Genetic Characterization and Phylogenetic Analysis of Small Ruminant Lentiviruses Detected in Spanish Assaf Sheep with Different Mammary Lesions

 ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Samples
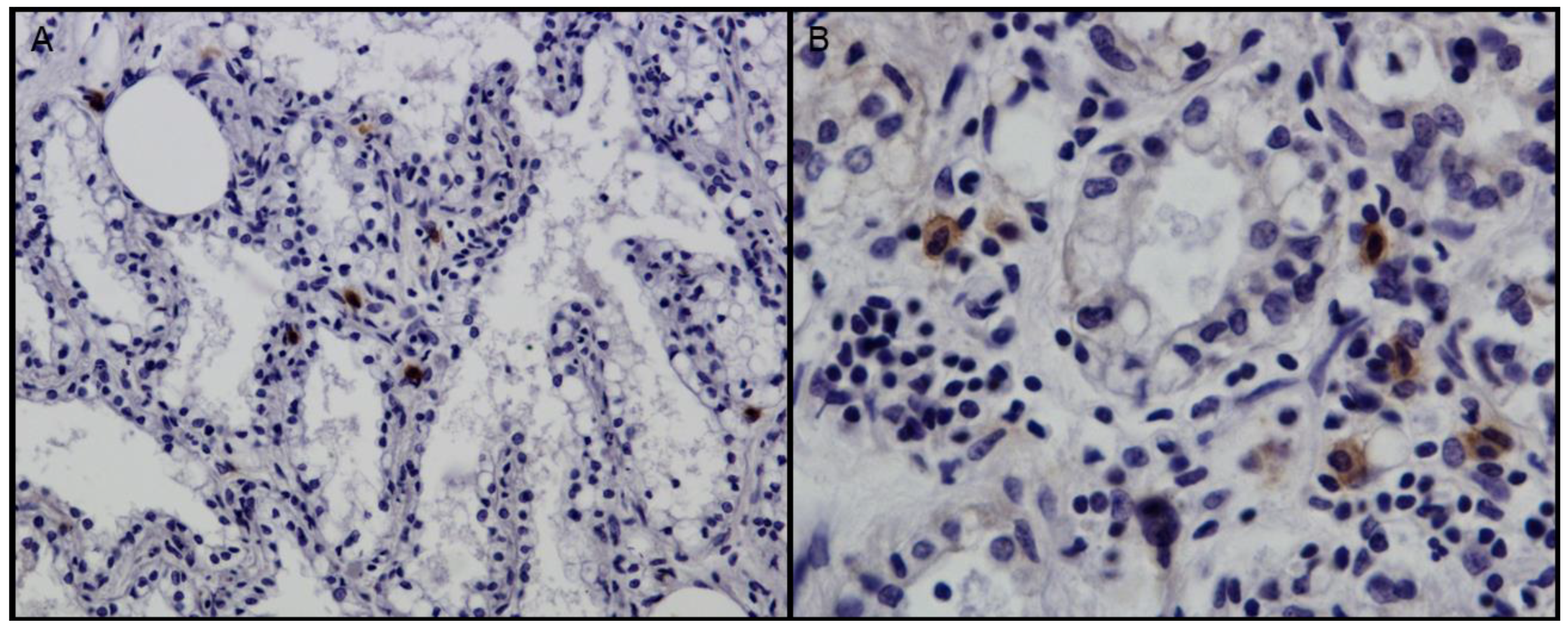
2.2. Histopathology and Immunohistochemistry
2.3. PCR
2.4. Sequence Analysis
3. Results
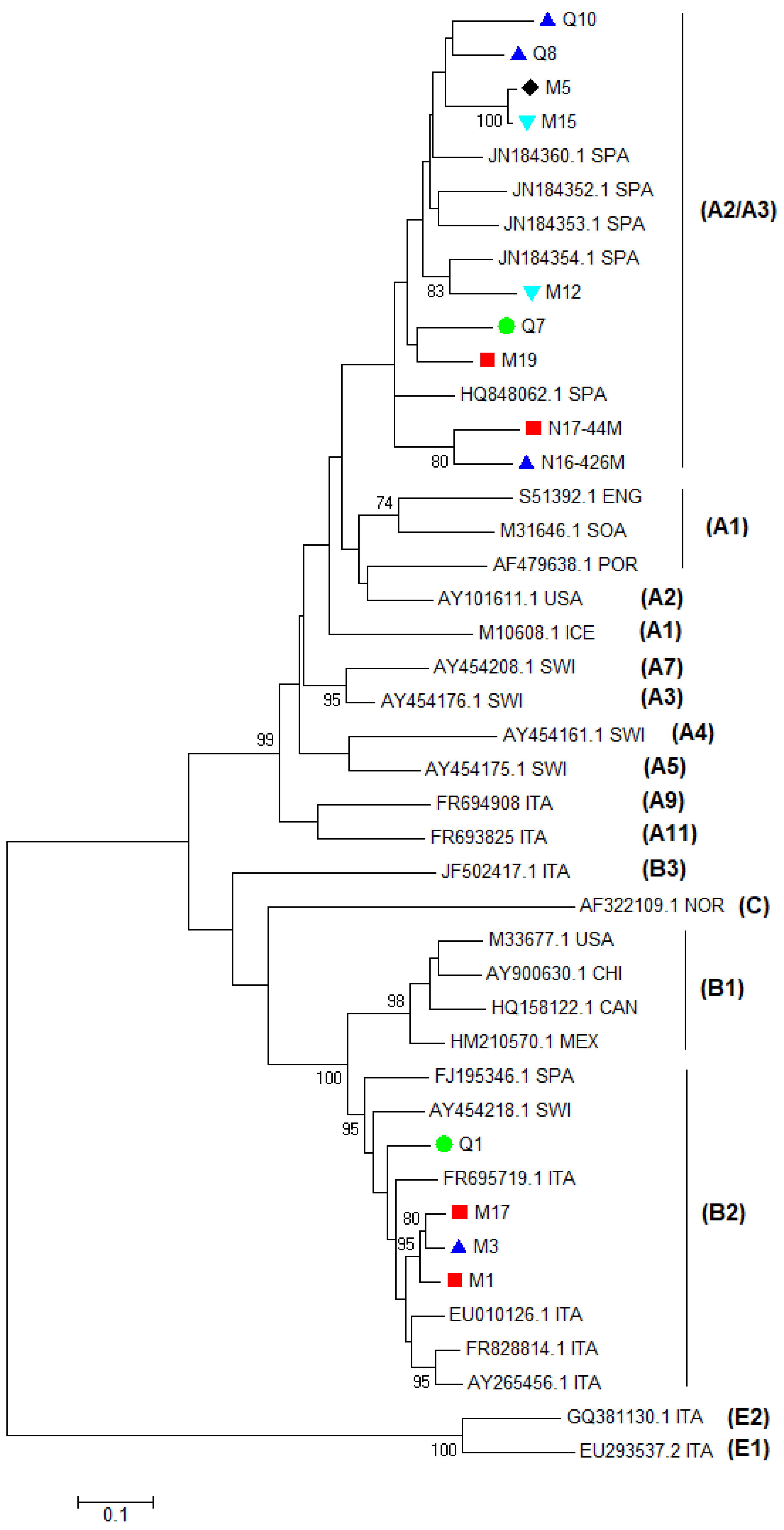
3.1. Analysis of Gag Sequences
3.2. Analysis of LTR Sequences
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Peterhans, E.; Greenland, T.; Badiola, J.; Harkiss, G.; Bertoni, G.; Amorena, B.; Eliaszewicz, M.; Juste, R.A.; Krassnig, R.; Lafont, J.P.; et al. Routes of transmission and consequences of small ruminant lentiviruses (SRLVs) infection and eradication schemes. Vet. Res. 2004, 35, 257–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preziuso, S.; Renzoni, G.; Allen, T.E.; Taccini, E.; Rossi, G.; DeMartini, J.C.; Braca, G. Colostral transmission of maedi visna virus: Sites of viral entry in lambs born from experimentally infected ewes. Vet. Microbiol. 2004, 104, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Shah, C.; Böni, J.; Huder, J.B.; Vogt, H.R.; Mühlherr, J.; Zanoni, R.; Miserez, R.; Lutz, H.; Schüpbach, J. Phylogenetic analysis and reclassification of caprine and ovine lentiviruses based on 104 new isolates: Evidence for regular sheep-to-goat transmission and worldwide propagation through livestock trade. Virology 2004, 319, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Grego, E.; Bertolotti, L.; Quasso, A.; Profiti, M.; Lacerenza, D.; Muz, D.; Rosati, S. Genetic characterization of small ruminant lentivirus in Italian mixed flocks: Evidence for a novel genotype circulating in a local goat population. J. Gen. Virol. 2007, 88, 3423–3427. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, H.; Reina, R.; Amorena, B.; de Andrés, D.; Martínez, H.A. Small ruminant lentiviruses: Genetic variability; tropism and diagnosis. Viruses 2013, 5, 1175–1207. [Google Scholar] [CrossRef] [PubMed]
- Leroux, C.; Chastang, J.; Greenland, T.; Mornex, J.F. Genomic heterogeneity of small ruminant lentiviruses: Existence of heterogeneous populations in sheep and of the same lentiviral genotypes in sheep and goats. Arch. Virol. 1997, 142, 1125–1137. [Google Scholar] [CrossRef] [PubMed]
- Minguijón, E.; Reina, R.; Pérez, M.; Polledo, L.; Villoria, M.; Ramírez, H.; Leginagoikoa, I.; Badiola, J.J.; García-Marín, J.F.; de Andrés, D.; et al. Small ruminant lentivirus infections and diseases. Vet. Microbiol. 2015, 181, 75–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lujan, L.; García Marin, J.F.; Fernandez de Luco, D.; Vargas, A.; Badiola, J.J. Pathological changes in the lungs and mammary glands of sheep and their relationship with maedi-visna infection. Vet. Rec. 1991, 129, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Benavides, J.; Fuertes, M.; García-Pariente, C.; Otaola, J.; Delgado, L.; Giraldez, J.; García Marín, J.F.; Carmen Ferreras, M.; Pérez, V. Impact of maedi-visna in intensively managed dairy sheep. Vet. J. 2013, 197, 607–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gayo, E.; Polledo, L.; Balseiro, A.; Martínez, C.P.; García Iglesias, M.J.; Preziuso, S.; Rossi, G.; García Marín, J.F. Inflammatory Lesion Patterns in Target Organs of Visna/Maedi in Sheep and their Significance in the Pathogenesis and Diagnosis of the Infection. J. Comp. Pathol. 2018, 159, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Anderson, B.C.; Bulgin, M.S.; Adams, S.; Duelke, B. Firm udder in periparturient ewes with lymphocytic accumulations; retrovirus infection; and milk unavailable at the teat. J. Am. Vet. Med. Assoc. 1985, 186, 391–393. [Google Scholar] [PubMed]
- Ryan, D.P.; Greenwood, P.L.; Nicholls, P.J. Effect of caprine arthritis-encephalitis virus infection on milk cell count and N-acetyl-beta-glucosaminidase activity in dairy goats. J. Dairy Res. 1993, 60, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Navalon, B.; Peris, C.; Gomez, E.A.; Peris, B.; Roche, M.L.; Caballero, C.; Goyena, E.; Berriatua, E. Quantitative estimation of the impact of caprine arthritis encephalitis virus infection on milk production by dairy goats. Vet. J. 2013, 197, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Sihvonen, L.; Nuotio, L.; Rikula, U.; Hirvelä-Koski, V.; Kokkonen, U. Preventing the spread of maedi-visna in sheep through a voluntary control programme in Finland. Prev. Vet. Med. 2000, 47, 213–220. [Google Scholar] [CrossRef]
- Tavella, A.; Bettini, A.; Ceol, M.; Zambotto, P.; Stifter, E.; Kusstatscher, N.; Lombardi, R.; Nardeli, S.; Beato, M.S.; Capello, K.; et al. Achievements of an eradication programme against caprine arthritis encephalitis virus in South Tyrol; Italy. Vet. Rec. 2018, 182, 51. [Google Scholar] [CrossRef] [PubMed]
- Deubelbeiss, M.; Blatti-Cardinaux, L.; Zahno, M.L.; Zanoni, R.; Vogt, H.R.; Posthaus, H.; Bertoni, G. Characterization of small ruminant lentivirus A4 subtype isolates and assessment of their pathogenic potential in naturally infected goats. Virol. J. 2014, 11, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, M.; Biescas, E.; Reina, R.; Glaria, I.; Marín, B.; Marquina, A.; Salazar, E.; Álvarez, N.; de Andrés, D.; Fantova, E.; et al. Small ruminant lentivirus-induced arthritis: Clinicopathologic findings in sheep infected by a highly replicative SRLV B2 genotype. Vet. Pathol. 2015, 52, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulou, K.; Poutahidis, T.; Brellou, G.D.; Greenland, T.; Vlemmas, I. A deletion in the R region of long terminal repeats in small ruminant lentiviruses is associated with decreased pathology in the lung. Vet. J. 2008, 175, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Preziuso, S.; Taccini, E.; Rossi, G.; Renzoni, G.; Braca, G. Experimental Maedi Visna Virus Infection in sheep: A morphological; immunohistochemical and PCR study after three years of infection. Eur. J. Histochem. 2003, 47, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Ryan, S.; Tiley, L.; McConnell, I.; Blacklaws, B. Infection of dendritic cells by the Maedi-Visna lentivirus. J. Virol. 2000, 74, 10096–10103. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Glaria, I.; Reina, R.; Ramírez, H.; de Andrés, X.; Crespo, H.; Jauregui, P.; Salazar, E.; Luján, L.; Pérez, M.M.; Benavides, J.; et al. Visna/Maedi virus genetic characterization and serological diagnosis of infection in sheep from a neurological outbreak. Vet. Microbiol. 2012, 155, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Sutton, K.A.; Lin, C.T.; Harkiss, G.D.; McConnell, I.; Sargan, D.R. Regulation of the long terminal repeat in visna virus by a transcription factor related to the AML/PEBP2/CBF superfamily. Virology 1997, 229, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Kuhar, U.; Barlic-Maganja, D.; Grom, J. Phylogenetic analysis of small ruminant lentiviruses detected in Slovenia. Vet. Microbiol. 2012, 162, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Olech, M.; Rachid, A.; Croise, B.; Kuźmak, J.; Valas, S. Genetic and antigenic characterization of small ruminant lentiviruses circulating in Poland. Virus Res. 2012, 163, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Fras, M.; Leboeuf, A.; Labrie, F.M.; Laurin, M.A.; Singh Sohal, J.; L’Homme, Y. Phylogenetic analysis of small ruminant lentiviruses in mixed flocks: Multiple evidence of dual infection and natural transmission of types A2 and B1 between sheep and goats. Infect. Genet. Evol. 2013, 19, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Santry, L.A.; de Jong, J.; Gold, A.C.; Walsh, S.R.; Menzies, P.I.; Wootton, S.K. Genetic characterization of small ruminant lentiviruses circulating in naturally infected sheep and goats in Ontario, Canada. Virus Res. 2013, 175, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Reina, R.; Mora, M.I.; Glaria, I.; García, I.; Solano, C.; Luján, L.; Badiola, J.J.; Contreras, A.; Berriatua, E.; Juste, R.; et al. Molecular characterization and phylogenetic study of Maedi Visna and Caprine Arthritis Encephalitis viral sequences in sheep and goats from Spain. Virus Res. 2006, 121, 189–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glaria, I.; Reina, R.; Crespo, H.; de Andrés, X.; Ramírez, H.; Biescas, E.; Pérez, M.M.; Badiola, J.; Luján, L.; Amorena, B.; et al. Phylogenetic analysis of SRLV sequences from an arthritic sheep outbreak demonstrates the introduction of CAEV-like viruses among Spanish sheep. Vet. Microbiol. 2009, 138, 156–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giammarioli, M.; Bazzucchi, M.; Puggioni, G.; Brajon, G.; Dei Giudici, S.; Taccori, F.; Feliziani, F.; De Mia, G.M. Phylogenetic analysis of small ruminant lentivirus (SRLV) in Italian flocks reveals the existence of novel genetic subtypes. Virus Genes 2011, 43, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Cardinaux, L.; Zahno, M.L.; Deubelbeiss, M.; Zanoni, R.; Vogt, H.R.; Bertoni, G. Virological and phylogenetic characterization of attenuated small ruminant lentivirus isolates eluding efficient serological detection. Vet. Microbiol. 2013, 162, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Purdy, J.G.; Flanagan, J.M.; Ropson, I.J.; Rennoll-Bankert, K.E.; Craven, R.C. Critical role of conserved hydrophobic residues within the major homology region in mature retroviral capsid assembly. J. Virol. 2008, 82, 5951–5961. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.H.; Chang, Y.F.; Wang, C.T. Mutations in the alpha-helix directly C-terminal to the major homology region of human immunodeficiency virus type 1 capsid protein disrupt Gag multimerization and markedly impair virus particle production. J. Biomed. Sci. 2006, 13, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulou, K.; Brellou, G.D.; Greenland, T.; Vlemmas, I. A novel deletion in the LTR region of a Greek small ruminant lentivirus may be associated with low pathogenicity. Virus Res. 2006, 118, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Kokawa, S.; Oba, M.; Hirata, T.; Tamaki, S.; Omura, M.; Tsuchiaka, S.; Nagai, M.; Omatsu, T.; Mizutani, T. Molecular characteristics and prevalence of small ruminant lentiviruses in goats in Japan. Arch. Virol. 2017, 162, 3007–3015. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Grade of Mastitis | IHC | LTR PCR | Gag-pol PCR | Genotype |
|---|---|---|---|---|---|
| M1 | moderate | + | + | + | B2 |
| M2 | minimal | + | + | − | − |
| M3 | severe | + | + | + | B2 |
| M4 | moderate | + | + | − | − |
| M5 | not classified | + | + | + | A2/A3 |
| M6 | moderate | + | + | − | − |
| M7 | moderate | + | + | + | − |
| M8 | moderate | + | + | + | − |
| M9 | severe | + | + | + | − |
| M10 | minimal | + | + | − | − |
| M11 | moderate | + | + | + | − |
| M12 | minimal | + | + | + | A2/A3 |
| M13 | no | − | − | − | − |
| M14 | moderate | + | + | − | − |
| M15 | minimal | + | + | + | A2/A3 |
| M16 | minimal | + | + | − | − |
| M17 | moderate | + | + | + | B2 |
| M18 | no | − | − | − | − |
| M19 | moderate | + | + | + | A2/A3 |
| Q1 | no | + | + | + | B2 |
| Q2 | no | − | − | − | − |
| Q3 | minimal | + | − | − | − |
| Q4 | minimal | + | + | − | − |
| Q5 | minimal | + | + | − | − |
| Q6 | severe | + | + | − | − |
| Q7 | no | + | + | + | A2/A3 |
| Q8 | severe | + | + | + | A2/A3 |
| Q9 | moderate | + | + | − | − |
| Q10 | severe | + | + | + | A2/A3 |
| Q11 | moderate | + | + | − | − |
| Q12 | moderate | + | + | + | − |
| Q13 | minimal | + | − | − | − |
| Q14 | moderate | + | + | − | − |
| Q15 | minimal | + | + | − | − |
| Q16 | minimal | + | + | − | − |
| N16-426 | severe | + | + | + | A2/A3 |
| N17-44 | moderate | + | + | + | A2/A3 |
| Sample | Genotype | M5 | M12 | M15 | M19 | Q7 | Q8 | Q10 | N16-426 | N17-44 | M1 | M3 | M17 | Q1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M12 | 0.112 | - | ||||||||||||
| M15 | 0.013 | 0.013 | - | |||||||||||
| M19 | 0.013 | 0.122 | 0.104 | - | ||||||||||
| Q7 | 0.130 | 0.133 | 0.129 | 0.109 | - | |||||||||
| Q8 | 0.100 | 0.115 | 0.097 | 0.122 | 0.122 | - | ||||||||
| Q10 | 0.109 | 0.119 | 0.105 | 0.129 | 0.127 | 0.105 | - | |||||||
| N16-426 | 0.127 | 0.138 | 0.129 | 0.115 | 0.137 | 0.127 | 0.135 | - | ||||||
| N17-44 | 0.152 | 0.155 | 0.152 | 0.138 | 0.163 | 0.157 | 0.152 | 0.094 | - | |||||
| M1 | 0.208 | 0.213 | 0.208 | 0.213 | 0.216 | 0.209 | 0.216 | 0.217 | 0.216 | - | ||||
| M3 | 0.216 | 0.222 | 0.211 | 0.221 | 0.231 | 0.211 | 0.221 | 0.224 | 0.217 | 0.040 | - | |||
| M17 | 0.214 | 0.221 | 0.209 | 0.226 | 0.232 | 0.217 | 0.217 | 0.231 | 0.227 | 0.048 | 0.041 | - | ||
| Q1 | 0.209 | 0.216 | 0.203 | 0.217 | 0.222 | 0.221 | 0.219 | 0.216 | 0.216 | 0.081 | 0.086 | 0.076 | - | |
| M10608.1 | A1 | 0.171 | 0.160 | 0.168 | 0.137 | 0.157 | 0.166 | 0.145 | 0.175 | 0.175 | 0.214 | 0.208 | 0.229 | 0.224 |
| S51392 | A1 | 0.173 | 0.165 | 0.168 | 0.173 | 0.181 | 0.171 | 0.163 | 0.176 | 0.168 | 0.242 | 0.247 | 0.245 | 0.244 |
| AY101611.1 | A2 | 0.155 | 0.135 | 0.153 | 0.150 | 0.147 | 0.153 | 0.152 | 0.165 | 0.165 | 0.186 | 0.188 | 0.191 | 0.201 |
| HQ848062.1 | A2/A3 | 0.117 | 0.135 | 0.120 | 0.105 | 0.122 | 0.127 | 0.135 | 0.142 | 0.124 | 0.206 | 0.209 | 0.219 | 0.213 |
| AY454176.1 | A3 | 0.130 | 0.138 | 0.130 | 0.130 | 0.137 | 0.133 | 0.145 | 0.145 | 0.152 | 0.178 | 0.191 | 0.193 | 0.198 |
| AY454161.1 | A4 | 0.138 | 0.166 | 0.135 | 0.153 | 0.185 | 0.143 | 0.150 | 0.173 | 0.171 | 0.211 | 0.216 | 0.214 | 0.214 |
| AY454175.1 | A5 | 0.153 | 0.145 | 0.155 | 0.148 | 0.171 | 0.160 | 0.147 | 0.176 | 0.165 | 0.209 | 0.203 | 0.208 | 0.206 |
| AY454208.1 | A7 | 0.176 | 0.157 | 0.170 | 0.145 | 0.158 | 0.158 | 0.166 | 0.161 | 0.140 | 0.198 | 0.209 | 0.201 | 0.209 |
| FR694908 | A9 | 0.166 | 0.168 | 0.170 | 0.166 | 0.178 | 0.180 | 0.171 | 0.181 | 0.161 | 0.213 | 0.224 | 0.224 | 0.231 |
| FR693825 | A11 | 0.183 | 0.171 | 0.180 | 0.152 | 0.157 | 0.176 | 0.183 | 0.189 | 0.178 | 0.213 | 0.217 | 0.231 | 0.232 |
| M33677 | B1 | 0.204 | 0.214 | 0.201 | 0.211 | 0.224 | 0.211 | 0.211 | 0.222 | 0.211 | 0.120 | 0.119 | 0.120 | 0.129 |
| FJ195346.1 | B2 | 0.226 | 0.231 | 0.221 | 0.214 | 0.232 | 0.226 | 0.237 | 0.221 | 0.217 | 0.092 | 0.092 | 0.102 | 0.100 |
| EU010126.1 | B2 | 0.208 | 0.222 | 0.206 | 0.226 | 0.232 | 0.219 | 0.227 | 0.224 | 0.216 | 0.064 | 0.068 | 0.068 | 0.081 |
| JF502417.1 | B3 | 0.213 | 0.221 | 0.214 | 0.209 | 0.217 | 0.219 | 0.232 | 0.213 | 0.224 | 0.185 | 0.186 | 0.196 | 0.178 |
| AF322109.1 | C | 0.262 | 0.252 | 0.257 | 0.250 | 0.252 | 0.245 | 0.255 | 0.244 | 0.236 | 0.213 | 0.214 | 0.214 | 0.216 |
| EU293537.2 | E1 | 0.292 | 0.295 | 0.292 | 0.297 | 0.301 | 0.293 | 0.300 | 0.293 | 0.290 | 0.293 | 0.285 | 0.290 | 0.303 |
| GQ381130.1 | E2 | 0.290 | 0.301 | 0.293 | 0.290 | 0.313 | 0.293 | 0.298 | 0.288 | 0.290 | 0.290 | 0.290 | 0.297 | 0.293 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gayo, E.; Cuteri, V.; Polledo, L.; Rossi, G.; García Marín, J.F.; Preziuso, S. Genetic Characterization and Phylogenetic Analysis of Small Ruminant Lentiviruses Detected in Spanish Assaf Sheep with Different Mammary Lesions. Viruses 2018, 10, 315. https://doi.org/10.3390/v10060315
Gayo E, Cuteri V, Polledo L, Rossi G, García Marín JF, Preziuso S. Genetic Characterization and Phylogenetic Analysis of Small Ruminant Lentiviruses Detected in Spanish Assaf Sheep with Different Mammary Lesions. Viruses. 2018; 10(6):315. https://doi.org/10.3390/v10060315
Chicago/Turabian StyleGayo, Elena, Vincenzo Cuteri, Laura Polledo, Giacomo Rossi, Juan F. García Marín, and Silvia Preziuso. 2018. "Genetic Characterization and Phylogenetic Analysis of Small Ruminant Lentiviruses Detected in Spanish Assaf Sheep with Different Mammary Lesions" Viruses 10, no. 6: 315. https://doi.org/10.3390/v10060315