 46, 303–316. [Google Scholar] [CrossRef]
- Cerqueira, A.; Alves, A.; Berenguer, H.; Correia, B.; Gómez-Cadenas, A.; Diez, J.J.; Monteiro, P.; Pinto, G. Phosphite shifts physiological and hormonal profile of Monterey pine and delays Fusarium circinatum progression. Plant Physiol. Biochem. 2017, 114, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Martín-García, J.; Paraschiv, M.; Flores-Pacheco, J.A.; Chira, D.; Diez, J.J.; Fernández, M. Susceptibility of several northeastern conifers to Fusarium circinatum and strategies for biocontrol. Forests 2017, 8, 318. [Google Scholar] [CrossRef]
- Plíva, A. Typologický Klasifikační Systém ÚHÚL; ÚHÚL (Ústav pro hospodářskou úpravu lesů): Brandýs nad Labem, Czech Republic, 1987. [Google Scholar]
- Martínez-Álvarez, P.; Fernández-González, R.A.; Sanz-Ros, A.V.; Pando, V.; Diez, J.J. Two fungal endophytes reduce the severity of pitch canker disease in Pinus radiata seedlings. Biol. Control 2016, 94, 1–10. [Google Scholar] [CrossRef]
- Correll, J.C.; Gordon, T.R.; Mccain, A.H.; Fox, J.W.; Koehler, C.S.; Wood, D.L.; Schultz, M.E. Pitch canker disease in California: Pathogenicity, distribution, and canker development on Monterey pine (Pinus radiata). Plant Dis. 1991, 75, 676–682. [Google Scholar] [CrossRef]
- Martínez-Álvarez, P. Environmentally Friendly Methods for the Integrated Management of Pine Pitch Canker (PPC) Disease. Ph.D. Thesis, University of Valladolid, Palencia, Spain, 2015. [Google Scholar]
- Leslie, J.; Summerell, B. The Fusarium Laboratory Manual; Blackwell Publishing Professional: Ames, IA, USA, 2006. [Google Scholar]
- Prieto-Recio, C.; Martín-García, J.; Diez, J.J. Pathogenicity of Spanish isolates of Heterobasidion annosum s. s. in Pinus pinaster seedlings. For. Pathol. 2014, 44, 163–165. [Google Scholar] [CrossRef]
- Zamora-Ballesteros, C.; Haque, M.M.U.; Diez, J.J.; Martín-García, J. Pathogenicity of Phytophthora alni complex and P. plurivora in Alnus glutinosa seedlings. For. Pathol. 2016. [Google Scholar] [CrossRef]
- García-Pérez, A. Métodos Avanzados de Estadística Aplicada. Métodos Robustos y de Remuestreo; García-Pérez, A., Ed.; UNED Universidad Nacional a Distancia: Madrid, Spain, 2010. [Google Scholar]
- Kaplan, E.L.; Meier, P. Nonparametric estimation from incomplete observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Therneau, T. A Package for Survival Analysis in S. R Package Version 2.41-3. Available online: https://CRAN.R-project.org/package=survival (accessed on 18 December 2017).
- Pérez-Sierra, A.; Landeras, E.; León, M.; Berbegal, M.; García-Jiménez, J.; Armengol, J. Characterization of Fusarium circinatum from Pinus spp. in northern Spain. Mycol. Res. 2007, 111, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Adalia, E.J.; Flores-Pacheco, J.A.; Martínez-Álvarez, P.; Martín-García, J.; Fernández, M.; Diez, J.J. Effect of mycoviruses on the virulence of Fusarium circinatum and laccase activity. Physiol. Mol. Plant Pathol. 2016, 94, 8–15. [Google Scholar] [CrossRef]
- Mullett, M.; Pérez-Sierra, A.; Armengol, J.; Berbegal, M. Phenotypical and Molecular Characterisation of Fusarium circinatum: Correlation with Virulence and Fungicide Sensitivity. Forests 2017, 8, 458. [Google Scholar] [CrossRef]
- Dvorak, W.S.; Hodge, G.R.; Kietzka, J.E. Genetic variation in survival, growth, and stem form of Pinus leiophylla in Brazil and South Africa and provenance resistance to pitch canker. South. Hemisph. For. J. 2007, 69, 125–135. [Google Scholar] [CrossRef]
- Dvorak, W.S.; Potter, K.M.; Hipkins, V.D.; Hodge, G.R. Genetic Diversity and Gene Exchange in Pinus oocarpa, a Mesoamerican Pine with Resistance to the Pitch Canker Fungus (Fusarium circinatum). Int. J. Plant Sci. 2009, 170, 609–626. [Google Scholar] [CrossRef]
- Mitchell, R.G.; Wingfield, M.J.; Hodge, G.R.; Steenkamp, E.T.; Coutinho, T.A. Selection of Pinus spp. in South Africa for tolerance to infection by the pitch canker fungus. New For. 2012, 43, 473–489. [Google Scholar] [CrossRef]
- Cheddadi, R.; Vendramin, G.G.; Litt, T.; Francois, L.; Kageyama, M.; Lorentz, S.; Laurent, J.; De Beaulieu, J.; Sadori, L.; Jost, A.; et al. Imprints of glacial refugia in the modern genetic diversity of Pinus sylvestris. Glob. Ecol. Biogeogr. 2006, 15, 271–282. [Google Scholar] [CrossRef]
- Prus-Glowacki, W.; Stephan, B.R. Genetic Variation of Pinus sylvestris from Spain in Relation to Other European Populations. Silva Fenn. 1994, 43, 7–14. [Google Scholar]
- CAB International (The Centre for Agriculture and Bioscience International). Gibberella circinata (Pitch Canker)—Pinus; Crop Protection Compendium; CAB International: Wallingford, UK, 2007. [Google Scholar]
Figure 1.
Plot of survival probability determined using the Kaplan-Meier estimate of the survival function for Pinus sylvestris, Picea abies, and Larix decidua seedlings infected with Fusarium circinatum. Note: no mortality was recorded for inoculated seedlings of P. abies and L. decidua and all control seedlings. For this reason, all of these curves overlap in a straight line making it difficult to distinguish them.
Figure 1.
Plot of survival probability determined using the Kaplan-Meier estimate of the survival function for Pinus sylvestris, Picea abies, and Larix decidua seedlings infected with Fusarium circinatum. Note: no mortality was recorded for inoculated seedlings of P. abies and L. decidua and all control seedlings. For this reason, all of these curves overlap in a straight line making it difficult to distinguish them.

Figure 2.
Type of symptomatology showed by Pinus sylvestris, Picea abies, and Larix decidua seedlings at the end of the experiment following the Correll et al. [53] scale, where 0 = healthy plant, 1 = resin and/or necrosis at the point of inoculation and healthy foliage, 2 = resin and/or necrosis beyond the point of inoculation, 3 = accentuated wilting and appreciable dieback, 4 = dead plant.
Figure 2.
Type of symptomatology showed by Pinus sylvestris, Picea abies, and Larix decidua seedlings at the end of the experiment following the Correll et al. [53] scale, where 0 = healthy plant, 1 = resin and/or necrosis at the point of inoculation and healthy foliage, 2 = resin and/or necrosis beyond the point of inoculation, 3 = accentuated wilting and appreciable dieback, 4 = dead plant.
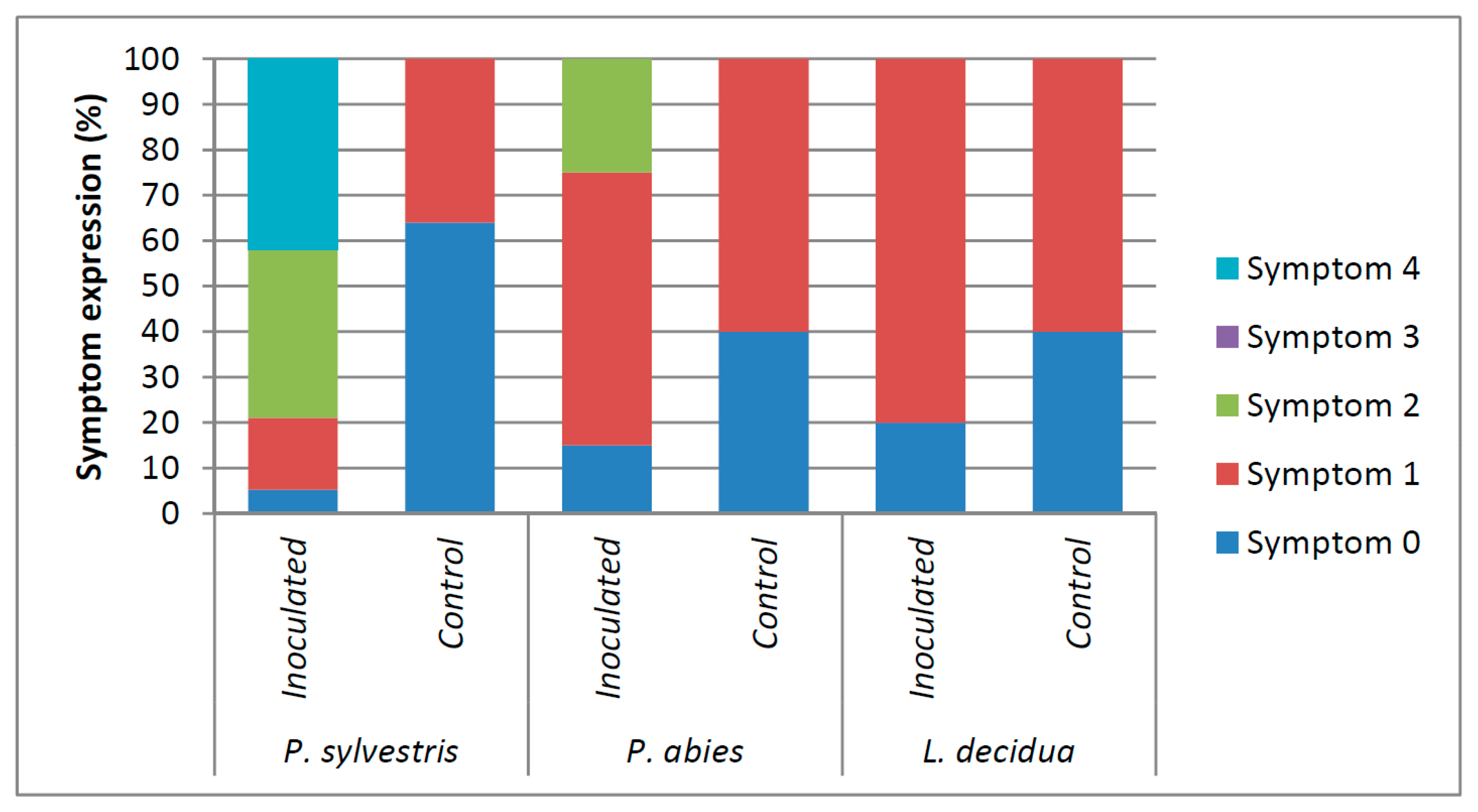
Figure 3.
Comparison of area under the disease progress curve (AUDPC) for Pinus sylvestris, Picea abies, and Larix decidua seedlings. Different letters above bars indicate significantly different means (Generalized Welch procedure 0.1 trimmed means, p < 0.05).
Figure 3.
Comparison of area under the disease progress curve (AUDPC) for Pinus sylvestris, Picea abies, and Larix decidua seedlings. Different letters above bars indicate significantly different means (Generalized Welch procedure 0.1 trimmed means, p < 0.05).
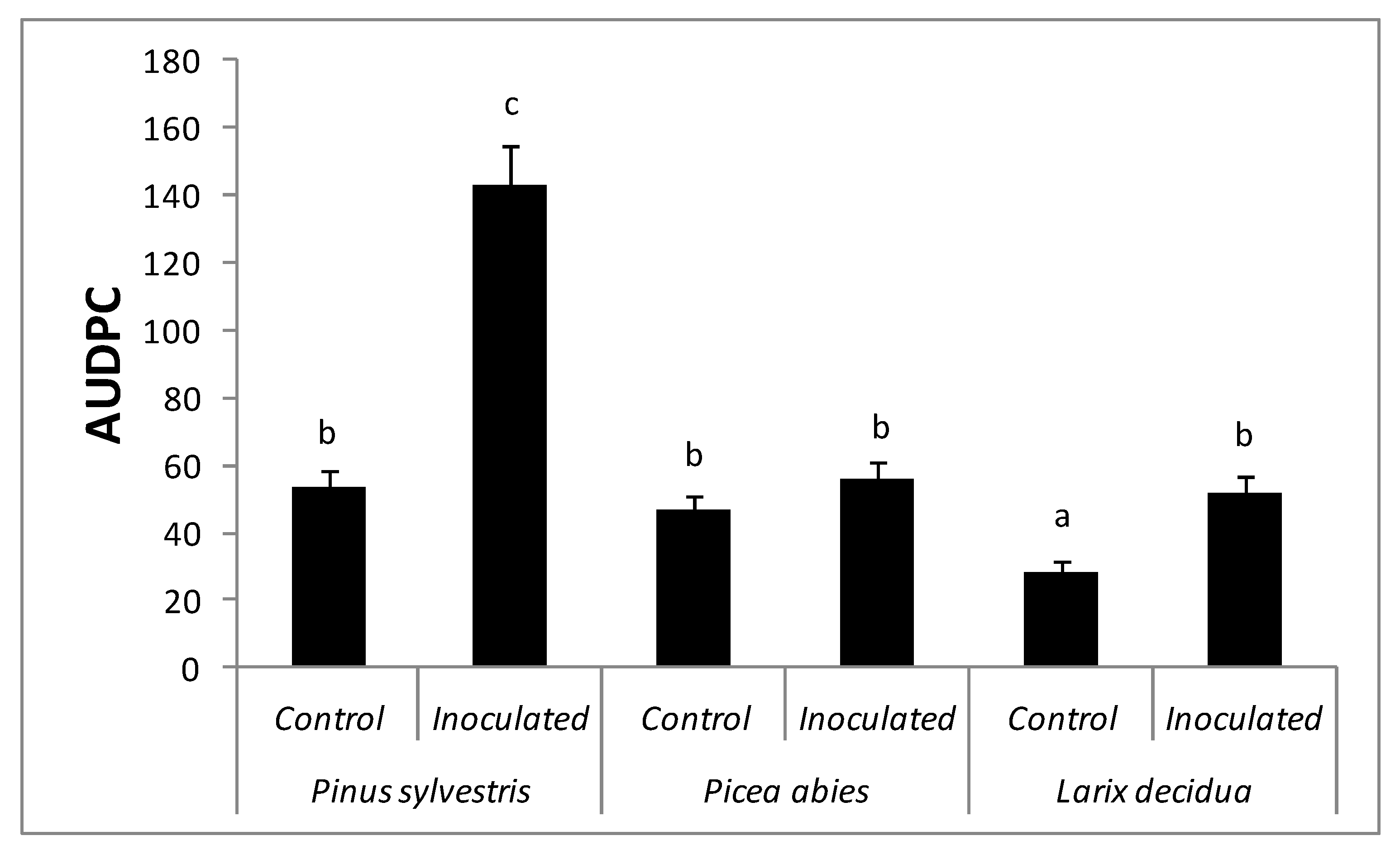
Figure 4.
Ratio of positive reisolations of Fusarium circinatum from inoculated seedlings at 2, 5, and 8.5 months after inoculation. (*) All the seedlings were already dead 8.5 months after inoculation and reisolation could not be carried out.
Figure 4.
Ratio of positive reisolations of Fusarium circinatum from inoculated seedlings at 2, 5, and 8.5 months after inoculation. (*) All the seedlings were already dead 8.5 months after inoculation and reisolation could not be carried out.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Martín-García, J.; Lukačevičová, A.; Flores-Pacheco, J.A.; Diez, J.J.; Dvořák, M. Evaluation of the Susceptibility of Several Czech Conifer Provenances to Fusarium circinatum. Forests 2018, 9, 72. https://doi.org/10.3390/f9020072
AMA Style
Martín-García J, Lukačevičová A, Flores-Pacheco JA, Diez JJ, Dvořák M. Evaluation of the Susceptibility of Several Czech Conifer Provenances to Fusarium circinatum. Forests. 2018; 9(2):72. https://doi.org/10.3390/f9020072
Chicago/Turabian StyleMartín-García, Jorge, Aneta Lukačevičová, Juan Asdrúbal Flores-Pacheco, Julio Javier Diez, and Miloň Dvořák. 2018. "Evaluation of the Susceptibility of Several Czech Conifer Provenances to Fusarium circinatum" Forests 9, no. 2: 72. https://doi.org/10.3390/f9020072
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.