Impacts of a High Nitrogen Load on Foliar Nutrient Status, N Metabolism, and Photosynthetic Capacity in a Cupressus lusitanica Mill. Plantation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design and Sample Collection
2.3. Photosynthesis Measurements
2.4. Leaf Soluble Protein and Free Amino Acid Determination
2.5. Determination of Foliar Nutrient Status
2.6. Statistical Analysis
3. Results
3.1. Effects of Different N Depositions on Foliar Nutrient Status
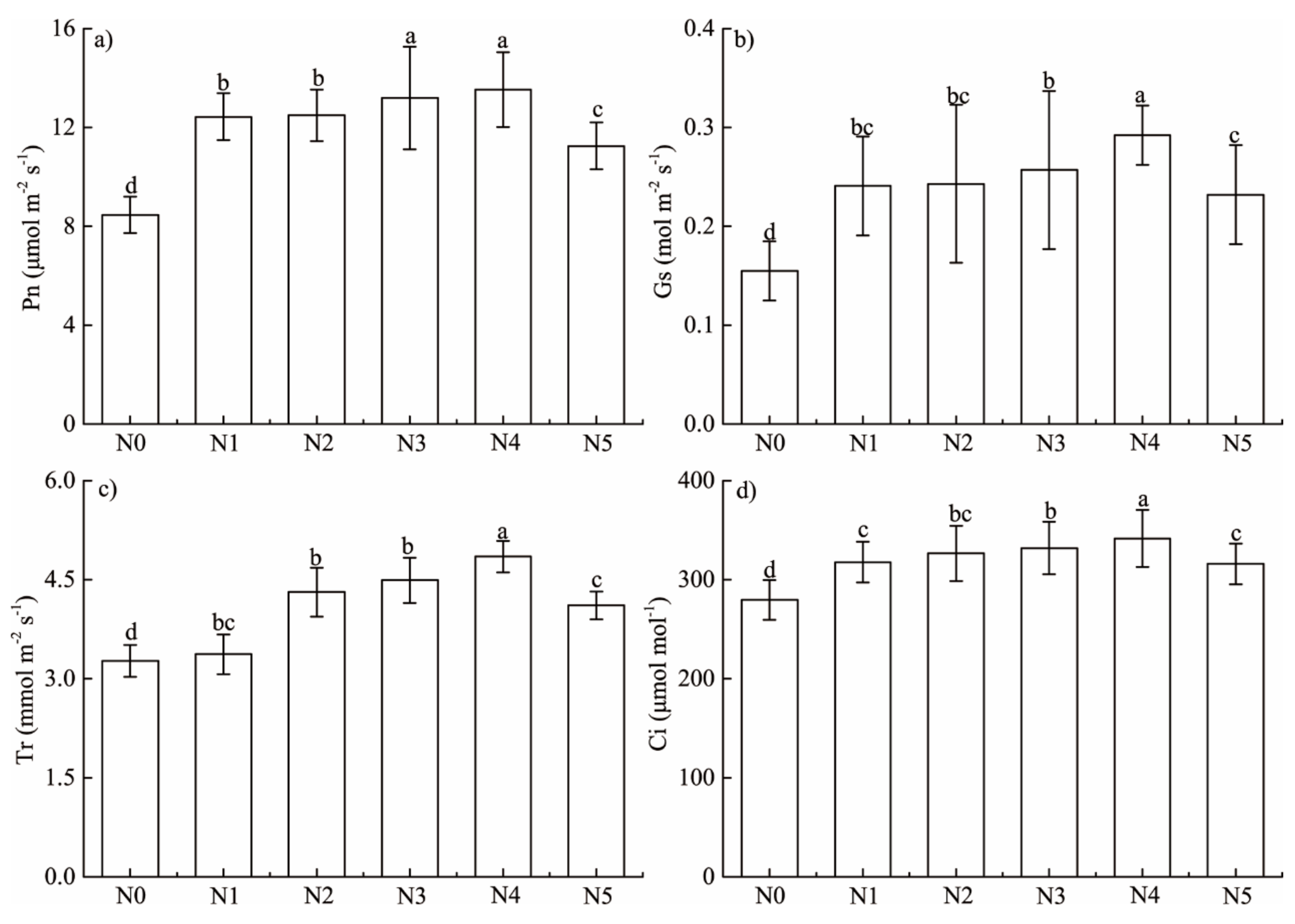
3.2. Responses of Leaf Photosynthetic Parameter and Pigment to Stimulated N Deposition
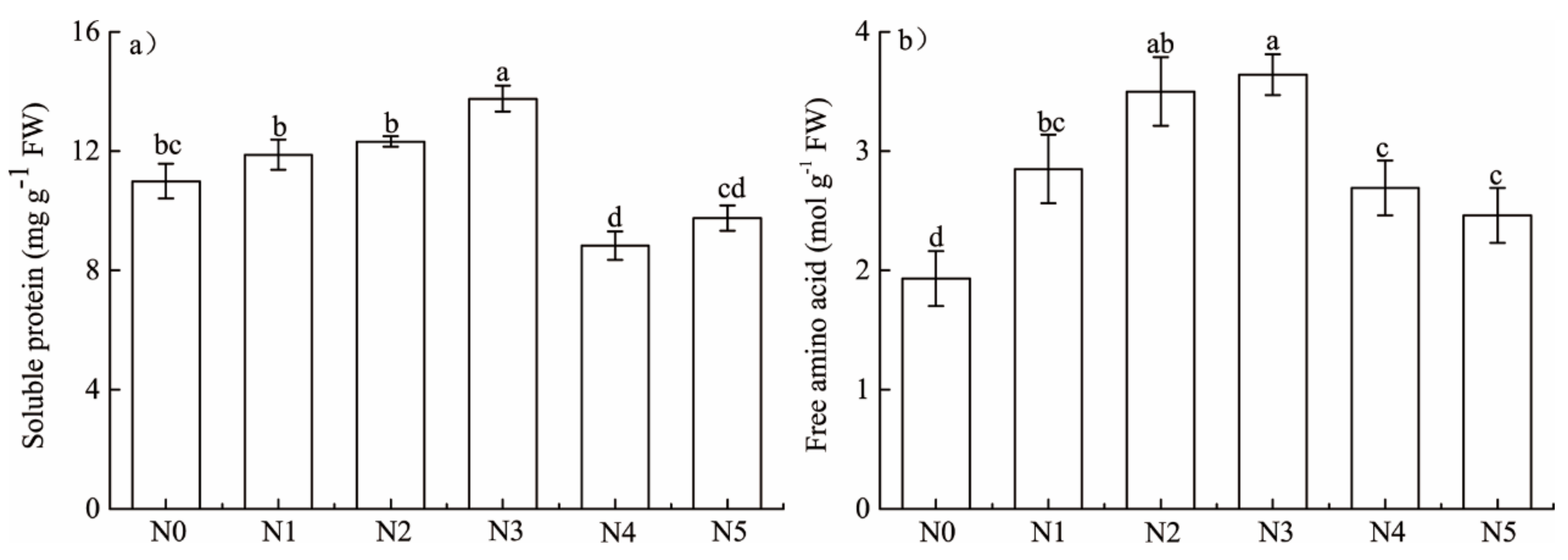
3.3. Effects of N Addition on Foliar Soluble Protein and Free Amino Acid
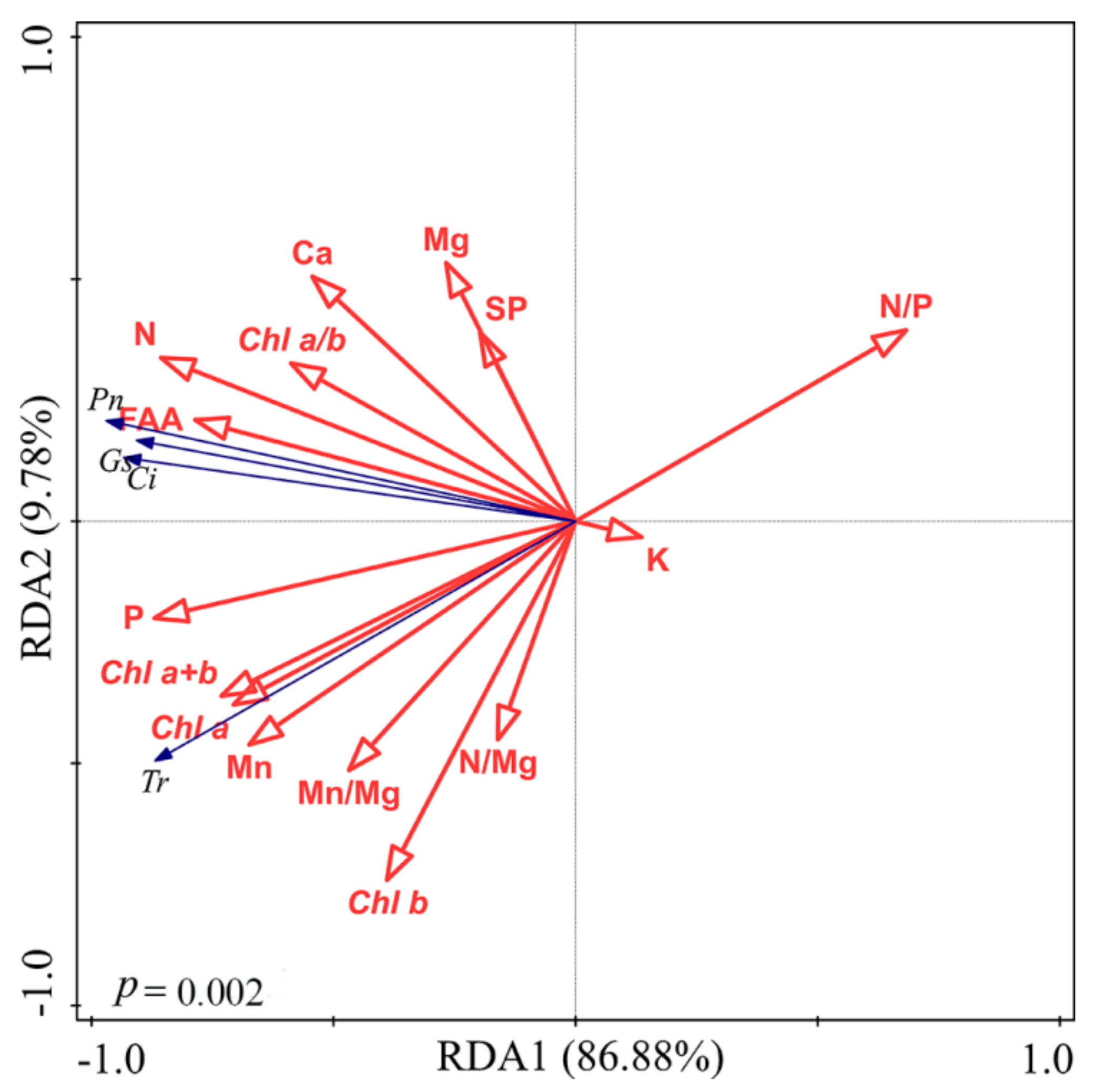
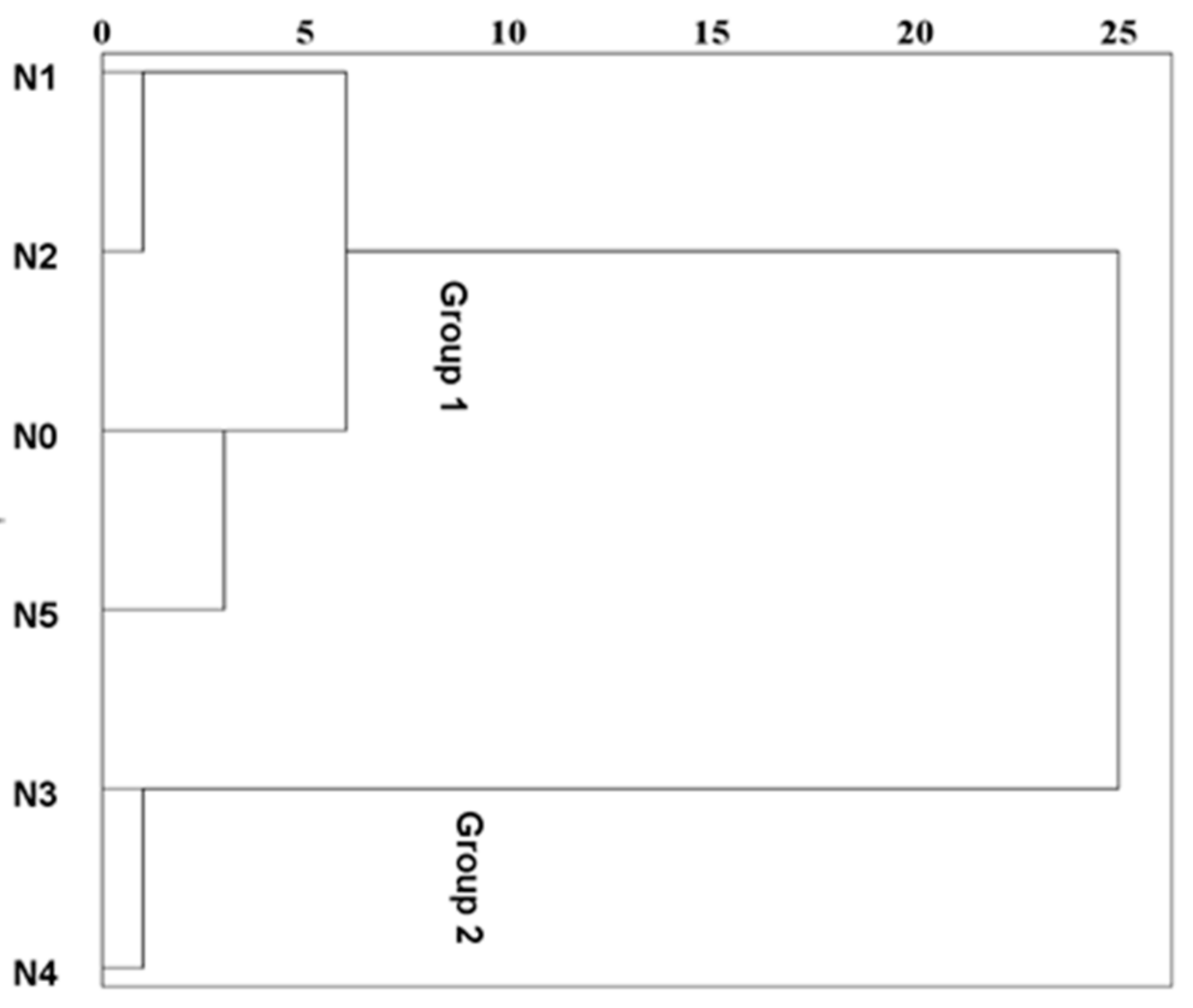
3.4. Multivariate Analysis
4. Discussion
4.1. Responses of Foliar Elements and N Metabolites
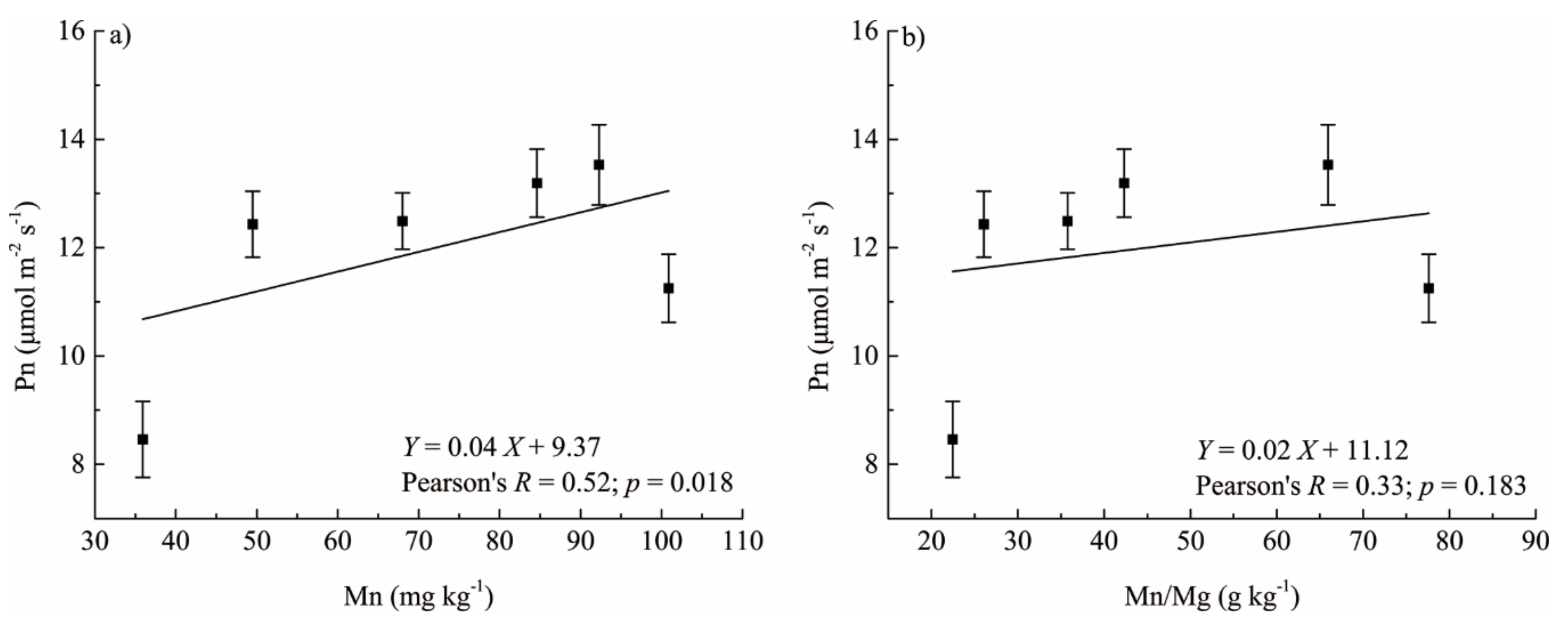
4.2. Responses of Foliar Photosynthetic Capacity
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Erisman, J.W.; Galloway, J.N.; Seitzinger, S.; Bleeker, A.; Dise, N.B.; Petrescu, A.R.; Leach, A.M.; de Vries, W. Consequences of human modification of the global nitrogen cycle. Philos. Trans. R. Soc. B 2013, 368. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Lee, K.; Najjar, R.G.; Jeong, H.; Jeong, H.J. Increasing N abundance in the northwestern Pacific Ocean due to atmospheric nitrogen deposition. Science 2011, 334, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Y.; Han, W.; Tang, A.; Shen, J.; Cui, Z.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P. Enhanced nitrogen deposition over China. Nature 2013, 494, 459. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Duan, L.; Mo, J.; Du, E.; Shen, J.; Lu, X.; Zhang, Y.; Zhou, X.; He, C.; Zhang, F. Nitrogen deposition and its ecological impact in China: An overview. Environ. Pollut. 2011, 159, 2251–2264. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, L.; Chen, Y.; Liu, X.; Xu, W.; Pan, Y.; Duan, L. Atmospheric nitrogen deposition to China: A model analysis on nitrogen budget and critical load exceedance. Atmos. Environ. 2017, 153, 32–40. [Google Scholar] [CrossRef]
- Jia, Y.; Yu, G.; He, N.; Zhan, X.; Fang, H.; Sheng, W.; Zuo, Y.; Zhang, D.; Wang, Q. Spatial and decadal variations in inorganic nitrogen wet deposition in China induced by human activity. Sci. Rep. 2014, 4, 3763. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Gilliam, F.S.; Yu, G.; Li, L. Long-term nitrogen addition decreases carbon leaching in nitrogen-rich forest ecosystems. Biogeosciences 2013, 10, 3931–3941. [Google Scholar] [CrossRef]
- Lü, C.; Tian, H. Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data. J. Geophys. Res. Atmos. 2007, 112. [Google Scholar] [CrossRef] [Green Version]
- Clark, C.M.; Tilman, D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 2008, 451, 712. [Google Scholar] [CrossRef] [PubMed]
- Richter, A.; Burrows, J.P.; Nüß, H.; Granier, C.; Niemeier, U. Increase in tropospheric nitrogen dioxide over China observed from space. Nature 2005, 437, 129. [Google Scholar] [CrossRef] [PubMed]
- Matson, P.; Lohse, K.A.; Hall, S.J. The globalization of nitrogen deposition: Consequences for terrestrial ecosystems. AMBIO 2002, 31, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D.G. Human alteration of the global nitrogen cycle: Sources and consequences. Ecol. Appl. 1997, 7, 737–750. [Google Scholar] [CrossRef]
- Hawkins, B.J.; Jones, M.D.; Kranabetter, J.M. Ectomycorrhizae and tree seedling nitrogen nutrition in forest restoration. New For. 2015, 46, 747–771. [Google Scholar] [CrossRef]
- Soudzilovskaia, N.A.; Douma, J.C.; Akhmetzhanova, A.A.; Bodegom, P.M.; Cornwell, W.K.; Moens, E.J.; Treseder, K.K.; Tibbett, M.; Wang, Y.P.; Cornelissen, J.H.C. Global patterns of plant root colonization intensity by mycorrhizal fungi explained by climate and soil chemistry. Glob. Ecol. Biogeogr. 2015, 24, 371–382. [Google Scholar] [CrossRef]
- Rai, M.; Rathod, D.; Agarkar, G.; Dar, M.; Brestic, M.; Pastore, G.M.; Junior, M.R.M. Fungal growth promotor endophytes: A pragmatic approach towards sustainable food and agriculture. Symbiosis 2014, 62, 63–79. [Google Scholar] [CrossRef]
- Simard, S.W.; Beiler, K.J.; Bingham, M.A.; Deslippe, J.R.; Philip, L.J.; Teste, F.P. Mycorrhizal networks: Mechanisms, ecology and modelling. Fungal Biol. Rev. 2012, 26, 39–60. [Google Scholar] [CrossRef]
- Taylor, A.F.S.; Gebauer, G.; Read, D.J. Uptake of nitrogen and carbon from double-labelled (15N and 13C) glycine by mycorrhizal pine seedlings. New Phytol. 2004, 164, 383–388. [Google Scholar] [CrossRef]
- Binkley, D.; Högberg, P. Tamm Review: Revisiting the influence of nitrogen deposition on Swedish forests. For. Ecol. Manag. 2016, 368, 222–239. [Google Scholar] [CrossRef]
- Lu, X.; Mao, Q.; Mo, J.; Gilliam, F.S.; Zhou, G.; Luo, Y.; Zhang, W.; Huang, J. Divergent responses of soil buffering capacity to long-term N deposition in three typical tropical forests with different land-use history. Environ. Sci. Technol. 2015, 49, 4072–4080. [Google Scholar] [CrossRef] [PubMed]
- Talhelm, A.F.; Pregitzer, K.S.; Burton, A.J. No evidence that chronic nitrogen additions increase photosynthesis in mature sugar maple forests. Ecol. Appl. 2011, 21, 2413–2424. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.K.; Mo, J.M.; Franks, G.; Zhou, G.; Fang, Y. Effects of experimental nitrogen additions on plant diversity in an old-growth tropical forest. Glob. Chang. Biol. 2010, 16, 2688–2700. [Google Scholar] [CrossRef] [Green Version]
- Bauer, G.A.; Bazzaz, F.A.; Minocha, R.; Long, S.; Magill, A.; Aber, J.; Berntson, G.M. Effects of chronic N additions on tissue chemistry, photosynthetic capacity, and carbon sequestration potential of a red pine (Pinus resinosa Ait.) stand in the NE United States. For. Ecol. Manag. 2004, 196, 173–186. [Google Scholar] [CrossRef]
- Mao, Q.; Lu, X.; Mo, H.; Gundersen, P.; Mo, J. Effects of stimulated N deposition on foliar nutrient status, N metabolism and photosynthetic capacity of three dominant understory plant species in a mature tropical forest. Sci. Total Environ. 2018, 610, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.; Lu, X.; Wang, C.; Zhou, K.; Mo, J. Responses of understory plant physiological traits to a decade of nitrogen addition in a tropical reforested ecosystem. For. Ecol. Manag. 2017, 401, 65–74. [Google Scholar] [CrossRef]
- Lu, X.; Mao, Q.; Gilliam, F.S.; Luo, Y.; Mo, J. Nitrogen deposition contributes to soil acidification in tropical ecosystems. Glob. Chang. Biol. 2014, 20, 3790–3801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bubier, J.L.; Smith, R.; Juutinen, S.; Moore, T.R.; Minocha, R.; Long, S.; Minocha, S. Effects of nutrient addition on leaf chemistry, morphology, and photosynthetic capacity of three bog shrubs. Oecologia 2011, 167, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.K.; Mo, J.M.; Gundersern, P.; Zhu, W.X.; Zhou, G.Y.; De-Jun, L.I.; Xu, Z. Effect of stimulated N deposition on soil exchangeable cations in three forest types of subtropical China. Pedosphere 2009, 19, 189–198. [Google Scholar] [CrossRef]
- Warren, C.R.; Mcgrath, J.F.; Adams, M.A. Differential effects of N, P and K on photosynthesis and partitioning of N in Pinus pinaster needles. Ann. For. Sci. 2005, 62, 1–8. [Google Scholar] [CrossRef]
- Magill, A.H.; Aber, J.D.; Berntson, G.M.; McDowell, W.H.; Nadelhoffer, K.J.; Melillo, J.M.; Steudler, P. Long-term nitrogen additions and nitrogen saturation in two temperate forests. Ecosystems 2000, 3, 238–253. [Google Scholar] [CrossRef]
- Pérezharguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bretharte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E. New handbook for standardized measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, W.; Li, N.; Liu, Y.; Zheng, X.; Hao, G. Photosynthesis and growth responses of Fraxinus mandshurica Rupr. seedlings to a gradient of simulated nitrogen deposition. Ann. For. Sci. 2018, 75, 1. [Google Scholar] [CrossRef]
- Zeng, J.; Sheng, H.; Liu, Y.; Wang, Y.; Wang, Y.; Kang, H.; Fan, X.; Sha, L.; Yuan, S.; Zhou, Y. High nitrogen supply induces physiological responsiveness to long photoperiod in Barley. Front. Plant Sci. 2017, 8, 569. [Google Scholar] [CrossRef] [PubMed]
- Tavarini, S.; Pagano, I.; Guidi, L.; Angelini, L.G. Impact of nitrogen supply on growth, steviol glycosides and photosynthesis in Stevia rebaudiana Bertoni. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2016, 150, 953–962. [Google Scholar]
- Nakaji, T.; Takenaga, S.; Kuroha, M.; Izuta, T. Photosynthetic response of Pines densiflora seedlings to high nitrogen load. Environ. Sci. 2002, 9, 269–282. [Google Scholar]
- Zhang, X.; Davidson, E.A.; Mauzerall, D.L.; Searchinger, T.D.; Dumas, P.; Shen, Y. Managing nitrogen for sustainable development. Nature 2015, 528, 51. [Google Scholar] [CrossRef] [PubMed]
- Garnier, E.; Navas, M. A trait-based approach to comparative functional plant ecology: Concepts, methods and applications for agroecology. A review. Agron. Sustain. Dev. 2012, 32, 365–399. [Google Scholar] [CrossRef]
- Brussaard, L.; Caron, P.; Campbell, B.; Lipper, L.; Mainka, S.; Rabbinge, R.; Babin, D.; Pulleman, M. Reconciling biodiversity conservation and food security: Scientific challenges for a new agriculture. Curr. Opin. Environ. Sustain. 2010, 2, 34–42. [Google Scholar] [CrossRef]
- Rose, L.; Rubarth, M.C.; Hertel, D.; Leuschner, C. Management alters interspecific leaf trait relationships and trait-based species rankings in permanent meadows. J. Veg. Sci. 2013, 24, 239–250. [Google Scholar] [CrossRef]
- Elvir, J.A.; Wiersma, G.B.; Day, M.E.; Greenwood, M.S.; Fernandez, I.J. Effects of enhanced nitrogen deposition on foliar chemistry and physiological processes of forest trees at the Bear Brook Watershed in Maine. For. Ecol. Manag. 2006, 221, 207–214. [Google Scholar] [CrossRef]
- Hood, I.A.; Gardner, J.F.; Hood, R.J.; Smith, B.M.; Phillips, G.D. Pruning and cypress canker in New Zealand. Australas. Plant Pathol. 2009, 38, 472–477. [Google Scholar] [CrossRef]
- Cai, Q.; Wan, F.-X. Research on cold tolerance of Cupressus lusitanica Mill. seedling. J. Anhui Agric. Sci. 2014, 9, 51. (In Chinese) [Google Scholar]
- Shen, F.; Yuan, Y.; Fan, H.; Liu, W.; Liu, Y. Effects of elevated nitrogen deposition on soil organic carbon mineralization and soil enzyme activities in a Chinese fir plantation. Acta Ecol. Sin. 2012, 32, 517–527. (In Chinese) [Google Scholar] [CrossRef]
- Qin, Y.W.; Wan, F.X. Effect of fertilization on seedlings growth of provenience of Cupressus lusitanica Mill. For. Sci. Tech. 2016, 5, 115–119. (In Chinese) [Google Scholar]
- Gu, T.H. Research on Salt Tolerance and Adaptability of Cupressus lusitanica Mill. Master’s Thesis, Nanjing Forestry University, Nanjing, China, 2011. [Google Scholar]
- Bao, H. Studies on Growth Rhytm and Photosynthesis Characteristics of Cupressus lusitanica Mill. Master’s Thesis, Nanjing Forestry University, Nanjing, China, 2012. [Google Scholar]
- Hu, L. A Study on Fertilizer Effect of Populus tomentosa Carr. Plantation. Master’s Thesis, Beijing Forestry University, Beijing, China, 2010. [Google Scholar]
- Li, W. Growth and Physiological Responses of Betula platyphylla to Fertilization in Juvenile Plantation. Master’s Thesis, Northeast Forestry University, Changchun, China, 2015. [Google Scholar]
- Dang, Y.C. Soil Microorganism and Soil Enzyme Response of Fertilization in Young Poplar Forest. Master’s Thesis, Sichuan Agriculture University, Sichuan, China, 2012. [Google Scholar]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Xu, Z.Z.; Zhou, G.S. Nitrogen metabolism and photosynthesis in Leymus chinensis in response to long-term soil drought. J. Plant Growth Regul. 2006, 25, 252–266. [Google Scholar] [CrossRef]
- Barnett, N.M.; Naylor, A.W. Amino acid and protein metabolism in Bermuda grass during water stress. Plant Physiol. 1966, 41, 1222–1230. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, C.C.; Townsend, A.R.; Taylor, P.; Alvarez Clare, S.; Bustamante, M.; Chuyong, G.; Dobrowski, S.Z.; Grierson, P.; Harms, K.E.; Houlton, B.Z. Relationships among net primary productivity, nutrients and climate in tropical rain forest: A pan-tropical analysis. Ecol. Lett. 2011, 14, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Matson, P.A.; McDowell, W.H.; Townsend, A.R.; Vitousek, P.M. The globalization of N deposition: Ecosystem consequences in tropical environments. Biochemistry 1999, 46, 67–83. [Google Scholar]
- Lu, X.; Mo, J.; Gilliam, F.S.; Fang, H.; Zhu, F.; Fang, Y.; Zhang, W.; Huang, J. Nitrogen addition shapes soil phosphorus availability in two reforested tropical forests in southern China. Biotropica 2012, 44, 302–311. [Google Scholar] [CrossRef]
- Wallander, H.; Nylund, J. Effects of excess nitrogen and phosphorus starvation on the extramatrical mycelium of ectomycorrhizas of Pinus sylvestris L. New Phytol. 1992, 120, 495–503. [Google Scholar] [CrossRef]
- Schulze, E. Air pollution and forest decline in a spruce (Picea abies) forest. Science 1989, 244, 776–783. [Google Scholar] [CrossRef] [PubMed]
- VanderSchaaf, C.L.; Moore, J.A.; Kingery, J.L. The effect of multi-nutrient fertilization on understory vegetation nutrient concentrations in inland Northwest conifer stands. For. Ecol. Manag. 2004, 190, 201–218. [Google Scholar] [CrossRef]
- Jeong, J.; Jo, C.G.; Baek, G.W.; Park, J.; Ma, H.; Yoo, B.O.; Kim, C. Soil and the foliage nutrient status following soil amendment applications in a Japanese cypress (Chamaecyparis obtusa Endlicher) plantation. J. Sustain. For. 2017, 36, 289–303. [Google Scholar] [CrossRef]
- Wortman, E.; Tomaszewski, T.; Waldner, P.; Schleppi, P.; Thimonier, A.; Eugster, W.; Buchmann, N.; Sievering, H. Atmospheric nitrogen deposition and canopy retention influences on photosynthetic performance at two high nitrogen deposition Swiss forests. Tellus B Chem. Phys. Meteorol. 2012, 64. [Google Scholar] [CrossRef]
- Nakaji, T.; Fukami, M.; Dokiya, Y.; Izuta, T. Effects of high nitrogen load on growth, photosynthesis and nutrient status of Cryptomeria japonica and Pinus densiflora seedlings. Trees 2001, 15, 453–461. [Google Scholar]
- Diem, B.; Godbold, D.L. Potassium, calcium and magnesium antagonism in clones of Populus trichocarpa. Plant Soil 1993, 155, 411–414. [Google Scholar] [CrossRef]
- Gurmesa, G.A.; Lu, X.; Gundersen, P.; Mao, Q.; Zhou, K.; Fang, Y.; Mo, J. High retention of 15N-labeled nitrogen deposition in a nitrogen saturated old-growth tropical forest. Glob. Chang. Biol. 2016, 22, 3608–3620. [Google Scholar] [CrossRef] [PubMed]
- Näsholm, T.; Ericsson, A.; Nordén, L.G. Accumulation of amino acids in some boreal forest plants in response to increased nitrogen availability. New Phytol. 1994, 126, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Hák, R.; Rinderle-Zimmer, U.; Lichtenthaler, H.K.; Nátr, L. Chlorophyll a fluorescence signatures of nitrogen deficient barley leaves. Photosynthetica 1993, 28, 151–159. [Google Scholar]
- Brown, H.R. A difference in N use efficiency in C3 and C4 plants and its implications in adaptation and evolution. Crop Sci. 1978, 18, 105–115. [Google Scholar] [CrossRef]
- Guo, J.; Wu, Y.; Wang, B.; Lu, Y.; Cao, F.; Wang, G. The effects of fertilization on the growth and physiological characteristics of Ginkgo biloba L. Forests 2016, 7, 293. [Google Scholar] [CrossRef]
- Yan, H.; Wu, Q.; Ding, J.; Zhang, S.R. Effects of precipitation and nitrogen addition on photosynthetically eco-physiological characteristics and biomass of four tree seedlings in Gutian Mountain, Zhejiang Province, China. Acta Ecol. Sin. 2013, 33, 4226–4236. [Google Scholar] [CrossRef]
- Boussadia, O.; Steppe, K.; Zgallai, H.; El Hadj, S.B.; Braham, M.; Lemeur, R.; Van Labeke, M. Effects of nitrogen deficiency on leaf photosynthesis, carbohydrate status and biomass production in two olive cultivars ‘Meski’and ‘Koroneiki’. Sci. Hortic. 2010, 123, 336–342. [Google Scholar] [CrossRef]
- Von Wettstein, D.; Gough, S.; Kannangara, C.G. Chlorophyll biosynthesis. Plant Cell 1995, 7, 1039. [Google Scholar] [CrossRef] [PubMed]
- Shah, J.M.; Bukhari, S.A.H.; Zeng, J.; Quan, X.; Ali, E.; Muhammad, N.; Zhang, G. Nitrogen (N) metabolism related enzyme activities, cell ultrastructure and nutrient contents as affected by N level and barley genotype. J. Integr. Agric. 2017, 16, 190–198. [Google Scholar] [CrossRef]
- Brestic, M.; Zivcak, M.; Olsovska, K.; Shao, H.B.; Kalaji, H.M.; Allakhverdiev, S.I. Reduced glutamine synthetase activity plays a role in control of photosynthetic responses to high light in barley leaves. Plant Physiol. Biochem. 2014, 81, 74–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswal, A.K.; Pattanayak, G.K.; Pandey, S.S.; Leelavathi, S.; Reddy, V.S.; Govindjee, X.; Tripathy, B.C. Light intensity-dependent modulation of chlorophyll b biosynthesis and photosynthesis by overexpression of chlorophyll a oxygenase (CAO) in tobacco. Plant Physiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Porra, R.J.; Schäfer, W.; Cmiel, E.; Katheder, I.; Scheer, H. Derivation of the formyl-group oxygen of chlorophyll b from molecular oxygen in greening leaves of a higher plant (Zea mays). FEBS Lett. 1993, 323, 31–34. [Google Scholar] [CrossRef] [Green Version]
- Cechin, I.; Fumis, T. Effect of nitrogen supply on growth and photosynthesis of sunflower plants grown in the greenhouse. Plant Sci. 2004, 166, 1379–1385. [Google Scholar] [CrossRef] [Green Version]
- Kalaji, H.M.; Oukarroum, A.; Alexandrov, V.; Kouzmanova, M.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Allakhverdiev, S.I.; Goltsev, V. Identification of nutrient deficiency in maize and tomato plants by in vivo chlorophyll a fluorescence measurements. Plant Physiol. Biochem. 2014, 81, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Laing, W.; Greer, D.; Sun, O.; Beets, P.; Lowe, A.; Payn, T. Physiological impacts of Mg deficiency in Pinus radiata: Growth and photosynthesis. New Phytol. 2000, 146, 47–57. [Google Scholar] [CrossRef]
- McHargue, J.S. The role of manganese in plants 1. J. Am. Chem. Soc. 1922, 44, 1592–1598. [Google Scholar] [CrossRef]
- Sherman, J.; Heerema, R.J.; VanLeeuwen, D.; Hilaire, R.S. Optimal manganese nutrition increases photosynthesis of immature Pecan trees. HortScience 2017, 52, 634–640. [Google Scholar] [CrossRef]
- Ohki, K. Manganese deficiency and toxicity effects on photosynthesis, chlorophyll, and transpiration in Wheat 1. Crop Sci. 1985, 25, 187–191. [Google Scholar] [CrossRef]
- Flückiger, W.; Braun, S. Nitrogen deposition in Swiss forests and its possible relevance for leaf nutrient status, parasite attacks and soil acidification. In Proceedings of the Nitrogen, the Confer-N-s: First International Nitrogen Conference, Noordwijkerhout, The Netherlands, 23–27 March 1998; pp. 69–76. [Google Scholar] [CrossRef]
- Wilson, E.J.; Skeffington, R.A. The effects of excess nitrogen deposition on young Norway spruce trees. Part II The vegetation. Environ. Pollut. 1994, 86, 153–160. [Google Scholar] [CrossRef]
- Houtz, R.L.; Nable, R.O.; Cheniae, G.M. Evidence for effects on the in vivo activity of ribulose-bisphosphate carboxylase/oxygenase during development of Mn toxicity in tobacco. Plant Physiol. 1988, 86, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Jordan, D.B.; Ogren, W.L. A sensitive assay procedure for simultaneous determination of ribulose-1, 5-bisphosphate carboxylase and oxygenase activities. Plant Physiol. 1981, 67, 237–245. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | N (%) | P (%) | K (%) | Ca (%) | Mg (%) | Mn (mg kg−1) | N/P | N/Mg | Mn/Mg (g kg−1) |
|---|---|---|---|---|---|---|---|---|---|
| N0 | 1.24 ± 0.01 c | 0.14 ± 0.01 c | 0.68 ± 0.05 a | 0.64 ± 0.03 c | 0.16 ± 0.02 b | 35.93 ± 3.30 d | 8.86 ± 1.01 a | 7.75 ± 0.28 b | 22.46 ± 2.17 e |
| N1 | 1.40 ± 0.04 a | 0.16 ± 0.01 ab | 0.63 ± 0.03 a | 1.19 ± 0.14 a | 0.19 ± 0.02 a | 49.51 ± 6.78 cd | 8.75 ± 1.10 ab | 7.37 ± 1.00 b | 26.06 ± 2.65 de |
| N2 | 1.40 ± 0.03 a | 0.17 ± 0.01 ab | 0.61 ± 0.02 a | 1.22 ± 0.03 a | 0.19 ± 0.01 a | 67.99 ± 7.31 bc | 8.23 ± 0.99 ab | 7.37 ± 0.22 b | 35.78 ± 4.00 cd |
| N3 | 1.39 ± 0.01 a | 0.19 ± 0.02 ab | 0.60 ± 0.02 a | 1.08 ± 0.15 ab | 0.20 ± 0.02 a | 84.61 ± 8.00 ab | 7.31 ± 1.00 ab | 6.95 ± 0.49 b | 42.31 ± 5.37 c |
| N4 | 1.39 ± 0.07 a | 0.22 ± 0.02 a | 0.58 ± 0.02 a | 0.94 ± 0.04 b | 0.14 ± 0.01 c | 92.29 ± 5.76 a | 6.32 ± 0.50 b | 9.93 ± 0.93 a | 65.92 ± 8.21 b |
| N5 | 1.33 ± 0.04 b | 0.15 ± 0.01 b | 0.58 ± 0.01 a | 0.52 ± 0.02 c | 0.13 ± 0.08 c | 100.87 ± 6.13 a | 8.86 ± 0.73 ab | 10.23 ± 0.56 a | 77.59 ± 7.13 a |
| Treatment | Chl a (mg g−1) | Chl b (mg g−1) | Chl a + b (mg g−1) | Chl a/b |
|---|---|---|---|---|
| N0 | 0.51 ± 0.01 bc | 0.12 ± 0.000 cd | 0.63 ± 0.01 c | 4.38 ± 0.11 bc |
| N1 | 0.58 ± 0.05 c | 0.09 ± 0.003 d | 0.67 ± 0.05 c | 6.80 ± 0.41 a |
| N2 | 0.59 ± 0.04 c | 0.13 ± 0.007 cd | 0.72 ± 0.04 c | 4.59 ± 0.37 b |
| N3 | 0.97 ± 0.07 a | 0.14 ± 0.008 b | 1.11 ± 0.10 a | 7.07 ± 0.45 a |
| N4 | 1.01 ± 0.01 a | 0.19 ± 0.002 a | 1.20 ± 0.01 a | 7.11 ± 0.19 a |
| N5 | 0.76 ± 0.05 b | 0.13 ± 0.01 bc | 0.89 ± 0.05 b | 3.97 ± 0.22 c |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, R.; Lu, Y.; Wan, F.; Wang, Y.; Pan, X. Impacts of a High Nitrogen Load on Foliar Nutrient Status, N Metabolism, and Photosynthetic Capacity in a Cupressus lusitanica Mill. Plantation. Forests 2018, 9, 483. https://doi.org/10.3390/f9080483
Li R, Lu Y, Wan F, Wang Y, Pan X. Impacts of a High Nitrogen Load on Foliar Nutrient Status, N Metabolism, and Photosynthetic Capacity in a Cupressus lusitanica Mill. Plantation. Forests. 2018; 9(8):483. https://doi.org/10.3390/f9080483
Chicago/Turabian StyleLi, Ruirui, Yi Lu, Fuxu Wan, Yiming Wang, and Xiaocheng Pan. 2018. "Impacts of a High Nitrogen Load on Foliar Nutrient Status, N Metabolism, and Photosynthetic Capacity in a Cupressus lusitanica Mill. Plantation" Forests 9, no. 8: 483. https://doi.org/10.3390/f9080483