Fungal Frequency and Mite Load Trends Interact with a Declining Mountain Pine Beetle Population

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. The Sampling and Identification of the Studied Organism
2.3. Molecular Identification of Fungi
2.4. Statistical Analysis
3. Results
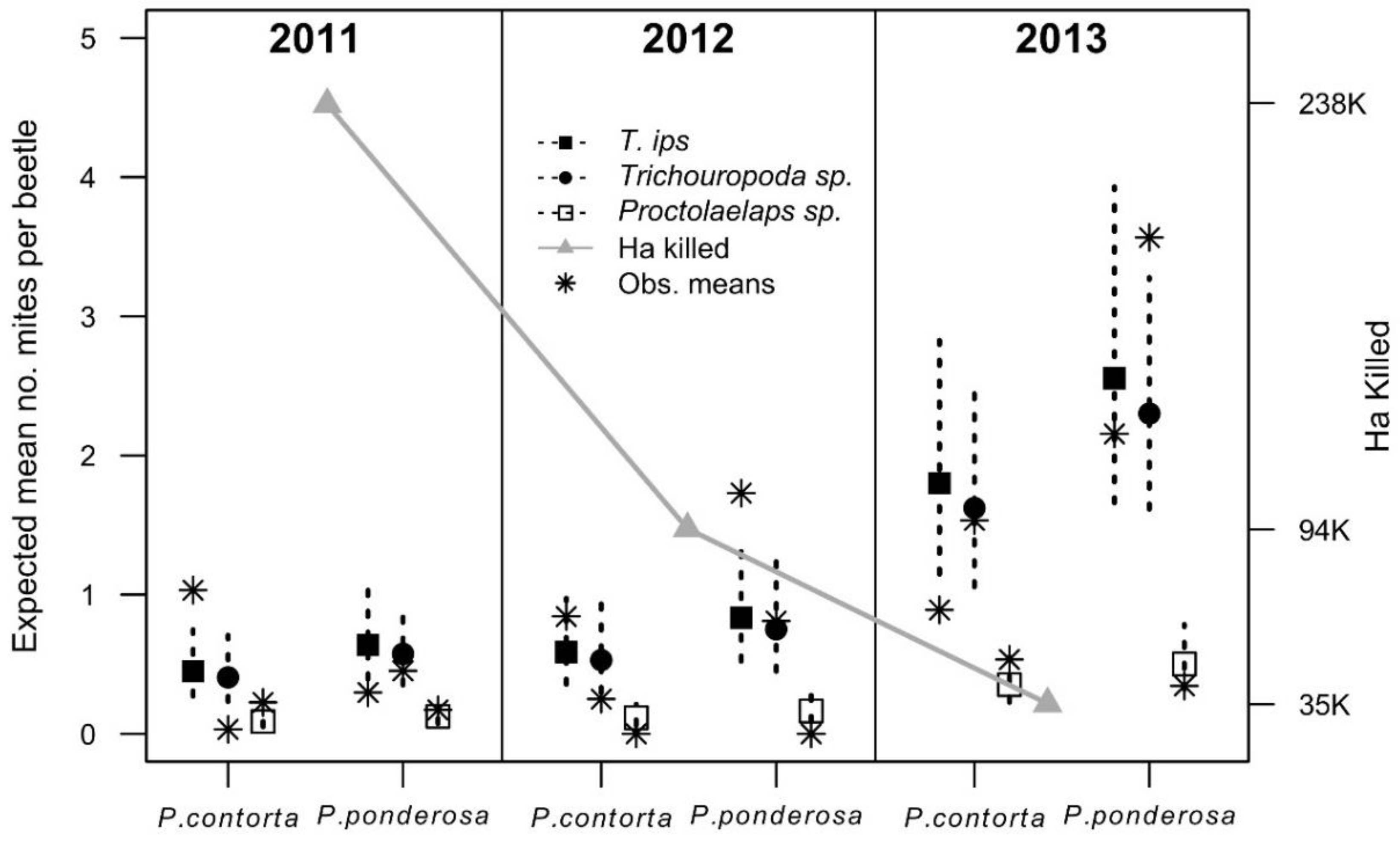
3.1. Mite Trends
3.1.1. Phoretic Mite Diversity and Yearly Trends
3.1.2. Influence of Attacked Pine Host on Mite Trends
3.1.3. Influence of Beetle Sex on Mite Trends
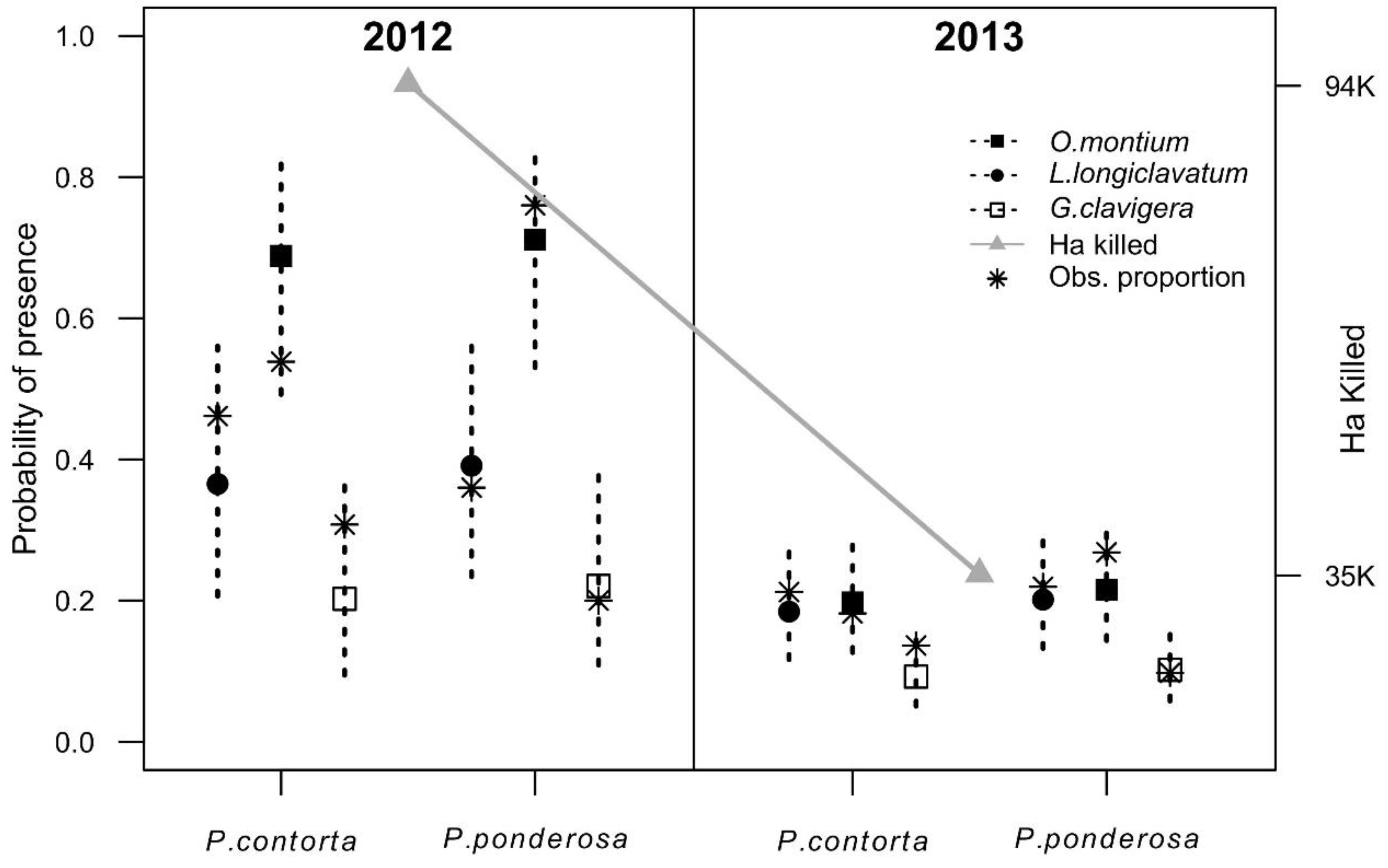
3.2. Trends of Transported Fungi
3.2.1. Fungal Transport Trends in Different Beetle Structures
3.2.2. The Year and Host Effects on Fungal Trends
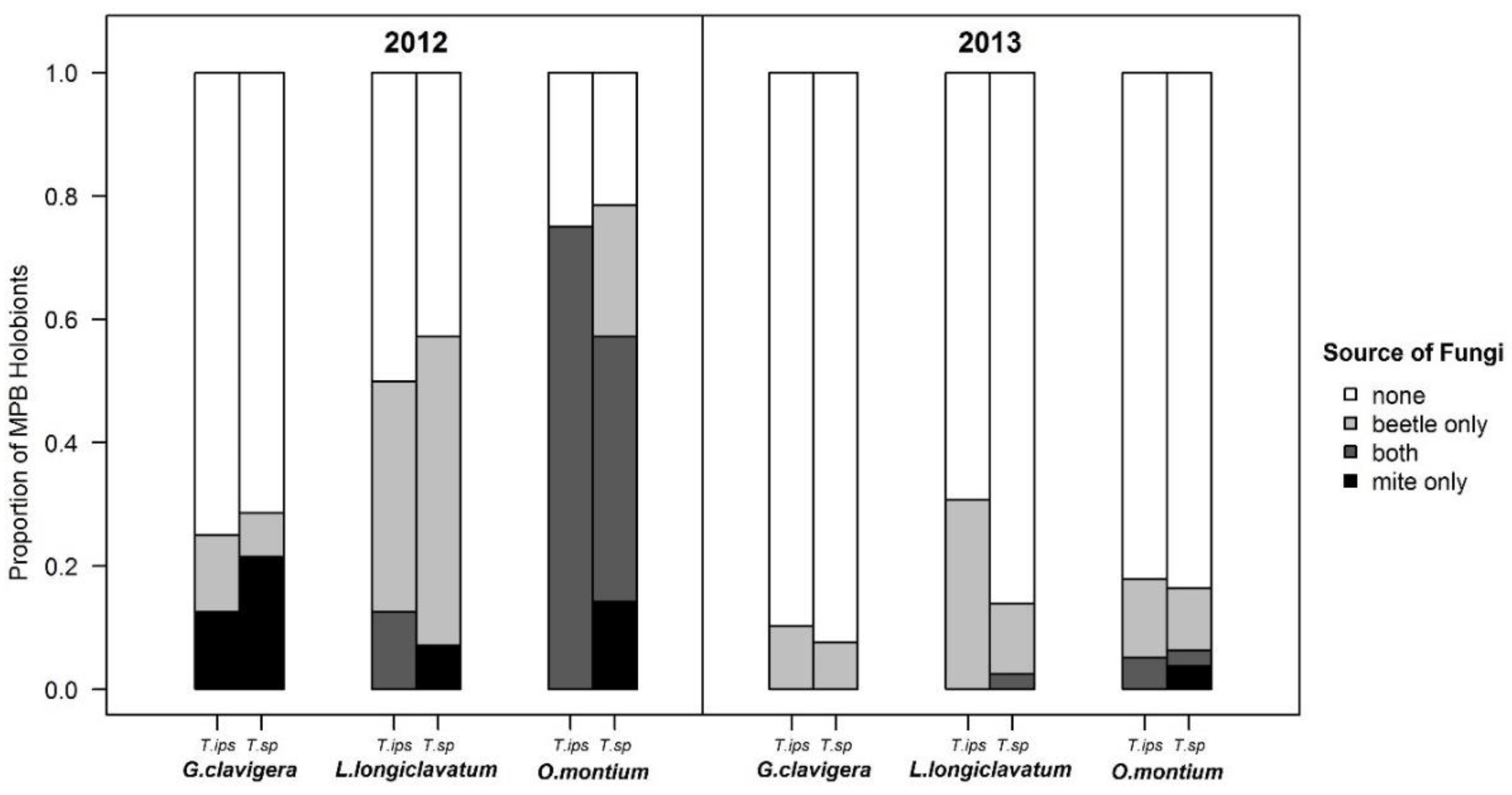
3.2.3. Fungal Trends Observed in Phoretic Mites
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jarvis, D.S.; Kulakowski, D. Long-term history and synchrony of mountain pine beetle outbreaks in lodgepole pine forests. J. Biogeogr. 2015, 42, 1029–1039. [Google Scholar] [CrossRef]
- Franklin, J.F.; Shugart, H.H.; Harmon, M.E. Tree death as an ecological process. Bioscience 1987, 37, 550–556. [Google Scholar] [CrossRef]
- Ballard, R.G.; Walsh, M.A.; Cole, W.E. Blue-stain fungi in xylem of lodgepole pine—A light-microscope study on extent of hyphal distribution. Can. J. Bot. Rev. Can. Bot. 1982, 60, 2334–2341. [Google Scholar] [CrossRef]
- Ballard, R.G.; Walsh, M.A.; Cole, W.E. The penetration and growth of blue-stain fungi in the sapwood of lodgepole pine attacked by mountain pine-beetle. Can. J. Bot. Rev. Can. Bot. 1984, 62, 1724–1729. [Google Scholar] [CrossRef]
- Hubbard, R.M.; Rhoades, C.C.; Elder, K.; Negron, J. Changes in transpiration and foliage growth in lodgepole pine trees following mountain pine beetle attack and mechanical girdling. For. Ecol. Manag. 2013, 289, 312–317. [Google Scholar] [CrossRef]
- Solheim, H. Early stages of blue-stain fungus invasion of lodgepole pine sapwood following mountain pine-beetle attack. Can. J. Bot. Rev. Can. Bot. 1995, 73, 70–74. [Google Scholar] [CrossRef]
- Callaham, R.Z.; Shifrine, M. The yeasts associated with bark beetles. For. Sci. 1960, 6, 146–154. Available online: https://www.fs.fed.us/psw/publications/callaham/psw_1960_callaham001.pdf (accessed on 10 December 2017).
- Davis, T.S. The ecology of yeasts in the bark beetle holobiont: A century of research revisited. Microb. Ecol. 2015, 69, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Six, D.L. The bark beetle holobiont: Why microbes matter. J. Chem. Ecol. 2013, 39, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Bleiker, K.P.; Potter, S.E.; Lauzon, C.R.; Six, D.L. Transport of fungal symbionts by mountain pine beetles. Can. Entomol. 2009, 141, 503–514. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J.J.; Breuil, C. Diversity of fungi associated with the mountain pine beetle, Dendroctonus ponderosae and infested lodgepole pines in British Columbia. Fungal Divers. 2006, 22, 91–105. [Google Scholar] [CrossRef]
- Whitney, H.; Farris, S. Maxillary mycangium in the mountain pine beetle. Science 1970, 167, 54–55. [Google Scholar] [CrossRef] [PubMed]
- Hunt, D.W.A.; Borden, J.H. Conversion of verbenols to verbenone by yeasts isolated from Dendroctonus-ponderosae (Coleoptera, Scolytidae). J. Chem.l Ecol. 1990, 16, 1385–1397. [Google Scholar] [CrossRef] [PubMed]
- Farmer, L.J. Phloem-Yeast Complex during Infestations of the Mountain Pine Beetle in Lodgepole Pine; Available through Interlibrary Loan (Received on 13 December 2017); FAO: Rome, Italy, 1965. [Google Scholar]
- Whitney, H.S. Association of Dendroctonus-ponderosae (Coleoptera-Scolytidae) with blue stain fungi and yeasts during brood development in lodgepole pine. Can. Entomol. 1971, 103, 1495–1503. [Google Scholar] [CrossRef]
- Six, D.L. A comparison of mycangial and phoretic fungi of individual mountain pine beetles. Can. J. For. Res. Rev. Can. Rech. For. 2003, 33, 1331–1334. [Google Scholar] [CrossRef]
- Six, D.L.; Bentz, B.J. Temperature determines symbiont abundance in a multipartite bark beetle-fungus ectosymbiosis. Microb. Ecol. 2007, 54, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Reid, R.W.; Whitney, H.S.; Watson, J.A. Reactions of lodgepole pine to attack by Dendroctonus ponderosae Hopkins and blue stain fungi. Can. J. Bot. 1967, 45, 1115–1126. [Google Scholar] [CrossRef]
- Myrholm, C.L.; Langor, D.W. Assessment of the impact of symbiont Ophiostomatales (Fungi) on mountain pine beetle (Coleoptera: Curculionidae) performance on a jack pine (Pinaceae) diet using a novel in vitro rearing method. Can. Entomol. 2016, 148, 68–82. [Google Scholar] [CrossRef]
- Six, D.L.; Paine, T.D. Effects of mycangial fungi and host tree species on progeny survival and emergence of Dendroctonus ponderosae (Coleoptera: Scolytidae). Environ. Entomol. 1998, 27, 1393–1401. [Google Scholar] [CrossRef]
- Roe, A.D.; James, P.M.A.; Rice, A.V.; Cooke, J.E.K.; Sperling, F.A.H. Spatial community structure of mountain pine beetle fungal symbionts across a latitudinal gradient. Microb. Ecol. 2011, 62, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lim, L.; Madilao, L.; Lah, L.; Bohlmann, J.; Breuil, C. Gene discovery for enzymes involved in limonene modification or utilization by the mountain pine beetle-associated pathogen Grosmannia clavigera. Appl. Environ. Microbiol. 2014, 80, 4566–4576. [Google Scholar] [CrossRef] [PubMed]
- Coyne, J.; Lott, L. Toxicity of substances in pine oleoresin to southern pine beetles [Dendroctonus frontalis, insect pests]. J. Georg. Entomol. Soc. 1976, 11, 301–305. [Google Scholar] [CrossRef]
- Raffa, K.F.; Berryman, A.A. Physiological differences between lodgepole pines resistant and susceptible to the mountain pine-beetle (Coleoptera, Scolytidae) and associated microorganisms. Environ. Entomol. 1982, 11, 486–492. [Google Scholar] [CrossRef]
- Smith, R.H. Formula for describing effect of insect and host tree factors on resistance to western pine beetle attack. J. Econ. Entomol. 1975, 68, 841–844. [Google Scholar] [CrossRef]
- Alamouti, S.M.; Wang, V.; DiGuistini, S.; Six, D.L.; Bohlmann, J.; Hamelin, R.C.; Feau, N.; Breuil, C. Gene genealogies reveal cryptic species and host preferences for the pine fungal pathogen Grosmannia clavigera. Mol. Ecol. 2011, 20, 2581–2602. [Google Scholar] [CrossRef] [PubMed]
- Khadempour, L.; LeMay, V.; Jack, D.; Bohlmann, J.; Breuil, C. The relative abundance of mountain pine beetle fungal associates through the beetle life cycle in pine trees. Microb. Ecol. 2012, 64, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Allen, E.A.; Humble, L.M.; Breuil, C. Ophiostomatoid and basidiomycetous fungi associated with green, red, and grey lodgepole pines after mountain pine beetle (Dendroctonus ponderosae) infestation. Can. J. For. Res. Rev. Can. Rech. For. 2005, 35, 274–284. [Google Scholar] [CrossRef]
- Moore, M.L.; Six, D.L. Effects of temperature on growth, sporulation, and competition of mountain pine beetle fungal symbionts. Microb. Ecol. 2015, 70, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.S.; Six, D.L. Temporal variation in mycophagy and prevalence of fungi associated with developmental stages of Dendroctonus ponderosae (Coleoptera: Curculionidae). Environ. Entomol. 2007, 36, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Addison, A.L.; Powell, J.A.; Six, D.L.; Moore, M.; Bentz, B.J. The role of temperature variability in stabilizing the mountain pine beetle-fungus mutualism. J. Theor. Biol. 2013, 335, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Hofstetter, R.W.; Dinkins-Bookwalter, J.; Davis, T.S.; Klepzig, K.D. Symbiotic associations of bark beetles. In Bark Beetles; Elsevier: New York, NY, USA, 2015; pp. 209–245. ISBN 9780124171565. [Google Scholar]
- Bridges, J.R.; Moser, J.C. Role of two phoretic mites in transmission of bluestain fungus, Ceratocystis minor. Ecol. Entomol. 1983, 8, 9–12. [Google Scholar] [CrossRef]
- Levieux, J.; Lieutier, F.; Moser, J.C.; Perry, T.J. Transportation of phytopathogenic fungi by the bark beetle Ips sexdentatus Boerner and associated mites. J. Appl. Entomol. Z. Angew. Entomol. 1989, 108, 1–11. [Google Scholar] [CrossRef]
- Moser, J.C.; Konrad, H.; Blomquist, S.R.; Kirisits, T. Do mites phoretic on elm bark beetles contribute to the transmission of Dutch elm disease? Naturwissenschaften 2010, 97, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Moser, J.C.; Perry, T.J.; Solheim, H. Ascospores hyperphoretic on mites associated with Ips typographus. Mycol. Res. 1989, 93, 513–517. [Google Scholar] [CrossRef]
- Klepzig, K.; Moser, J.; Lombardero, M.; Ayres, M.; Hofstetter, R.; Walkinshaw, C. Chapter 13, Mutualism and Antagonism: Ecological Interactions Among Bark Beetles, Mites and Fungi. In Biotic Interactions in Plant-Pathogen Associations; Jeger, M., Ed.; Wye College, University of London: Ashford, UK, 2001; pp. 237–267, ISBN-10: 0851995128. [Google Scholar]
- Hofstetter, R.W.; Klepzig, K.D.; Moser, J.C.; Ayres, M.P. Seasonal dynamics of mites and fungi and their interaction with Southern Pine Beetle. Environ. Entomol. 2006, 35, 22–30. [Google Scholar] [CrossRef]
- Ojeda Alayon, D.I.; Tsui, C.K.; Feau, N.; Capron, A.; Dhillon, B.; Zhang, Y.; Massoumi Alamouti, S.; Boone, C.K.; Carroll, A.L.; Cooke, J.E. Genetic and genomic evidence of niche partitioning and adaptive radiation in mountain pine beetle fungal symbionts. Mol. Ecol. 2017, 26, 2077–2091. [Google Scholar] [CrossRef] [PubMed]
- Mercado, J.E.; Ortiz-Santana, B. Mountain pine beetle mutualist Leptographium longiclavatum presence in the southern Rocky Mountains during a record warm period. Sydowia 2018, 70, 1–10. [Google Scholar]
- McCambridge, W.F. Emergence Period of Black Hills Beetles from Ponderosa Pine in the Central Rocky Mountains; Rocky Mountain Forest and Range Experiment Station, Forest Service, US Department of Agriculture: Washington, WA, USA, 1964; Volume 32. [Google Scholar] [CrossRef]
- Bright, D.E., Jr. The Insects and Arachnids of Canada. Part 2. The Bark Beetles of Canada and Alaska. Coleoptera: Scolytidae; The Insects and Arachnids of Canada: Otawa, ON, Canada, 1976; ISBN 978-0660013206. [Google Scholar]
- Wood, S.L. The Bark and Ambrosia Beetles of North and Central America (Coleoptera: Scolytidae), a Taxonomic Monograph; Great Basin Naturalist Memoirs: Provo, UT, USA, 1982; pp. 1–1359. Available online: https://scholarsarchive.byu.edu/gbnm/vol6/iss1/9 (accessed on 1 December 2017).
- Hirschmann, W. Gange, Teilgange, Stadien von 13 neuen Trichouropoda-Arten (Trichouropodini, Uropodinae). Acarol. Folge 1972, 17, 3–8. [Google Scholar]
- Hirschmann, W. Teilgange, Stadium von 6 neuen Trichouropoda -Arten aus der Verwandtschaft um Tricouropoda dalarnaensis (Sellnick 1952 i. 1.) Hirschmann u. Zirngiebl-Nicol 1961 aus Polen, Mexiko und Kanada (Trichouropodini, Uropodinae). Acarol. Folge 1978, 24, 23–27. [Google Scholar]
- Hirschmann, W.; Wisniewski, J. Weltweite Revision der Ganggattung Trichouropoda Berlese 1916; IV. Die dalarnaensis-Gruppe (Trichouropodini, Uropodinae). Acarologie 1986, 33, 117–148. [Google Scholar]
- Hirschmann, W.; Wisniewski, J. Weltweite Revision der Ganggattung Trichouropoda Berlese 1916. VII. Die sociata-Gruppe (Trichouropodini, Uropodinae). Acarologie 1987, 34, 51–132. [Google Scholar]
- Hirschmann, W.; Wisniewski, J. Weltweite Revision der Ganggattung Trichouropoda Berlese 1916; Nachträge zu den von 1986 bis 1988 revidierten Gruppen (Trichouropodini, Uropodinae). Acarologie 1988, 35, 85–115. [Google Scholar]
- Hirschmann, W.; Wiśniewski, J. Weltweite Revision der Ganggattung Trichouropoda Berlese 1916. I. Die ovalis-Gruppe (Trichouropodini, Uropodinae). Acarologie 1986, 33, 1–81. [Google Scholar]
- Kinn, D.N.; Linit, M.J. A key to Phoretic Mites Commonly Found on Long-Horned Beetles Emerging from Southern Pines; Southern Forest Experiment Station: New Orleans, LA, USA, 1989; Volume 357. [Google Scholar] [CrossRef]
- Kinn, D.N.; Swanston, D.N. Key to Mites Commonly Associated with the Southern Pine Beetle; Dept. of Agriculture, Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1976. Available online: https://www.fs.usda.gov/treesearch/pubs/2018 (accessed on 1 December 2017).
- Lindquist, E.E. New species of Tarsonemus (Acarina-Tarsonemidae) associated with bark beetles. Can. Entomol. 1969, 101, 1291–1314. [Google Scholar] [CrossRef]
- Lindquist, E.E. New species of Ascidae (Acarina-Mesostigmata) associated with forest insect pests. Can. Entomol. 1971, 103, 919–942. [Google Scholar] [CrossRef]
- Lindquist, E.E.; Hunter, P.E. Some mites of the genus Proctolaelaps Berlese (Acarina: Blattisociidae) associated with forest insect pests. Can. Entomol. 1965, 97, 15–32. [Google Scholar] [CrossRef]
- Grylls, B.; Seifert, K. A Synoptic Key to Species of Ophiostoma, Ceratocystis, and Ceratocystiopsis. Ceratocystis and Ophiostoma: Taxonomy, Ecology, and Pathogenicity; American Phytopathological Association: St. Paul, MN, USA, 1993; pp. 161–172. ISBN 978-0890541562. [Google Scholar]
- Lee, S.; Kim, J.J.; Breuil, C. Leptographium longiclavatum sp nov., a new species associated with the mountain pine beetle, Dendroctonus ponderosae. Mycol. Res. 2005, 109, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Robinson-Jeffrey, R.C.; Davidson, R.W. Three new Europhium species with Verticicladiella imperfect states on blue-stained pine. Can. J. Bot. 1968, 46, 1523–1527. [Google Scholar] [CrossRef]
- Rumbold, C.T. A blue stain fungus, Ceratostomella montium n. sp., and some yeasts associated with two species of Dendroctonus. J. Agric. Res. 1941, 62, 0589–0601. [Google Scholar]
- Upadhyay, H.P. A Monograph of Ceratocystis and Ceratocystiopsis; University of Georgia Press: Athens, GA, USA, 1981; ISBN 978-0820305394. [Google Scholar]
- Lewinsohn, D.; Lewinsohn, E.; Bertagnolli, C.L.; Patridge, A.D. Blue-stain fungi and their transport structures on the Douglas-fir beetle. Canadian J. For. Res. 1994, 24, 2275–2283. [Google Scholar] [CrossRef]
- Solheim, H.; Krokene, P. Growth and virulence of mountain pine beetle associated blue-stain fungi, Ophiostoma clavigerum and Ophiostoma montium. Can. J. Bot. 1998, 76, 561–566. [Google Scholar] [CrossRef]
- Bleiker, K.; Six, D. Competition and coexistence in a multi-partner mutualism: Interactions between two fungal symbionts of the mountain pine beetle in beetle-attacked trees. Microb. Ecol. 2009, 57, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S; Springer: New York, NY, USA, 2002. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S.; Christensen, R.H.B.; Singmann, H.; Dai, B.; Grothendieck, G. Package ‘lme4’; R Foundation for Statistical Computing: Vienna, Austria, 2014; p. 12. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P.; Heiberger, R. Multcomp: Simultaneous inference for general linear hypotheses. Biometr. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R package Version 1.1. 2018. Available online: https://rdrr.io/cran/emmeans/ (accessed on 1 January 2018).
- Lombardero, M.J.; Ayres, M.P.; Hofstetter, R.W.; Moser, J.C.; Lepzig, K.D. Strong indirect interactions of Tarsonemus mites (Acarina: Tarsonemidae) and Dendroctonus frontalis (Coleoptera: Scolytidae). Oikos 2003, 102, 243–252. [Google Scholar] [CrossRef]
- Lombardero, M.J.; Klepzig, K.D.; Moser, J.C.; Ayres, M.P. Biology, demography and community interactions of Tarsonemus (Acarina: Tarsonemidae) mites phoretic on Dendroctonus frontalis (Coleoptera: Scolytidae). Agric. For. Entomol. 2000, 2, 193–202. [Google Scholar] [CrossRef]
- Mori, B.A.; Proctor, H.C.; Walter, D.E.; Evenden, M.L. Phoretic mite associates of mountain pine beetle at the leading edge of an infestation in northwestern Alberta, Canada. Can. Entomol. 2011, 143, 44–55. [Google Scholar] [CrossRef]
- Mercado, J.E.; Hofstetter, R.W.; Reboletti, D.M.; Negron, J.F. Phoretic symbionts of the mountain pine beetle (Dendroctonus ponderosae Hopkins). For. Sci. 2014, 60, 512–526. [Google Scholar] [CrossRef]
- Reboletti, D.M. A multi-Partite Mutualism: Bark Beetles, Fungi and Mites; MS Thesis, Northern Arizona University: Flagstaff, AZ, USA, 2008. [Google Scholar]
- Phillips, B.L.; Kelehear, C.; Pizzatto, L.; Brown, G.P.; Barton, D.; Shine, R. Parasites and pathogens lag behind their host during periods of host range advance. Ecology 2010, 91, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Poulin, R. Are there general laws in parasite ecology? Parasitology 2007, 134, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Tsui, C.K.M.; DiGuistini, S.; Wang, Y.; Feau, N.; Dhillon, B.; Bohlmann, J.; Hamelin, R.C. Unequal recombination and evolution of the mating-type (MAT) loci in the pathogenic fungus Grosmannia clavigera and relatives. G3 Genes Genomes Genet. 2013, 3, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Mitton, J.B.; Ferrenberg, S.M. Mountain Pine Beetle Develops an Unprecedented Summer Generation in Response to Climate Warming. Am. Nat. 2012, 179, E163–E171. [Google Scholar] [CrossRef] [PubMed]
- Moser, J.C. Use of sporothecae by phoretic Tarsonemus mites to transport ascospores of coniferous bluestain fungi. Trans. Br. Mycol. Soc. 1985, 84, 750–753. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MPB Mites | Feeding Guild | 2011 n = 95 | 2012 n = 69 | 2013 n = 248 |
|---|---|---|---|---|
| Histiogaster arborsignis Woodring | fungivore | 2 (2.1%) | 1 (1.4%) | 2 (1.2%) |
| Proctolaelaps sp. Berlese | nematophagous | 18 (7.4%) | 0 (0%) | 127 (18.1%) |
| Tarsonemus endophloeus Lindquist | fungivore | NA | NA | 95 (16.1%) |
| Tarsonemus ips Lindquist | fungivore | 51 (21.1%) | 91 (42%) | 330 (14.9%) |
| Trichouropoda sp. Berlese | omnivore | 30 (7.4%) | 38 (26.1%) | 747 (40.3%) |
| Year | Blue-Stain | Maxilla | Elytra | Max. & Ely. | Any |
|---|---|---|---|---|---|
| 2012 n = 38 | O. montium | 15 (39%) | 25 (66%) | 14 (37%) | * 26 (68%) |
| L. longiclavatum | 15 (39%) | 4 (11%) | 4 (11%) | 15 (39%) | |
| G. clavigera | 5 (13%) | 4 (11%) | 0 (0%) | 9 (24%) | |
| 2013 n = 235 | O. montium | 26 (11%) | 28 (12%) | 13 (6%) | * 41 (17%) |
| L. longiclavatum | 29 (12%) | 15 (6%) | 4 (2%) | 40 (17%) | |
| G. clavigera | 9 (4%) | 13 (6%) | 1 (0.004%) | 21 (9%) | |
| 2012–2013 n = 273 | O. montium | 41 (15%) | 53 (19%) | 27 (10%) | 67 (25%) |
| L. longiclavatum | 44 (16%) | 19 (7%) | 8 (3%) | 55 (20%) | |
| G. clavigera | 14 (5%) | 17 (6%) | 1 (0.004%) | 30 (11%) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mercado, J.E.; Ortiz-Santana, B.; Kay, S.L. Fungal Frequency and Mite Load Trends Interact with a Declining Mountain Pine Beetle Population. Forests 2018, 9, 484. https://doi.org/10.3390/f9080484
Mercado JE, Ortiz-Santana B, Kay SL. Fungal Frequency and Mite Load Trends Interact with a Declining Mountain Pine Beetle Population. Forests. 2018; 9(8):484. https://doi.org/10.3390/f9080484
Chicago/Turabian StyleMercado, Javier E., Beatriz Ortiz-Santana, and Shannon L. Kay. 2018. "Fungal Frequency and Mite Load Trends Interact with a Declining Mountain Pine Beetle Population" Forests 9, no. 8: 484. https://doi.org/10.3390/f9080484
APA StyleMercado, J. E., Ortiz-Santana, B., & Kay, S. L. (2018). Fungal Frequency and Mite Load Trends Interact with a Declining Mountain Pine Beetle Population. Forests, 9(8), 484. https://doi.org/10.3390/f9080484