Multi-Environment Evaluation and Genetic Characterisation of Common Bean Breeding Lines for Organic Farming Systems

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
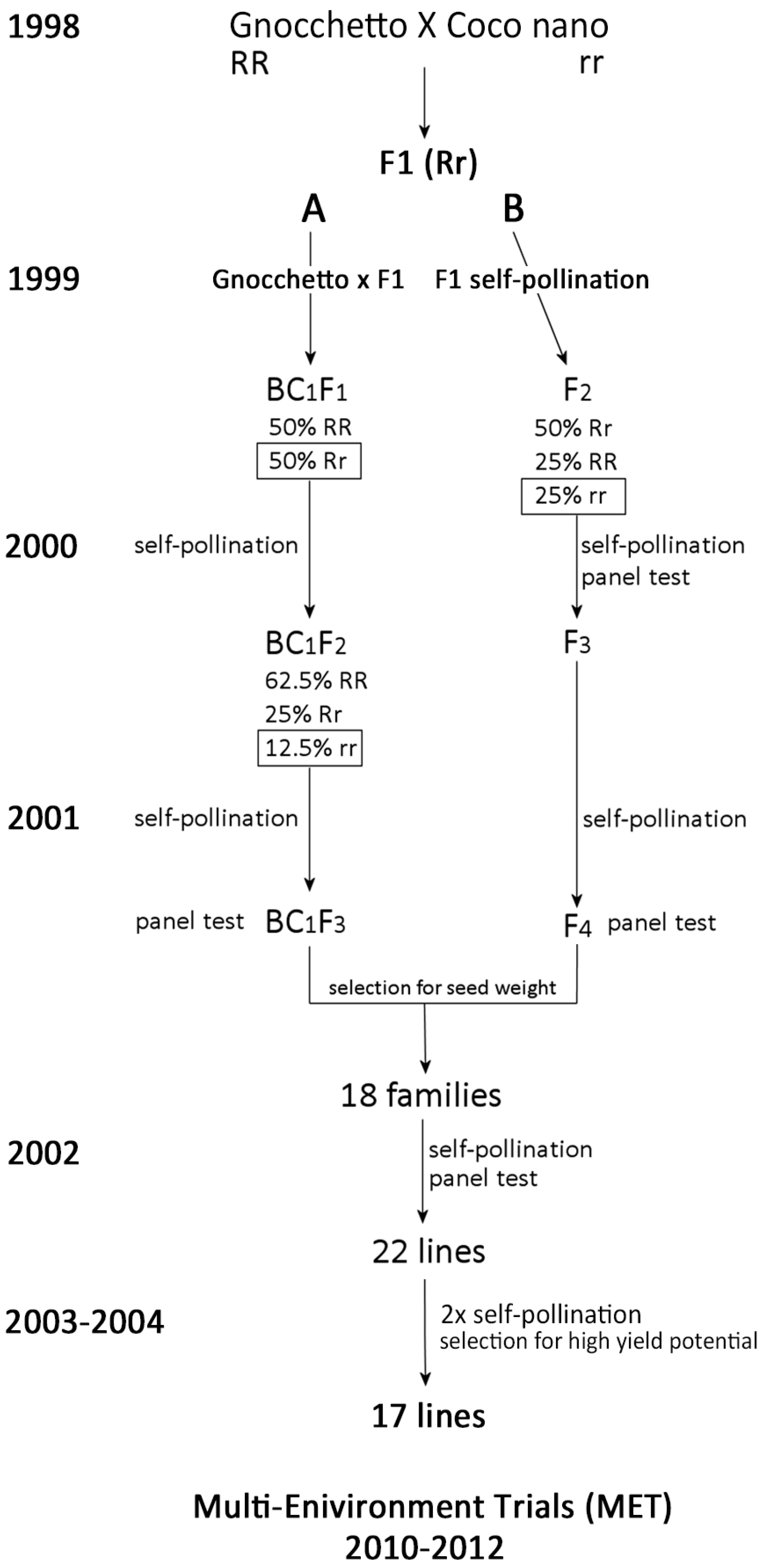
2.1. The Breeding Programme and Lines
2.2. Multi-Environment Field Trials
2.3. Statistical Analysis of Yield Data
2.4. DNA Isolation, Markers Selection and Genetic Characterization
2.5. Genetic Diversity Analysis
3. Results
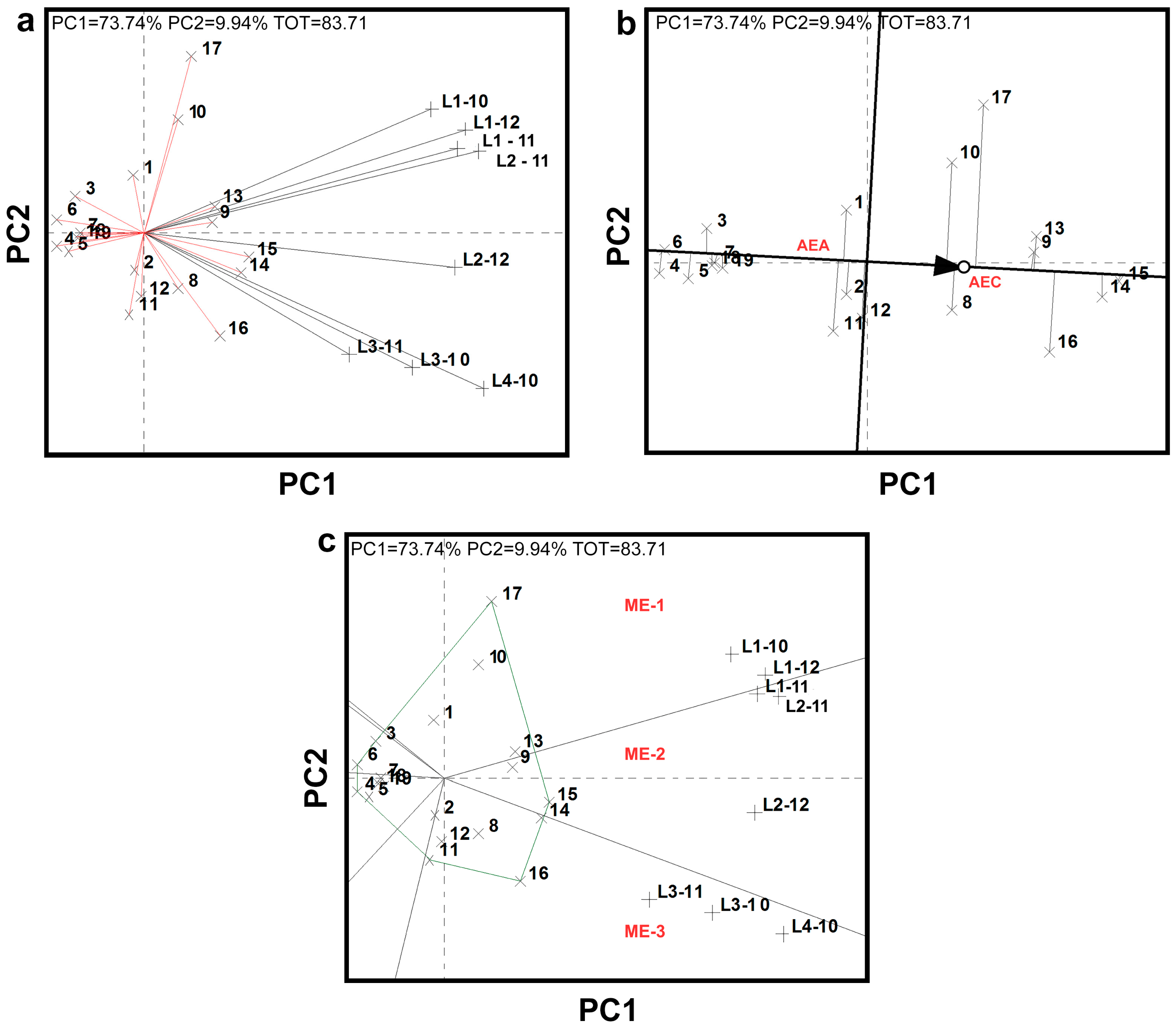
3.1. Multi-Environment Field Trials and Statistical Analysis
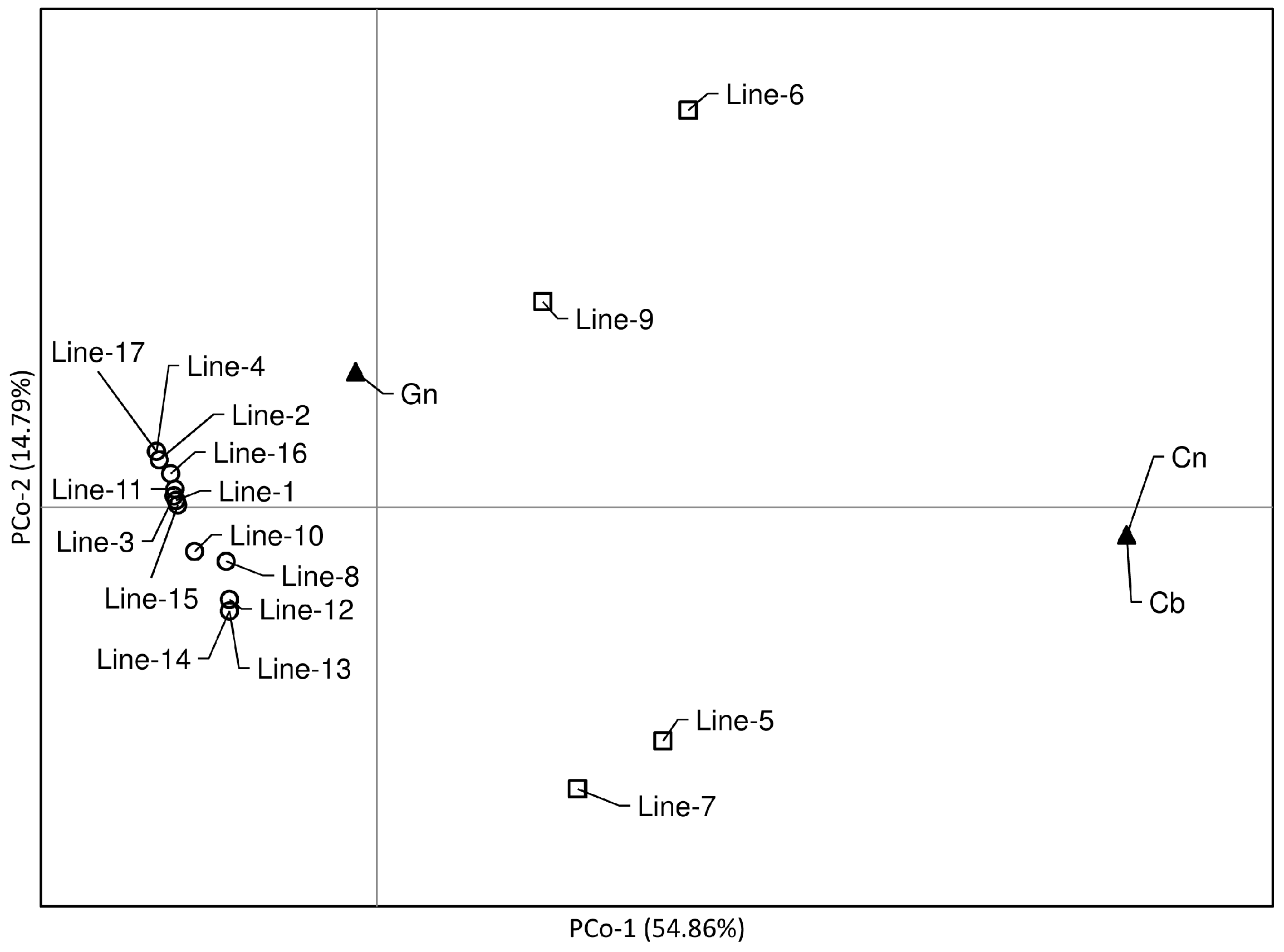
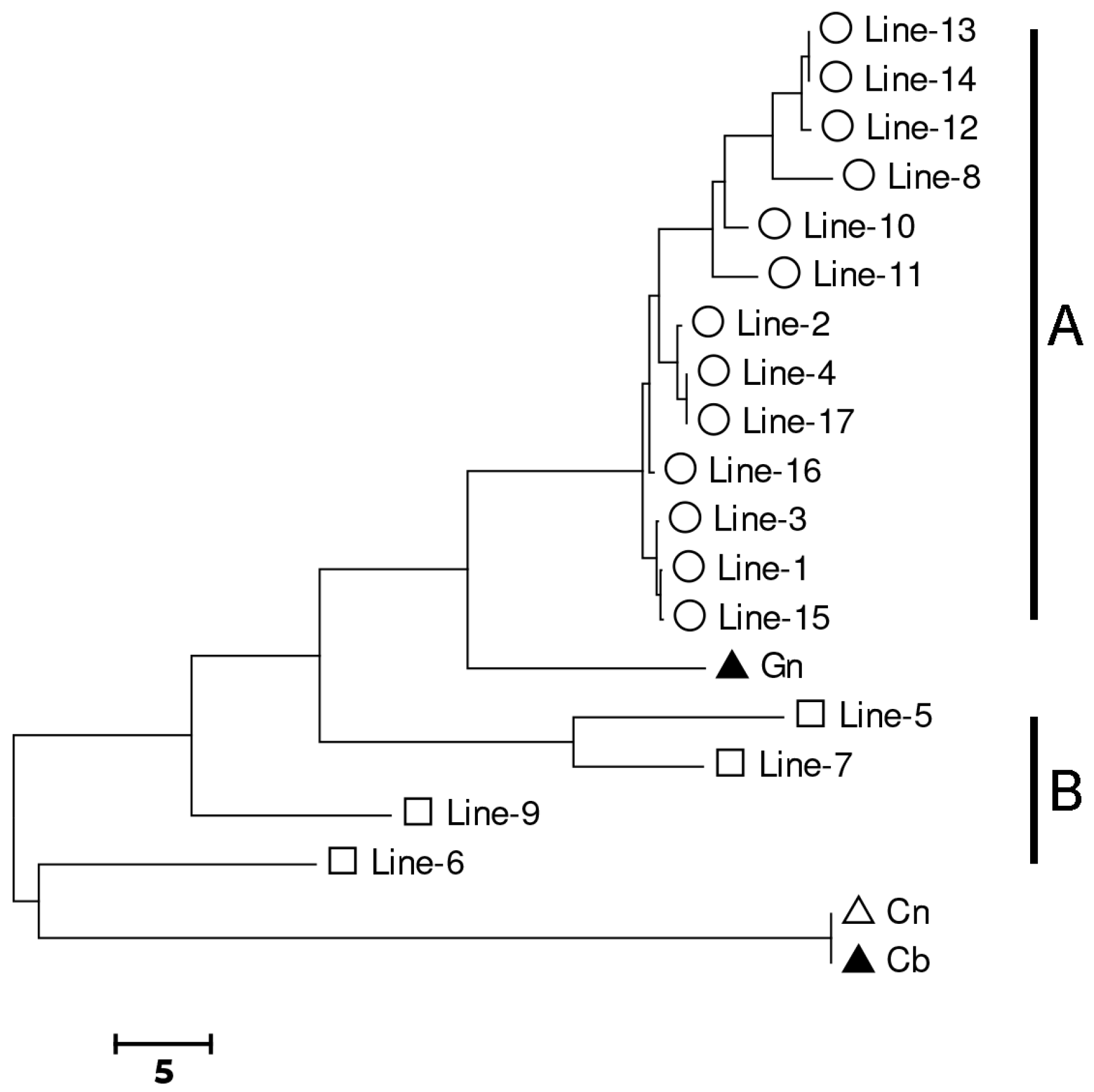
3.2. Genotyping and Genetic Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Willer, H.; Kilcher, L. The World of Organic Agriculture—Statistics and Emerging Trends 2012; Willer, H., Kilcher, L., Eds.; IFOAM: Bonn, Germany; FiBL: Frick, Switzerland, 2012. [Google Scholar]
- Shafie, F.A.; Rennie, D. Consumer Perceptions Towards Organic Food. Procedia Soc. Behav. Sci. 2012, 49, 360–367. [Google Scholar] [CrossRef]
- Stanhill, G. The comparative productivity of organic agriculture. Agric. Ecosyst. Environ. 1990, 30, 1–26. [Google Scholar] [CrossRef]
- Padel, S.; Lampkin, N. Conversion to organic farming: An overview. In The Economics of Organic Farming; Lampkin, N.H., Padel, S., Eds.; CAB: Wallingford, UK, 1994; pp. 295–313. [Google Scholar]
- Mäder, P.; Fliessbach, A.; Dubois, D.; Gunst, L.; Fried, P.; Niggli, U. Soil fertility and biodiversity in organic farming. Science 2002, 296, 1694–1697. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.H.; Derrick, J.W.; Dann, P.R. Grain mineral concentrations and yield of wheat grown under organic and conventional management. J. Sci. Food Agric. 2004, 84, 207–216. [Google Scholar] [CrossRef]
- Smolik, J.; Dobbs, T. Crop yields and economic returns accompanying the transition to alternative farming systems. J. Prod. Agric. 1991, 4, 153–161. [Google Scholar] [CrossRef]
- Clark, M.S.; Horwath, W.R.; Shennan, C.; Scow, K.M.; Lantni, W.T.; Ferris, H. Nitrogen, weeds and water as yield-limiting factors in conventional, low-input, and organic tomato systems. Agric. Ecosyst. Environ. 1999, 73, 257–270. [Google Scholar] [CrossRef]
- Reganold, J.P.; Glover, J.D.; Andrews, P.K.; Hinman, H.R. Sustainability of three apple production systems. Nature 2001, 410, 926–930. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, D.; Hepperly, P.; Hanson, J.; Douds, D.; Seidel, R. Environmental, Energetic, and Economic Comparisons of Organic and Conventional Farming Systems. Bioscience 2005, 55, 573–582. [Google Scholar] [CrossRef]
- Posner, J.L.; Baldock, J.O.; Hedtcke, J.L. Organic and Conventional Production Systems in the Wisconsin Integrated Cropping Systems Trials: I. Productivity 1990–2002. Agron. J. 2008, 100, 253–260. [Google Scholar] [CrossRef]
- De Ponti, T.; Rijk, B.; Van Ittersum, M.K. The crop yield gap between organic and conventional agriculture. Agric. Syst. 2012, 108, 1–9. [Google Scholar] [CrossRef]
- Seufert, V.; Ramankutty, N.; Foley, J.A. Comparing the yields of organic and conventional agriculture. Nature 2012, 485, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Ponisio, L.C.; M’Gonigle, L.K.; Mace, K.C.; Palomino, J.; De Valpine, P.; Kremen, C. Diversification practices reduce organic to conventional yield gap. Proc. R. Soc. B Biol. Sci. 2015, 282, 1–7. [Google Scholar] [CrossRef]
- Lammerts Van Bueren, E.T.; Jones, S.S.; Tamm, L.; Murphy, K.M.; Myers, J.R.; Leifert, C.; Messmer, M.M. The need to breed crop varieties suitable for organic farming, using wheat, tomato and broccoli as examples: A review. NJAS Wagening. J. Life Sci. 2011, 58, 193–205. [Google Scholar] [CrossRef]
- Murphy, K.M.; Campbell, K.G.; Lyon, S.R.; Jones, S.S. Evidence of varietal adaptation to organic farming systems. Field Crop. Res. 2007, 102, 172–177. [Google Scholar] [CrossRef]
- Lammerts van Bueren, E.T.; Struik, P.C.; Jacobsen, E. Ecological concepts in organic farming and their consequences for an organic crop ideotype. NJAS Wagening. J. Life Sci. 2002, 50, 1–26. [Google Scholar] [CrossRef]
- Wolfe, M.S.; Baresel, J.P.; Desclaux, D.; Goldringer, I.; Hoad, S.; Kovacs, G.; Löschenberger, F.; Miedaner, T.; Østergård, H.; Lammerts Van Bueren, E.T. Developments in breeding cereals for organic agriculture. Euphytica 2008, 163, 323–346. [Google Scholar] [CrossRef]
- Hildermann, I.; Thommen, A.; Dubois, D.; Boller, T.; Wiemken, A.; Mader, P. Yield and baking quality of winter wheat cultivars in different farming systems of the DOK long-term trial. J. Sci. Food Agric. 2009, 2477–2491. [Google Scholar] [CrossRef]
- Saastamoinen, M.; Hietaniemi, V.; Pihlava, J.-M.; Merja, E.; Kontturi, M.; Tuuri, H.; Niskanen, M.; Kangas, A. β-Glucan contents of groats of different oat cultivars in official variety, in organic cultivation, and in nitrogen ferilization trials in Finland. Agric. Food Sci. 2004, 13, 68–79. [Google Scholar] [CrossRef]
- Heilig, J.A.; Kelly, J.D. Performance of Dry Bean Genotypes Grown under Organic and Conventional Production Systems in Michigan. Agron. J. 2012, 104, 1485. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Chiapparino, E.; Perenzin, M. Response of common wheat varieties to organic and conventional production systems across Italian locations, and implications for selection. Field Crop. Res. 2010, 116, 230–238. [Google Scholar] [CrossRef]
- Raggi, L.; Ciancaleoni, S.; Torricelli, R.; Terzi, V.; Ceccarelli, S.; Negri, V. Field Crops Research Evolutionary breeding for sustainable agriculture: Selection and multi-environmental evaluation of barley populations and lines. Field Crop. Res. 2017, 204, 76–88. [Google Scholar] [CrossRef]
- Lammerts van Bueren, E.T.; Backes, G.; de Vriend, H.; Østergård, H. The role of molecular markers and marker assisted selection in breeding for organic agriculture. Euphytica 2010, 175, 51–64. [Google Scholar] [CrossRef]
- Fess, T.L.; Kotcon, J.B.; Benedito, V.A. Crop breeding for low input agriculture: A sustainable response to feed a growing world population. Sustainability 2011, 3, 1742–1772. [Google Scholar] [CrossRef]
- Tiranti, B.; Negri, V. Selective microenvironmental effects play a role in shaping genetic diversity and structure in a Phaseolus vulgaris L. landrace: Implications for on-farm conservation. Mol. Ecol. 2007, 16, 4942–4955. [Google Scholar] [CrossRef] [PubMed]
- Brush, S. The issues of in situ conservation of crop genetic resources. In Genes in the Field. On Farm Conservation of Crop Diversity; Brush, S., Ed.; IPGRI: Rome, Italy, 2000; pp. 3–28. [Google Scholar]
- Klaedtke, S.; Caproni, L.; Klauck, J.; de la Grandville, P.; Dutartre, M.; Stassart, P.; Chable, V.; Negri, V.; Raggi, L. Short-Term Local Adaptation of Historical Common Bean (Phaseolus vulgaris L.) Varieties and Implications for In Situ Management of Bean Diversity. Int. J. Mol. Sci. 2017, 18, 493. [Google Scholar] [CrossRef] [PubMed]
- Negri, V.; Tosti, N. Phaseolus genetic diversity maintained on-farm in central Italy. Genet. Resour. Crop Evol. 2002, 49, 511–520. [Google Scholar] [CrossRef]
- Broughton, W.; Hernandez, G.; Blair, M. Beans (Phaseolus spp.)—model food legumes. Plant Soil 2003, 252, 55–128. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTAT Database. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 20 August 2017).
- United States Department of Agriculture (USDA). Economic Research Service. 2013. Available online: https://www.ers.usda.gov/data-products/organic-production.aspx (accessed on 20 August 2017).
- European Commission Statistics Database (EUROSTAT). Available online: http://ec.europa.eu/eurostat/data/database (accessed on 20 August 2017).
- Graham, P.H.; Ranalli, P. Common bean (Phaseolus vulgaris L.). Field Crop. Res. 1997, 53, 131–146. [Google Scholar] [CrossRef]
- Negri, V.; Tiranti, B. Effectiveness of in situ and ex situ conservation of crop diversity. What a Phaseolus vulgaris L. landrace case study can tell us. Genetica 2010, 138, 985–998. [Google Scholar] [CrossRef] [PubMed]
- Wiesinger, J.A.; Cichy, K.A.; Glahn, R.P.; Grusak, M.A.; Brick, M.A.; Thompson, H.J.; Tako, E. Demonstrating a Nutritional Advantage to the Fast-Cooking Dry Bean (Phaseolus vulgaris L.). J. Agric. Food Chem. 2016, 64, 8592–8603. [Google Scholar] [CrossRef] [PubMed]
- Kwak, M.; Toro, O.; Debouck, D.G.; Gepts, P. Multiple origins of the determinate growth habit in domesticated common bean (Phaseolus vulgaris). Ann. Bot. 2012, 110, 1573–1580. [Google Scholar] [CrossRef] [PubMed]
- Kornegay, J.; White, J.W.; de la Cruz, O.O. Growth habit and gene pool effects on inheritance of yield in common bean. Euphytica 1992, 62, 171–180. [Google Scholar] [CrossRef]
- Kwak, M.; Velasco, D.; Gepts, P. Mapping homologous sequences for determinacy and photoperiod sensitivity in common bean (Phaseolus vulgaris). J. Hered. 2008, 99, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Torricelli, R.; Lorenzetti, S.; Falcinelli, M. La valorizzazione di una varietà locale di fagiolo attraverso il miglioramento genetico. Sementi Elette 2003, 6, 35–39. [Google Scholar]
- Garruti, R.D.S.; Bourne, M.C. Effect of Storage Conditions of Dry Bean Seeds ( Phaseolus on Texture Profile Parameters After Cooking vulgaris L.). J. Food Sci. 1985, 50, 1067–1071. [Google Scholar] [CrossRef]
- Yan, W.; Tinker, N.A. Biplot analysis of multi-environment trial data: Principles and applications. Can. J. Plant Sci. 2006, 86, 623–645. [Google Scholar] [CrossRef]
- Voysest, O.; Dessert, M. Bean cultivars: Classes and commercial seed types. In Common Beans: Research for Crop Improvement; van Schoonhoven, A., Yovest, O., Eds.; CAB International, in association with CIAT: Wallingford, UK, 1991; pp. 119–162. [Google Scholar]
- De Ron, A.M.; Rodiño, A.P.; Santalla, M.; González, A.M.; Lema, M.J.; Martín, I.; Kigel, J. Seedling Emergence and Phenotypic Response of Common Bean Germplasm to Different Temperatures under Controlled Conditions and in Open Field. Front. Plant Sci. 2016, 7, 1087. [Google Scholar] [CrossRef] [PubMed]
- Thung, M. Bean Agronomy in Monoculture. In Common Beans: Research for Crop Improvement; van Schoonhoven, A., Yovest, O., Eds.; CAB International, in association with CIAT: Wallingford, UK, 1991; pp. 737–834. [Google Scholar]
- Yu, K.; Park, S.J.; Poysa, V.; Gepts, P. Integration of simple sequence repeat (SSR) markers into a molecular linkage map of common bean (Phaseolus vulgaris L.). J. Hered. 2000, 91, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Gaitán-Solís, E.; Duque, M.C.C.; Edwards, K.J.J.; Tohme, J. Microsatellite Repeats in Common Bean (Phaseolus vulgaris): Isolation, Characterization, and Cross-Species Amplification in Phaseolus spp. Crop Sci. 2002, 42, 2128–2136. [Google Scholar] [CrossRef]
- Guerra-Sanz, J.M. Short Communication—New SSR markers of Phaseolus vulgaris from sequence databases. Plant Breed. 2004, 89, 87–90. [Google Scholar] [CrossRef]
- Blair, M.W.; Pedraza, F.; Buendia, H.F.; Gaitán-Solís, E.; Beebe, S.E.; Gepts, P.; Tohme, J. Development of a genome-wide anchored microsatellite map for common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2003, 107, 1362–1374. [Google Scholar] [CrossRef] [PubMed]
- Blair, M.W.; Muñoz-Torres, M.C.; Pedraza, F.; Giraldo, M.C.; Buendía, H.F.; Hurtado, N. Development of microsatellite markers for common bean (Phaseolus vulgaris L.) based on screening of non-enriched, small-insert genomic libraries. Genome 2009, 52, 772–782. [Google Scholar] [CrossRef] [PubMed]
- Córdoba, J.M.; Chavarro, C.; Rojas, F.; Muñoz, C.; Blair, M.W. Identification and Mapping of Simple Sequence Repeat Markers from Common Bean (Phaseolus vulgaris L.) Bacterial Artificial Chromosome End Sequences for Genome Characterization and Genetic–Physical Map Integration. Plant Genome J. 2010, 3, 154. [Google Scholar] [CrossRef]
- Raggi, L.; Tissi, C.; Mazzucato, A.; Negri, V. Molecular polymorphism related to flowering trait variation in a Phaseolus vulgaris L. collection. Plant Sci. 2014, 215–216, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Martins, W.S.; Lucas, D.C.S.; de Souza Neves, K.F.; Bertioli, D.J. WebSat—A web software for microsatellite marker development. Bioinformation 2009, 3, 282–283. [Google Scholar] [CrossRef] [PubMed]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. In Methods in Molecular Biology: Bioinformatics Methods and Protocols; Krawetz, S., Misener, Eds.; Humana Press Inc.: Totowa, NJ, USA, 1999; pp. 365–386. [Google Scholar]
- Shen, Z.; Qu, W.; Wang, W.; Lu, Y.; Wu, Y.; Li, Z.; Hang, X.; Wang, X. MPprimer: A program for reliable multiplex PCR primer design. BMC Bioinform. 2010, 11, 143. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Crow, J.F. The Number of Alleles That Can Be Maintained in a Finite Population. Genetics 1964, 49, 725–738. [Google Scholar] [PubMed]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Mcclean, P.E.; Burridge, J.; Beebe, S.E.; Rao, I.M.; Porch, T.G. Crop improvement in the era of climate change: An integrated, multi-disciplinary approach for common bean (Phaseolus vulgaris). Funct. Plant Biol. 2011, 38, 927–933. [Google Scholar] [CrossRef]
- Rainey, K.M.; Griffi, P.D. Inheritance of Heat Tolerance during Reproductive Development in Snap Bean (Phaseolus vulgaris L.). Differences 2005, 130, 700–706. [Google Scholar]
- Omae, H.; Kumar, A.; Shono, M. Adaptation to high temperature and water deficit in the common bean (Phaseolus vulgaris L.) during the reproductive period. J. Bot. 2012, 2012, 803413. [Google Scholar] [CrossRef]
- Porch, T.G.; Smith, J.R.; Beaver, J.S.; Griffiths, P.D.; Station, E.; Street, W.N.; Canaday, C.H. TARS-HT1 and TARS-HT2 Heat-tolerant Dry Bean Germplasm. Hortic. Sci. 2010, 45, 1278–1280. [Google Scholar]
- Tolessa, T.T.; Keneni, G.; Sefera, T.; Jarso, M.; Bekele, Y. Genotype × Environment Interaction and Performance Stability for Grain Yield in Field Pea (Pisum sativum L.) Genotypes. Int. J. Plant Breed. 2013, 7, 116–123. [Google Scholar]
- Edugbo, R.E.; Nwofia, G.E.; Fayeun, L.S. An Assessment of Soybean (Glycine max, L. Merrill ) Grain Yield in Different Environments Using AMMI and GGE Biplot Models in Humidorest Fringes of Southeast Nigeria. Agric. Trop. Subtrop. 2015, 48, 82–90. [Google Scholar] [CrossRef]
- Mehari, M.; Sentayehu, A.; Berhane, L. Genotype X Environment Interaction and Yield Stability of Malt Barley Genotypes Evaluated in Tigray, Ethiopia Using the Ammi Analysis. Asian J. Plant Sci. 2014, 13, 73–79. [Google Scholar]
- Yahiaoui, S.; Cuesta-Marcos, A.; Gracia, M.P.; Medina, B.; Lasa, J.M.; Casas, A.M.; Ciudad, F.J.; Montoya, J.L.; Moralejo, M.; Molina-Cano, J.L.; et al. Spanish barley landraces outperform modern cultivars at low-productivity sites. Plant Breed. 2014, 133, 218–226. [Google Scholar] [CrossRef]
- Ceccarelli, B.S.; Erskine, W.; Hamblin, J.; Grando, S. Genotype by environment interaction and international breeding programmes. Exp. Agric. 1994, 30, 177–187. [Google Scholar] [CrossRef]
- Rao, I.M.; Beebe, S.E.; Polania, J.; Grajales, M.; Cajiao, C.; Ricaurte, J.; García, R.; Rivera, M. Evidence for genotypic differences among elite lines of common bean in the ability to remobilize photosynthate to increase yield under drought. J. Agric. Sci. 2017, 155, 857–875. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Name | Biological Status | Pedigree Note |
|---|---|---|---|
| Line-1 | Solibam-01 | Breeding line | BC1F1 offspring |
| Line-2 | Solibam-02 | Breeding line | BC1F1 offspring |
| Line-3 | Solibam-03 | Breeding line | BC1F1 offspring |
| Line-4 | Solibam-04 | Breeding line | BC1F1 offspring |
| Line-5 | Solibam-05 | Breeding line | F2 offspring |
| Line-6 | Solibam-06 | Breeding line | F2 offspring |
| Line-7 | Solibam-07 | Breeding line | F2 offspring |
| Line-8 | Solibam-08 | Breeding line | BC1F1 offspring |
| Line-9 | Solibam-09 | Breeding line | F2 offspring |
| Line-10 | Solibam-10 | Breeding line | BC1F1 offspring |
| Line-11 | Solibam-11 | Breeding line | BC1F1 offspring |
| Line-12 | Solibam-12 | Breeding line | BC1F1 offspring |
| Line-13 | Solibam-13 | Breeding line | BC1F1 offspring |
| Line-14 | Solibam-14 | Breeding line | BC1F1 offspring |
| Line-15 | Solibam-15 | Breeding line | BC1F1 offspring |
| Line-16 | Solibam-16 | Breeding line | BC1F1 offspring |
| Line-17 | Solibam-17 | Breeding line | BC1F1 offspring |
| Gn * | Gnocchetto | Landrace | Parental line |
| Cn | Coco nano | Cultivar | Parental line |
| Cb | Coco blanc Gautier | Cultivar | Control |
| San Martino in Campo, Perugia, IT | Rispescia, Grosseto, IT | Fressingfield, Suffolk, UK | Bellegarde du Razes Limoux, FR | |||||
|---|---|---|---|---|---|---|---|---|
| Coordinates | 43°02′35.34′′ N; 12°25′00.94′′ E | 42°42′11.81′′N; 11°08′31.33′′E | 52°21′14.15′′N; 1°21′16.73′′E | 43°06′31.07′′N; 2°02′44.43′′E | ||||
| Altitude (m a.s.l.) | 174 | 26 | 50 | 310 | ||||
| Environment | L1-10 | L1-11 | L1-12 | L2-11 | L2-12 | L3-10 | L3-11 | L4-10 |
| Year | 2010 | 2011 | 2012 | 2011 | 2012 | 2010 | 2011 | 2010 |
| Tmin a SF b | 13.0 | 15.1 | 13.1 | 15.2 | 14.7 | 11.5 | 9.0 | 14.1 |
| Tmax c SF | 26.8 | 28.4 | 28.0 | 27.4 | 27.1 | 20.9 | 18.4 | 25.1 |
| Rh d SF | 62.3 | 59.4 | 56.9 | 67.0 | 44.8 | 68.9 | 70.3 | 68.1 |
| Rnfl e SF | 95.8 | 86.8 | 63.4 | 41.8 | 24.6 | 49.0 | 38.0 | 81.3 |
| Tmin FM f | 16.8 | 16.7 | 17.6 | 17.3 | 18.6 | 11.1 | 11.6 | 17.2 |
| Tmax FM | 31.9 | 31.5 | 34.1 | 29.6 | 31.5 | 18.7 | 20.0 | 29.2 |
| Rh FM | 51.3 | 51.3 | 41.1 | 66.8 | 42.3 | 77.3 | 75.8 | 59.0 |
| Rnfl FM | 87.2 | 63.6 | 53.6 | 116.2 | 32.6 | 111.8 | 82.0 | 65.4 |
| Sowing date | 20 May | 24 May | 16 May | 20 May | 21 May | 2 June | 12 May | 21 May |
| Average dtf g | 45 | 44 | 47 | 47 | 40 | 43 | 53 | 41 |
| Average dtm h | 112 | 103 | 107 | 98 | 107 | 136 | 121 | 101 |
| ID | Environment | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L1-10 | L1-11 | L1-12 | L2-11 | L2-12 | L3-10 | L3-11 | L4-10 | Mean | ||||||||||
| Line-1 | 65.3 | AB | 14.6 | AB | 27.7 | A | 39.5 | CD | 18.8 | D | 72.2 | BCDE | 119.3 | A | 13.0 | AB | 46.31 | ABC |
| Line-2 | 64.4 | ABC | 14.7 | AB | 15.7 | AB | 56.0 | ABCD | 28.1 | ABCD | 69.6 | BCDE | 110.6 | AB | 20.5 | AB | 47.44 | ABC |
| Line-3 | 68.4 | A | 13.9 | AB | 6.4 | B | 52.5 | ABCD | 18.2 | D | 85.5 | ABCD | 94.8 | AB | 24.6 | A | 45.53 | BC |
| Line-4 | 57.7 | ABCD | 19.6 | AB | 24.3 | AB | 65.3 | ABC | 41.7 | ABC | 106.5 | A | 84.9 | AB | 17.6 | AB | 52.21 | AB |
| Line-5 | 23.8 | DE | 9.9 | AB | 11.1 | AB | 43.3 | BCD | 29.7 | ABCD | 61.7 | CDE | 82.4 | B | 19.2 | AB | 35.12 | DEF |
| Line-6 | 11.7 | E | 3.2 | B | 15.0 | AB | 35.1 | D | 18.9 | D | 44.3 | E | 101.3 | AB | 20.7 | AB | 31.26 | F |
| Line-7 | 15.3 | E | 4.6 | AB | 9.2 | AB | 55.1 | ABCD | 29.9 | ABCD | 57.8 | DE | 48.7 | C | 15.3 | AB | 29.50 | F |
| Line-8 | 52.7 | ABCD | 11.7 | AB | 13.7 | AB | 57.6 | ABCD | 30.6 | ABCD | 79.6 | ABCD | 107.9 | AB | 20.9 | AB | 46.85 | ABC |
| Line-9 | 16.3 | DE | 4.9 | AB | 7.8 | AB | 35.4 | D | 24.8 | CD | 48.3 | E | 80.3 | BC | 11.9 | B | 28.71 | F |
| Line-10 | 48.4 | ABCD | 25.3 | A | 4.3 | B | 66.2 | ABC | 33.1 | ABCD | 89.6 | ABCD | 85.4 | AB | 19.5 | AB | 46.48 | ABC |
| Line-11 | 59.5 | ABC | 21.9 | AB | 29.9 | A | 74.5 | A | 45.8 | A | 83.0 | ABCD | 109.4 | AB | 21.2 | AB | 55.66 | A |
| Line-12 | 54.2 | ABCD | 20.3 | AB | 39.0 | A | 67.3 | ABC | 44.9 | AB | 75.2 | ABCDE | 103.3 | AB | 16.4 | AB | 52.58 | AB |
| Line-13 | 41.1 | BCD | 9.9 | AB | 15.5 | AB | 46.2 | BCD | 33.8 | ABCD | 71.4 | BCDE | 78.8 | BC | 17.1 | AB | 39.24 | CDE |
| Line-14 | 36.7 | BCDE | 11.8 | AB | 22.2 | AB | 46.6 | ABCD | 19.7 | D | 66.5 | BCDE | 100.9 | AB | 13.6 | AB | 39.74 | CDE |
| Line-15 | 54.4 | ABCD | 20.5 | AB | 17.3 | AB | 53.5 | ABCD | 30.9 | ABCD | 94.1 | ABC | 82.0 | B | 17.1 | AB | 46.24 | ABC |
| Line-16 | 35.5 | CDE | 15.0 | AB | 19.8 | AB | 53.6 | ABCD | 36.6 | ABCD | 69.7 | BCDE | 90.9 | AB | 19.1 | AB | 42.53 | BCD |
| Line-17 | 35.8 | BCDE | 20.1 | AB | 42.7 | A | 59.0 | ABCD | 26.7 | BCD | 91.2 | ABC | 95.5 | AB | 19.0 | AB | 48.75 | ABC |
| Cn | 46.5 | ABCD | 17.2 | AB | 21.4 | AB | 65.9 | ABC | 27.9 | ABCD | 96.5 | AB | 98.3 | AB | 19.1 | AB | 49.10 | AB |
| Cb | 28.9 | DE | 11.9 | AB | 8.9 | AB | 67.9 | AB | 32.2 | ABCD | 89.8 | ABC | 103.6 | AB | 19.9 | AB | 45.38 | BC |
| Mean | 42.98 | 14.27 | 18.53 | 54.76 | 30.12 | 76.44 | 93.60 | 18.19 | 43.61 | |||||||||
| Source of Variation | d.f. a | SS b | SS (%) b | F c | p-Value |
|---|---|---|---|---|---|
| E d | 7 | 665,932.4 | 55.5 | 207.2 | *** |
| G e | 18 | 56,264.2 | 4.7 | 6.8 | *** |
| G x E f | 126 | 85,050.6 | 7.1 | 1.5 | *** |
| Error | 1368 | 393,525.0 | 32.8 | ||
| Total | 1519 | 1,200,772.1 | 100.0 |
| Source of Variation | d.f. a | SS b | v.c. c | v d |
|---|---|---|---|---|
| Among lines | 16 | 449.9 | 2.5 | 94.0% |
| Among individuals within lines | 76 | 19.2 | 0.1 | 3.4% |
| Within Individuals | 93 | 6.5 | 0.1 | 2.6% |
| Total | 185 | 475.7 | 2.7 | 100.0% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caproni, L.; Raggi, L.; Tissi, C.; Howlett, S.; Torricelli, R.; Negri, V. Multi-Environment Evaluation and Genetic Characterisation of Common Bean Breeding Lines for Organic Farming Systems. Sustainability 2018, 10, 777. https://doi.org/10.3390/su10030777
Caproni L, Raggi L, Tissi C, Howlett S, Torricelli R, Negri V. Multi-Environment Evaluation and Genetic Characterisation of Common Bean Breeding Lines for Organic Farming Systems. Sustainability. 2018; 10(3):777. https://doi.org/10.3390/su10030777
Chicago/Turabian StyleCaproni, Leonardo, Lorenzo Raggi, Carlo Tissi, Sally Howlett, Renzo Torricelli, and Valeria Negri. 2018. "Multi-Environment Evaluation and Genetic Characterisation of Common Bean Breeding Lines for Organic Farming Systems" Sustainability 10, no. 3: 777. https://doi.org/10.3390/su10030777
APA StyleCaproni, L., Raggi, L., Tissi, C., Howlett, S., Torricelli, R., & Negri, V. (2018). Multi-Environment Evaluation and Genetic Characterisation of Common Bean Breeding Lines for Organic Farming Systems. Sustainability, 10(3), 777. https://doi.org/10.3390/su10030777