Understanding Oxidative Stress in Aedes during Chikungunya and Dengue Virus Infections Using Integromics Analysis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquitoes Rearing and Virus Infection
2.2. NMR Spectroscopy
2.3. NMR Data Analysis
2.4. -Omics Data Collection
2.5. Integrated Pathway Annotation
3. Results
3.1. Infection of CHIKV and DENV in Mosquitoes
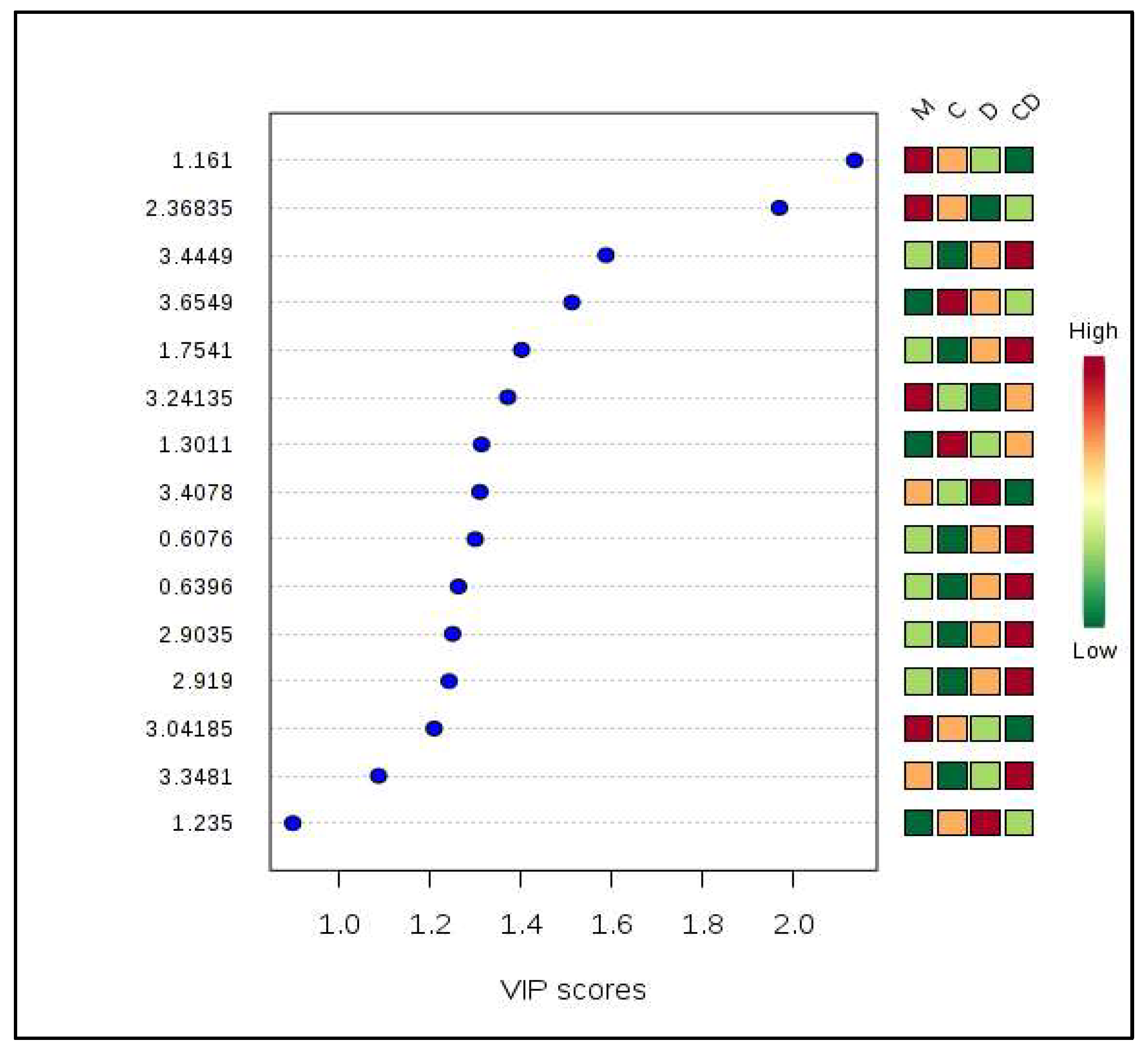
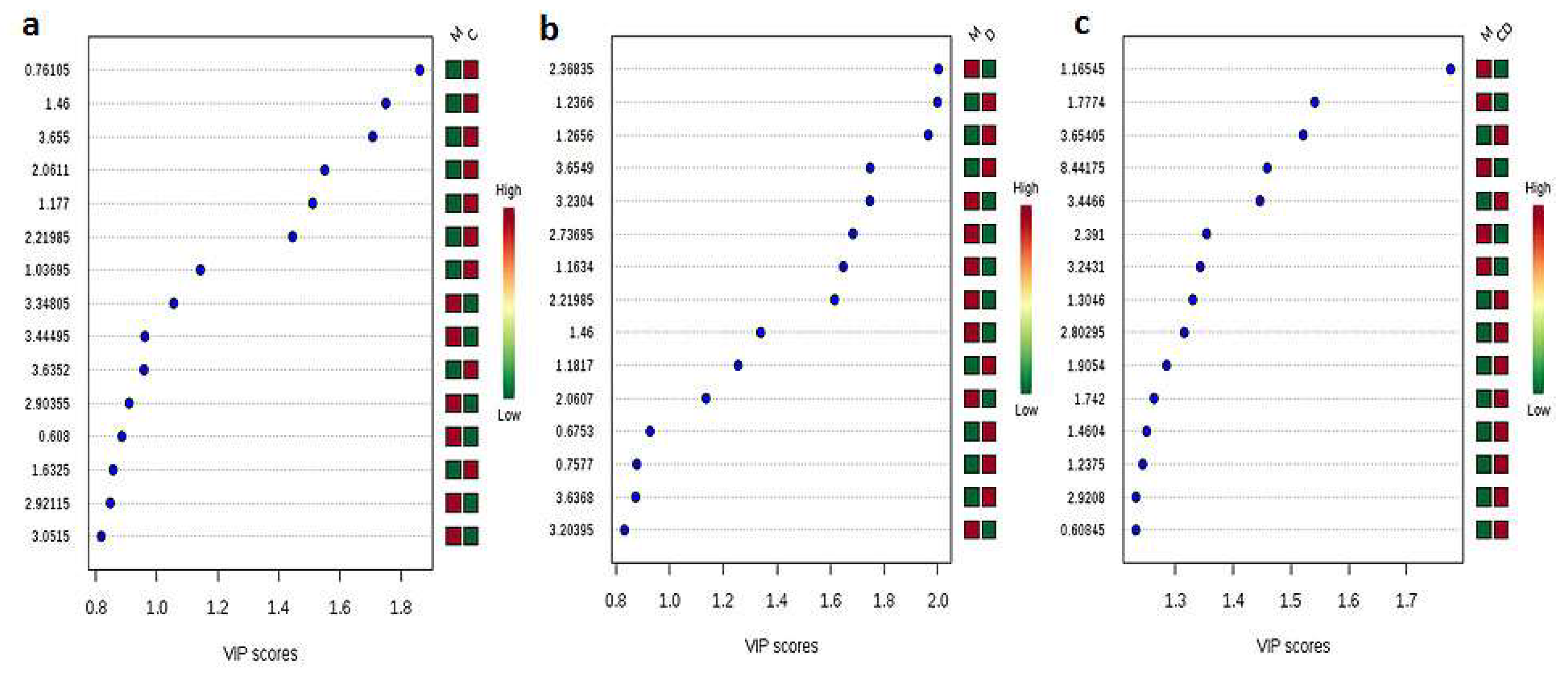
3.2. Metabolites Profiling of CHIKV, DENV, and CHIKV/DENV Co-Infected Hemolymph of Aedes Mosquitoes
3.3. Pair-Wise Metabolites Profiling of CHIKV, DENV, and Co-Infected Hemolymph of Aedes Mosquitoes in Comparison to Control Mosquitoes
3.4. Integration of -Omics Data
3.4.1. Data Selected for Study
3.4.2. Differentially Regulated Transcripts and Pathway Analysis
3.4.3. Differentially Regulated Proteins and Pathway Analysis
3.4.4. Integration of Pathways to Understand Mechanism of Oxidative Stress
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Harrington, L.C.; Edman, J.D.; Scott, T.W. Why do female Aedes aegypti (Diptera: Culicidae) feed preferentially and frequently on human blood? J. Med. Entomol. 2001, 38, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Woke, P.A.; Ally, M.S.; Rosenberger, C.R., Jr. The numbers of eggs developed related to the quantities of human blood ingested in Aedes aegypti (L.) (Diptera: Culicidae). Ann. Entomol. Soc. Am. 1956, 49, 435–441. [Google Scholar] [CrossRef]
- Saeaue, L.; Morales, N.P.; Komalamisra, N.; Vargas, R.E.M. Antioxidative systems defense against oxidative stress induced by blood meal in Aedes aegypti. Southeast Asian J. Trop. Med. Public Health 2011, 42, 542. [Google Scholar] [PubMed]
- Caragata, E.P.; Rezende, F.O.; Simões, T.C.; Moreira, L.A. Diet-induced nutritional stress and pathogen interference in wolbachia-infected Aedes aegypti. PLoS Negl. Trop. Dis. 2016, 10, e0005158. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J. The global emergence/resurgence of arboviral diseases as public health problems. Arch. Med. Res. 2002, 33, 330–342. [Google Scholar] [CrossRef]
- Furuya-Kanamori, L.; Liang, S.; Milinovich, G.; Magalhaes, R.J.S.; Clements, A.C.A.; Hu, W.; Brasil, P.; Frentiu, F.D.; Dunning, R.; Yakob, L. Co-distribution and co-infection of chikungunya and dengue viruses. BMC Infect. Dis. 2016, 16, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavrini, F.; Gaibani, P.; Pierro, A.M.; Rossini, G.; Landini, M.P.; Sambri, V. Chikungunya: An emerging and spreading arthropod-borne viral disease. J. Infect. Dev. Ctries. 2009, 3, 744–752. [Google Scholar] [PubMed]
- Chambers, T.J.; Hahn, C.S.; Galler, R.; Rice, C.M. Flavivirus genome organization, expression, and replication. Ann. Rev. Microbiol. 1990, 44, 649–688. [Google Scholar] [CrossRef] [PubMed]
- Kielian, M.; Helenius, A. Entry of alphaviruses. In The Togaviridae and Flaviviridae; Springer: Boston, MA, USA, 1986; pp. 91–119. [Google Scholar]
- Schmaljohn, A.L.; McClain, D. Alphaviruses (togaviridae) and flaviviruses (flaviviridae). In Medical Microbiology, 4th ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Solignat, M.; Gay, B.; Higgs, S.; Briant, L.; Devaux, C. Replication cycle of chikungunya: A re-emerging arbovirus. Virology 2009, 393, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Tuiskunen Back, A.; Lundkvist, A. Dengue viruses-an overview. Infect. Ecol. Epidemiol. 2013, 3, 19839. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, F.; Nie, L. Integrating multiple ‘omics’ analysis for microbial biology: Application and methodologies. Microbiology 2010, 156, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Gubb, E.; Matthiesen, R. Introduction to omics. Bioinform. Methods Clin. Res. 2010, 1–23. [Google Scholar]
- Beltran, P.M.J.; Federspiel, J.D.; Sheng, X.; Cristea, I.M. Proteomics and integrative omic approaches for understanding host-pathogen interactions and infectious diseases. Mol. Syst. Biol. 2017, 13, 922. [Google Scholar] [CrossRef] [PubMed]
- Shrinet, J.; Srivastava, P.; Sunil, S. Transcriptome analysis of Aedes aegypti in response to mono-infections and co-infections of dengue virus-2 and chikungunya virus. Biochem. Biophys. Res. Commun. 2017, 492, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Shrinet, J.; Shastri, J.S.; Gaind, R.; Bhavesh, N.S.; Sunil, S. Serum metabolomics analysis of patients with chikungunya and dengue mono/co-infections reveals distinct metabolite signatures in the three disease conditions. Sci. Rep. 2016, 6, 36833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, D.; Arensburger, P.; Atkinson, P.; Besansky, N.J.; Bruggner, R.V.; Butler, R.; Campbell, K.S.; Christophides, G.K.; Christley, S.; Dialynas, E. Vectorbase: A data resource for invertebrate vector genomics. Nucleic Acids Res. 2009, 37, D583–D587. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Mao, X.; Cai, T.; Luo, J.; Wei, L. Kobas server: A web-based platform for automated annotation and pathway identification. Nucleic Acids Res. 2006, 34, W720–W724. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, C.; Behura, S.K.; Lovin, D.D.; Harker, B.W.; Gomez-Machorro, C.; Mori, A.; Romero-Severson, J.; Severson, D.W. Comparative expression profiles of midgut genes in dengue virus refractory and susceptible Aedes aegypti across critical period for virus infection. PLoS ONE 2012, 7, e47350. [Google Scholar] [CrossRef] [PubMed]
- Luplertlop, N.; Surasombatpattana, P.; Patramool, S.; Dumas, E.; Wasinpiyamongkol, L.; Saune, L.; Hamel, R.; Bernard, E.; Sereno, D.; Thomas, F. Induction of a peptide with activity against a broad spectrum of pathogens in the Aedes aegypti salivary gland, following infection with dengue virus. PLoS Pathog. 2011, 7, e1001252. [Google Scholar] [CrossRef] [PubMed]
- Colpitts, T.M.; Cox, J.; Vanlandingham, D.L.; Feitosa, F.M.; Cheng, G.; Kurscheid, S.; Wang, P.; Krishnan, M.N.; Higgs, S.; Fikrig, E. Alterations in the Aedes aegypti transcriptome during infection with west nile, dengue and yellow fever viruses. PLoS Pathog. 2011, 7, e1002189. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, H.; Hanley, K.A.; Sundararajan, A.; Devitt, N.P.; Schilkey, F.D.; Hansen, I.A. Dengue virus serotype 2 infection alters midgut and carcass gene expression in the asian tiger mosquito, Aedes albopictus. PLoS ONE 2017, 12, e0171345. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.; Ramirez, J.L.; Dimopoulos, G. Dengue virus infection of the Aedes aegypti salivary gland and chemosensory apparatus induces genes that modulate infection and blood-feeding behavior. PLoS Pathog. 2012, 8, e1002631. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef] [PubMed]
- Bonizzoni, M.; Dunn, W.A.; Campbell, C.L.; Olson, K.E.; Marinotti, O.; James, A.A. Complex modulation of the Aedes aegypti transcriptome in response to dengue virus infection. PLoS ONE 2012, 7, e50512. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.; Dimopoulos, G. Dengue virus inhibits immune responses in Aedes aegypti cells. PLoS ONE 2010, 5, e10678. [Google Scholar] [CrossRef] [PubMed]
- Behura, S.K.; Gomez-Machorro, C.; Harker, B.W.; Lovin, D.D.; Hemme, R.R.; Mori, A.; Romero-Severson, J.; Severson, D.W. Global cross-talk of genes of the mosquito Aedes aegypti in response to dengue virus infection. PLoS Negl. Trop. Dis. 2011, 5, e1385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zheng, X.; Wu, Y.; Gan, M.; He, A.; Li, Z.; Zhang, D.; Wu, X.; Zhan, X. Differential proteomics of Aedes albopictus salivary gland, midgut and c6/36 cell induced by dengue virus infection. Virology 2013, 444, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Tchankouo-Nguetcheu, S.; Khun, H.; Pincet, L.; Roux, P.; Bahut, M.; Huerre, M.; Guette, C.; Choumet, V. Differential protein modulation in midguts of Aedes aegypti infected with chikungunya and dengue 2 viruses. PLoS ONE 2010, 5, e13149. [Google Scholar] [CrossRef] [PubMed]
- Tchankouo-Nguetcheu, S.; Bourguet, E.; Lenormand, P.; Rousselle, J.-C.; Namane, A.; Choumet, V. Infection by chikungunya virus modulates the expression of several proteins in Aedes aegypti salivary glands. Parasites Vectors 2012, 5, 264. [Google Scholar] [CrossRef] [PubMed]
- Chisenhall, D.M.; Christofferson, R.C.; McCracken, M.K.; Johnson, A.-M.F.; Londono-Renteria, B.; Mores, C.N. Infection with dengue-2 virus alters proteins in naturally expectorated saliva of NOx mosquitoes. Parasites Vectors 2014, 7, 252. [Google Scholar] [CrossRef] [PubMed]
- Chisenhall, D.M.; Londono, B.L.; Christofferson, R.C.; McCracken, M.K.; Mores, C.N. Effect of dengue-2 virus infection on protein expression in the salivary glands of Aedes aegypti mosquitoes. Am. J. Trop. Med. Hyg. 2014, 90, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Patramool, S.; Surasombatpattana, P.; Luplertlop, N.; Seveno, M.; Choumet, V.; Thomas, F.; Misse, D. Proteomic analysis of an Aedes albopictus cell line infected with dengue serotypes 1 and 3 viruses. Parasites Vectors 2011, 4, 138. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.H.; Chu, J.J.H. Proteomics profiling of chikungunya-infected Aedes albopictus c6/36 cells reveal important mosquito cell factors in virus replication. PLoS Negl. Trop. Dis. 2015, 9, e0003544. [Google Scholar] [CrossRef] [PubMed]
- El-Bacha, T.; Da Poian, A.T. Virus-induced changes in mitochondrial bioenergetics as potential targets for therapy. Int. J. Biochem. Cell Biol. 2013, 45, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Ringrose, J.H.; Jeeninga, R.E.; Berkhout, B.; Speijer, D. Proteomic studies reveal coordinated changes in t-cell expression patterns upon infection with human immunodeficiency virus type 1. J. Virol. 2008, 82, 4320–4330. [Google Scholar] [CrossRef] [PubMed]
- Thai, M.; Graham, N.A.; Braas, D.; Nehil, M.; Komisopoulou, E.; Kurdistani, S.K.; McCormick, F.; Graeber, T.G.; Christofk, H.R. Adenovirus e4orf1-induced myc activation promotes host cell anabolic glucose metabolism and virus replication. Cell Metab. 2014, 19, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Besancon, F.; Just, J.; Bourgeade, M.F.; Van Weyenbergh, J.; Solomon, D.; Guillozo, H.; Wietzerbin, J.; Cayre, Y.E. Hiv-1 p17 and ifn-î³ both induce fructose 1, 6-bisposphatase. J. Interferon Cytokine Res. 1997, 17, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Coombs, K.; Berkhout, B. Global Host Proteomic Responses to Virus Infection; Frontiers E-books: Lausanne Switzerland, 2012. [Google Scholar]
- Deng, L.; Shoji, I.; Ogawa, W.; Kaneda, S.; Soga, T.; Jiang, D.-P.; Ide, Y.-H.; Hotta, H. Hepatitis c virus infection promotes hepatic gluconeogenesis through an ns5a-mediated, foxo1-dependent pathway. J. Virol. 2011, 85, 8556–8568. [Google Scholar] [CrossRef] [PubMed]
- Klemperer, H. Glucose breakdown in chick embryo cells infected with influenza virus. Virology 1961, 13, 68–77. [Google Scholar] [CrossRef]
- Ritter, J.B.; Wahl, A.S.; Freund, S.; Genzel, Y.; Reichl, U. Metabolic effects of influenza virus infection in cultured animal cells: Intra-and extracellular metabolite profiling. BMC Syst. Biol. 2010, 4, 61. [Google Scholar] [CrossRef] [PubMed]
- Janke, R.; Genzel, Y.; Wetzel, M.; Reichl, U. Effect of influenza virus infection on key metabolic enzyme activities in mdck cells. BMC Proc. 2011, 5, P129. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.; Riley, C.; Isaac, G.; Hopf-Jannasch, A.S.; Moore, R.J.; Weitz, K.W.; Pasa-Tolic, L.; Metz, T.O.; Adamec, J.; Kuhn, R.J. Dengue virus infection perturbs lipid homeostasis in infected mosquito cells. PLoS Pathog. 2012, 8, e1002584. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, C.; LeBreton, A.; Ng, C.Y.; Lim, J.Y.H.; Liu, W.; Vasudevan, S.; Labow, M.; Gu, F.; Gaither, L.A. Cholesterol biosynthesis modulation regulates dengue viral replication. Virology 2009, 389, 8–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kielian, M.; Chanel-Vos, C.; Liao, M. Alphavirus entry and membrane fusion. Viruses 2010, 2, 796–825. [Google Scholar] [CrossRef] [PubMed]
- Munger, J.; Bennett, B.D.; Parikh, A.; Feng, X.-J.; McArdle, J.; Rabitz, H.A.; Shenk, T.; Rabinowitz, J.D. Systems-level metabolic flux profiling identifies fatty acid synthesis as a target for antiviral therapy. Nat. Biotechnol. 2008, 26, 1179. [Google Scholar] [CrossRef] [PubMed]
- Noor, E.; Eden, E.; Milo, R.; Alon, U. Central carbon metabolism as a minimal biochemical walk between precursors for biomass and energy. Mol. Cell 2010, 39, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Cossart, P.; Toledo-Arana, A. Listeria monocytogenes, a unique model in infection biology: An overview. Microbes Infect. 2008, 10, 1041–1050. [Google Scholar] [CrossRef] [PubMed]
- Beuzón, C.R.; Méresse, S.; Unsworth, K.E.; Ruíz-Albert, J.; Garvis, S.; Waterman, S.R.; Ryder, T.A.; Boucrot, E.; Holden, D.W. Salmonella maintains the integrity of its intracellular vacuole through the action of sifa. EMBO J. 2000, 19, 3235–3249. [Google Scholar] [CrossRef] [PubMed]
- Ray, K.; Marteyn, B.; Sansonetti, P.J.; Tang, C.M. Life on the inside: The intracellular lifestyle of cytosolic bacteria. Nat. Rev. Microbiol. 2009, 7, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Eisenreich, W.; Dandekar, T.; Heesemann, J.; Goebel, W. Carbon metabolism of intracellular bacterial pathogens and possible links to virulence. Nat. Rev. Microbiol. 2010, 8, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Geiler, J.; Michaelis, M.; Naczk, P.; Leutz, A.; Langer, K.; Doerr, H.-W.; Cinatl, J. N-acetyl-l-cysteine (nac) inhibits virus replication and expression of pro-inflammatory molecules in a549 cells infected with highly pathogenic h5n1 influenza a virus. Biochem. Pharmacol. 2010, 79, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Makipour, K.; Friedenberg, F.K. The potential role of N-acetylcysteine for the treatment of helicobacter pylori. J. Clin. Gastroenterol. 2011, 45, 841. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Yan, Q.; Liu, R.-H.; Zhang, L. Preventive and therapeutic effect of N-acetyl-l-cysteine on infection-associated preterm labor in mice. Asian Pac. J. Trop. Med. 2016, 9, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Ripps, H.; Shen, W. Taurine: A very essential amino acid. Mol. Vis. 2012, 18, 2673. [Google Scholar] [PubMed]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. Ros and ros-mediated cellular signaling. Oxid. Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Zhou, G.; Wu, J.; Bian, G.; Lu, P.; Raikhel, A.S.; Xi, Z. Wolbachia induces reactive oxygen species (ros)-dependent activation of the toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2012, 109, E23–E31. [Google Scholar] [CrossRef] [PubMed]
- Zele, F.; Nicot, A.; Berthomieu, A.; Weill, M.; Duron, O.; Rivero, A. Wolbachia increases susceptibility to plasmodium infection in a natural system. Proc. R. Soc. B 2014, 281, 20132837. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Oberley, L.W.; Murhammer, D.W. Evidence of oxidative stress following the viral infection of two lepidopteran insect cell lines. Free Radic. Biol. Med. 2001, 31, 1448–1455. [Google Scholar] [CrossRef]
- Paes, M.C.; Oliveira, M.B.; Oliveira, P.L. Hydrogen peroxide detoxification in the midgut of the blood-sucking insect, rhodnius prolixus. Arch. Insect Biochem. Physiol. 2001, 48, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.H.M.; Talyuli, O.A.; Goncalves, R.L.; Paiva-Silva, G.O.; Sorgine, M.H.F.; Alvarenga, P.H.; Oliveira, P.L. Catalase protects NOx from oxidative stress and increases midgut infection prevalence of dengue but not zika. PLoS Negl. Trop. Dis. 2017, 11, e0005525. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.H.M.; Gonçalves, R.L.; Lara, F.A.; Dias, F.A.; Gandara, A.C.P.; Menna-Barreto, R.F.; Edwards, M.C.; Laurindo, F.R.M.; Silva-Neto, M.A.C.; Sorgine, M.H.F. Blood meal-derived heme decreases ros levels in the midgut of Aedes aegypti and allows proliferation of intestinal microbiota. PLoS Pathog. 2011, 7, e1001320. [Google Scholar] [CrossRef] [PubMed]
- Penin, F.; Dubuisson, J.; Rey, F.A.; Moradpour, D.; Pawlotsky, J.M. Structural biology of hepatitis C virus. Hepatology 2004, 39, 5–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Peaks (ppm) | Chi-Squared | p-Value | −log 10(p) | FDR |
|---|---|---|---|---|---|
| 1 | 3.697 | 6.69 | 0.08 | 1.08 | 0.65 |
| 2 | 3.655 | 6.59 | 0.09 | 1.06 | 0.65 |
| 3 | 1.161 | 6.49 | 0.09 | 1.04 | 0.65 |
| 4 | 1.635 | 6.23 | 0.1 | 1 | 0.65 |
| 5 | 2.061 | 6.08 | 0.11 | 0.97 | 0.65 |
| 6 | 1.713 | 5.82 | 0.12 | 0.92 | 0.65 |
| 7 | 2.368 | 5.62 | 0.13 | 0.88 | 0.65 |
| Sample | Pathway Name | Total | Hits | p-Value | Compounds |
|---|---|---|---|---|---|
| Global metabolite profiling | Taurine and hypotaurine metabolism | 6 | 3 | 1.88 × 10−5 | l-Cysteine, taurine, hypotaurine |
| Thiamine metabolism | 6 | 1 | 0.063562 | l-Cysteine | |
| CHIKV (CM) | Taurine and hypotaurine metabolism | 6 | 2 | 0.0015716 | l-Cysteine, hypotaurine |
| Thiamine metabolism | 6 | 1 | 0.063562 | l-Cysteine | |
| DENV (DM) | Glycosylphosphatidylinositol-anchor biosynthesis | 11 | 1 | 0.093946 | Ethanol |
| Glycolysis or Gluconeogenesis | 25 | 1 | 0.20213 | Ethanol | |
| Co-infection (CD) | Taurine and hypotaurine metabolism | 6 | 1 | 0.063562 | Taurine |
| Pantothenate and CoA biosynthesis | 12 | 1 | 0.12343 | Dihydrouracil |
| S. No. | Organism | Body Part/Source | Technique | Virus(es) | References |
|---|---|---|---|---|---|
| Transcriptomics | |||||
| 1 | Aedes aegypti | Midgut | cDNA Microarray | DENV2 | [20] |
| 2 | Aedes aegypti | Salivary gland | DGE | DENV2 | [21] |
| 3 | Aedes aegypti | Whole mosquito | Microarray | DENV2 | [22] |
| 4 | Aedes albopictus | Midgut | RNA-seq | DENV2 | [23] |
| Carcass | RNA-seq | ||||
| 5 | Aedes aegypti | Salivary gland | Microarray | DENV2 | [24] |
| Chemosensory apparatus | |||||
| 6 | Aedes aegypti | Midgut | Microarray | DENV2 | [25] |
| Carcass | |||||
| 7 | Aedes aegypti | Midgut | RNA-seq | DENV2 | [26] |
| Salivary gland | |||||
| Carcass | |||||
| 8 | Aedes aegypti | Aag2 cells | Microarray | DENV2 | [27] |
| 9 | Aedes aegypti | Whole mosquito | Microarray | DENV2 | [28] |
| 10 | Aedes aegypti | Whole mosquito | RNA-seq | CHIKV and DENV2 | [16] |
| Proteomics | |||||
| 1 | Aedes albopictus | Salivary gland | 2D-DIGE; MALDI TOF/TOF | DENV2 | [29] |
| Midgut | DENV2 | ||||
| C6/36 | DENV2 | ||||
| 2 | Aedes aegypti | Midgut | 2D-DIGE | CHIKV and DENV2 infection | [30] |
| 3 | Aedes aegypti | Salivary gland | 2D-DIGE; MALDI TOF/TOF | CHIKV | [31] |
| 4 | Aedes aegypti | Saliva | 2-D; Nano LC-MS/MS | DENV2 | [32] |
| 5 | Aedes aegypti | Salivary glands | 2-D; Nano LC-MS/MS | DENV2 | [33] |
| 6 | Aedes albopictus | C6/36 | 2D-DIGE | DENV1; DENV3 | [34] |
| 7 | Aedes albopictus | C6/36 | 2D-PAGE; Maldi TOF/TOF | CHIKV | [35] |
| 8 | Aedes aegypti | Whole body | LC-MS/MS | CHIKV and DENV2 | Data link: 10.6084/m9.figshare.5746134 |
| Metabolomics | |||||
| 1 | Aedes aegypti | Hemolymph | NMR | CHIKV and DENV2 | This study |
| Pathways | Input Number | Total Number of Transcripts/Genes | p-Value |
|---|---|---|---|
| Metabolic pathways | 85 | 929 | 56 × 10−21 |
| Oxidative phosphorylation | 34 | 142 | 1.36 × 10−19 |
| Biosynthesis of amino acids | 12 | 57 | 3.14 × 10−7 |
| Lysosome | 12 | 81 | 8.21 × 10−6 |
| Citrate cycle (TCA cycle) | 8 | 34 | 1.50 × 10−5 |
| Carbon metabolism | 11 | 98 | 0.00019 |
| 2-Oxocarboxylic acid metabolism | 4 | 16 | 0.001865 |
| Glycosphingolipid biosynthesis—globo series | 3 | 9 | 0.003683 |
| Arginine and proline metabolism | 5 | 37 | 0.005454 |
| Glycine, serine, and threonine metabolism | 5 | 38 | 0.006036 |
| Fatty acid biosynthesis | 3 | 12 | 0.007168 |
| Caffeine metabolism | 3 | 13 | 0.008645 |
| Arginine biosynthesis | 3 | 14 | 0.010289 |
| One carbon pool by folate | 3 | 14 | 0.010289 |
| Glycosaminoglycan degradation | 3 | 15 | 0.0121 |
| Other glycan degradation | 3 | 19 | 0.021077 |
| Pyruvate metabolism | 4 | 37 | 0.024909 |
| Glycosphingolipid biosynthesis—ganglio series | 2 | 8 | 0.028854 |
| Tryptophan metabolism | 3 | 26 | 0.043514 |
| Pathways | Input Number | Total Number of Transcripts/Genes | p-Value |
|---|---|---|---|
| Glycine, serine, and threonine metabolism | 5 | 38 | 0.001091 |
| One carbon pool by folate | 3 | 14 | 0.00343 |
| Metabolic pathways | 27 | 929 | 0.017225 |
| Lysosome | 5 | 81 | 0.020921 |
| Biosynthesis of amino acids | 4 | 57 | 0.025549 |
| Nitrogen metabolism | 2 | 13 | 0.030062 |
| Phototransduction—fly | 3 | 35 | 0.032108 |
| FoxO signaling pathway | 4 | 62 | 0.032892 |
| Arginine and proline metabolism | 3 | 37 | 0.03662 |
| Carbon metabolism | 5 | 98 | 0.041077 |
| Starch and sucrose metabolism | 3 | 40 | 0.043962 |
| Pathways | Input Number | Total Number of Transcripts/Genes | p-Value |
|---|---|---|---|
| Metabolic pathways | 111 | 929 | 2.84 × 10−12 |
| Oxidative phosphorylation | 34 | 142 | 7.48× 10−11 |
| Biosynthesis of amino acids | 15 | 57 | 72 × 10−6 |
| Pyruvate metabolism | 11 | 37 | 3.94× 10−5 |
| Carbon metabolism | 18 | 98 | 3 × 10−5 |
| Citrate cycle (TCA cycle) | 10 | 34 | 9.52 × 10−5 |
| One carbon pool by folate | 6 | 14 | 0.000499 |
| Glycolysis/gluconeogenesis | 9 | 44 | 0.001988 |
| Glycine, serine, and threonine metabolism | 8 | 38 | 0.00299 |
| Lysosome | 12 | 81 | 0.004325 |
| FoxO signaling pathway | 10 | 62 | 0.00524 |
| Neuroactive ligand–receptor interaction | 8 | 51 | 0.013724 |
| 2-Oxocarboxylic acid metabolism | 4 | 16 | 0.020663 |
| Glyoxylate and dicarboxylate metabolism | 5 | 30 | 0.038834 |
| Fatty acid biosynthesis | 3 | 12 | 0.04422 |
| Histidine metabolism | 3 | 12 | 0.04422 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shrinet, J.; Bhavesh, N.S.; Sunil, S. Understanding Oxidative Stress in Aedes during Chikungunya and Dengue Virus Infections Using Integromics Analysis. Viruses 2018, 10, 314. https://doi.org/10.3390/v10060314
Shrinet J, Bhavesh NS, Sunil S. Understanding Oxidative Stress in Aedes during Chikungunya and Dengue Virus Infections Using Integromics Analysis. Viruses. 2018; 10(6):314. https://doi.org/10.3390/v10060314
Chicago/Turabian StyleShrinet, Jatin, Neel Sarovar Bhavesh, and Sujatha Sunil. 2018. "Understanding Oxidative Stress in Aedes during Chikungunya and Dengue Virus Infections Using Integromics Analysis" Viruses 10, no. 6: 314. https://doi.org/10.3390/v10060314
APA StyleShrinet, J., Bhavesh, N. S., & Sunil, S. (2018). Understanding Oxidative Stress in Aedes during Chikungunya and Dengue Virus Infections Using Integromics Analysis. Viruses, 10(6), 314. https://doi.org/10.3390/v10060314