Soil Microbial Communities and Enzyme Activities after Long-Term Application of Inorganic and Organic Fertilizers at Different Depths of the Soil Profile

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Sampling and Soil Parameters
2.4. Phosphilipid Fatty Acid Analyses
2.5. Soil Enzyme Assays
2.6. Data Analysis
3. Results and Discussion
3.1. Structure of Soil Microbial Communities
3.2. Effects of Fertilizers on Microbial Communities
3.3. Analysis of Data Variability
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Drinkwater, L.E.; Letourneau, D.K.; Workneh, F.; Vanbruggen, A.H.C.; Shennan, C. Fundamental differences between conventional and organic tomato agroecosystems in California. Ecol. Appl. 1995, 5, 1098–1112. [Google Scholar] [CrossRef]
- Vasseur, C.; Joannon, A.; Aviron, S.; Burel, F.; Meynard, J.M.; Baudry, J. The cropping systems mosaic: How does the hidden heterogeneity of agricultural landscapes drive arthropod populations? Agric. Ecosyst. Environ. 2013, 166, 3–14. [Google Scholar] [CrossRef]
- Acosta-Martinez, V.; Acosta-Mercado, D.; Sotomayor-Ramirez, D.; Cruz-Rodriguez, L. Microbial communities and enzymatic activities under different management in semiarid soils. Appl. Soil Ecol. 2008, 38, 249–260. [Google Scholar] [CrossRef]
- Schipanski, M.E.; Drinkwater, L.E. Nitrogen fixation in annual and perennial legume-grass mixtures across a fertility gradient. Plant Soil 2012, 357, 147–159. [Google Scholar] [CrossRef]
- Bowles, T.M.; Acosta-Martinez, V.; Calderon, F.; Jackson, L.E. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Stamenov, D.; Đurić, S.; Jafari, T.H.; Ćirić, V.; Manojlović, M. Microbiological Activity in the Soil of Various Agricultural Crops in Organic Production. Contemp. Agric. 2018, 67, 34–39. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; He, Z.; Wilson, M.J.; Campbell, C.D. Microbial biomass and community structure in a sequence of soils with increasing fertility and changing land use. Microb. Ecol. 2000, 40, 223–237. [Google Scholar]
- Rochette, P.; Angers, D.A.; Flanagan, L.B. Maize residue decomposition measurement using soil surface carbon dioxide fluxes and natural abundance of carbon-13. Soil Sci. Soc. Am. J. 1999, 63, 1385–1396. [Google Scholar] [CrossRef]
- Gong, W.; Yan, X.Y.; Wang, J.Y.; Hu, T.X.; Gong, Y.B. Long-term manure and fertilizer effects on soil organic matter fractions and microbes under a wheat-maize cropping system in northern China. Geoderma 2009, 149, 318–324. [Google Scholar] [CrossRef]
- Guo, S.; Wu, J.; Coleman, K.; Zhu, H.; Li, Y.; Liu, W. Soil organic carbon dynamics in a dryland cereal cropping system of the Loess Plateau under long-term nitrogen fertilizer applications. Plant Soil 2012, 353, 321–332. [Google Scholar] [CrossRef]
- Smith, J.; Smith, P.; Wattenbach, M.; Zaehle, S.; Hiederer, R.; Jones, R.J.A.; Montanarella, L.; Rounsevell, M.D.A.; Reginster, I.; Ewert, F. Projected changes in mineral soil carbon of European croplands and grasslands, 1990–2080. Glob. Chang. Biol. 2005, 11, 2141–2152. [Google Scholar] [CrossRef]
- Ludwig, B.; Geisseler, D.; Michel, K.; Joergensen, R.G.; Schulz, E.; Merbach, I.; Raupp, J.; Rauber, R.; Hu, K.; Niu, L.; et al. Effects of fertilization and soil management on crop yields and carbon stabilization in soils. A review. Agron. Sustain. Dev. 2011, 31, 361–372. [Google Scholar] [CrossRef]
- Malhi, S.S.; Nyborg, M.; Goddard, T.; Puurveen, D. Long-term tillage, straw management and N fertilization effects on quantity and quality of organic C and N in a Black Chernozem soil. Nutr. Cycl. Agroecosyst. 2011, 90, 227–241. [Google Scholar] [CrossRef]
- Grandy, A.S.; Salam, D.S.; Wickings, K.; McDaniel, M.D.; Culman, S.W.; Snapp, S.S. Soil respiration and litter decomposition responses to nitrogen fertilization rate in no-till corn systems. Agric. Ecosyst. Environ. 2013, 179, 35–40. [Google Scholar] [CrossRef]
- Zhao, S.C.; Li, K.J.; Zhou, W.; Qiu, S.J.; Huang, S.W.; He, P. Changes in soil microbial community, enzyme activities and organic matter fractions under long-term straw return in north-central China. Agric. Ecosyst. Environ. 2016, 216, 82–88. [Google Scholar] [CrossRef]
- Nemergut, D.R.; Townsend, A.R.; Sattin, S.R.; Freeman, K.R.; Fierer, N.; Neff, J.C.; Bowman, W.D.; Schadt, C.W.; Weintraub, M.N.; Schmidt, S.K. The effects of chronic nitrogen fertilization on alpine tundra soil microbial communities: Implications for carbon and nitrogen cycling. Environ. Microbiol. 2008, 10, 3093–3105. [Google Scholar] [CrossRef] [PubMed]
- Jangid, K.; Williams, M.A.; Franzluebbers, A.J.; Sanderlin, J.S.; Reeves, J.H.; Jenkins, M.B.; Endale, D.M.; Coleman, D.C.; Whitman, W.B. Relative impacts of land-use, management intensity and fertilization upon soil microbial community structure in agricultural systems. Soil Biol. Biochem. 2008, 40, 2843–2853. [Google Scholar] [CrossRef]
- Ding, J.L.; Jiang, X.; Ma, M.C.; Zhou, B.K.; Guan, D.W.; Zhao, B.S.; Zhou, J.; Cao, F.M.; Li, L.; Li, J. Effect of 35 years inorganic fertilizer and manure amendment on structure of bacterial and archaeal communities in black soil of northeast China. Appl. Soil Ecol. 2016, 105, 187–195. [Google Scholar] [CrossRef]
- Simonsen, A.K.; Han, S.; Rekret, P.; Rentschler, C.S.; Heath, K.D.; Stinchcombe, J.R. Short-term fertilizer application alters phenotypic traits of symbiotic nitrogen fixing bacteria. PeerJ 2015, 3, e1291. [Google Scholar] [CrossRef] [PubMed]
- Hu, A.; Lu, Y.H. The differential effects of ammonium and nitrate on methanotrophs in rice field soil. Soil Biol. Biochem. 2015, 85, 31–38. [Google Scholar] [CrossRef]
- Fan, F.L.; Li, Z.J.; Wakelin, S.A.; Yu, W.T.; Liang, Y.C. Mineral fertilizer alters cellulolytic community structure and suppresses soil cellobiohydrolase activity in a long-term fertilization experiment. Soil Biol. Biochem. 2012, 55, 70–77. [Google Scholar] [CrossRef]
- Tian, W.; Wang, L.; Li, Y.; Zhuang, K.M.; Li, G.; Zhang, J.B.; Xiao, X.J.; Xi, Y.G. Responses of microbial activity, abundance, and community in wheat soil after three years of heavy fertilization with manure-based compost and inorganic nitrogen. Agric. Ecosyst. Environ. 2015, 213, 219–227. [Google Scholar] [CrossRef]
- Cusack, D.F.; Silver, W.L.; Torn, M.S.; Burton, S.D.; Firestone, M.K. Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests. Ecology 2011, 92, 621–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romaniuk, R.; Giuffre, L.; Costantini, A.; Nannipieri, P. Assessment of soil microbial diversity measurements as indicators of soil functioning in organic and conventional horticulture systems. Ecol. Indic. 2011, 11, 1345–1353. [Google Scholar] [CrossRef]
- Marschner, P.; Umar, S.; Baumann, K. The microbial community composition changes rapidly in the early stages of decomposition of wheat residue. Soil Biol. Biochem. 2011, 43, 445–451. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Tiemann, L.K.; Billings, S.A. Indirect Effects of Nitrogen Amendments on Organic Substrate Quality Increase Enzymatic Activity Driving Decomposition in a Mesic Grassland. Ecosystems 2011, 14, 234–247. [Google Scholar] [CrossRef]
- Wang, R.Z.; Dorodnikov, M.; Yang, S.; Zhang, Y.Y.; Filley, T.R.; Turco, R.F.; Zhang, Y.G.; Xu, Z.W.; Li, H.; Jiang, Y. Responses of enzymatic activities within soil aggregates to 9-year nitrogen and water addition in a semi-arid grassland. Soil Biol. Biochem. 2015, 81, 159–167. [Google Scholar] [CrossRef]
- Li, W.T.; Liu, M.; Jiang, C.Y.; Wu, M.; Chen, X.F.; Ma, X.Y.; Li, Z.P. Changes in soil aggregate-associated enzyme activities and nutrients under long-term chemical fertilizer applications in a phosphorus-limited paddy soil. Soil Use Manag. 2017, 33, 25–33. [Google Scholar] [CrossRef]
- Mehlich, A. New extractant for soil test evaluation of phosphorus, potassium, magnesium, calcium, sodium, manganese and zinc. Commun. Soil Sci. Plant Anal. 1978, 9, 477–492. [Google Scholar] [CrossRef]
- Kukla, J.; Holec, M.; Trogl, J.; Holcova, D.; Hofmanova, D.; Kuran, P.; Popelka, J.; Pacina, J.; Krizenecka, S.; Ust’ak, S.; et al. Tourist Traffic Significantly Affects Microbial Communities of Sandstone Cave Sediments in the Protected Landscape Area “Labske Piskovce” (Czech Republic): Implications for Regulatory Measures. Sustainability 2018, 10. [Google Scholar] [CrossRef]
- International Standard Organization. Soil Quality—Determination of Soil Microbial Diversity—Part 2: Method by Phospholipid Fatty Acid Analysis (PLFA) Using the Simple PLFA Extraction Method; International Standard Organization: Geneva, Switzerland, 2011; Volume ISO/TS 29843-2. [Google Scholar]
- Zelles, L.; Bai, Q.Y.; Rackwitz, R.; Chadwick, D.; Beese, F. Determination of phospholipid-derived and lipopolysaccharide-derived fatty-acids as an estimate of microbial biomass and community structures in soils. Biol. Fertil. Soils 1995, 19, 115–123. [Google Scholar] [CrossRef]
- Nebeska, D.; Trogl, J.; Pidlisnyuk, V.; Popelka, J.; Danova, P.V.; Ust’ak, S.; Honzik, R. Effect of Growing Miscanthus x giganteus on Soil Microbial Communities in Post-Military Soil. Sustainability 2018, 10. [Google Scholar] [CrossRef]
- Kuran, P.; Trogl, J.; Novakova, J.; Pilarova, V.; Danova, P.; Pavlorkova, J.; Kozler, J.; Novak, F.; Popelka, J. Biodegradation of Spilled Diesel Fuel in Agricultural Soil: Effect of Humates, Zeolite, and Bioaugmentation. Sci. World J. 2014. [Google Scholar] [CrossRef]
- Kakosova, E.; Hrabak, P.; Cernik, M.; Novotny, V.; Czinnerova, M.; Trogl, J.; Popelka, J.; Kuran, P.; Zoubkova, L.; Vrtoch, L. Effect of various chemical oxidation agents on soil microbial communities. Chem. Eng. J. 2017, 314, 257–265. [Google Scholar] [CrossRef]
- Trogl, J.; Pavlorkova, J.; Packova, P.; Sejak, J.; Kuran, P.; Popelka, J.; Pacina, J. Indication of Importance of Including Soil Microbial Characteristics into Biotope Valuation Method. Sustainability 2016, 8. [Google Scholar] [CrossRef]
- Federici, E.; Giubilei, M.A.; Cajthaml, T.; Petruccioli, M.; D’Annibale, A. Lentinus (Panus) tigrinus augmentation of a historically contaminated soil: Matrix decontamination and structure and function of the resident bacterial community. J. Hazard. Mater. 2011, 186, 1263–1270. [Google Scholar] [CrossRef]
- Moore-Kucera, J.; Dick, R.P. PLFA profiling of microbial community structure and seasonal shifts in soils of a Douglas-fir chronosequence. Microb. Ecol. 2008, 55, 500–511. [Google Scholar] [CrossRef]
- International Standard Organization. Soil Quality—Determination of Polycyclic Aromatic Hydrocarbons (PAH) by Gas Chromatography (GC) and High Performance Liquid Chromatography (HPLC); International Standard Organization: Geneva, Switzerland, 2014; Volume ISO 13859:2014. [Google Scholar]
- Frouz, J.; Cajthaml, T.; Kribek, B.; Schaeffer, P.; Bartuska, M.; Galertova, R.; Rojik, P.; Kristufek, V. Deep, subsurface microflora after excavation respiration and biomass and its potential role in degradation of fossil organic matter. Folia Microbiol. 2011, 56, 389–396. [Google Scholar] [CrossRef]
- Bailey, V.L.; Peacock, A.D.; Smith, J.L.; Bolton, H. Relationships between soil microbial biomass determined by chloroform fumigation-extraction, substrate-induced respiration, and phospholipid fatty acid analysis. Soil Biol. Biochem. 2002, 34, 1385–1389. [Google Scholar] [CrossRef]
- Kaur, A.; Chaudhary, A.; Choudhary, R.; Kaushik, R. Phospholipid fatty acid—A bioindicator of environment monitoring and assessment in soil ecosystem. Curr. Sci. 2005, 89, 1103–1112. [Google Scholar]
- Blanchet, G.; Gavazov, K.; Bragazza, L.; Sinaj, S. Responses of soil properties and crop yields to different inorganic and organic amendments in a Swiss conventional farming system. Agric. Ecosyst. Environ. 2016, 230, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Zhong, W.H.; Gu, T.; Wang, W.; Zhang, B.; Lin, X.G.; Huang, Q.R.; Shen, W.S. The effects of mineral fertilizer and organic manure on soil microbial community and diversity. Plant Soil 2010, 326, 511–522. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Dong, W.Y.; Dai, X.Q.; Schaeffer, S.; Yang, F.T.; Radosevich, M.; Xu, L.L.; Liu, X.Y.; Sun, X.M. Responses of absolute and specific soil enzyme activities to long term additions of organic and mineral fertilizer. Sci. Total Environ. 2015, 536, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Ai, C.; Liang, G.Q.; Sun, J.W.; Wang, X.B.; Zhou, W. Responses of extracellular enzyme activities and microbial community in both the rhizosphere and bulk soil to long-term fertilization practices in a fluvo-aquic soil. Geoderma 2012, 173, 330–338. [Google Scholar] [CrossRef]
- Bohme, L.; Bohme, F. Soil microbiological and biochemical properties affected by plant growth and different long-term fertilisation. Eur. J. Soil Biol. 2006, 42, 1–12. [Google Scholar] [CrossRef]
- Wang, Y.; Ji, H.F.; Gao, C.Q. Differential responses of soil bacterial taxa to long-term P, N, and organic manure application. J. Soils Sediments 2016, 16, 1046–1058. [Google Scholar] [CrossRef]
- Liu, E.K.; Yan, C.R.; Mei, X.R.; He, W.Q.; Bing, S.H.; Ding, L.P.; Liu, Q.; Liu, S.A.; Fan, T.L. Long-term effect of chemical fertilizer, straw, and manure on soil chemical and biological properties in northwest China. Geoderma 2010, 158, 173–180. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Wang, P.; Li, J.L.; Chen, Y.R.; Ying, X.Z.; Liu, S.Y. The effects of two organic manures on soil properties and crop yields on a temperate calcareous soil under a wheat-maize cropping system. Eur. J. Agron. 2009, 31, 36–42. [Google Scholar] [CrossRef]
- Dhull, S.; Goyal, S.; Kapoor, K.; Mundra, M. Microbial biomass carbon and microbial activities of soils receiving chemical fertilizers and organic amendments. Arch. Agron Soil Sci. 2004, 50, 641–647. [Google Scholar] [CrossRef]
- Saha, S.; Gopinath, K.A.; Mina, B.L.; Gupta, H.S. Influence of continuous application of inorganic nutrients to a Maize-Wheat rotation on soil enzyme activity and grain quality in a rainfed Indian soil. Eur. J. Soil Biol. 2008, 44, 521–531. [Google Scholar] [CrossRef]
- Saha, S.; Prakash, V.; Kundu, S.; Kumar, N.; Mina, B.L. Soil enzymatic activity as affected by long term application of farm yard manure and mineral fertilizer under a rainfed soybean-wheat system in N-W Himalaya. Eur. J. Soil Biol. 2008, 44, 309–315. [Google Scholar] [CrossRef]
- Liang, Q.; Chen, H.Q.; Gong, Y.S.; Yang, H.F.; Fan, M.S.; Kuzyakov, Y. Effects of 15 years of manure and mineral fertilizers on enzyme activities in particle-size fractions in a North China Plain soil. Eur. J. Soil Biol. 2014, 60, 112–119. [Google Scholar] [CrossRef]
- Li, J.; Cooper, J.M.; Lin, Z.A.; Li, Y.T.; Yang, X.D.; Zhao, B.Q. Soil microbial community structure and function are significantly affected by long-term organic and mineral fertilization regimes in the North China Plain. Appl. Soil Ecol. 2015, 96, 75–87. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, W.; Liang, G.Q.; Sun, J.W.; Wang, X.B.; He, P. Distribution of soil nutrients, extracellular enzyme activities and microbial communities across particle-size fractions in a long-term fertilizer experiment. Appl. Soil Ecol. 2015, 94, 59–71. [Google Scholar] [CrossRef]
- Banashree, S.; Smrita, B.; Nath, D.J.; Nirmali, G. Temporal responses of soil biological characteristics to organic inputs and mineral fertilizers under wheat cultivation in inceptisol. Arch. Agron. Soil Sci. 2017, 63, 35–47. [Google Scholar] [CrossRef]
- Zhao, J.; Ni, T.; Li, J.; Lu, Q.; Fang, Z.Y.; Huang, Q.W.; Zhang, R.F.; Li, R.; Shen, B.; Shen, Q.R. Effects of organic-inorganic compound fertilizer with reduced chemical fertilizer application on crop yields, soil biological activity and bacterial community structure in a rice-wheat cropping system. Appl. Soil Ecol. 2016, 99, 1–12. [Google Scholar] [CrossRef]



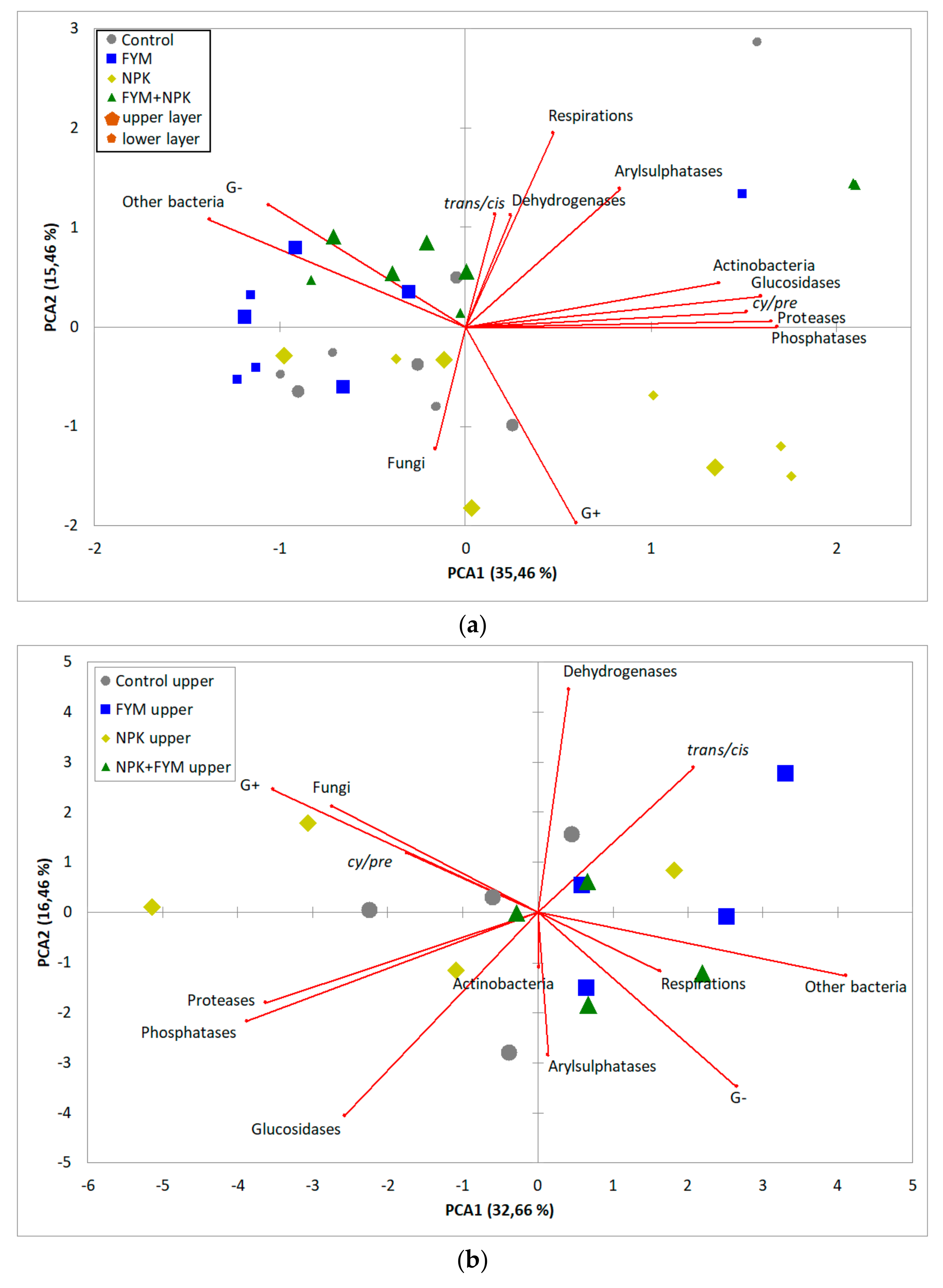{kind=link}
{kind=link}
| Plot | Ct (%) | Nt (%) | C/N | pH/KCl | pH/H2O | Available Nutrients (mg/kg) * | |||
|---|---|---|---|---|---|---|---|---|---|
| P | K | Ca | Mg | ||||||
| Control | 1.01 | 0.129 | 7.82 | 5.22 | 6.32 | 24.2 | 150 | 3297 | 187 |
| NPK | 1.21 | 0.145 | 8.34 | 5.12 | 6.12 | 71.7 | 177 | 2711 | 133 |
| FYM | 1.21 | 0.143 | 8.46 | 5.50 | 6.62 | 35.6 | 149 | 3483 | 189 |
| FYM + NPK | 1.33 | 0.166 | 7.89 | 5.22 | 6.28 | 76.2 | 207 | 3094 | 170 |
| Plot (Depth cm) | PLFAtot (mg.kg−1 dwt) | PLFAfun (mg.kg−1 dwt) | PLFAbac (mg.kg−1 dwt) | PLFAG+ (mg.kg−1 dwt) | PLFAG- (mg.kg−1 dwt) | PLFAAc (mg.kg−1 dwt) | cy/pre () | trans/cis () |
|---|---|---|---|---|---|---|---|---|
| Control U | 14.5 ± 1.0 | 0.51 ± 0.05 | 9.5 ± 0.7 | 3.7 ± 0.3 | 2.9 ± 0.2 | 1.35 ± 0.08 | 0.59 ± 0.05 | 0.00 ± 0.00 |
| Control L | 13.3 ± 2.0 | 0.44 ± 0.08 | 8.9 ± 1.3 | 3.2 ± 0.6 | 2.7 ± 0.4 | 1.44 ± 0.09 | 0.77 ± 0.37 | 0.00 ± 0.00 |
| NPK U | 14.1 ± 1.3 | 0.52 ± 0.05 | 9.0 ± 0.8 | 3.6 ± 0.4 | 2.7 ± 0.2 | 1.35 ± 0.11 | 1.06 ± 0.52 | 0.00 ± 0.00 |
| NPK L | 12.1 ± 1.2 | 0.40 ± 0.03 | 7.2 ± 0.3 | 2.8 ± 0.2 | 2.1 ± 0.1 | 1.63 ± 0.14 | 1.73 ± 0.15 | 0.00 ± 0.00 |
| FYM U | 16.3 ± 0.6 | 0.49 ± 0.06 | 10.7 ± 0.4 | 4.0 ± 0.1 | 3.3 ± 0.2 | 1.58 ± 0.04 | 0.64 ± 0.04 | 0.01 ± 0.03 |
| FYM L | 16.4 ± 1.7 | 0.64 ± 0.08 | 10.5 ± 1.1 | 3.9 ± 0.4 | 3.4 ± 0.4 | 1.64 ± 0.17 | 0.81 ± 0.50 | 0.06 ± 0.11 |
| FYM + NPK U | 14.9 ± 0.4 | 0.44 ± 0.06 | 9.8 ± 0.3 | 3.7 ± 0.1 | 3.0 ± 0.1 | 1.41 ± 0.03 | 0.70 ± 0.06 | 0.00 ± 0.00 |
| FYM + NPK L | 13.5 ± 2.0 | 0.47 ± 0.08 | 8.7 ± 1.3 | 3.2 ± 0.5 | 2.8 ± 0.5 | 1.61 ± 0.13 | 1.18 ± 0.63 | 0.00 ± 0.00 |
| Plot Depth (cm) | PLFAtot (mg.kg−1 dwt) | PLFAfun (mg.kg−1 dwt) | PLFAbac (mg.kg−1 dwt) | PLFAG+ (mg.kg−1 dwt) | PLFAG- (mg.kg−1 dwt) | PLFAAc (mg.kg−1 dwt) | cy/pre () | trans/cis () |
|---|---|---|---|---|---|---|---|---|
| Control | 13.9 ± 1.1 | 0.48 ± 0.05 | 9.2 ± 0.7 ab | 3.5 ± 0.3 | 2.8 ± 0.2 ab | 1.4 ± 0.1 | 0.68 ± 0.26 | 0.00 ± 0.00 |
| NPK | 13.1 ± 0.9 | 0.46 ± 0.04 | 8.1 ± 0.5 a | 3.2 ± 0.2 | 2.4 ± 0.2 a | 1.5 ± 0.1 | 1.39 ± 0.50 | 0.00 ± 0.00 |
| FYM | 16.3 ± 0.9 | 0.56 ± 0.06 | 10.6 ± 0.5 b | 3.9 ± 0.2 | 3.3 ± 0.2 b | 1.6 ± 0.1 | 0.73 ± 0.34 | 0.04 ± 0.08 |
| FYM+NPK | 14.2 ± 1.0 | 0.45 ± 0.05 | 9.3 ± 0.6 ab | 3.4 ± 0.2 | 2.9 ± 0.2 ab | 1.5 ± 0.1 | 0.94 ± 0.49 | 0.00 ± 0.00 |
| Plot Depth (cm) | Phosphatases µU.g−1 Dry Soil | Arylsulphatases µU.g−1 Dry Soil | Glucosidases µU.g−1 Dry Soil | Proteases µU.g−1 Dry Soil | Dehydrogenases µU.g−1 Dry Soil | Respiration mU.g−1 Dry Soil |
|---|---|---|---|---|---|---|
| Control U | 40.9 ± 1.8 | 2.19 abc ± 0.23 | 3.76 ± 0.14 | 11.3 ± 0.4 | 0.14 ± 0.09 | 1.79 a ± 0.23 |
| Control L | 34.2 ± 0.9 | 2.06 abc ± 0.23 | 3.47 ± 0.37 | 10.0 ± 1.1 | 0.21 ± 0.21 | 1.00 a ± 0.21 |
| NPK U | 41.4 ± 2.5 | 1.54 b ± 0.10 | 3.93 ± 0.16 | 9.9 ± 1.2 | 0.02 ± 0.02 | 2.19 a ± 0.48 |
| NPK L | 38.1 ± 3.3 | 1.4 bc ± 0.07 | 3.76 ± 0.13 | 9.1 ± 0.6 | 0.01 ± 0.01 | 1.55 a ± 0.25 |
| FYM U | 33.9 ± 1.4 | 2.46 a ± 0.10 | 3.93 ± 0.22 | 8.3 ± 0.5 | 0.17 ± 0.17 | 2.1 a ± 0.30 |
| FYM L | 33.6 ± 0.5 | 2.38 a ± 0.16 | 4.28 ± 0.09 | 9.4 ± 0.8 | 0.06 ± 0.06 | 2.22 a ± 0.10 |
| FYM+NPK U | 38.8 ± 2.8 | 1.85 abc ± 0.17 | 4.33 ± 0.18 | 10.1 ± 0.8 | 0.27 ± 0.16 | 6.79 b ± 0.25 |
| FYM + NPK L | 43.9 ± 3.9 | 2.20 ac ± 0.20 | 4.24 ± 0.17 | 11.8 ± 0.9 | 0.11 ± 0.09 | 6.8 b ± 0.13 |
| Plot Depth (cm) | Phosphatases µU.g−1 dwt | Arylsulphatases µU.g−1 dwt | Glucosidases µU.g−1 dwt | Proteases µU.g−1 dwt | Dehydrogenases µU.g−1 dwt | Respiration mU.g−1 dwt |
|---|---|---|---|---|---|---|
| Control | 37.6 ab ± 1.56 | 2.12 a ± 0.15 | 3.6 a ± 0.19 | 10.6 ± 0.6 | 0.17 ± 0.10 | 1.40 a ± 0.21 |
| NPK | 39.7 ab ± 2.02 | 1.48 b ± 0.06 | 3.84 ab ± 0.10 | 9.5 ± 0.7 | 0.06 ± 0.01 | 1.87 ab ± 0.28 |
| FYM | 33.7 a ± 0.69 | 2.42 a ± 0.09 | 4.10 ab ± 0.13 | 8.8 ± 0.5 | 0.12 ± 0.09 | 2.17 b ± 0.15 |
| FYM+NPK | 41.3 b ± 2.39 | 2.03 a ± 0.14 | 4.29 b ± 0.11 | 11.0 ± 0.6 | 0.19 ± 0.09 | 6.80 c ± 0.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holík, L.; Hlisnikovský, L.; Honzík, R.; Trögl, J.; Burdová, H.; Popelka, J. Soil Microbial Communities and Enzyme Activities after Long-Term Application of Inorganic and Organic Fertilizers at Different Depths of the Soil Profile. Sustainability 2019, 11, 3251. https://doi.org/10.3390/su11123251
Holík L, Hlisnikovský L, Honzík R, Trögl J, Burdová H, Popelka J. Soil Microbial Communities and Enzyme Activities after Long-Term Application of Inorganic and Organic Fertilizers at Different Depths of the Soil Profile. Sustainability. 2019; 11(12):3251. https://doi.org/10.3390/su11123251
Chicago/Turabian StyleHolík, Ladislav, Lukáš Hlisnikovský, Roman Honzík, Josef Trögl, Hana Burdová, and Jan Popelka. 2019. "Soil Microbial Communities and Enzyme Activities after Long-Term Application of Inorganic and Organic Fertilizers at Different Depths of the Soil Profile" Sustainability 11, no. 12: 3251. https://doi.org/10.3390/su11123251