Combining High Yields and Blast Resistance in Rice (Oryza spp.): A Screening under Upland and Lowland Conditions in Benin


 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Field Trials for Agronomic Evaluation
2.3. Data Collection
2.4. Statistical Analysis
3. Results
3.1. Phenotypic Variability for 15 Agronomic Measured Traits
3.2. Performance Evaluation of 42 Rice Accessions and 7 Reference Varieties for Yield and Yield Components in Upland Conditions
3.3. Performance Evaluation of 37 Rice Accessions and 7 Reference Varieties for Yield and Yield Components in Lowland Conditions
3.4. Grain Yield Performance of the Rice Accessions in Lowland and Upland Agro-Ecology
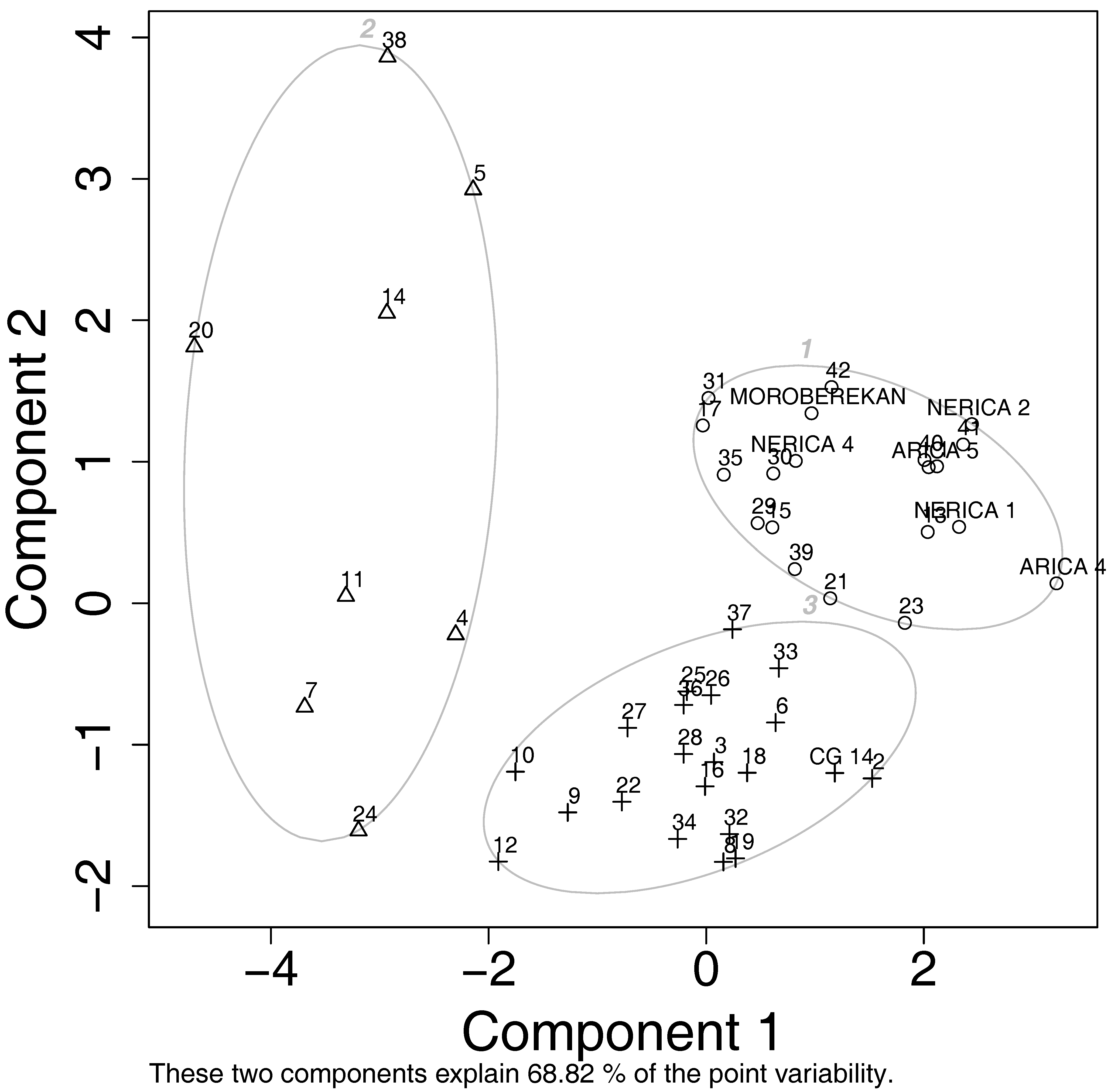
3.5. Relationships of Yield Components with Blast Resistance and Genetic Population Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| S/N | Accessions Number | Accession Name | Species Name | Country of Origin | Blast Resistance Pattern * |
|---|---|---|---|---|---|
| 1 | WAB0015772 | BEN11-126-A | O. sativa | Benin | Resistant |
| 2 | WAB0006684 | BEN11-45 | O. sativa | Benin | Resistant |
| 3 | WAB0035038 | BEN11-84-A | O. sativa | Benin | Resistant |
| 4 | WAB0035055 | BEN11-90-A | O. sativa | Benin | Resistant |
| 5 | WAB0035059 | BEN11-71-A | O. sativa | Benin | Resistant |
| 6 | WAB0029182 | DION KIVO | O. glaberrima | Mali | Resistant |
| 7 | WAB0029335 | ISSABGO | O. glaberrima | Nigeria | Moderately resistant |
| 8 | WAB0030263 | MACINA-B | O. glaberrima | Nigeria | Susceptible |
| 9 | WAB0023837 | RAM 100 | O. glaberrima | Mali | Resistant |
| 10 | WAB0024105 | RAM 131 | O. glaberrima | Mali | Resistant |
| 11 | WAB0024116 | RAM 154 | O. glaberrima | Mali | Resistant |
| 12 | WAB0029194 | TIERO OUSSI DIOUBOU KOUNTI | O. glaberrima | Mali | Resistant |
| 13 | WAB0032487 | TIOLY | O. glaberrima | Mali | Moderately resistant |
| 14 | WAB0032298 | TIRO SI DIOUMDOU | O. glaberrima | Mali | Resistant |
| 15 | WAB0008589 | TOG 5380 | O. glaberrima | Nigeria | Moderately resistant |
| 16 | WAB0020477 | TOG 5392 | O. glaberrima | Nigeria | Moderately resistant |
| 17 | WAB0032397 | TOG 5439-C | O. glaberrima | Nigeria | Resistant |
| 18 | WAB0029323 | TOG 5480 | O. glaberrima | Nigeria | Moderately resistant |
| 19 | WAB0029333 | TOG 5509 | O. glaberrima | Nigeria | Moderately resistant |
| 20 | WAB0019882 | TOG 5538 | O. glaberrima | Nigeria | Resistant |
| 21 | WAB0029315 | TOG 5673 | O. glaberrima | Nigeria | Moderately resistant |
| 22 | WAB0020505 | TOG 5693 | O. glaberrima | Nigeria | Moderately susceptible |
| 23 | WAB0032497 | TOG 5786-A | O. glaberrima | Liberia | Resistant |
| 24 | WAB0015703 | TOG 5951 | O. glaberrima | Nigeria | Resistant |
| 25 | WAB0001360 | TOG 5978 | O. glaberrima | Nigeria | Susceptible |
| 26 | WAB0029342 | TOG 6029 | O. glaberrima | Nigeria | Susceptible |
| 27 | WAB0032230 | TOG 6142-A | O. glaberrima | Nigeria | Susceptible |
| 28 | WAB0008937 | TOG 6201 | O. glaberrima | Guinea | Resistant |
| 29 | WAB0032848 | TOG 6228-A | O. glaberrima | Mali | Resistant |
| 30 | WAB0032550 | TOG 6804-A | O. glaberrima | Nigeria | Resistant |
| 31 | WAB0026783 | TOG 7106 | O. glaberrima | Mali | Resistant |
| 32 | WAB0002093 | TOG 7183 | O. glaberrima | Mali | Moderately resistant |
| 33 | WAB0002136 | TOG 7232 | O. glaberrima | Mali | Resistant |
| 34 | WAB0002143 | TOG 7239 | O. glaberrima | Mali | Resistant |
| 35 | WAB0002145 | TOG 7243 | O. glaberrima | Mali | Moderately susceptible |
| 36 | WAB0032345 | TOG 7250-A | O. glaberrima | Mali | Moderately resistant |
| 37 | WAB0009280 | TOG 7393 | O. glaberrima | Nigeria | Moderately susceptible |
| 38 | WAB0015043 | TOS 16746 | O. glaberrima | Ivory Coast | Resistant |
| 39 | WAB0026176 | TOS 6447 | O. glaberrima | Mali | Resistant |
| 40 | WAB0032495 | TOS 6454-A | O. glaberrima | Liberia | Resistant |
| 41 | WAB0008956 | TOS 6457 | O. glaberrima | Liberia | Resistant |
| 42 | WAB0032394 | W 1032 | O. glaberrima | Nigeria | Resistant |
| Traits | Codes Used | Description |
|---|---|---|
| Total number of tillers/plant | Tillers_Total | Counted number of tillers at the maturity stage |
| Number of fertile tillers/plant | Tillers_Fertile | Counted fertile tillers at the maturity stage |
| Percentage of fertile tillers/plant | % Fertile_Tillers | Calculated in Excel |
| Panicle length | Length_Pan | Measured from the base of the lowest spikelet to the tip of the latest spikelet on the panicle, excluding awn |
| Plant height at maturity (cm) | Plant_height | Measured height from the base of the plant to the top of the latest spikelet on the panicle, excluding awn |
| Total number of spikelets/panicle | Spikelets_TotalNum | Counted total number of grains in sampled panicles |
| Number of filled spikelets/panicle | Filled_Spikelets | Counted filled grains in sampled panicles |
| Percentage of filled spikelets | % Fertile_Spikelets | Calculated in Excel |
| Number of primary branching | Ram_Iaire | Counted number of primary branches of the panicle |
| Number of secondary branching | Ram_IIaire | Counted number of secondary branches of the panicle |
| Ratio secondary branching/primary branching | Ratio_RamIIRamIaire | Calculated in Excel |
| Total number of panicles/m2 | Panicules_Num | Counted the number of panicles per square meter |
| Number of days to 80% flowering | Number of days to 80% flowering (CSE) | Recorded the number of days from effective seeding to 80% heading |
| Number of days to 80% maturity | Number of days to 80% maturity (CSM) | Recorded the number of days from effective seeding to 80% maturity |
| Grain yield (g/36 plants) | Yield | Harvested a square meter (1 m × 1 m) of each plot |

References
- Dipti, S.S.; Bergman, C.; Indrasari, S.D.; Herath, T.; Hall, R.; Lee, H.; Habibi, F.; Bassinello, P.Z.; Gratero, E.; Ferraz, J.; et al. The potential of rice to offer solutions for malnutrition and chronic diseases. Rice 2012, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, V.; Sie, M.; Hijmmans, R.J.; Otsuka, K. Increasing rice production in sub-Saharan Africa: Challenges and opportunities. Adv. Agron. 2007, 94, 55–133. [Google Scholar]
- Seck, P.A.; Diagne, A.; Mohanty, S.; Wopereis, M.C.S. Crops that feed the world 7: Rice. Food Secur. 2012, 4, 7–24. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Organisation of the United Nations; FAOSTAT Statistics Database: Rome, Italy. Available online: http://faostat3.fao.org/ (accessed on 26 June 2018).
- Seck, P.; Touré, A.; Coulibaly, J.; Diagne, A.; Wopereis, M. Africa’s rice economy before and after the 2008 rice crisis. In Realizing Africa’s Rice Promise; CABI: Wallingford, UK, 2013; 138p. [Google Scholar]
- OECD/Food and Agriculture Organization of the United Nations. OECD-FAO Agricultural Outlook 2015; OECD Publishing: Paris, France, 2015. [Google Scholar] [CrossRef]
- Edgerton, M.D. Increasing Crop Productivity to Meet Global Needs for Feed, Food, and Fuel. Plant Physiol. 2009, 149, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Pham, J.L. Evaluation des Ressources génétiques des riz Cultivés en Afrique par Hybridation intra et Interspecifique. Ph.D. Thesis, Université de Paris XI ORSAY, Paris, France, 1992; 236p. [Google Scholar]
- Jones, M.P.; Dingkuhn, M.; Aluko, G.K.; Semon, M. Interspecific Oryza Sativa L. X O. Glaberrima Steud. Progenies in upland rice improvement. Euphytica 1997, 92, 237–246. [Google Scholar] [CrossRef]
- Futakuchi, K.; Sié, M. Better exploitation of African rice (Oryza glaberrima Steud.) in varietal development for resource-poor farmers in West and Central Africa. Agric. J. 2009, 4, 96–102. [Google Scholar]
- Sié, M.; Ogunbayo, S.A.; Dakouo, D.; Sanou, I.; Dembélé, Y.; N’dri, B.; Dramé, K.N.; Sanni, K.A.; Toulou, B.; Glele, R.K. Evaluation of intra and interspecific rice varieties adapted to valley bottom conditions in Burkina Faso. Afr. J. Plant Sci. 2010, 4, 308–318. [Google Scholar]
- Thiémélé, D.; Boisnard, A.; Ndjiondjop, M.; Chéron, S.; Séré, Y.; Aké, S.; Ghesquière, A.; Albar, L. Identification of a second major resistance gene to Rice yellow mottle virus, RYMV2, in the African cultivated rice species, O. glaberrima. Theor. Appl. Genet. 2010, 121, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Adegbola, P.Y.; Arouna, A.; Diagne, A. Evaluation de l’impact économique des nouvelles variétés de riz NERICA au Bénin: Une évidence avec les modèles basés sur l’approche “contre factuel”. Paper presented at the First Africa Rice Congress, Dar es Salaam, Tanzania, 31 July–4 August 2006. [Google Scholar]
- Rodenburg, J.; Diagne, A.; Oikeh, S.; Futakuchi, K.; Kormawa, P.M.; Semon, M.; Akintayo, I.; Cissé, B.; Sié, M.; Narteh, L.T.; et al. Achievements and impact of NERICA on sustainable rice production in sub-Saharan Africa. Int. Rice Comm. Newslett. 2006, 55, 45–58. Available online: https://www.researchgate.net/profile/Jonne_Rodenburg/publication/313078439_Achievements_and_impact_of_NERICA_on_sustainable_rice_production_in_subSaharan_Africa/links/58b04e0d45851503be97dbbc/Achievements-and-impact-of-NERICA-on-sustainable-rice-production-in-sub-Saharan-Africa.pdf (accessed on 1 July 2018).
- Kijima, Y.; Otsuka, K.; Sserunkuuma, D. ‘Assessing the impact of NERICA on income and poverty in central and western Uganda’. Agric. Econ. 2008, 38, 327–337. [Google Scholar] [CrossRef]
- Nguezet, P.M.D.; Diagne, A.; Okoruwa, V.O.; Ojehomon, V. Impact of improved rice technology on income and poverty among rice farming household in nigeria: A local average treatment effect (late) approach. Q. J. Int. Agric. 2011, 50, 267–291. [Google Scholar]
- Dibba, L.; Zeller, M.; Diagne, A. The impact of new Rice for Africa (NERICA) adoption on household food security and health in the Gambia. Food Secur. 2017, 9, 929–944. [Google Scholar] [CrossRef]
- Wang, X.; Jia, M.H.; Ghai, P.; Lee, F.N.; Jia, Y. Genome-wide association of rice blast disease resistance ad yield-related components of rice. MPMI 2015, 28, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- MAEP: Ministere de L’Agriculture, de l’Elevage et de la Peche. Stratégie Nationale Pour le Développement de la Riziculture au Bénin; MAEP: Cotonou, Benin, 2010; 26p, Available online: http://extwprlegs1.fao.org/docs/pdf/ben149179.pdf (accessed on 5 May 2018).
- FAOSTAT: Food and Agriculture Organisation of the United Nations. FAOSTAT Statistics Database: Rome, Italy. Available online: http://faostat.fao.org/ (accessed on 2 June 2018).
- Adégbola, P.Y.; Singbo, A.G. Impact de L’importation du riz sur la Compétitivité et la Rentabilité de la Production Nationale au Bénin. Communication à L’atelier Régional de L’ADRAO sur le Thème: Politique et Stratégies Pour la Promotion de la Production Rizicole et la Sécurité Alimentaire en Afrique. November 2005. Available online: http://www.brmnbenin.org/base/docs_de_rech/Impact_de_l_importation_du_riz_sur_la_competitivite_et_la_rentabilite_de_la_production_nationale_au_Benin_Adegbola_Benin.pdf (accessed on 2 June 2018).
- Diagne, A.; Amovin-Assagba, E.; Koichi, F.; Wopereis, M.C.S. Estimation of cultivated area; number of farming households and yield for major rice-growing environments in Africa. In Realizing Africa’s Rice Promise; Wopereis, M.C.S., Johnson, D.E., Ahmadi, N., Tollens, E., Jalloh, A., Eds.; CAB International: Boston, MA, USA, 2013; pp. 35–45. [Google Scholar]
- FAOSTAT Food and Agriculture Organisation of the United Nations. GIEWS—Global Information and Early Warning System; FAOSTAT Statistics Database: Rome, Italy, 2018; Available online: http://www.fao.org/giews/countrybrief/country.jsp?code=BEN (accessed on 16 May 2018).
- Odjo, T.C.; Dossou-Aminon, I.; Dansi, A.; Djengue, W. Diversity, Genetic Erosion and Participatory Evaluation of Rice (Oryza sativa L. and Oryza glaberrima Steud.) Varieties in Benin. Int. J. Curr. Res. Biosci. Plant Biol. 2017, 4, 147–164. [Google Scholar] [CrossRef]
- Yokouchi, T.; Saito, K. Why did farmers stop cultivating NERICA upland rice varieties in central Benin? Int. J. Agric. Sustain. 2017, 15, 724–734. [Google Scholar] [CrossRef]
- Couch, B.C.; Kohn, L.M. A multilocus gene genealogy concordant with host preference indicates segregation of a new species, Magnaporthe oryzae, from M. grisea. Mycologia 2002, 94, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Afouda, L.; Gnikpo, S.; Sere, Y.; Gumedzoe, Y. Prevalence des principales maladies sur des variétes de riz cultivees dans le departement de l’alibori au Nord-Benin. J. de la Recherche Scientifique de l’Universite de Lome 2007, 9, 1027–1988. [Google Scholar]
- Vodouhe, S.R.; Ojegui, M.; Amadji, F. Impact of blast on rice cultivation in People’s Republic of Benin. In Proceedings of the Symposium on Rice Resistance to Blast, Montpellier, France, 18–21 March 1981; Service de Pathologie Végétale: Montpellier, France, 1981; pp. 27–33. [Google Scholar]
- AfricaRice. Lessons from the Rice Crisis: Policies for Food Security in Africa; Africa Rice Center: Cotonou, Benin, 2011. [Google Scholar]
- Norman, J.C.; Kebe, B. African Smallholder Farmers: Rice Production and Sustainable Livelihoods; International Rice Commission Newsletter (FAO): Accra, Ghana, 2006. [Google Scholar]
- Yelome, O.I.; Audenaert, K.; Landschoot, S.; Dansi, A.; Vanhove, W.; Silue, D.; Van Damme, P.; Haesaert, G. Exploring genetic diversity and disease response of cultivated rice accessions against Pyricularia oryzae under rainfed upland conditions in Benin. Genet. Resour. Crop Evol. 2018, 65, 1615–1624. [Google Scholar] [CrossRef]
- Ogunbayo, S.A.; Sie, M.; Odjo, D.K.; Sanni, K.A.; Akinwale, M.G.; Toulou, B.; Shittu, A.; Idehen, E.O.; Popoola, A.R.; Daniel, I.O.; et al. Genetic variation and heritability of yield and related traits in promising rice genotypes (Oryza sativa). J. Plant Breed. Crop Sci. 2014, 6, 153–159. [Google Scholar]
- Acquaah, G. Principles of Plant Genetics and Breeding; Blackwell Publishing: Oxford, UK, 2007. [Google Scholar]
- Ruelle, P.; Bruggers, R.L. Traditional approaches for protecting cereal crops from birds in Africa. In Vertebrate Pest Conference Proceedings Collection, Proceedings of the Tenth Vertebrate Pest Conference, Monterey, CA, USA, 23–25 February 1982; Marsh, R.E., Ed.; University of California: Nebraska, CA, USA, 1982; pp. 80–86. Available online: https://digitalcommons.unl.edu/cgi/viewcontent.cgi?article=1036&context=vpc10 (accessed on 2 June 2018).
- Tréca, B. Les dégâts d’oiseaux d’eau sur les rizières aménagées du delta central du Niger au Mali. J. D’agriculture Traditionnelle et de Botanique Appliquée 1987, 34, 153–170. [Google Scholar] [CrossRef]
- Montcho, D.; Futakuchi, K.; Agbangla, C.; Semon, M.; Dieng, I.; Sie, M. Morphological and phenological response of African rice (Oryza glaberrima Steud.) under different hydrological conditions. Asian J. Plant Sci. 2013, 12, 79–86. [Google Scholar] [CrossRef]
- Zhao, M.; Sun, J.; Xiao, Z.; Cheng, F.; Xu, H.; Tang, L.; Chen, W.; Xu, Z.; Xu, Q. Variations in DENSE AND ERECT PANICLE 1 (DEP1) contribute to the diversity of the panicle trait in high-yielding japonica rice varieties in northern China. Breed. Sci. 2016, 66, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Ashikari, M.; Sakakibara, H.; Lin, S.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Cytokinin oxidase regulates rice grain production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Mokuwa, A.; Nuijten, E.; Okry, F.; Teeken, B.; Maat, H.; Richards, P.; Struik, P.C. Robustness and Strategies of Adaptation among Farmer Varieties of African Rice (Oryza glaberrima) and Asian Rice (Oryza sativa) across West Africa. PLoS ONE 2013, 8, e34801. [Google Scholar] [CrossRef] [PubMed]
- Lafitte, R.; Torres, R.; Chen, J. Managing drought tress in aerobic (Oryza sativa) rice systems. Philos. J. Crop Sci. 2003, 18, 21. [Google Scholar]
- Laborte, A.G.; Paguirigan, N.C.; Moya, P.F.; Nelson, A.; Sparks, A.H.; Gregorio, G.B. Farmers’ Preference for Rice Traits: Insights from Farm Surveys in Central Luzon, Philippines, 1966–2012. PLoS ONE 2015, 10, e0136562. [Google Scholar] [CrossRef] [PubMed]
- Richards, P. Coping with Hunger: Hazard and Experiment in an African Rice-Farming System; Allen and Unwin: London, UK, 1986. [Google Scholar]
- Sarla, N.; Swamy, B.P.M. Oryza glaberrima: A source for the improvement of Oryza sativa. Curr. Sci. 2005, 89, 955–963. [Google Scholar]
- Ndjiondjop, M.N.; Seck, P.A.; Lorieux, M.; Futakuchi, K.; Yao, K.N.; Djedatin, G.; Sow, M.E.; Bocco, R.; Cisse, F.; Fatondji, B. Effect of drought on Oryza glaberrima rice accessions and Oryza glaberrima derived-lines. Asian J. Agric. Res. 2012, 6, 144–157. [Google Scholar] [CrossRef]
- Linares, O.F. African rice (Oryza glaberrima): History and future potential. Proc. Natl. Acad. Sci. USA 2002, 99, 16360–16365. [Google Scholar] [CrossRef] [PubMed]
- Odjo, T.; Ahohuendo, B.C.; Onasanya, A.; Akator, K.; Séré, Y. Analysis of Magnaporthe oryzae population structure in Benin. Afr. J. Agric. Res. 2011, 6, 6183–6188. [Google Scholar] [CrossRef]
- Zhu, M.; Wang, L.; Pan, Q. Identification and characterization of a new blast resistance gene located on rice chromosome 1 through linkage and differential analyses. Phytopathology 2004, 94, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Roy-Chowdhury, M.; Jia, Y.; Jackson, A.; Jia, M.H.; Fjellstrom, R.; Cartwright, R.D. Analysis of rice blast resistance gene Pi-z in rice germplasm using pathogenicity assays and DNA markers. Euphytica 2012, 184, 35–47. [Google Scholar] [CrossRef]
- Yelome, O.I.; Audenaert, K.; Landschoot, S.; Dansi, A.; Vanhove, W.; Silue, D.; Van Damme, P.; Haesaert, G. Analysis of population structure and genetic diversity reveals gene flow and geographic patterns in cultivated rice (Oryza. sativa & O. glaberrima) in West Africa. Euphytica 2018. submitted. [Google Scholar]
- Sie, M.; Sanni, K.; Futakuchi, K.; Manneh, B.; Mande, S.; Vodouhe, R.; Dogbe, S.; Drame, K.; Ogunbayo, A.; Ndjiondjop, M.; et al. Towards a rational use of African rice (Oryza glaberrima Steud.) for breeding in Sub-Saharan Africa. Genes Gen. Genom. 2012, 6, 1–7. [Google Scholar]


| Trait | Lowland | Upland | ||||
|---|---|---|---|---|---|---|
| Access. (df = 48) | Rep. (df = 2) | Error (df = 96) | Access (df = 44) | Rep. (df = 2) | Error (df = 88) | |
| Tillers_Total | 19 *** | 9.99 * | 2.84 | 35 *** | 4.92 | 1.85 |
| Tillers_Fertile | 19 *** | 9.65 * | 2.98 | 34 *** | 3.59 | 2.29 |
| %Fertile_Tillers | 36.45 *** | 33.91 | 17.04 | 46.19 | 4.93 | 33.20 |
| Length_Pan | 37 *** | 31.31 | 16.53 | 17 *** | 1.48 | 1.22 |
| Plant_Height | 444 *** | 419.70 *** | 44.39 | 238 *** | 5.14 | 21.20 |
| Spikelets_TotalNum | 2180 *** | 313.1 | 233.8 | 2368 *** | 17.43 | 85.67 |
| Filled_Spikelets | 1804 *** | 93.39 | 248.77 | 2110 *** | 262.20 | 143.5 |
| %Fertile_Spikelets | 49.96 *** | 77.71 * | 16.69 | 355.37 *** | 246.43 ** | 37.45 |
| Ram_Iaire | 18 *** | 8.72 | 4.85 | 8 *** | 0.15 | 0.70 |
| Ram_IIaire | 197 *** | 4.88 | 16.38 | 184 *** | 9.64 | 8.15 |
| Ratio_RamIIRamIaire | 1.88 *** | 0.14 | 0.19 | 1.69 *** | 0.05 | 0.05 |
| Panicules_Num | 34,223 *** | 8434 | 3348 | 31,117 *** | 503.1 | 2141.7 |
| FLW | 440 *** | 36.33 | 12.16 | 1150 *** | 9.64 | 9.77 |
| MAT | 475 *** | 36.18 | 12.20 | 961 *** | 3.11 | 13.48 |
| Yield | 42,975 *** | 16,730 | 11,962 | 62,184 *** | 8141 | 5963 |
| Trait | Lowland | Upland | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Min. | Max. | Std. | Min. | Max. | SD | ||
| Tillers_Total | 5 | 20 | 2.89 | 3 | 21 | 3.56 | 0.4125 |
| Tillers_Fertile | 5 | 20 | 2.88 | 3 | 21 | 3.56 | 0.8075 |
| %Fertile_Tillers | 61.45 | 100.00 | 4.86 | 60.98 | 100.00 | 5.97 | 0.001 ** |
| Long_Pan | 15 | 69 | 4.85 | 18 | 30 | 2.52 | 0.8641 |
| Plant_Height | 76 | 134 | 13.46 | 82 | 135 | 9.60 | 0.5462 |
| Spikelets_TotalNumber | 54 | 185 | 29.57 | 62 | 193 | 28.90 | 0.0001 *** |
| Filled_Spikelets | 51 | 172 | 27.51 | 35 | 163 | 28.14 | 0.1483 |
| %Fertile_Spikelets | 72.21 | 98.65 | 5.34 | 29.42 | 98.82 | 12.03 | 0.0000 *** |
| Ram_Iaire | 4 | 29 | 3.02 | 6 | 16 | 1.75 | 0.1866 |
| Ram_IIaire | 1 | 39 | 8.70 | 2 | 40 | 8.13 | 0.0039 ** |
| Ratio_RamIIRamIaire | 0.02 | 3.35 | 0.86 | 0.16 | 3.20 | 0.77 | 0.0518 |
| Panicules_Num | 105 | 714 | 117.16 | 114 | 590 | 107.91 | 0.9754 |
| FLW | 75 | 136 | 12.63 | 64 | 156 | 19.74 | 0.0071 ** |
| MAT | 94 | 155 | 13.10 | 86 | 173 | 18.03 | 0.0208 * |
| Yield | 96 | 783 | 158.01 | 31 | 869 | 159.92 | 0.0002 *** |
| Traits | %Fertile_Tillers | Spikelets_TotalNumber | %Fertile Spikelets | Ram_IIaire | FLW | MAT | Yield |
|---|---|---|---|---|---|---|---|
| %Fertile_Tillers | 0.01 | −0.17 | −0.04 | 0.21 | 0.19 | −0.27 | |
| Spikelets_TotalNumber | 0.03 | −0.09 | 0.69 * | 0.01 | −0.01 | 0.09 | |
| %Fertile Spikelets | 0.12 | −0.12 | −0.03 | −0.70 * | −0.69 | 0.57 * | |
| Ram_IIaire | −0.16 | 0.61 * | −0.17 | −0.15 | −0.10 | 0.27 | |
| FLW | 0.16 | 0.17 | −0.41 * | −0.23 | 0.96 * | −0.52 * | |
| MAT | 0.1 | 0.12 | −0.44 * | −0.22 | 0.94 * | −0.49 * | |
| Yield | 0.17 | 0.08 | 0.06 | 0.24 | −0.05 | −0.03 |
| Ecology | Upland | Lowland | |||
|---|---|---|---|---|---|
| Traits | PCA 1 | PCA 2 | PCA 1 | PCA 2 | PCA 3 |
| %Fertile_Tillers | −0.20 | −0.11 | −0.74 * | ||
| Spikelets_TotalNumber | 0.69 * | 0.11 | 0.62 * | −0.07 | |
| %Fertile Spikelets | 0.47 * | 0.17 | −0.40 * | −0.22 | −0.32 |
| Ram_IIaire | 0.13 | 0.69 * | −0.13 | 0.70 * | 0.06 |
| FLW | −0.53 * | 0.63 * | −0.08 | ||
| MAT | −0.52 * | 0.63 * | −0.04 | ||
| Yield | 0.42 * | 0.11 | 0.26 | −0.58 * | |
| Upland | Lowland | |||||||
|---|---|---|---|---|---|---|---|---|
| Accessions | Species | Grain Yield | MAT | Rank | Grain Yield | MAT | Rank | Presumed Ecology |
| WAB0015772 | SATIVA | 257.35 a* | 172.33 a | 30 | 489.26 b | 140 b | 14 | Lowland |
| WAB0006684 | SATIVA | 319.56 a | 95.33 a | 25 | 168.46 a | 99 b | 35 | Upland |
| WAB0035055 | SATIVA | 494.39 a | 126 a | 5 | 569.18 a | 107.67 b | 4 | BOTH |
| WAB0035059 | SATIVA | 669.49 a | 124.67 a | 1 | 498.34 b | 107.67 b | 12 | Upland |
| WAB0029182 | GLA | 573.59 a | 117.33 a | 3 | 510.32 a | 108 b | 11 | BOTH |
| WAB0029335 | GLA | 338.57 a | 132.67 a | 21 | 474.73 a | 118.67 b | 16 | BOTH |
| WAB0030263 | GLA | 327.22 a | 103 a | 23 | 168.28 a | 94 b | 36 | Upland |
| WAB0023837 | GLA | 395.73 a | 134.67 a | 13 | 555.75 a | 125.67 b | 6 | BOTH |
| WAB0024105 | GLA | 100.09 a | 140 a | 35 | 358.15 b | 145 a | 27 | Lowland |
| WAB0024116 | GLA | 322.16 a | 157 a | 24 | 421.12 a | 135 b | 25 | Lowland |
| WAB0029194 | GLA | 603.35 a | 129.67 a | 2 | 476.51 a | 128.33 a | 15 | BOTH |
| WAB0032487 | GLA | 190.69 a | 168.33 a | 33 | 258.56 a | 151.67 b | 34 | Lowland |
| WAB0032298 | GLA | 371.9 a | 132 a | 17 | 343.35 a | 126 a | 29 | BOTH |
| WAB0008589 | GLA | 409.93 a | 123.67 a | 12 | 471.15 a | 117.33 b | 18 | BOTH |
| WAB0020477 | GLA | 425.17 a | 127.33 a | 9 | 295.3 a | 120.67 b | 32 | Upland |
| WAB0029323 | GLA | 363.39 a | 137.67 a | 19 | 427.34 a | 127.33 a | 24 | BOTH |
| WAB0019882 | GLA | 212.5 a | 135.33 a | 32 | 567.78 b | 127 b | 5 | Lowland |
| WAB0029315 | GLA | 259.95 a | 131.33 a | 29 | 493.8 b | 122 a | 13 | Lowland |
| WAB0020505 | GLA | 269.31 a | 133.33 a | 26 | 430.31 a | 135.67 a | 21 | BOTH |
| WAB0032497 | GLA | 469.19 a | 129.67 a | 8 | 446.11 a | 129 a | 20 | BOTH |
| WAB0015703 | GLA | 374.68 a | 115 a | 16 | 526.31 b | 112.67 a | 8 | Lowland |
| WAB0001360 | GLA | 411.85 a | 131 a | 10 | 427.65 a | 124 b | 23 | BOTH |
| WAB0029342 | GLA | 365.09 a | 132 a | 18 | 644.95 b | 135 a | 2 | Lowland |
| WAB0008937 | GLA | 491.98 a | 123.67 a | 6 | 536.48 a | 107.67 b | 7 | BOTH |
| WAB0032848 | GLA | 268.96 a | 138.33 a | 27 | 260.76 a | 136 a | 33 | BOTH |
| WAB0032550 | GLA | 220.5 a | 125 a | 31 | 318.77 a | 120 a | 31 | BOTH |
| WAB0002093 | GLA | 98.18 a | 150.67 a | 36 | 333 b | 150 a | 30 | Lowland |
| WAB0002136 | GLA | 103.2 a | 147.33 a | 34 | 155.56 a | 143 b | 37 | BOTH |
| WAB0002143 | GLA | 540.5 a | 112.67 a | 4 | 358.03 b | 111.33 a | 28 | Upland |
| WAB0002145 | GLA | 263.51 a | 147.33 a | 28 | 454.26 a | 140 b | 19 | Lowland |
| WAB0032345 | GLA | 333.42 a | 125.67 a | 22 | 512.78 a | 117.33 b | 9 | BOTH |
| WAB0009280 | GLA | 348.65 a | 122.67 a | 20 | 510.61 b | 126.5 a | 10 | Lowland |
| WAB0015043 | GLA | 410.95 a | 126.33 a | 11 | 600.05 a | 127 a | 3 | BOTH |
| WAB0026176 | GLA | 375.64 a | 125.33 a | 15 | 429.71 a | 120.67 a | 22 | BOTH |
| WAB0032495 | GLA | 389.67 a | 127.33 a | 14 | 413.82 a | 125.67 a | 26 | BOTH |
| WAB0008956 | GLA | 469.41 a | 132 a | 7 | 698.9 b | 124 b | 1 | Lowland |
| WAB0032394 | GLA | 51.07 a | 169.67 a | 37 | 471.73 b | 143 b | 17 | Lowland |
| WAB0035038 | SATIVA | 717.18 | 134.67 | - | - | - | - | Upland |
| WAB0029333 | GLA | 238.42 | 125.33 | - | - | - | - | - |
| WAB0032230 | GLA | 50.63 | 146 | - | - | - | - | - |
| WAB0032397 | GLA | 346.98 | 129.67 | - | - | - | - | - |
| WAB0026783 | GLA | 270.22 | 152.33 | - | - | - | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yelome, O.I.; Audenaert, K.; Landschoot, S.; Dansi, A.; Vanhove, W.; Silue, D.; Damme, P.V.; Haesaert, G. Combining High Yields and Blast Resistance in Rice (Oryza spp.): A Screening under Upland and Lowland Conditions in Benin. Sustainability 2018, 10, 2500. https://doi.org/10.3390/su10072500
Yelome OI, Audenaert K, Landschoot S, Dansi A, Vanhove W, Silue D, Damme PV, Haesaert G. Combining High Yields and Blast Resistance in Rice (Oryza spp.): A Screening under Upland and Lowland Conditions in Benin. Sustainability. 2018; 10(7):2500. https://doi.org/10.3390/su10072500
Chicago/Turabian StyleYelome, Octaviano Igor, Kris Audenaert, Sofie Landschoot, Alexandre Dansi, Wouter Vanhove, Drissa Silue, Patrick Van Damme, and Geert Haesaert. 2018. "Combining High Yields and Blast Resistance in Rice (Oryza spp.): A Screening under Upland and Lowland Conditions in Benin" Sustainability 10, no. 7: 2500. https://doi.org/10.3390/su10072500
APA StyleYelome, O. I., Audenaert, K., Landschoot, S., Dansi, A., Vanhove, W., Silue, D., Damme, P. V., & Haesaert, G. (2018). Combining High Yields and Blast Resistance in Rice (Oryza spp.): A Screening under Upland and Lowland Conditions in Benin. Sustainability, 10(7), 2500. https://doi.org/10.3390/su10072500