Changes in Phylogenetic Community Structure of the Seedling Layer Following Hurricane Disturbance in a Human-Impacted Tropical Forest

 ,
, 


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Seedling Census
2.3. Analysis of Phylogenetic Community Structure
2.4. Phylogenetic Neighborhood Effects on Seedling Survival
2.5. Temporal Variation in Community Mean Seed Mass
3. Results
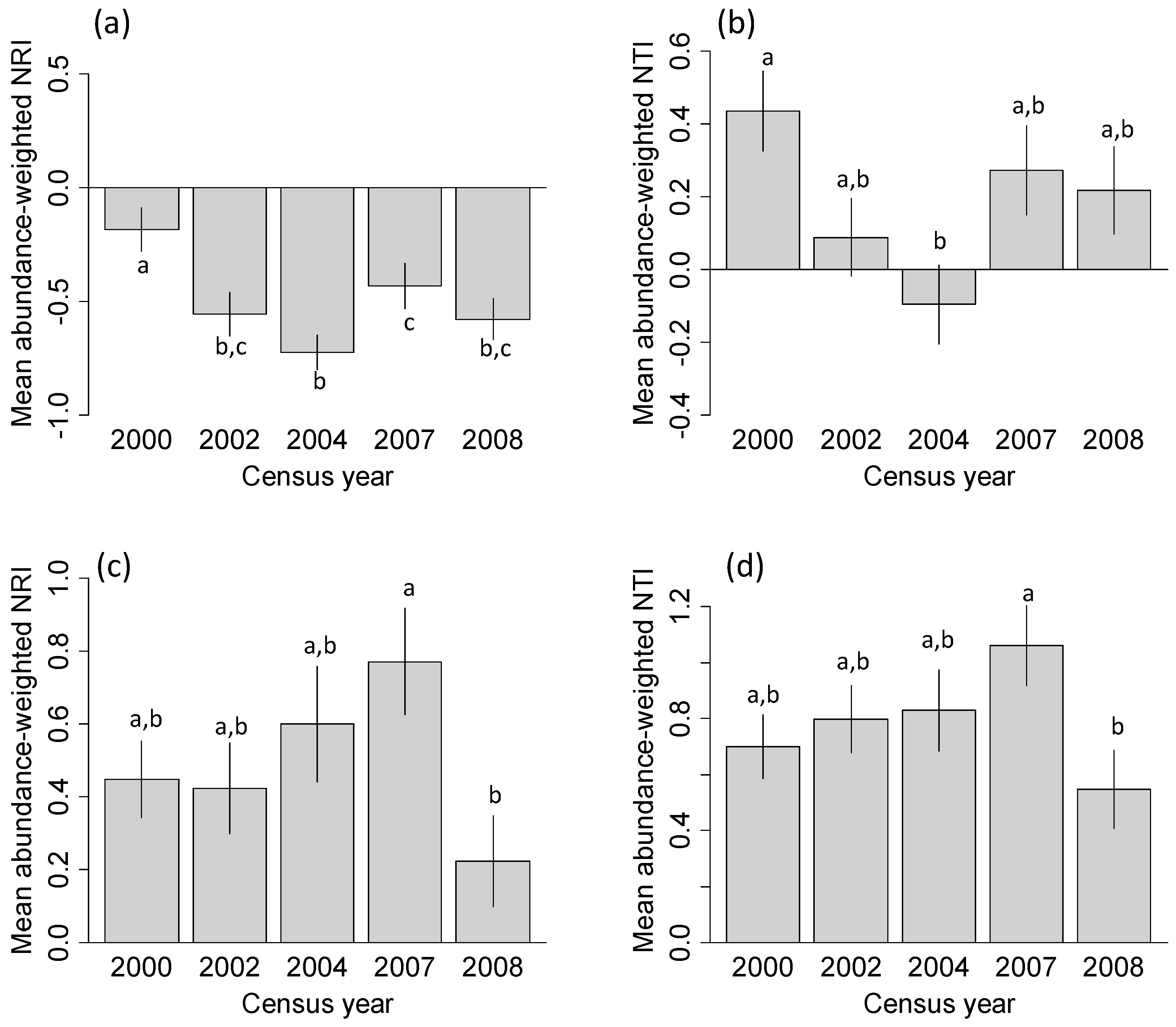
3.1. Phylogenetic Community Structure Following Hurricane Disturbance
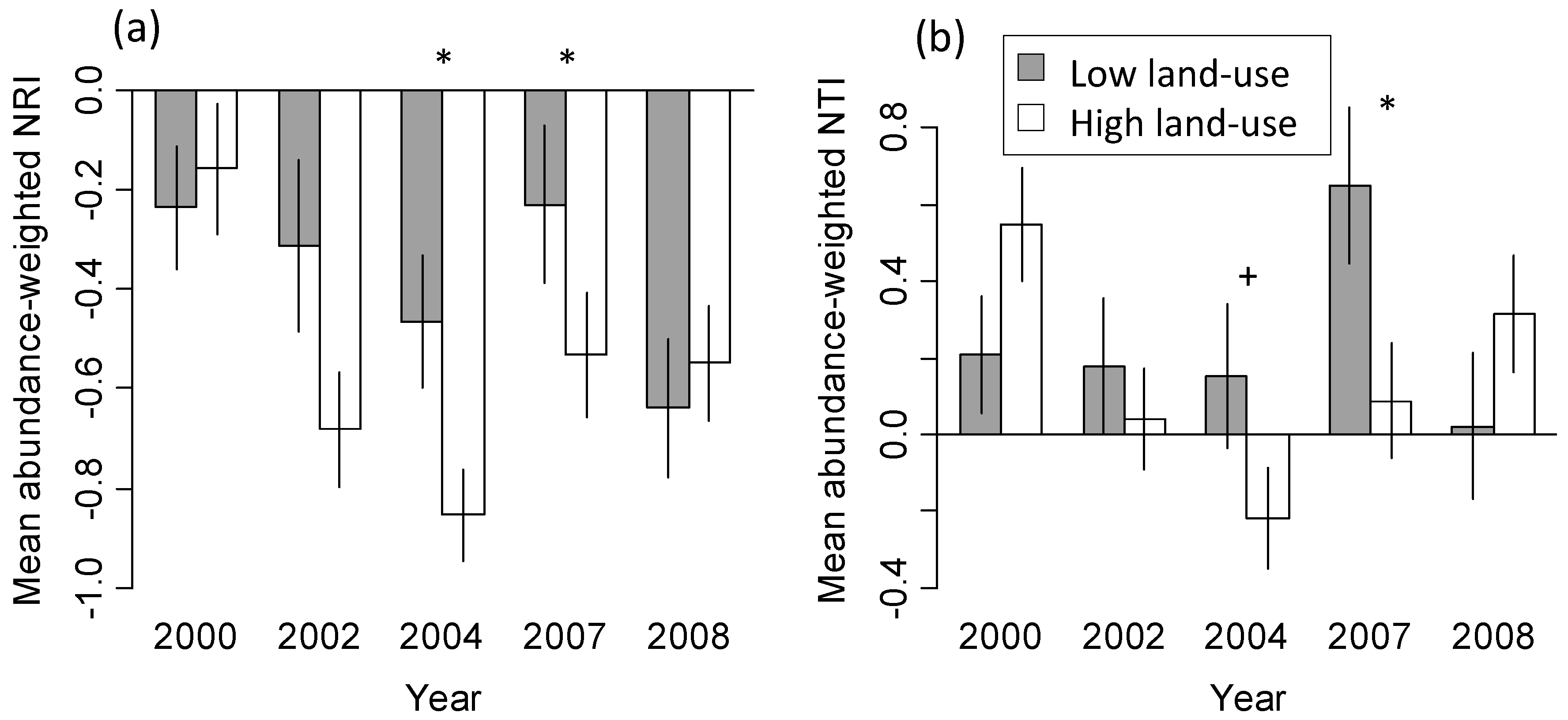
3.2. Phylogenetic Community Structure and Land-Use History
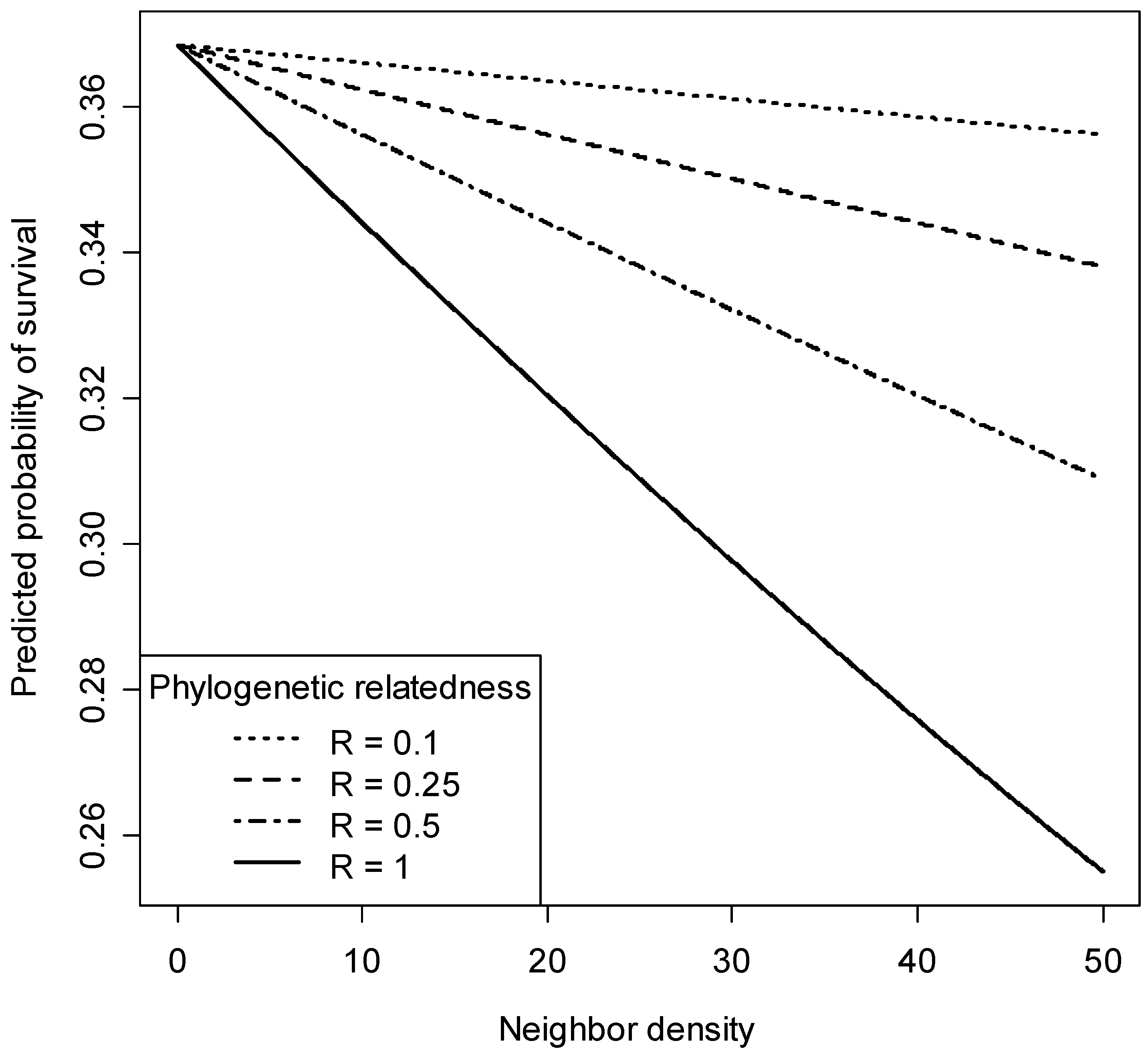
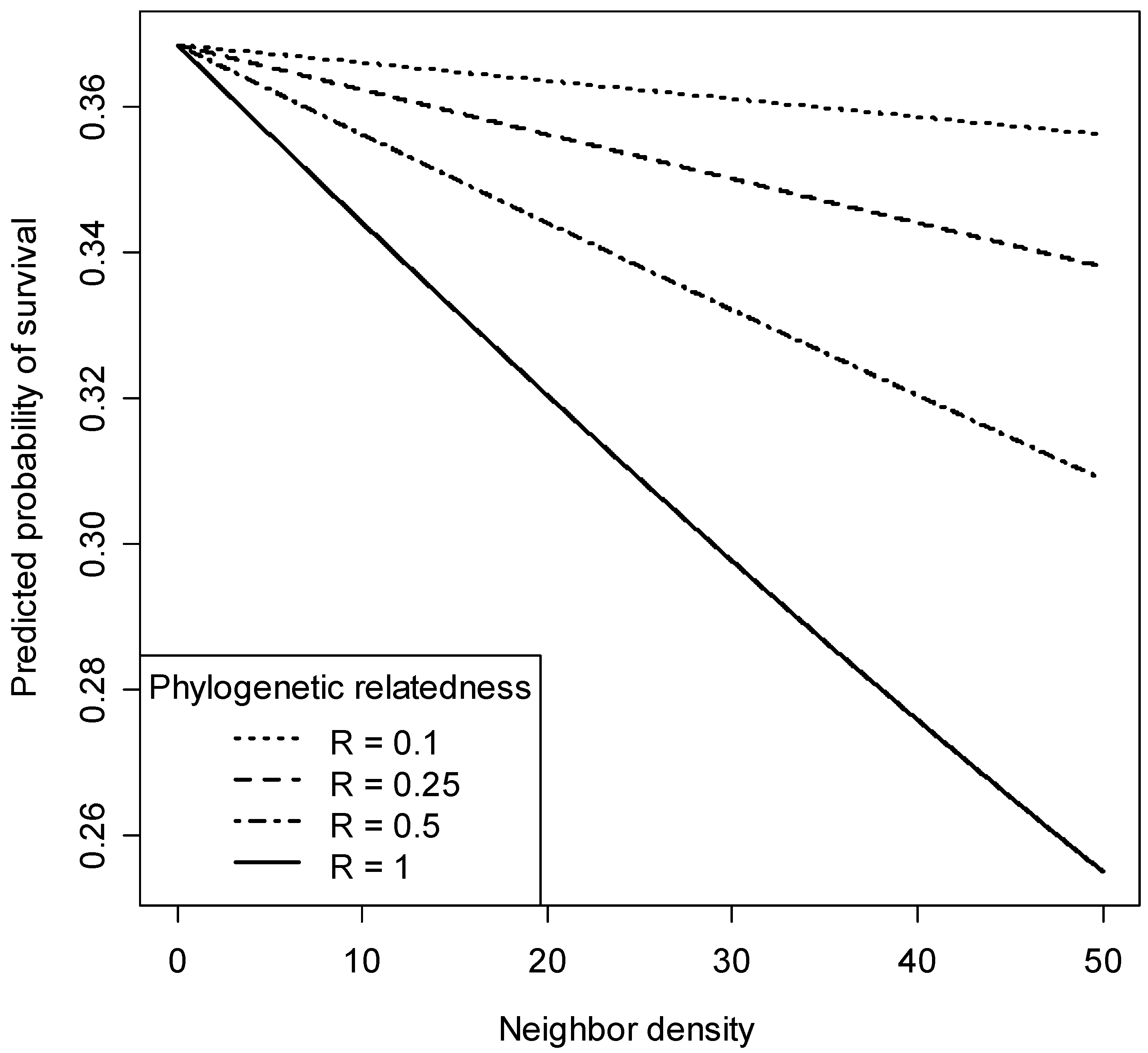
3.3. Phylogenetic Neighborhood Effects on Seedling Survival
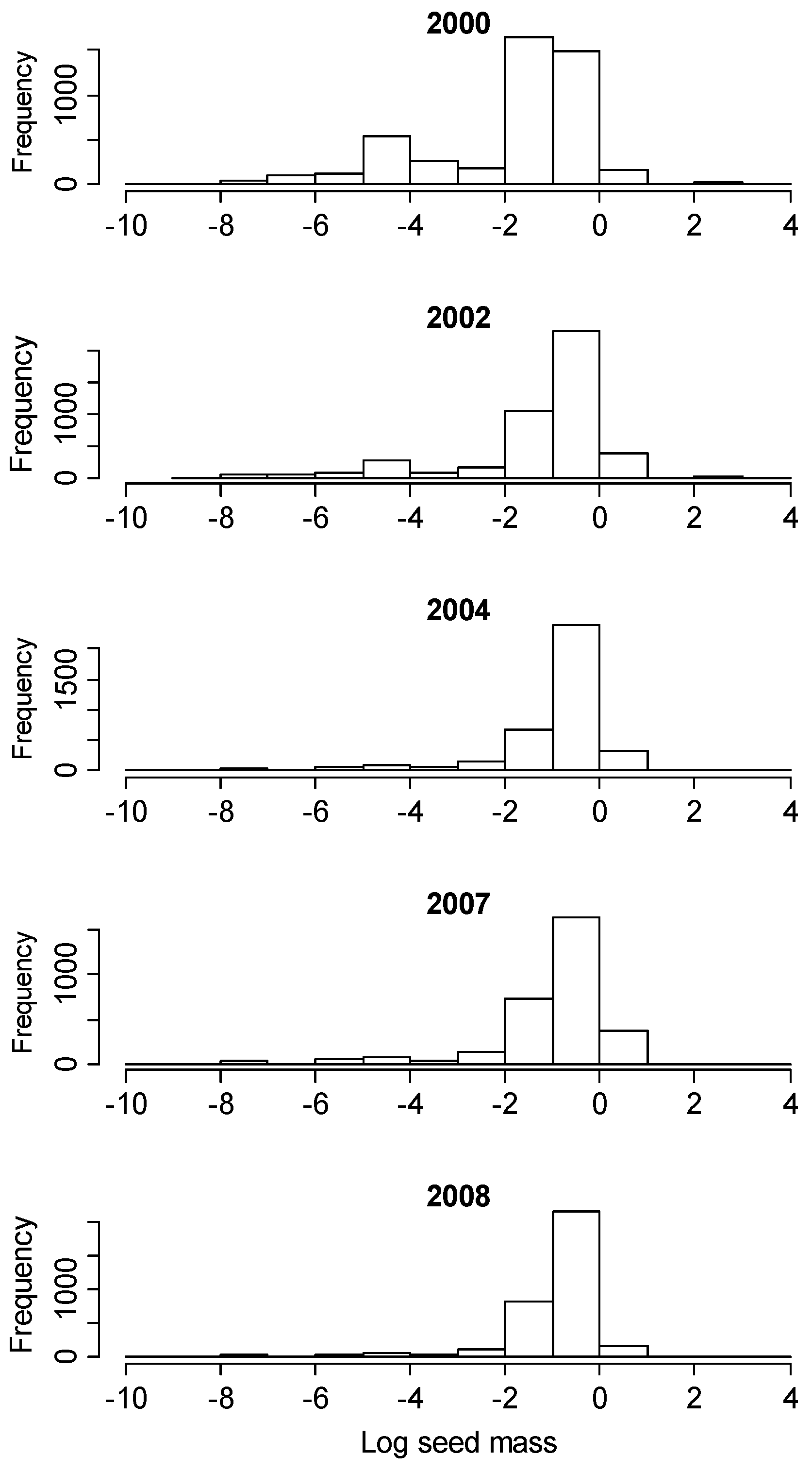
3.4. Changes in Seed Mass Values Following Hurricane Disturbance
4. Discussion
4.1. Phylogenetic Community Dynamics Following Hurricane Disturbance
4.2. Signature of Past Land-Use on Phylogenetic Community Structure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sousa, W.P. The role of disturbance in natural communities. Annu. Rev. Ecol. Syst. 1984, 15, 353–391. [Google Scholar] [CrossRef]
- Canham, C.D.; Marks, P.L. The response of woody plants to disturbance patterns of establishment and growth. In The Ecology of Natural Disturbance and Patch Dynamics; White, P.S., Pickett, S.T.A., Eds.; Academic Press: New York, NY, USA, 1985; pp. 197–216. [Google Scholar]
- Grubb, P.J. The maintenance of species-richness in plant communities: The importance of regeneration niche. Biol. Rev. 1977, 52, 107–145. [Google Scholar] [CrossRef]
- Clark, J.S. Testing disturbance theory with long-term data: Alternative life-history solutions to the distribution of events. Am. Nat. 1996, 148, 976–996. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in Tropical Rain Forests and Coral Reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Pacala, S.W.; Rees, M. Models suggesting field experiments to test two hypotheses explaining successional diversity. Am. Nat. 1998, 152, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Loehle, C. Strategy Space and the Disturbance Spectrum: A Life-History Model for Tree Species Coexistence. Am. Nat. 2000, 156, 14–33. [Google Scholar] [CrossRef] [PubMed]
- Meiners, S.J.; Cadotte, M.W.; Fridley, J.D.; Pickett, S.T.A.; Walker, L.R. Is successional research nearing its climax? New approaches for understanding dynamic communities. Funct. Ecol. 2015, 29, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Verdu, M.; Gomez-Aparicio, L.; Valiente-Banuet, A. Phylogenetic relatedness as a tool in restoration ecology: A meta-analysis. Proc. R. Soc. Lond. B Biol. Sci. 2012, 279, 1761–1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdú, M.; Pausas, J.G. Fire drives phylogenetic clustering in Mediterranean Basin woody plant communities. J. Ecol. 2007, 95, 1316–1323. [Google Scholar] [CrossRef] [Green Version]
- Knapp, S.; Kühn, I.; Schweiger, O.; Klotz, S. Challenging urban species diversity: Contrasting phylogenetic patterns across plant functional groups in Germany. Ecol. Lett. 2008, 11, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Dinnage, R. Disturbance alters the phylogenetic composition and structure of plant communities in an old field system. PLoS ONE 2009, 4, e7071. [Google Scholar] [CrossRef] [PubMed]
- Letcher, S.G. Phylogenetic structure of angiosperm communities during tropical forest succession. Proc. R. Soc. Lond. B Biol. Sci. 2010, 277, 97–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitfeld, T.J.S.; Kress, W.J.; Erickson, D.L.; Weiblen, G.D. Change in community phylogenetic structure during tropical forest succession: Evidence from New Guinea. Ecography 2012, 35, 821–830. [Google Scholar] [CrossRef]
- Purschke, O.; Schmid, B.C.; Sykes, M.T.; Poschlod, P.; Michalski, S.G.; Durka, W.; Kühn, I.; Winter, M.; Prentice, H.C. Contrasting changes in taxonomic, phylogenetic and functional diversity during a long-term succession: Insights into assembly processes. J. Ecol. 2013, 101, 857–866. [Google Scholar] [CrossRef]
- Feng, G.; Svenning, J.C.; Mi, X.; Jia, Q.; Rao, M.; Ren, H.; Bebber, D.P.; Ma, K. Anthropogenic disturbance shapes phylogenetic and functional tree community structure in a subtropical forest. For. Ecol. Manag. 2014, 313, 188–198. [Google Scholar] [CrossRef]
- Pastore, A.I.; Scherer, B.P. Changes in community phylogenetic structure in a North American forest chronosequence. Ecosphere 2016, 7, e01592. [Google Scholar] [CrossRef] [Green Version]
- Norden, N.; Letcher, S.G.; Boukili, V.; Swenson, N.G.; Chazdon, R. Demographic drivers of successional changes in phylogenetic structure across life-history stages in plant communities. Ecology 2012, 93, S70–S82. [Google Scholar] [CrossRef]
- Whitmore, T.C. Canopy gaps and the two major groups of forest trees. Ecology 1989, 70, 536–538. [Google Scholar] [CrossRef]
- Swenson, N.G.; Enquist, B.J. Ecological and evolutionary determinants of a key plant functional trait: Wood density and its community. Am. J. Bot. 2007, 94, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Queenborough, S.A.; Mazer, S.J.; Vamosi, S.M.; Garwood, N.C.; Valencia, R.; Freckleton, R.P. Seed mass, abundance and breeding system among tropical forest species: Do dioecious species exhibit compensatory reproduction or abundances? J. Ecol. 2009, 97, 555–566. [Google Scholar] [CrossRef]
- Uriarte, M.; Swenson, N.G.; Chazdon, R.L.; Comita, L.S.; Kress, W.J.; Erickson, D.; Forero-Montaña, J.; Zimmerman, J.K.; Thompson, J. Trait similarity, shared ancestry and the structure of neighbourhood interactions in a subtropical wet forest: Implications for community assembly. Ecol. Lett. 2010, 13, 1503–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moles, A.T.; Westoby, M. Cycle Seed size and plant strategy across the whole life. OIKOS 2006, 113, 91–105. [Google Scholar] [CrossRef]
- Westoby, M.; Leishman, M.; Lord, J. Comparative ecology of seed size and dispersal. Philos. Trans. R. Soc. Lond. Ser. B 1996, 351, 1309–1318. [Google Scholar] [CrossRef]
- Poorter, L.; Rose, S.A. Light-dependent changes in the relationship between seed mass and seedling traits: A meta-analysis for rain forest tree species. Oecologia 2005, 142, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Muller-Landau, H.C. The tolerance—Fecundity trade-off and the maintenance of diversity in seed size. Proc. Natl. Acad. Sci. USA 2010, 107, 4242–4247. [Google Scholar] [CrossRef] [PubMed]
- Coomes, D.A.; Grubb, P.J. Colonization, tolerance, competition and seed-size variation within functional groups. Trends Ecol. Evol. 2003, 18, 283–291. [Google Scholar] [CrossRef]
- Grubb, P.J.; Metcalfe, J.D. Adaptation and inertia in the Australian tropical lowland rain-forest flora: Contradictory trends in intergeneric and intrageneric comparisons of seed size in relation to light demand. Funct. Ecol. 1996, 10, 512–520. [Google Scholar] [CrossRef]
- Comita, L.S.; Uriarte, M.; Thompson, J.; Jonckheere, I.; Canham, C.D.; Zimmerman, J.K. Abiotic and biotic drivers of seedling survival in a hurricane-impacted tropical forest. J. Ecol. 2009, 97, 1346–1359. [Google Scholar] [CrossRef]
- Verdú, M.; Rey, P.J.; Alcántara, J.M.; Siles, G.; Valiente-Banuet, A. Phylogenetic signatures of facilitation and competition in successional communities. J. Ecol. 2009, 97, 1171–1180. [Google Scholar] [CrossRef] [Green Version]
- Novotny, V.; Basset, Y.; Miller, S.E.; Weiblen, G.D.; Bremer, B.; Cizek, L.; Drozd, P. Low host specificity of herbivorous insects in a tropical forest. Nature 2002, 416, 841–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, G.S.; Webb, C.O. Phylogenetic signal in plant pathogen-host range. Proc. Natl. Acad. Sci. USA 2007, 104, 4979–4983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahill, J.F.; Kembel, S.W.; Lamb, E.G.; Keddy, P.A. Does phylogenetic relatedness influence the strength of competition among vascular plants? Perspect. Plant Ecol. Evol. Syst. 2008, 10, 41–50. [Google Scholar] [CrossRef]
- Webb, C.O.; Gilbert, G.S.; Donoghue, M.J. Phylodiversity-dependent seedling mortality, size structure, and disease in a Bornean rain forest. Ecology 2006, 87, S123–S131. [Google Scholar] [CrossRef]
- Metz, M.R.; Sousa, W.P.; Valencia, R. Widespread density-dependent seedling mortality promotes species coexistence in a highly diverse Amazonian rain forest. Ecology 2010, 91, 3675–3685. [Google Scholar] [CrossRef] [PubMed]
- Paine, C.E.T.; Norden, N.; Chave, J.; Forget, P.-M.; Fortunel, C.; Dexter, K.G.; Baraloto, C. Phylogenetic density dependence and environmental filtering predict seedling mortality in a tropical forest. Ecol. Lett. 2012, 15, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Kress, W.J.; Erickson, D.L.; Swenson, N.G.; Thompson, J.; Uriarte, M.; Zimmerman, J.K. Advances in the use of DNA barcodes to build a community phylogeny for tropical trees in a Puerto Rican forest dynamics plot. PLoS ONE 2010, 5, e15409. [Google Scholar] [CrossRef] [PubMed]
- Boucher, D.H.; Vandermeer, J.H.; Mallona, M.A.; Zamora, N.; Perfecto, I. Resistance and resilience in a directly regenerating rainforest: Nicaraguan trees of the Vochysiaceae after Hurricane Joan. For. Ecol. Manag. 1994, 68, 127–136. [Google Scholar] [CrossRef]
- Walker, L.R. Tree damage and recovery from hurricane Hugo in Luquillo Experimental Forest, Puerto Rico. Biotropica 1991, 23, 379–385. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Kapos, V.; Varty, N.; Healey, J.R.; Tanner, E.V.J.; Kelly, D.L.; Dalling, J.W.; Burns, L.S.; Lee, D.; Sidrak, G. Hurricanes need not causes high mortality: The effects of Hurricane Gilbert on forest in Jamaica. J. Trop. Ecol. 1992, 8, 217–223. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Tanner, E.V.J.; Rich, P.M.; Goodland, T.C.R. Changes in light below the canopy of a Jamaican montane rainforest after a hurricane. J. Trop. Ecol. 1996, 12, 669–722. [Google Scholar] [CrossRef]
- Walker, L.R.; Lodge, D.J.; Guzman-Grajales, S.M.; Fetcher, N. Species-specific seedling responses to hurricane disturbance in a Puerto Rican rain forest. Biotropica 2003, 35, 472–485. [Google Scholar] [CrossRef]
- Norden, N.; Chazdon, R.L.; Chao, A.; Jiang, Y.-H.; Vilhez-Alvarado, B. Resilience of tropical rain forests: Tree community reassembly in secondary forests. Ecol. Lett. 2009, 12, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.; Brokaw, N.; Zimmerman, J.K.; Waide, R.B.; Everham, E.M.I.; Schaefer, D.A. Luquillo forest dynamics plot, Puerto Rico, United States. In Tropical Forest Diversisty Dynamism Findings from a Large-Scale Plot Network; University of Chicago Press: Chicago, IL, USA, 2004; pp. 540–550. [Google Scholar]
- Ostertag, R.; Scatena, F.N.; Silver, W.L. Forest floor decomposition following hurricane litter inputs in several Puerto Rican forests. Ecosystems 2003, 6, 261–273. [Google Scholar] [CrossRef]
- Thompson, J.; Brokaw, N.; Zimmerman, J.K.; Waide, R.B.; Everham, E.M., III; Lodge, D.J.; Taylor, C.M.; García-Montiel, D.; Fluet, M. Land use history, environment, and tree composition in a tropical forest. Ecol. Appl. 2002, 12, 1344–1363. [Google Scholar] [CrossRef]
- Zimmerman, J.K.; Mitchell Aide Bt, T.; Rosario, M.; Serrano, M.; Herrera, L. Effects of land management and a recent hurricane on forest structure and composition in the Luquillo Experimental Forest, Puerto Rico. For. Ecol. Manag. 1995, 77, 65–76. [Google Scholar] [CrossRef]
- Foster, D.; Swanson, F.; Aber, J.; Burke, I.; Brokaw, N.; Tilman, D.; Knapp, A. The importance of land-use legacies to ecology and conservation. Bioscience 2003, 53, 77–88. [Google Scholar] [CrossRef]
- Uriarte, M.; Rivera, L.W.; Zimmerman, J.K.; Aide, T.M.; Power, A.G.; Flecker, A. Effects of land use history on hurricane damage and recovery in a neotropical forest. Plant Ecol. 2004, 174, 49–58. [Google Scholar] [CrossRef]
- Uriarte, M.; Canham, C.D.; Thompson, J.; Zimmerman, J.K.; Brokaw, N. Seedling recruitment in a hurricane-driven tropical forest: Light limitation, density-dependence and the spatial distribution of parent trees. J. Ecol. 2005, 93, 291–304. [Google Scholar] [CrossRef]
- Erickson, D.L.; Jones, F.A.; Swenson, N.G.; Pei, N.; Bourg, N.; Chen, W.; Davies, S.J.; Ge, X.; Hao, Z.; Howe, R.W.; et al. Comparative evolutionary diversity and phylogenetic structure across multiple forest dynamics plots: A mega-phylogeny approach. Front. Genet. 2014, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cavender-Bares, J.; Kozak, K.H.; Fine, P.V.A.; Kembel, S.W. The merging of community ecology and phylogenetic biology. Ecol. Lett. 2009, 12, 693–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, C.O. Exploring the phylogenetic structure of ecological communities: An example for rain forest trees. Am. Nat. 2000, 156, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Kembel, S.W.; Hubbell, S.P. The phylogenetic structure of a neotropical forest tree community. Ecology 2006, 87, S86–S99. [Google Scholar] [CrossRef]
- Hardy, O.J. Testing the spatial phylogenetic structure of local communities: Statistical performances of different null models and test statistics on a locally neutral community. J. Ecol. 2008, 96, 914–926. [Google Scholar] [CrossRef]
- Kembel, S.W.; Ackerly, D.D.; Blomberg, S.P.; Cornwell, W.K.; Cowan, P.D.; Hel, M.R.; Morlon, H.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models using lme4. arXiv, 2014; arXiv:1046.5823. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 25 July 2018).
- Umaña, M.N.; Forero-Montaña, J.; Muscarella, R.; Nytch, C.J.; Thompson, J.; Uriarte, M.; Zimmerman, J.; Swenson, N.G. Interspecific functional convergence and divergence and intraspecific negative density dependence underlie the seed-to-seedling transition in tropical trees. Am. Nat. 2015, 187, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muscarella, R.; Uriarte, M.; Aide, T.M.; Erickson, D.L.; Forero-Montaña, J.; Kress, W.J.; Swenson, N.G.; Zimmerman, J.K. Functional convergence and phylogenetic divergence during secondary succession of subtropical wet forests in Puerto Rico. J. Veg. Sci. 2016, 27, 283–294. [Google Scholar] [CrossRef]
- Uriarte, M.; Muscarella, R.; Zimmerman, J.K. Environmental heterogeneity and biotic interactions mediate climate impacts on tropical forest regeneration. Glob. Chang. Biol. 2017, 24, e692–e704. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, R.; Gallery, R.E.; Gripenberg, S.; Gurr, S.J.; Narayan, L.; Addis, C.E.; Freckleton, R.P.; Lewis, O.T. Pathogens and insect herbivores drive rainforest plant diversity and composition. Nature 2014, 506, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.J. Plant diversity in tropical forests: A review of mechanisms of species coexistence. Oecologia 2002, 130, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.A.; Roger, A.; Courtois, E.A.; Jabot, F.; Norden, N.; Paine, C.E.T.; Baraloto, C.; Thébaud, C.; Chave, J. Shifts in species and phylogenetic diversity between sapling and tree communities indicate negative density dependence in a lowland rain forest. J. Ecol. 2010, 98, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Comita, L.S.; Hubbell, S.P.; Ma, K. Conspecific and phylogenetic density-dependent survival differs across life stages in a tropical forest. J. Ecol. 2015, 103, 957–966. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Comita, L.S.; Wright, S.J.; Swenson, N.G.; Zimmerman, J.K.; Mi, X.; Hao, Z.; Ye, W.; Hubbell, S.P.; Kress, W.J.; et al. Forest tree neighborhoods are structured more by negative conspecific density dependence than by interactions among closely related species. Ecography 2017, 41, 1114–1123. [Google Scholar] [CrossRef]
- Letcher, S.G.; Chazdon, R.L.; Andrade, A.C.S.; Bongers, F.; van Breugel, M.; Finegan, B.; Laurance, S.G.; Mesquita, R.C.G.; Martínez-Ramos, M.; Williamson, G.B. Phylogenetic community structure during succession: Evidence from three Neotropical forest sites. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 79–87. [Google Scholar] [CrossRef]
- Arnan, X.; Arcoverde, G.B.; Pie, M.R.; Ribeiro-Neto, J.D.; Leal, I.R. Increased anthropogenic disturbance and aridity reduce phylogenetic and functional diversity of ant communities in Caatinga dry forest. Sci. Total Environ. 2018, 631–632, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, J.K.; Comita, L.S.; Thompson, J.; Uriarte, M.; Brokaw, N. Patch dynamics and community metastability of a subtropical forest: Compound effects of natural disturbance and human land use. Landsc. Ecol. 2010, 25, 1099–1111. [Google Scholar] [CrossRef]
- Lugo, A.E.; Helmer, E. Emerging forests on abandoned land: Puerto Rico’s new forests. For. Ecol. Manag. 2004, 190, 145–161. [Google Scholar] [CrossRef] [Green Version]
- Uriarte, M.; Canham, C.D.; Thompson, J.; Zimmerman, J.K.; Murphy, L.; Sabat, A.M.; Fetcher, N.; Haines, B.L. Natural disturbance and human land use as determinants of tropical forest dynamics: Results from a forest simulator. Ecol. Monogr. 2009, 79, 423–443. [Google Scholar] [CrossRef]
- Zimmerman, J.K.; Everham, E.M.; Waide, R.B.; Lodge, D.J.; Taylor, C.M.; Brokaw, N.V.L. Responses of tree species to hurricane winds in subtropical wet forest in Puerto Rico: Implications for tropical tree life-histories. J. Ecol. 1994, 82, 911–922. [Google Scholar] [CrossRef]
- Comita, L.S.; Thompson, J.; Uriarte, M.; Jonckheere, I.; Canham, C.D.; Zimmerman, J.K. Interactive effects of land use history and natural disturbance on seedling dynamics in a subtropical forest. Ecol. Appl. 2010, 20, 1270–1284. [Google Scholar] [CrossRef] [PubMed]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Corlett, R.T. Climate change in the tropics: The end of the world as we know it? Biol. Conserv. 2012, 151, 22–25. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization). The State of the World’s Forests; FAO: Rome, Italy, 2017. [Google Scholar]
- Hu, T.; Smith, R.B. The Impact of hurricane Maria on the vegetation of Dominica and Puerto Rico using multispectral remote sensing. Remote Sens. 2018, 10, 827. [Google Scholar] [CrossRef]
- Liu, J.; Bowman, K.W.; Schimel, D.S.; Parazoo, N.C.; Jiang, Z.; Lee, M.; Bloom, A.A.; Wunch, D.; Frankenberg, C.; Sun, Y.; et al. Contrasting carbon cycle responses of the tropical continents to the 2015–2016 El Niño. Science 2017, 358, eaam5690. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Census Interval | ||||
|---|---|---|---|---|
| Model | 2000–2002 | 2002–2004 | 2004–2007 | 2007–2008 |
| Null (No neighbors) | 5064.9 | 42670.0 | 3210.5 | 3106.5 |
| Total neighbor density | 5063.1 | 4265.1 | 3210.7 | 3108.2 |
| Conspecific neighbor density | 5060.4 | 4261.0 | 3183.8 | 3108.5 |
| Phylogenetic density | 5049.8 | 4261.5 | 3182.3 | 3108.5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Comita, L.S.; Uriarte, M.; Forero-Montaña, J.; Kress, W.J.; Swenson, N.G.; Thompson, J.; Umaña, M.N.; Zimmerman, J.K. Changes in Phylogenetic Community Structure of the Seedling Layer Following Hurricane Disturbance in a Human-Impacted Tropical Forest. Forests 2018, 9, 556. https://doi.org/10.3390/f9090556
Comita LS, Uriarte M, Forero-Montaña J, Kress WJ, Swenson NG, Thompson J, Umaña MN, Zimmerman JK. Changes in Phylogenetic Community Structure of the Seedling Layer Following Hurricane Disturbance in a Human-Impacted Tropical Forest. Forests. 2018; 9(9):556. https://doi.org/10.3390/f9090556
Chicago/Turabian StyleComita, Liza S., María Uriarte, Jimena Forero-Montaña, W. John Kress, Nathan G. Swenson, Jill Thompson, María Natalia Umaña, and Jess K. Zimmerman. 2018. "Changes in Phylogenetic Community Structure of the Seedling Layer Following Hurricane Disturbance in a Human-Impacted Tropical Forest" Forests 9, no. 9: 556. https://doi.org/10.3390/f9090556
APA StyleComita, L. S., Uriarte, M., Forero-Montaña, J., Kress, W. J., Swenson, N. G., Thompson, J., Umaña, M. N., & Zimmerman, J. K. (2018). Changes in Phylogenetic Community Structure of the Seedling Layer Following Hurricane Disturbance in a Human-Impacted Tropical Forest. Forests, 9(9), 556. https://doi.org/10.3390/f9090556