Hydrolases Control Soil Carbon Sequestration in Alpine Grasslands in the Tibetan Plateau
by

Yuanye Zhang
1,2, 
Xia Wang
1,2,*,
Yuxin Sun
1,2,
Jinhong Wu
1,2,
Tao Deng
1,2,
Menghan Yuan
1,2,
Wenhui Duan
1,2 and
Yunfei Zhao
1,2 1
Gansu Key Laboratory for Environmental Pollution Prediction and Control, College of Earth and Environmental Sciences, Lanzhou University, Lanzhou 730000, China
2
Key Laboratory of Western China’s Environment System (Ministry of Education), College of Earth and Environmental Science, Lanzhou University, Lanzhou 730000, China
*
Author to whom correspondence should be addressed.
Sustainability 2024, 16(9), 3508; https://doi.org/10.3390/su16093508
Submission received: 11 March 2024
/
Revised: 18 April 2024
/
Accepted: 19 April 2024
/
Published: 23 April 2024
(This article belongs to the Special Issue Soil Carbon Sequestration and Greenhouse Gas Emission)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Microbial-sourced carbon is an important component of soil organic carbon (SOC) and influences SOC’s size and turnover. Soil extracellular enzymes can participate in the degradation of plants in the soil to produce substances needed by microorganisms, which in turn affects microbial sources of carbon. Most of the current studies focus on the effects of soil extracellular enzymes on SOC pools, while there is a lack of clarity regarding the effects on microbial sources of carbon during SOC pool formation. In this paper, three typical grassland types (alpine meadow, alpine grassland, and desert grassland, respectively) on the Tibetan Plateau were selected as research objects to investigate the effects of grassland type and soil depth on microbial-sourced carbon (amino sugars) and soil extracellular enzymes (hydrolytic enzymes: β-glucosidase and cellulase; oxidative enzymes: peroxidase and polyphenol oxidase) in the soil profiles. Our study shows that the content of amino sugars in the three grassland types followed the order: alpine meadow > alpine grassland > desert grassland; the content of hydrolytic enzyme followed the order of alpine meadow > alpine grassland > desert grassland; the content of oxidative enzyme followed the order of desert grassland > alpine grassland > alpine meadow; amino sugars content showed a positive correlation with hydrolytic enzymes and a negative correlation with oxidative enzymes; and the hydrolytic enzyme was the main factor promoting the accumulation of amino sugars. The environmental conditions of alpine meadows and alpine grasslands are more favorable for the formation of microbial-derived carbon and have greater sequestration potential, while desert grasslands are not favorable for the formation of microbial-derived carbon. The results of this study provide a reference basis for exploring the model of organic carbon sequestration in the Tibetan Plateau.
1. Introduction
Soil organic carbon (SOC) is the largest active carbon pool in terrestrial ecosystems, with a storage capacity of about 1500 Pg C, which is much larger than the sum of the atmospheric and vegetation carbon pools [1,2]. Secondly, SOC plays a central role in soil structure formation and retention, soil nutrient cycling, and soil biodiversity nourishment, and thus, research on SOC has been a core area in soil science [3].
As one of the largest terrestrial ecosystems, grassland has diverse ecosystem functions, and its organic carbon stock accounts for more than 25% of the global organic carbon, but its area accounts for only 26.91% of the total global land area, and it has huge potential for carbon sequestration [4]. A recent study suggested that the global grassland carbon pool is about 520 Pg C, with values of 50–120 Pg C and 279–592 Pg C for vegetation and soil carbon stocks, respectively [5]. However, the estimation results of grassland carbon sinks are still characterized by a high degree of certainty, and the mechanism of SOC sequestration in grassland ecosystems is still controversial [6,7].
With the increasing sophistication of biomarker identification techniques for molecular characterization, researchers have reached a consensus on the formation and stabilization of SOC with microbial-mediated soil carbon transformation and sequestration processes as the key mechanism for long-term organic carbon accumulation [8,9]. Soil extracellular enzymes are secreted into the soil by plant roots and soil microorganisms [10]. There are a variety of extracellular enzymes in the soil, and different extracellular enzymes have different functions, catalyze different reaction stages, and work together to maintain the overall function of the ecosystem [11]. The soil microbial carbon pump proposed by Chao Liang suggests that enzymes are capable of degrading plant-derived macromolecules into small fragments that can be directly absorbed and utilized by microorganisms for in vivo turnover, and that by assimilating ingested small molecular weight plant-derived carbon matrices, soil microorganisms synthesize their biomass and contribute microbial-sourced carbon to the soil through an iterative process of microbial cell growth, proliferation, and residue formation and accumulation [12,13]. Most of the current studies focus on the effects of soil extracellular enzymes on SOC pools, while there are few studies on the effects on microbial sources of carbon during SOC pool formation [14]. Therefore, it is necessary to carry out studies on soil extracellular enzymes in microbial sources of carbon.
The Tibetan Plateau is the largest plateau in China and the highest in the world, and is known as the “third pole” [15]. Because of its high altitude and low temperatures, the Tibetan Plateau stores a large amount of soil carbon, accounting for about 2.5 percent of the global soil carbon pool [16,17]. There are three main grassland types on the Tibetan Plateau: alpine meadow, alpine grassland, and desert grassland [18]. However, the effect of the same grassland types on soil carbon sequestration capacity still varies in different studies, and there is not a clear-cut result [19,20,21]. Therefore, clarifying the SOC formation and stability maintenance of alpine grassland on the Tibetan Plateau is of great significance to enhance the carbon conservation and sink capacity of the ecosystem.
In this study, three typical grassland types on the Tibetan Plateau were used to evaluate their sequestration effects on SOC. The objectives of our study were to investigate (1) the spatial distribution of extracellular enzymes and amino sugar content under different grassland types and soil depths; (2) the effects of different soil extracellular enzymes on microbial sources of carbon; and (3) the effects of basic soil physicochemical properties on the accumulation of soil extracellular enzymes and amino sugars and the mechanism of carbon sequestration in grassland soils.
2. Materials and Methods
2.1. Study Area
The Tibetan Plateau is located in southwestern China (between 26°00′ and 39°47′ N latitude and 73°19′ and 104°47′ E longitude) [22]. It extends from the south of the Himalayas South Court north to the Kunlun Mountains, Altun Shan, and Qilian Mountains, and at the northern edge, the average elevation is more than 4000 m. It is where East Asia, South Asia, and many other large rivers originate [18]. The average annual temperature on the Tibetan Plateau decreases from 20 °C in the southeast to below −6 °C in the northwest, while annual precipitation decreases from 2000 mm to below 50 mm [23]. Alpine grassland is the main vegetation type on the Tibetan Plateau, of which there are three grassland types, namely, alpine meadow (AM), alpine grassland (AG), and desert grassland (DG) [24].
2.2. Plant Surveys and Soil Sampling
The field survey was conducted from August to September 2021, and we chose the eastern alpine grassland of the Tibetan Plateau as the study area. The grassland in this region occupies a relatively large area, showing common characteristics in terms of the combination of temperature and moisture conditions, and is located at the intersection of warm humid and semi-arid zones, with typical grassland types of the Qinghai–Tibetan Plateau [25]. Soil and plant samples were collected from 21 alpine grassland sites (including 7 alpine meadows, 7 alpine grasslands, and 7 desert grasslands) (Figure 1). At each sample point, we set up a large area (30 m × 30 m) and then randomly arranged three 1 m × 1 m sub-areas, with the three sub-areas serving as three replicates of the sample point [26]. We recorded the number of plant species in each subzone and used the total number of plant species to represent species abundance at that sample site. All plants within each sampling square were mowed flush to harvest aboveground biomass. We collected soil from two depth bands (0–10 cm, 20–30 cm) in each subzone. Three replicate soils from the same layer were mixed to form a composite soil. Fresh soil samples were then stored at 4 °C in a closed cooler covered with ice packs and returned to the laboratory promptly after each sampling.
All soil samples were passed through a 2 mm sieve to remove visible stones as well as roots. The sieved soil was divided into two parts: one part was stored at 4 °C for soil physicochemical properties and amino sugars, and the other part was stored at −20 °C for soil extracellular enzyme activity analysis.
2.3. Soil Properties
We measured soil physicochemical properties including soil pH, total nitrogen (TN), total phosphorus (TP), cation exchange capacity (CEC), soil moisture content, soil texture, and electrical conductivity.
Soil pH was determined using the potentiometric method [27]. Soil total nitrogen and total phosphorus were determined using a fully automated chemical analyzer (Smart Chem 200, Westco Scientific Instruments, Brookfield, CT, USA) [28]. Soil cation exchange was determined using the hexaamminecobalt(III) chloride leaching-spectrophotometric method [29]. Soil moisture content was determined using the drying method [30]. Soil texture was determined using a laser particle size meter (Master Sizer 2000, Malvern Instruments, Malvern, UK) [31]. Soil conductivity was determined using the electrode method [32].
2.4. Determination of Soil Extracellular Enzymes
We measured the activities of soil hydrolases and oxidative enzymes. Hydrolytic enzymes included β-glucosidase (BG) and cellulase (CB), and oxidative enzymes included polyphenol oxidase (POX) and peroxidase (PER). The high-throughput fluorescence method was used for the determination of soil enzyme activities [33].
Substrate solution, standard control, and soil homogenate were prepared by adjusting the pH to 6.0 using hydrochloric acid at a concentration of 25 mM sodium maleate as buffer. The soil stored in the refrigerator at −20 °C was weighed in two portions of 0.2 g into two centrifuge tubes, both of which were added with 20 mL of buffer for cell fragmentation, and one of the solutions was poured into a round plastic box and then added with 50 mL of buffer for the determination of hydrolytic enzyme activity; the other portion was added with 10 mL of buffer, and then filtered through 0.45 μm filter paper to obtain the supernatant for the determination of oxidase activity. In each microtiter plate, 125 μL of substrate solution was added to 125 μL of soil homogenate. For each enzyme, enzyme activity was determined at a certain substrate concentration. The fluorescence signals of the hydrolases were read after 4 h of incubation at room temperature using an enzyme marker (BioTek Synergy H1 microplate reader, Winooski, VT, USA) under excitation at 360 nm and emission at 460 nm. Microtiter plate data of oxidase absorption at 410 nm were read after 24 h of oxidase incubation. Soil extracellular enzyme activity was expressed as mg g−1.
2.5. Amino Sugar Analysis
Soil amino sugars were determined using the acid hydrolysis method [34]. The main components were glucosamine, galactose, cytosolic acid, and mannose.
Specifically, 1 g of air-dried soil was placed into a hydrolysis flask and hydrolyzed with 10 mL of 6 M HCl at 105 °C for 8 h, after which we added 100 μL of inositol to the cooled sample. The solution was then filtered into a chicken heart flask and evaporated with a rotary evaporator at 52 °C. The residue was then dissolved in 5 mL of deionized water and adjusted to a pH of between 6.6 and 6.8 using 1 M potassium hydroxide. The precipitates were removed by centrifugation at 3000 rpm for 10 min. After freeze-drying, the supernatant was dissolved in anhydrous methanol and centrifuged again. We then transferred the supernatant to the derivatization flask and blow-dried the samples with nitrogen at 45 °C. Next, we added 1 mL of deionized water and 100 μL N-methyl-glucamine for freeze-drying again. After that, we added 300 μL of derivatization reagent (preparation method: 32 mg/mL hydroxylamine hydrochloride and 40 mg/mL 4-dimethylamino) pyridine solution, dissolved in the mixed solution of pyridine: methanol (4:1 volume ratio). Then, the mixture was covered and sealed, violently vibrated, and heated at 75–80 °C for 30 min. After cooling to room temperature, we added 1 mL of acetic anhydride and heated it at 75–80 °C for 20 min, then added 1.5 mL of dichloromethane. Excess derivatization reagents were extracted with 1 M of HCl and deionized water, after which the aqueous phase was discarded. After drying with nitrogen at 45 °C, the organic solution was dissolved in the mixed solvent of ethyl acetate and n-hexane (1:1 volume ratio).
We used a Trace 1300 gas chromatographer with a TG-1 MS (30 m × 0.25 mm × 0.25 μm) column and an FID detector (Thermo Fisher Scientific, Waltham, MA, USA) to quantify amino sugars. The oven temperature was held at 120 °C for 4 min, then raised to 230 °C at the rate of 10 °C min−1, then increased to 250 °C at a rate of 5 °C min−1 for 4 min, and finally increased to 290 °C at a rate of 40 °C min−1 for 5 min.
2.6. Parameters Related to Soil Microbial Source Carbon
Of the four amino sugars, it is not clear which microorganisms Mann and Gala originate from. Whereas fungi produce most of the Gluc, bacteria produce a small portion of Gluc in addition to Mura. We used Gluc to calculate carbon from fungal sources and Mura to calculate carbon from bacterial sources. The specific were as follows in (1) and (2):
In Equation (1), it is assumed that the ratio of Mura to Gluc in bacterial cells is 1:2; 9 is the conversion factor from Gluc to fungal-derived carbon; 179.17 is the relative molecular mass of Gluc; and 251.23 is the relative molecular mass of Mura. In Equation (2), 45 is the conversion factor from Mura to carbon of bacterial origin. In Equations (1) and (2), the units of Mura and Gluc are mg g−1.
Soil microbial-sourced carbon is the sum of soil fungal-sourced carbon and bacterial-sourced carbon, with the following formula:
In Equation (3), Microbial C: carbon of microbial origin; Fungal-derived C: carbon of fungal origin; and Bacterial-derived C: carbon of bacterial origin, all in mg g−1.
2.7. Statistical Analysis
First, differences were tested for amino sugars and oxidases, β-glucosidase, and cellulase. A one-way ANOVA test was used to evaluate the differences between the three grassland types at different soil depths, and a paired t-test was used to compare the differences between the topsoil and subsoil of each grassland type. For all tests, p < 0.05 was significant. Next, we explored the relationship between amino sugars and oxidase, β-glucosidase, and cellulase using linear regression. Then, we determined which environmental parameters affected the accumulation of amino sugars with oxidases, β-glucosidases, and cellulase. We used correlation heat maps and principal component analysis (PCA) to assess the relationships between amino sugars, oxidases, β-glucosidases, and cellulase with environmental parameters.
3. Results
3.1. Dynamics of Amino Sugar in Soil
As shown in Figure 2, we know that soil amino sugar content tends to decrease with the deepening of soil depth. The amino sugar content of alpine meadows is higher than that of alpine grassland and desert grassland. In the topsoil, both alpine meadow and alpine grassland had significantly higher content than desert grassland (p < 0.05); in the subsoil, alpine meadow was significantly higher than desert grassland (p < 0.05), and alpine grassland was not significantly different from the remaining two grassland types. Between different soil depths in the same grassland type, topsoil was significantly higher than subsoil in both alpine meadows and alpine grasslands (p < 0.05), and there was no significant difference in desert grasslands. The mean values of amino sugar content of the alpine meadow, alpine grassland, and desert grassland soils were 5.13, 4.27, and 1.2 mg g−1, respectively.
3.2. Soil Extracellular Enzyme
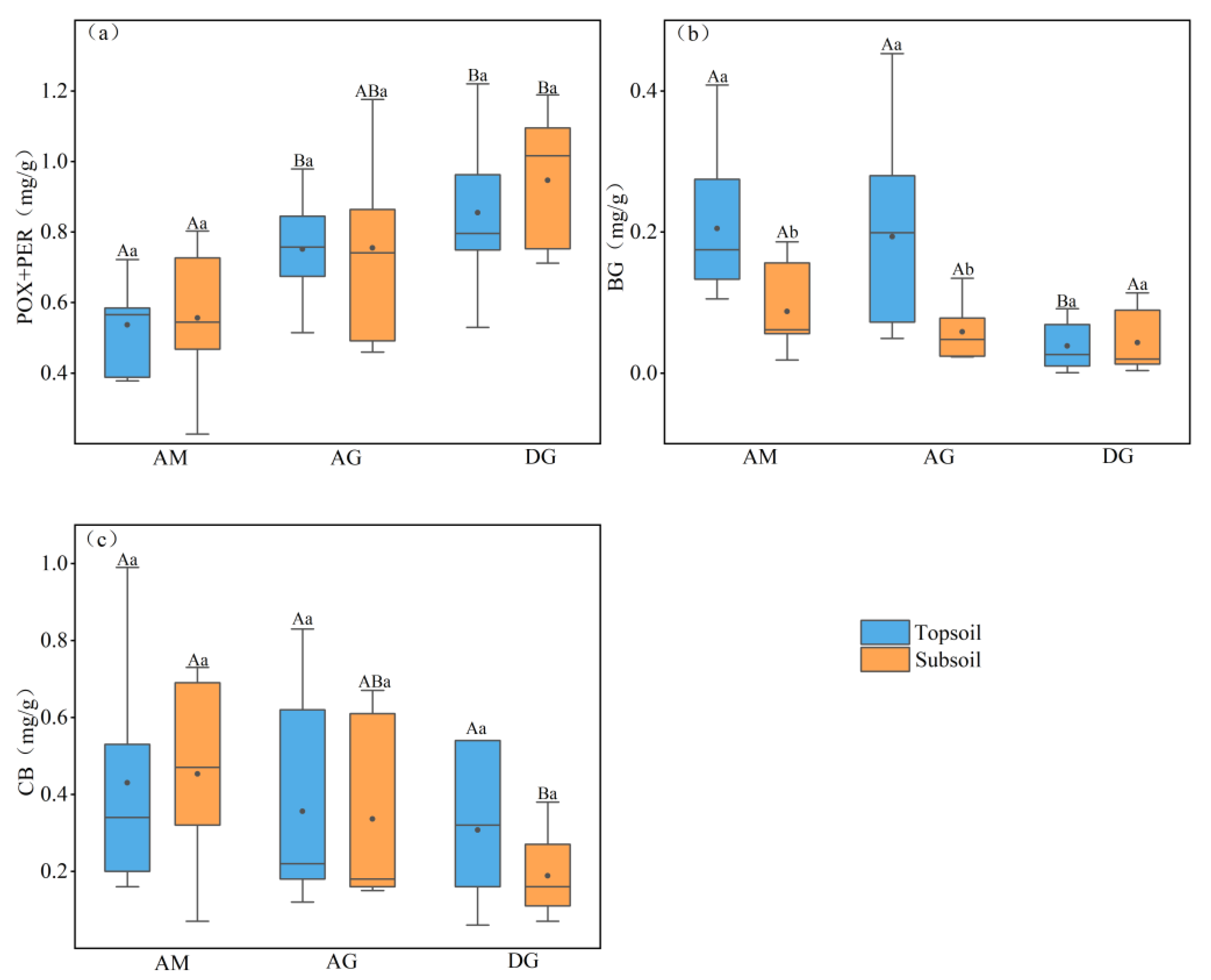
As shown in Figure 3, we know that all soil extracellular enzyme contents responded to depth as well as to changes in grassland type. Oxidase (POX+PER) content tended to increase with soil depth, and the average oxidase content of desert grassland was higher than that of alpine meadow and alpine steppe. In the topsoil, both the desert grassland and alpine grassland oxidase contents were significantly higher than that of alpine meadow (p < 0.05); in the subsoil, desert grassland oxidase contents were significantly higher than that of alpine meadow (p < 0.05). There was no significant relationship between different soil depths in the same grassland type for any of the three grassland types in terms of oxidase content. The mean values of oxidase content in alpine meadow, alpine grassland, and desert grassland soils were 0.55, 0.75, and 0.90 mg g−1, respectively.
The β-glucosidase (BG) content tended to decrease with the soil depth, and the average content of alpine meadow BG was higher than that of alpine grassland and desert grassland. In the topsoil, the BG content of both alpine meadow and alpine steppe was significantly higher than that of desert steppe (p < 0.05). In the subsoil, there were no significant differences between the three grassland types. It can be observed that, between different soil depths of the same grassland type, the topsoil BG content of alpine meadows and alpine grasslands was significantly higher than that of the subsoil (p < 0.05); there was no significant difference in desert grasslands. The mean values of soil BG content in alpine meadow, alpine grassland, and desert grassland were 0.15, 0.13, and 0.04 mg g−1, respectively.
The cellulase (CB) content tended to increase with soil depth in alpine meadows and decrease with soil depth in alpine grasslands and desert steppes, and the average CB content of the topsoil and subsoil in alpine meadows was higher than that in alpine grasslands and desert steppes. In the topsoil, there was no significant difference in CB content among the three grassland types; in the subsoil, the CB content of alpine meadow was significantly higher than that of desert grassland (p < 0.05), and there was no significant difference between alpine grassland and the remaining two grassland types. Between different soil depths of the same grassland type, there was no significant difference in CB content among the three grassland types. The mean values of soil CB content in alpine grassland and desert grassland were 0.44, 0.34, and 0.25 mg g−1, respectively.
In order to investigate the effect of soil enzyme activities on microbial-derived carbon, we selected oxidizing enzymes (polyphenol oxidase+peroxidase) and hydrolytic enzymes (BG + CB) for the study, respectively. As shown in Figure 4, we know that microbial-derived carbon showed a significant negative correlation with oxidizing enzymes (p < 0.01, Figure 4a), a significant positive correlation with BG (p < 0.01, Figure 4b), and a positive correlation with CB (Figure 4c).
3.3. Influence of Soil Physical and Chemical Properties on Extracellular Enzyme Activities of Amino Sugars and Soil Enzymes
Correlation analysis showed that in the soil surface layer, the amino sugar content was significantly positively correlated with TN, clay grain, CEC, and moisture content, and significantly negatively correlated with pH (p < 0.05, Figure 5); the oxidase content was significantly positively correlated with pH and significantly negatively correlated with TN and moisture content (p < 0.05, Figure 5); the BG content was significantly positively correlated with TN and CEC (p < 0.05, Figure 5); and the CB content was significantly positively correlated with conductivity (p < 0.05, Figure 5).
In the subsoil, the amino sugar content showed a significant positive correlation with TN, CEC, and moisture content (p < 0.05, Figure 5) and a significant negative correlation with pH (p < 0.05, Figure 5); the oxidase content showed a significant positive correlation with pH and sand (p < 0.05, Figure 5) and a significant negative correlation with TN, conductivity, CEC, and moisture content (p < 0.05, Figure 5); the BG content was significantly positively correlated with TN (p < 0.05, Figure 5) and negatively correlated with pH (p < 0.05, Figure 5); and the CB content was not significantly related to the physicochemical properties.
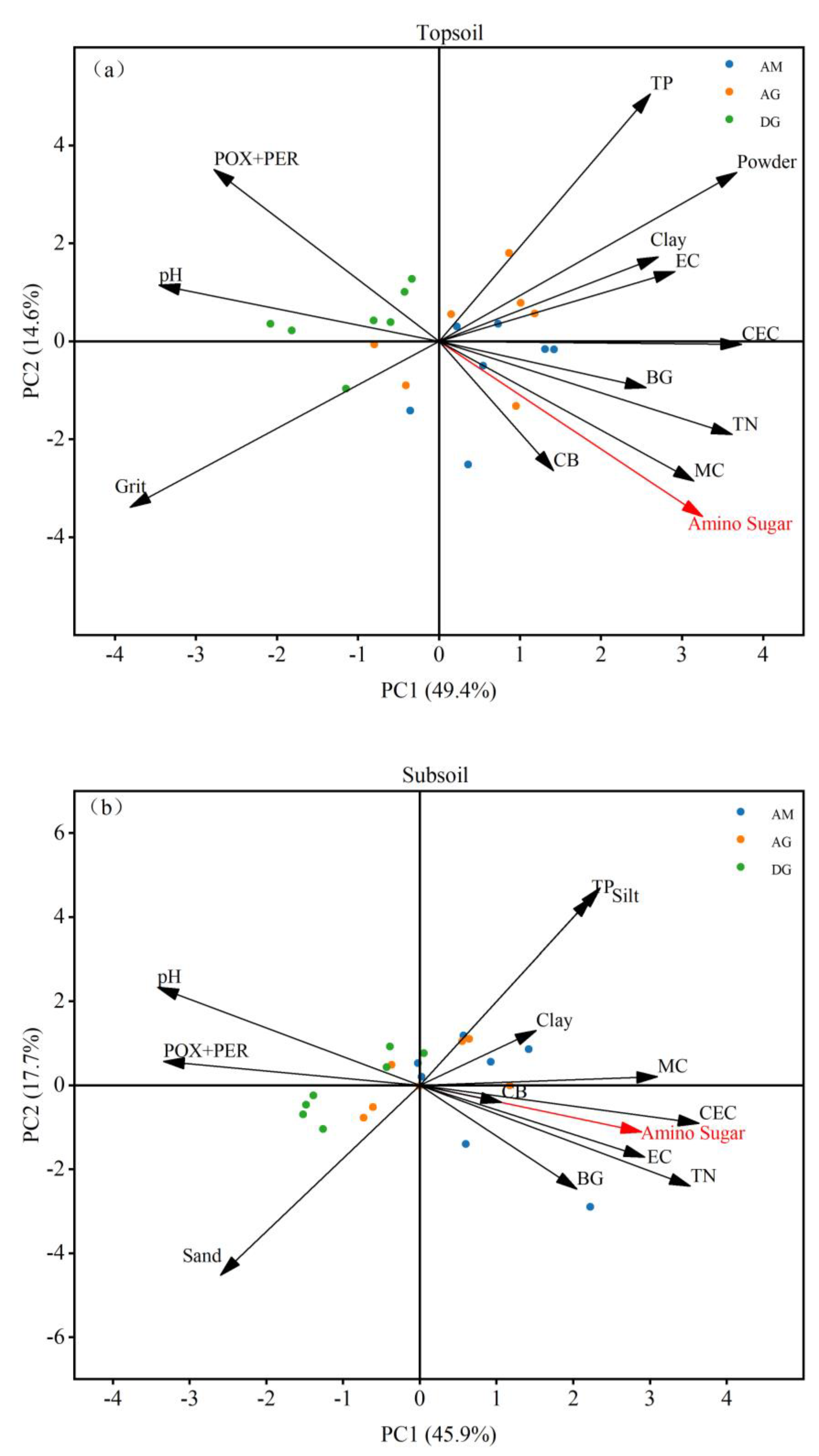
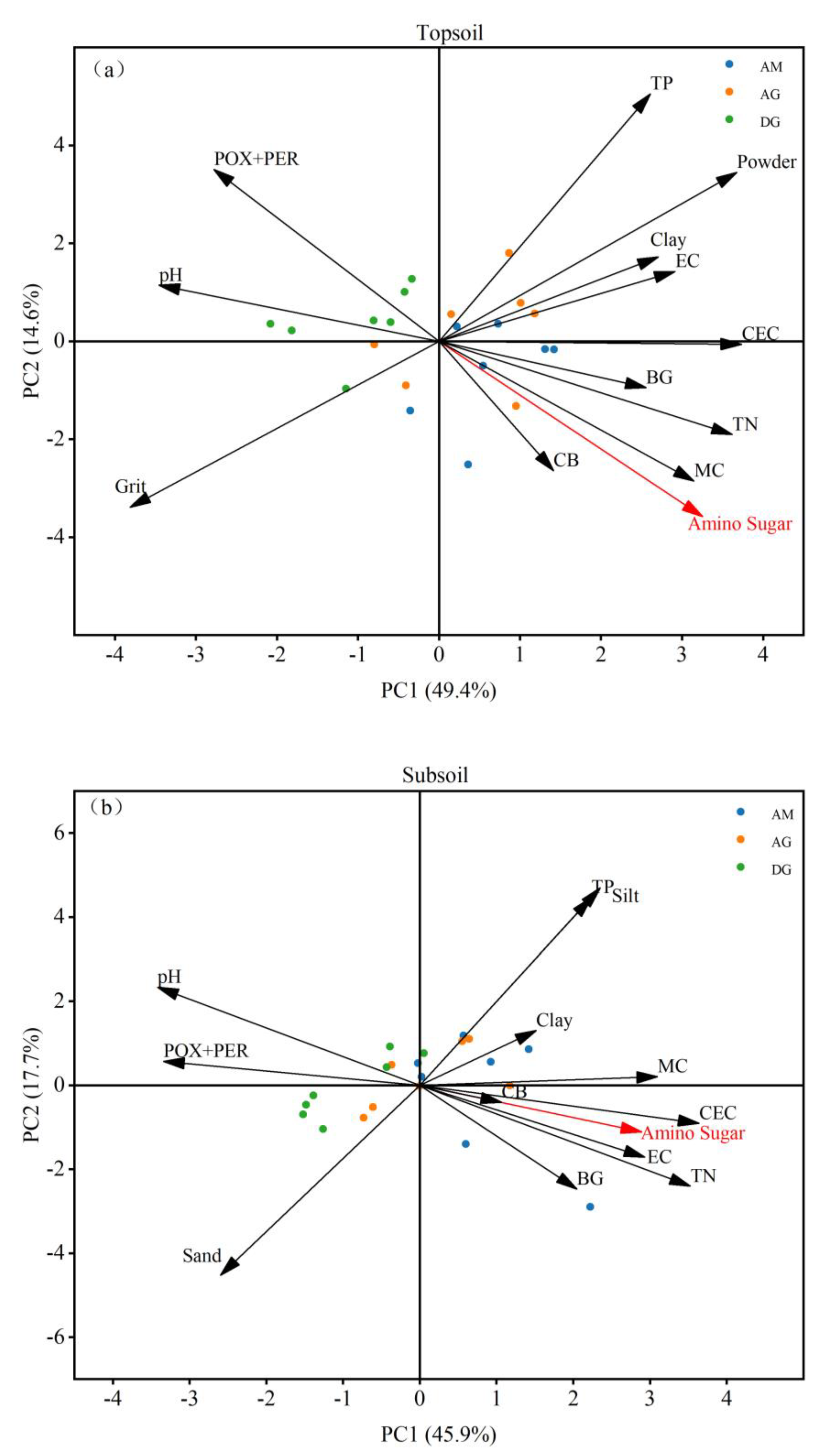
PCA analysis showed that in the soil surface layer, amino sugars, BG, CB, and POX+PER were mainly affected by TN, CEC, MC, and pH, and amino sugars, BG, and CB were significantly positively correlated with TN, CEC, and MC and negatively correlated with pH, while the correlation was opposite for POX+PER (Figure 6a). In the subsoil, amino sugars, BG, CB, and POX+PER were mainly affected by TN, CEC, and pH. Amino sugars, BG, and CB showed a significant positive correlation with TN and CEC and a negative correlation with pH, while the opposite correlation was observed for POX+PER (Figure 6b). As shown in Figure 6, the results of PCA analysis were the same as those of correlation analyses, which strengthened the illustration of the effect of soil physicochemical properties on amino sugars and extracellular enzyme activities.
4. Discussion
4.1. Influence of Different Grassland Types and Soil Depth on Soil Extracellular Enzymes and Amino Sugar Content
In all three grassland types, the soil amino sugar content was greater in the surface layer than in the subsoil layer (Figure 2), which is consistent with the global trend of decreasing soil amino sugar content in ecosystems with increasing soil depth [9,35]. This is mainly due to the fact that root biomass and apoplastic material are mainly distributed in the topsoil and carbon input decreases with deeper soil depth [36]. The higher amount of nutrients in the topsoil stimulates microbial growth and reproduction, resulting in a faster turnover of microbial biomass, which leads to more amino sugars in the topsoil than in the subsoil [37].
The soil amino sugar content was as follows: alpine meadows > alpine grassland > desert grassland (Figure 2), which may be due to the different levels of complexity of the ecosystem structure and function of the three grassland types [38]. Compared to desert grassland, alpine meadow and alpine grassland have higher species abundance and plant productivity, which can increase the availability of soil-dissolved organic carbon and nutrients and stimulate microbial activity, leading to an increase in microbial biomass and turnover of living microorganisms, which can lead to an increase in the accumulation of soil amino sugars [38,39].
Soil hydrolase activity was higher in alpine meadows and alpine grasslands than in desert grasslands, probably due to the higher content of soil amino sugars [14]. This finding is consistent with previous studies which found that the soil hydrolase activity increases with increasing soil organic matter content [40,41]. Similarly, all types of hydrolytic enzyme activities decreased with increasing soil depth, mainly due to the decrease in soil carbon effectiveness [42]. In contrast, oxidase activity was highest in desert grasslands, followed by alpine grasslands and alpine meadows. Previous studies have reported that microorganisms can produce enzymes only in nutrient-deficient conditions to avoid the production of relevant enzymes in nutrient-rich conditions, thus reducing manufacturing costs [43,44]. As a result, alpine meadow and alpine steppe soils are heavily invested with apomictic material, which produces higher levels of unstable C, resulting in lower oxidase activity compared to desert steppe soils [11]. The oxidase content was higher in the subsoil than topsoil, which is mainly due to the significant negative correlation between oxidase and TN (Figure 5): as depth increases, soil TN content decreases, changing the substrate effectiveness of the microbial community and soil properties, which in turn increases soil oxidase content [45].
4.2. Effect of Different Soil Extracellular Enzymes on Amino Sugars
The results of the study showed a highly significant positive correlation between BG (β-glucosidase) and amino sugars (R2 = 0.36, p < 0.01, Figure 4b), a highly significant negative correlation between oxidative enzymes and amino sugars (R2 = 0.25, p < 0.01, Figure 4a), and a positive correlation between CB (cellulase) and amino sugars. These results suggest that the accumulation of soil carbon from microbial sources in alpine grasslands of the Tibetan Plateau mainly depends on BG and oxidative enzymes.
Cellulose is preferentially degraded during the early stages of plant apoplastic decomposition [11]. In ecological studies, the most common cellulose hydrolases are CB and BG [11]. CB hydrolyzes fibrous dimers from the non-reducing ends of the cellulose molecule; BG hydrolyzes oligosaccharides to monosaccharides, providing usable substrates and energy for microorganisms, thus contributing microbial-sourced carbon to the soil through an iterative microbial process [13,46].
4.3. Influence of Basic Physical and Chemical Properties of Soil on the Accumulation of Extracellular Enzymes and Amino Sugar Content in Soil
The results of the study showed that TN, CEC, and pH were the main factors that mainly affected the content of amino sugars, BG, and oxidative enzymes. N is closely related to microbial growth and development, and only in nutrient-rich environments can microbes be more involved in biosynthesis than in catabolism [47]. Microbial communities are very large and active in the soil, and they require large amounts of nutrients to maintain normal life activities [48]. The soil cation exchange, which is a measure of the soil’s ability to retain fertilizer, has a positive effect on vegetation productivity, which reinforces the importance of soil nutrients in promoting the accumulation of amino sugars and BG [49,50]. Soil amino sugars and BG decreased with increasing pH in this study, which may be attributed to the fact that higher pH reduces SOC storage and limits nutrient availability, leading to reduced carbon inputs and resulting in a decrease in amino sugars and BG content [51].
On the contrary, for soil oxidase, the more soil nutrients there are, when the TN and CEC are higher, the more unstable C inputs in the soil, which will inhibit the activity of oxidase [52]. Some studies have shown that some enzymatic reactions are very sensitive to changes in pH and can only be carried out in a narrow range of pH; when the pH value is lower than 5.0, the activity of oxidase will be almost completely lost, so a high pH is more conducive to the improvement of oxidase activity and thus to the accumulation of oxidase content [53]. Therefore, a high pH value is more favorable to increase the activity of oxidase and thus to accumulate its content. Factors affecting soil oxidase content formed an inverse relationship with those affecting amino sugars and BG content, reinforcing the importance of the influence of both on amino sugar content accumulation.
4.4. Analysis of Soil Carbon Sequestration Mechanism in Alpine Grassland
Our results indicate that hydrolytic enzymes promote the accumulation of microbial-sourced carbon and that oxidative enzymes act as inhibitors of microbial-sourced carbon accumulation. In terms of soil properties, soil TN and soil CEC are the most important variables controlling the distribution of hydrolytic enzymes and microbial source carbon composition, which can increase the accumulation of microbial source carbon by increasing the activity of hydrolytic enzymes and accelerating the microbial iterative process, illustrating that soil nutrients are crucial for promoting microbial source carbon. Lower pH can favor the microbial source carbon by affecting the carbon inputs, microbial community structure, and functioning increase in hydrolase activity. These results suggest that hydrolytic enzymes play a dominant role in the accumulation of microbial-sourced carbon in alpine grasslands of the Tibetan Plateau, and that soil TN, CEC, and pH are the most important soil physicochemical properties affecting the accumulation process.
By comparing the three grassland types, we found that the ecological environments of alpine meadows and alpine grasslands are more favorable for the accumulation of microbial-sourced carbon, showing great potential for carbon sequestration. Although microbial sources of carbon exist in desert grasslands, their ecological environment is unfavorable for the accumulation of microbial sources of carbon. In the Tibetan Plateau, the degree of desertification is gradually increasing due to grazing, climate change, and anthropogenic activities, which is unfavorable for soil carbon sequestration [54]. Our results demonstrate the importance of soil nutrients and pH for carbon accumulation from microbial sources, and perhaps the application of fertilizers or changes in soil pH in desert grasslands could improve carbon loss in desert grasslands.
The grasslands of the Tibetan Plateau are the largest alpine biota in the world, and the flora and fauna of the Tibetan Plateau are historically linked to and similar to the Arctic, North America, Eurasia, and even mid-latitude regions of the entire Northern Hemisphere due to the north–south migration corridor of the Northern Hemisphere as well as the natural and climatic continuity of North America and Eurasia [55]. Therefore, our results not only reveal the mechanism of soil carbon sequestration in the alpine grasslands of the eastern Tibetan Plateau, but also guide carbon sequestration studies in other regions of the world.
5. Conclusions
Our results show that hydrolases can promote the accumulation of soil microbial-derived carbon. Soil nutrients and pH are the most important soil physicochemical properties affecting the accumulation process. Furthermore, alpine meadows and grasslands exhibit greater potential for carbon sequestration. Our results demonstrate the important role of hydrolases in carbon sequestration and reveal the effects of soil physical and chemical properties on carbon sequestration, which provides theoretical guidance for addressing climate change and regulating soil carbon sequestration. In future research on carbon sequestration in grasslands, the role of soil nutrients and pH should be comprehensively considered, starting from hydrolases.
Author Contributions
Conceptualization, Y.Z. (Yuanye Zhang) and X.W.; methodology, Y.Z. (Yuanye Zhang); software, J.W.; validation, Y.Z. (Yuanye Zhang), X.W. and Y.S.; formal analysis, Y.Z. (Yuanye Zhang); investigation, Y.Z. (Yuanye Zhang), X.W., J.W. and T.D.; resources, X.W.; data curation, M.Y., W.D. and Y.Z. (Yunfei Zhao); writing—original draft preparation, Y.Z. (Yuanye Zhang); writing—review and editing, X.W.; visualization, Y.Z. (Yuanye Zhang); supervision, X.W.; project administration, X.W.; funding acquisition, X.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (NSFC 42271079; 42301069), the Key Research and Development Program of Gansu Province (22YF7FA020) and the Fundamental Research Funds for the Central Universities (lzujbky-2023-pd01).
Institutional Review Board Statement
Not applicable since our study did not involve humans or animals.
Informed Consent Statement
Not applicable since our research did not involve humans.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We appreciate the financial support for this research provided by the National Natural Science Foundation of China (NSFC 42271079; 42301069), the Key Research and Development Program of Gansu Province (22YF7FA020) and the Fundamental Research Funds for the Central Universities (lzujbky-2023-pd01).
Conflicts of Interest
The authors declare they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Georgiou, K.; Jackson, R.B.; Vinduskova, O.; Abramoff, R.Z.; Ahlstrom, A.; Feng, W.; Harden, J.W.; Pellegrini, A.F.A.; Polley, H.W.; Soong, J.L.; et al. Global stocks and capacity of mineral-associated soil organic carbon. Nat. Commun. 2022, 13, 3797. [Google Scholar] [CrossRef] [PubMed]
- Friedlingstein, P.; O’Sullivan, M.; Jones, M.W.; Andrew, R.M.; Hauck, J.; Olsen, A.; Peters, G.P.; Peters, W.; Pongratz, J.; Sitch, S.; et al. Global Carbon Budget 2020. Earth Syst. Sci. Data 2020, 12, 3269–3340. [Google Scholar] [CrossRef]
- Gerke, J. The Central Role of Soil Organic Matter in Soil Fertility and Carbon Storage. Soil Syst. 2022, 6, 33. [Google Scholar] [CrossRef]
- Bai, Y.; Cotrufo, M.F. Grassland soil carbon sequestration: Current understanding, challenges, and solutions. Science 2022, 377, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Bullock, J.M.; Lavorel, S.; Manning, P.; Schaffner, U.; Ostle, N.; Chomel, M.; Durigan, G.; Fry, E.L.; Johnson, D.; et al. Combatting global grassland degradation. Nat. Rev. Earth Environ. 2021, 2, 720–735. [Google Scholar] [CrossRef]
- Yang, Y.; Dou, Y.; Wang, B.; Wang, Y.; Liang, C.; An, S.; Soromotin, A.; Kuzyakov, Y. Increasing contribution of microbial residues to soil organic carbon in grassland restoration chronosequence. Soil Biol. Biochem. 2022, 170, 108688. [Google Scholar] [CrossRef]
- Abdalla, M.; Hastings, A.; Chadwick, D.R.; Jones, D.L.; Evans, C.D.; Jones, M.B.; Rees, R.M.; Smith, P. Critical review of the impacts of grazing intensity on soil organic carbon storage and other soil quality indicators in extensively managed grasslands. Agric. Ecosyst. Environ. 2018, 253, 62–81. [Google Scholar] [CrossRef] [PubMed]
- Kallenbach, C.M.; Frey, S.D.; Grandy, A.S. Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat. Commun. 2016, 7, 13630. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; An, S.; Liang, C.; Liu, Y.; Kuzyakov, Y. Microbial necromass as the source of soil organic carbon in global ecosystems. Soil Biol. Biochem. 2021, 162, 108422. [Google Scholar] [CrossRef]
- Xiao, W.; Chen, X.; Jing, X.; Zhu, B. A meta-analysis of soil extracellular enzyme activities in response to global change. Soil Biol. Biochem. 2018, 123, 21–32. [Google Scholar] [CrossRef]
- Jian, S.; Li, J.; Chen, J.; Wang, G.; Mayes, M.A.; Dzantor, K.E.; Hui, D.; Luo, Y. Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: A meta-analysis. Soil Biol. Biochem. 2016, 101, 32–43. [Google Scholar] [CrossRef]
- Liang, C. Soil microbial carbon pump: Mechanism and appraisal. Soil Ecol. Lett. 2020, 2, 241–254. [Google Scholar] [CrossRef]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2017, 2, 17105. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Luo, Y.; van Groenigen, K.J.; Hungate, B.A.; Cao, J.; Zhou, X.; Wang, R.-w. A keystone microbial enzyme for nitrogen control of soil carbon storage. Sci. Adv. 2018, 4, eaaq1689. [Google Scholar] [CrossRef] [PubMed]
- Yao, T.; Xue, Y.; Chen, D.; Chen, F.; Thompson, L.; Cui, P.; Koike, T.; Lau, W.K.M.; Lettenmaier, D.; Mosbrugger, V.; et al. Recent Third Pole’s Rapid Warming Accompanies Cryospheric Melt and Water Cycle Intensification and Interactions between Monsoon and Environment: Multidisciplinary Approach with Observations, Modeling, and Analysis. Bull. Amer. Meteorol. Soc. 2019, 100, 423–444. [Google Scholar] [CrossRef]
- Chen, H.; Ju, P.; Zhu, Q.; Xu, X.; Wu, N.; Gao, Y.; Feng, X.; Tian, J.; Niu, S.; Zhang, Y.; et al. Carbon and nitrogen cycling on the Qinghai-Tibetan Plateau. Nat. Rev. Earth Environ. 2022, 3, 701–716. [Google Scholar] [CrossRef]
- Liu, S.; Zamanian, K.; Schleuss, P.-M.; Zarebanadkouki, M.; Kuzyakov, Y. Degradation of Tibetan grasslands: Consequences for carbon and nutrient cycles. Agric. Ecosyst. Environ. 2018, 252, 93–104. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, W.; Xue, K.; Wang, S.; Zhang, L.; Hu, R.; Zeng, H.; Xu, X.; Li, Y.; Jiang, L.; et al. Grassland changes and adaptive management on the Qinghai-Tibetan Plateau. Nat. Rev. Earth Environ. 2022, 3, 668–683. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, Z.; Zhang, J.; Zhao, M.; He, N. Changes in Soil Particulate Organic Carbon and Their Response to Changing Environments on the Tibetan Plateau, Mongolian Plateau, and Loess Plateau, China. J. Soil Sci. Plant Nutr. 2023, 23, 420–430. [Google Scholar] [CrossRef]
- Wei, X.; Wang, X.; Ma, T.; Huang, L.; Pu, Q.; Hao, M.; Zhang, X. Distribution and mineralization of organic carbon and nitrogen in forest soils of the southern Tibetan Plateau. CATENA 2017, 156, 298–304. [Google Scholar] [CrossRef]
- Yang, R.-M.; Zhang, G.-L.; Yang, F.; Zhi, J.-J.; Yang, F.; Liu, F.; Zhao, Y.-G.; Li, D.-C. Precise estimation of soil organic carbon stocks in the northeast Tibetan Plateau. Sci. Rep. 2016, 6, 21842. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Wang, S.; Jiang, N.; Sun, J.; Cao, R.; Ling, X.; Fang, B.; Zhang, L.; Zhang, L.; Xu, X.; et al. Plant phenology changes and drivers on the Qinghai-Tibetan Plateau. Nat. Rev. Earth Environ. 2022, 3, 633–651. [Google Scholar] [CrossRef]
- Xia, M.; Jia, K.; Zhao, W.; Liu, S.; Wei, X.; Wang, B. Spatio-temporal changes of ecological vulnerability across the Qinghai-Tibetan Plateau. Ecol. Indic. 2021, 123, 107274. [Google Scholar] [CrossRef]
- Tang, L.; Dong, S.; Sherman, R.; Liu, S.; Liu, Q.; Wang, X.; Su, X.; Zhang, Y.; Li, Y.; Wu, Y.; et al. Changes in vegetation composition and plant diversity with rangeland degradation in the alpine region of Qinghai-Tibet Plateau. Rangel. J. 2015, 37, 107–115. [Google Scholar] [CrossRef]
- Tian, P.; Lu, H.; Feng, W.; Guan, Y.; Xue, Y. Large decrease in streamflow and sediment load of Qinghai-Tibetan Plateau driven by future climate change: A case study in Lhasa River Basin. CATENA 2020, 187, 104340. [Google Scholar] [CrossRef]
- Ding, J.; Chen, L.; Zhang, B.; Liu, L.; Yang, G.; Fang, K.; Chen, Y.; Li, F.; Kou, D.; Ji, C.; et al. Linking temperature sensitivity of soil CO2 release to substrate, environmental, and microbial properties across alpine ecosystems. Glob. Biogeochem. Cycles 2016, 30, 1310–1323. [Google Scholar] [CrossRef]
- Naz, M.; Dai, Z.; Hussain, S.; Tariq, M.; Danish, S.; Khan, I.U.; Qi, S.; Du, D. The soil pH and heavy metals revealed their impact on soil microbial community. J. Environ. Manag. 2022, 321, 115770. [Google Scholar] [CrossRef]
- Shimizu, M.; Wariishi, H. Development of a sample preparation method for fungal proteomics. FEMS Microbiol. Lett. 2005, 247, 17–22. [Google Scholar] [CrossRef]
- Aran, D.; Maul, A.; Masfaraud, J.-F. A spectrophotometric measurement of soil cation exchange capacity based on cobaltihexamine chloride absorbance. C. R. Geosci. 2008, 340, 865–871. [Google Scholar] [CrossRef]
- Banerjee, S.; Helgason, B.; Wang, L.; Winsley, T.; Ferrari, B.C.; Siciliano, S.D. Legacy effects of soil moisture on microbial community structure and N2O emissions. Soil Biol. Biochem. 2016, 95, 40–50. [Google Scholar] [CrossRef]
- Chaudhari, S.K.; Singh, R.; Kundu, D.K. Rapid textural analysis for saline and alkaline soils with different physical and chemical properties. Soil Sci. Soc. Am. J. 2008, 72, 431–441. [Google Scholar] [CrossRef]
- Blanco-Canqui, H. Biochar and Soil Physical Properties. Soil Sci. Soc. Am. J. 2017, 81, 687–711. [Google Scholar] [CrossRef]
- Bell, C.W.; Fricks, B.E.; Rocca, J.D.; Steinweg, J.M.; McMahon, S.K.; Wallenstein, M.D. High-throughput Fluorometric Measurement of Potential Soil Extracellular Enzyme Activities. J. Vis. Exp. 2013, 81, e50961. [Google Scholar] [CrossRef]
- Zhang, X.D.; Amelung, W. Gas chromatographic determination of muramic acid, glucosamine, mannosamine, and galactosamine in soils. Soil Biol. Biochem. 1996, 28, 1201–1206. [Google Scholar] [CrossRef]
- Ma, T.; Dai, G.; Zhu, S.; Chen, D.; Chen, L.; Lu, X.; Wang, X.; Zhu, J.; Zhang, Y.; Ma, W.; et al. Distribution and Preservation of Root- and Shoot-Derived Carbon Components in Soils Across the Chinese-Mongolian Grasslands. J. Geophys. Res.-Biogeosci. 2019, 124, 420–431. [Google Scholar] [CrossRef]
- Prommer, J.; Walker, T.W.N.; Wanek, W.; Braun, J.; Zezula, D.; Hu, Y.; Hofhansl, F.; Richter, A. Increased microbial growth, biomass, and turnover drive soil organic carbon accumulation at higher plant diversity. Glob. Chang. Biol. 2020, 26, 669–681. [Google Scholar] [CrossRef]
- Hu, J.; Huang, C.; Zhou, S.; Liu, X.; Dijkstra, F.A. Nitrogen addition increases microbial necromass in croplands and bacterial necromass in forests: A global meta-analysis. Soil Biol. Biochem. 2022, 165, 108500. [Google Scholar] [CrossRef]
- Mellado-Vazquez, P.G.; Lange, M.; Bachmann, D.; Gockele, A.; Karlowsky, S.; Milcu, A.; Piel, C.; Roscher, C.; Roy, J.; Gleixner, G. Plant diversity generates enhanced soil microbial access to recently photosynthesized carbon in the rhizosphere. Soil Biol. Biochem. 2016, 94, 122–132. [Google Scholar] [CrossRef]
- Jia, Y.; Zhai, G.; Zhu, S.; Liu, X.; Schmid, B.; Wang, Z.; Ma, K.; Feng, X. Plant and microbial pathways driving plant diversity effects on soil carbon accumulation in subtropical forest. Soil Biol. Biochem. 2021, 161, 108375. [Google Scholar] [CrossRef]
- Zhang, B.; Cai, Y.; Hu, S.; Chang, S.X. Plant mixture effects on carbon-degrading enzymes promote soil organic carbon accumulation. Soil Biol. Biochem. 2021, 163, 108457. [Google Scholar] [CrossRef]
- Qi, R.; Li, J.; Lin, Z.; Li, Z.; Li, Y.; Yang, X.; Zhang, J.; Zhao, B. Temperature effects on soil organic carbon, soil labile organic carbon fractions, and soil enzyme activities under long-term fertilization regimes. Appl. Soil Ecol. 2016, 102, 36–45. [Google Scholar] [CrossRef]
- Erdel, E.; Simsek, U.; Kesimci, T.G. Effects of Fungi on Soil Organic Carbon and Soil Enzyme Activity under Agricultural and Pasture Land of Eastern Turkiye. Sustainability 2023, 15, 1765. [Google Scholar] [CrossRef]
- Kim, J.I.; Yang, Y.; Kang, H. Fluorometric assay for phenol oxidase activity in soils and its controlling variables. Appl. Soil Ecol. 2024, 195, 105240. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L. Phenol oxidase, peroxidase and organic matter dynamics of soil. Soil Biol. Biochem. 2010, 42, 391–404. [Google Scholar] [CrossRef]
- Wei, X.; Zhu, Z.; Liu, Y.; Luo, Y.; Deng, Y.; Xu, X.; Liu, S.; Richter, A.; Shibistova, O.; Guggenberger, G.; et al. C:N:P stoichiometry regulates soil organic carbon mineralization and concomitant shifts in microbial community composition in paddy soil. Biol. Fertil. Soils 2020, 56, 1093–1107. [Google Scholar] [CrossRef]
- Luo, L.; Meng, H.; Gu, J.-D. Microbial extracellular enzymes in biogeochemical cycling of ecosystems. J. Environ. Manag. 2017, 197, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Finzi, A.C.; Austin, A.T.; Cleland, E.E.; Frey, S.D.; Houlton, B.Z.; Wallenstein, M.D. Responses and feedbacks of coupled biogeochemical cycles to climate change: Examples from terrestrial ecosystems. Front. Ecol. Environ. 2011, 9, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Zhu, E.; Cao, Z.; Jia, J.; Liu, C.; Zhang, Z.; Wang, H.; Dai, G.; He, J.-S.; Feng, X. Inactive and inefficient: Warming and drought effect on microbial carbon processing in alpine grassland at depth. Glob. Change Biol. 2021, 27, 2241–2253. [Google Scholar] [CrossRef]
- Fang, K.; Kou, D.; Wang, G.; Chen, L.; Ding, J.; Li, F.; Yang, G.; Qin, S.; Liu, L.; Zhang, Q.; et al. Decreased Soil Cation Exchange Capacity Across Northern China’s Grasslands Over the Last Three Decades. J. Geophys. Res.-Biogeosci. 2017, 122, 3088–3097. [Google Scholar] [CrossRef]
- Hall, S.J.; Ye, C.; Weintraub, S.R.; Hockaday, W.C. Molecular trade-offs in soil organic carbon composition at continental scale. Nat. Geosci. 2020, 13, 687. [Google Scholar] [CrossRef]
- Chen, S.; Wang, W.; Xu, W.; Wang, Y.; Wan, H.; Chen, D.; Tang, Z.; Tang, X.; Zhou, G.; Xie, Z.; et al. Plant diversity enhances productivity and soil carbon storage. Proc. Natl. Acad. Sci. USA 2018, 115, 4027–4032. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Yuan, M.; Jian, S.; Gamage, L.; Parajuli, M.; Dzantor, K.E.; Hui, D.; Fay, P.A.; Li, J. Soil extracellular oxidases mediated nitrogen fertilization effects on soil organic carbon sequestration in bioenergy croplands. GCB Bioenergy 2021, 13, 1303–1318. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Van Horn, D.J.; Shah, J.J.F.; Findlay, S. Ecoenzymatic Stoichiometry in Relation to Productivity for Freshwater Biofilm and Plankton Communities. Microb. Ecol. 2010, 60, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guo, X.; Ma, Y.; Hu, B.; Yang, Y.; Tian, H.; Liu, X.; Meng, N.; Zhu, J.; Yan, D.; et al. The hidden risk: Changes in functional potentials of microbial keystone taxa under global climate change jeopardizing soil carbon storage in alpine grasslands. Environ. Int. 2024, 185, 108516. [Google Scholar] [CrossRef]
- Han, X.; Li, Y.; Du, X.; Li, Y.; Wang, Z.; Jiang, S.; Li, Q. Effect of grassland degradation on soil quality and soil biotic community in a semi-arid temperate steppe. Ecol. Process. 2020, 9, 63. [Google Scholar] [CrossRef]
Figure 1.
Description of sampling sites. Red circles represent sampling sites.
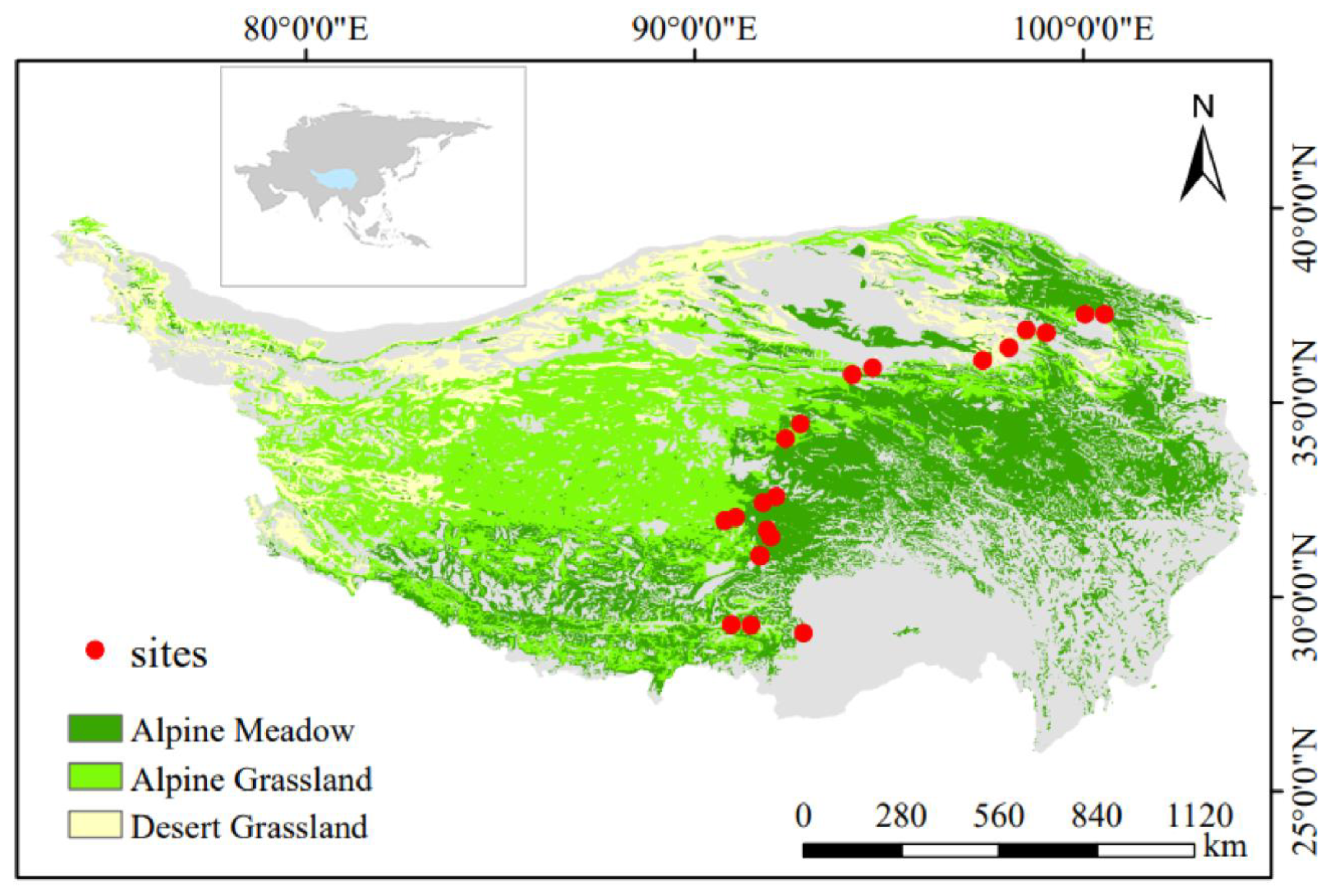
Figure 2.
Amino sugar content of surface and subsoil of different grassland types. Differences between grasslands at the same soil depth were tested using a one-way ANOVA test, and results are indicated by capital letters. Differences in soil depth under the same vegetation type were tested using a paired-sample t-test, and results are indicated by lowercase letters. The upper and lower boundaries of the box plots represent the 25% and 75% quantile values of the data; the top and bottom short lines of the box plots represent the maximum and minimum values of the data; and the solid lines and black dots in the box plots represent the median and the mean, respectively. p < 0.05 are significant differences. AM: alpine meadow; AG: alpine grassland; DG: desert grassland.
Figure 2.
Amino sugar content of surface and subsoil of different grassland types. Differences between grasslands at the same soil depth were tested using a one-way ANOVA test, and results are indicated by capital letters. Differences in soil depth under the same vegetation type were tested using a paired-sample t-test, and results are indicated by lowercase letters. The upper and lower boundaries of the box plots represent the 25% and 75% quantile values of the data; the top and bottom short lines of the box plots represent the maximum and minimum values of the data; and the solid lines and black dots in the box plots represent the median and the mean, respectively. p < 0.05 are significant differences. AM: alpine meadow; AG: alpine grassland; DG: desert grassland.
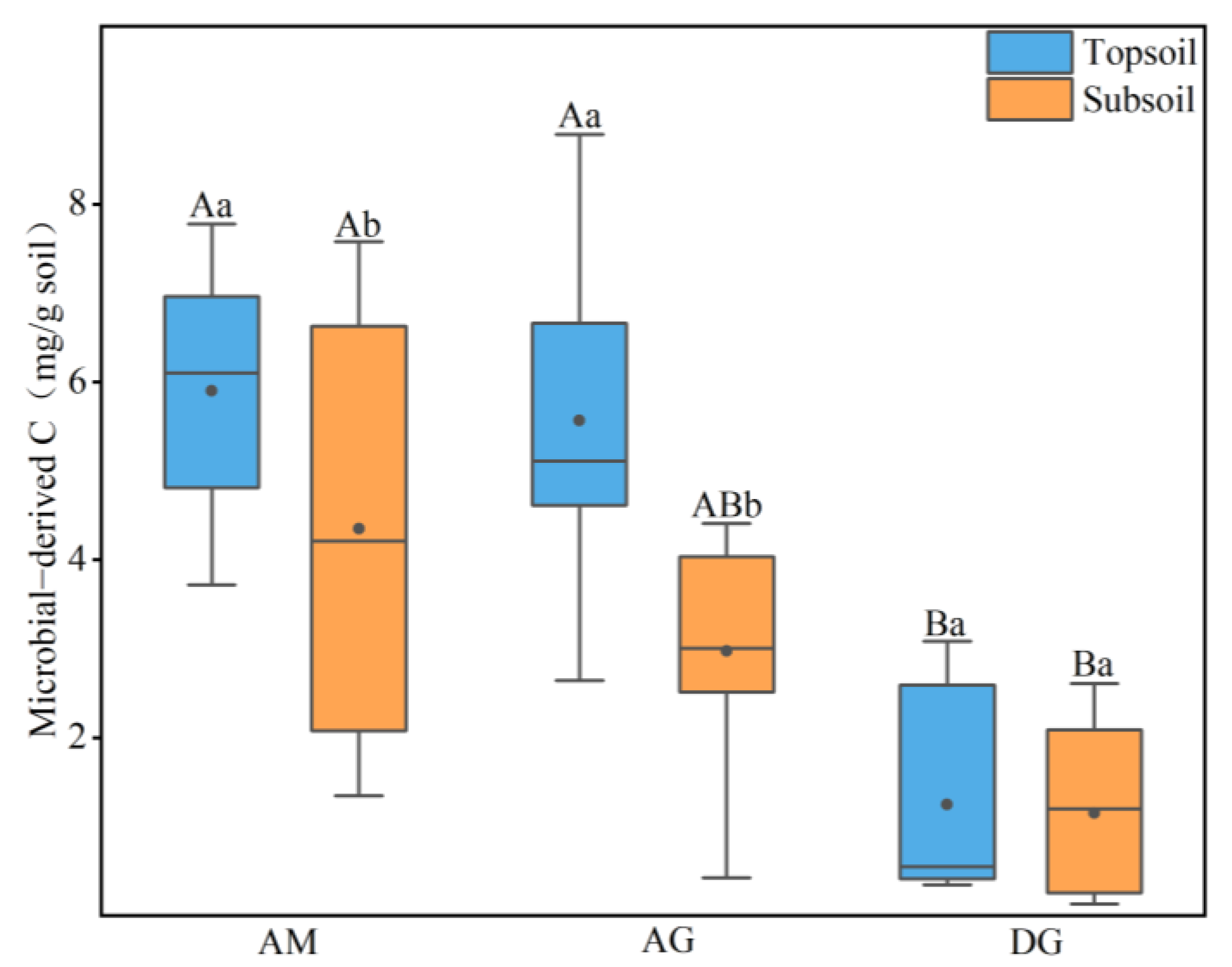
Figure 3.
Surface and subsoil enzyme activities in different grassland types. (a) soil oxidase; (b) soil β-glucosidase; (c) soil cellulase. Differences between grasslands at the same soil depth were tested using a one-way ANOVA test, with results indicated by capital letters. Differences in soil depth under the same vegetation type were tested using a paired-sample t-test, and results are indicated by lowercase letters. The upper and lower boundaries of the box plots represent the 25% and 75% quantile values of the data; the top and bottom shortest lines of the box plots represent the maximum and minimum values of the data; and the solid lines and black dots in the box plots represent the median and the mean values, respectively. p < 0.05 is significantly different. AM: alpine meadow; AG: alpine grassland; DG: desert grassland; POX+PER: polyphenol oxidase+peroxidase; BG: β-glucosidase; CB: cellulase.
Figure 3.
Surface and subsoil enzyme activities in different grassland types. (a) soil oxidase; (b) soil β-glucosidase; (c) soil cellulase. Differences between grasslands at the same soil depth were tested using a one-way ANOVA test, with results indicated by capital letters. Differences in soil depth under the same vegetation type were tested using a paired-sample t-test, and results are indicated by lowercase letters. The upper and lower boundaries of the box plots represent the 25% and 75% quantile values of the data; the top and bottom shortest lines of the box plots represent the maximum and minimum values of the data; and the solid lines and black dots in the box plots represent the median and the mean values, respectively. p < 0.05 is significantly different. AM: alpine meadow; AG: alpine grassland; DG: desert grassland; POX+PER: polyphenol oxidase+peroxidase; BG: β-glucosidase; CB: cellulase.
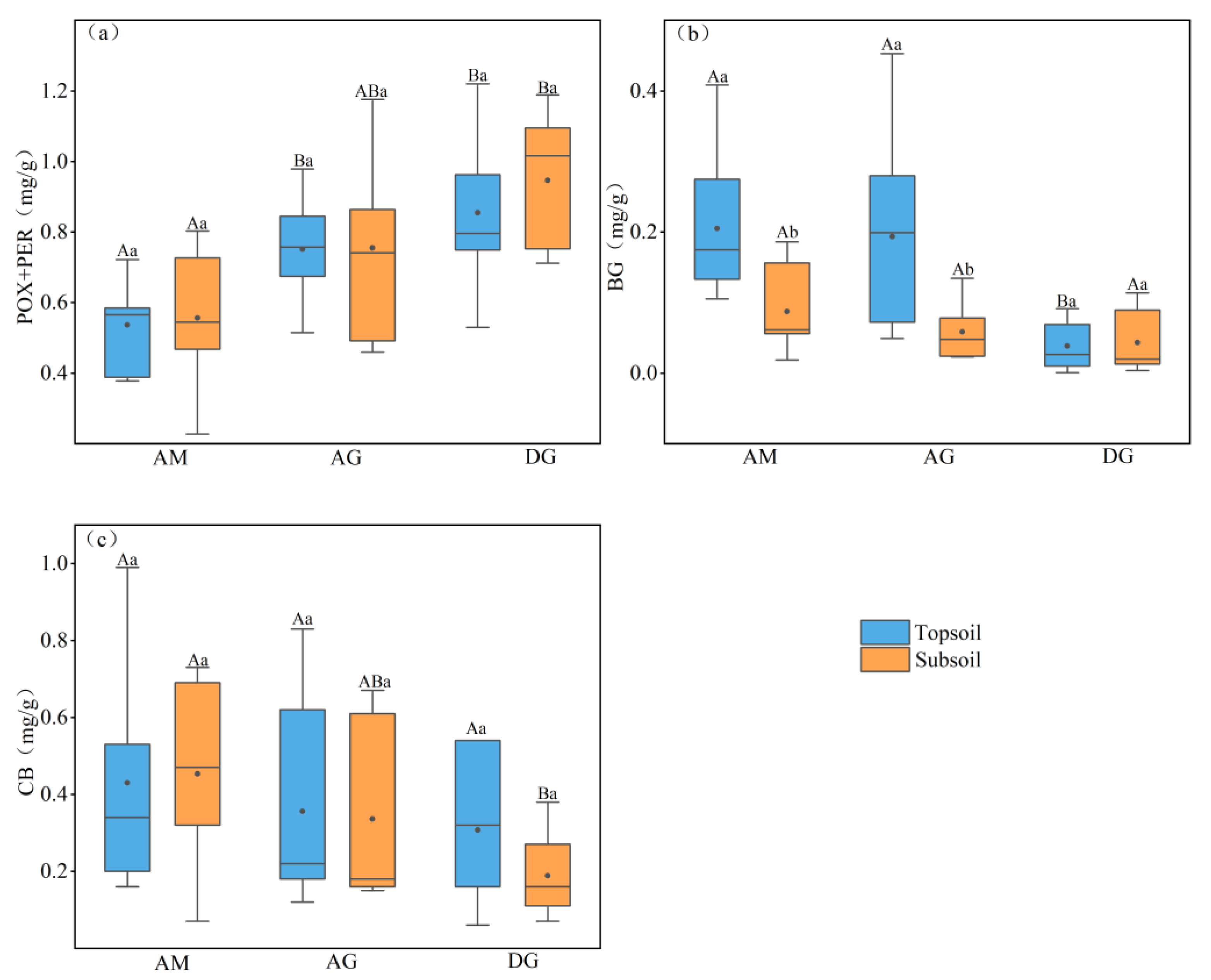
Figure 4.
Relationship between soil amino sugars content and soil extracellular enzyme activity. (a) soil polyphenol oxidase+peroxidase; (b) soil BG; (c) soil CB. Solid lines indicate the linear relationship between extracellular enzyme activities and amino sugars for each soil type. p < 0.05 is significantly different. AM: alpine meadow; AG: alpine grassland; DG: desert grassland; POX+PER: polyphenol oxidase+peroxidase; BG: β-glucosidase; CB: cellulase.
Figure 4.
Relationship between soil amino sugars content and soil extracellular enzyme activity. (a) soil polyphenol oxidase+peroxidase; (b) soil BG; (c) soil CB. Solid lines indicate the linear relationship between extracellular enzyme activities and amino sugars for each soil type. p < 0.05 is significantly different. AM: alpine meadow; AG: alpine grassland; DG: desert grassland; POX+PER: polyphenol oxidase+peroxidase; BG: β-glucosidase; CB: cellulase.
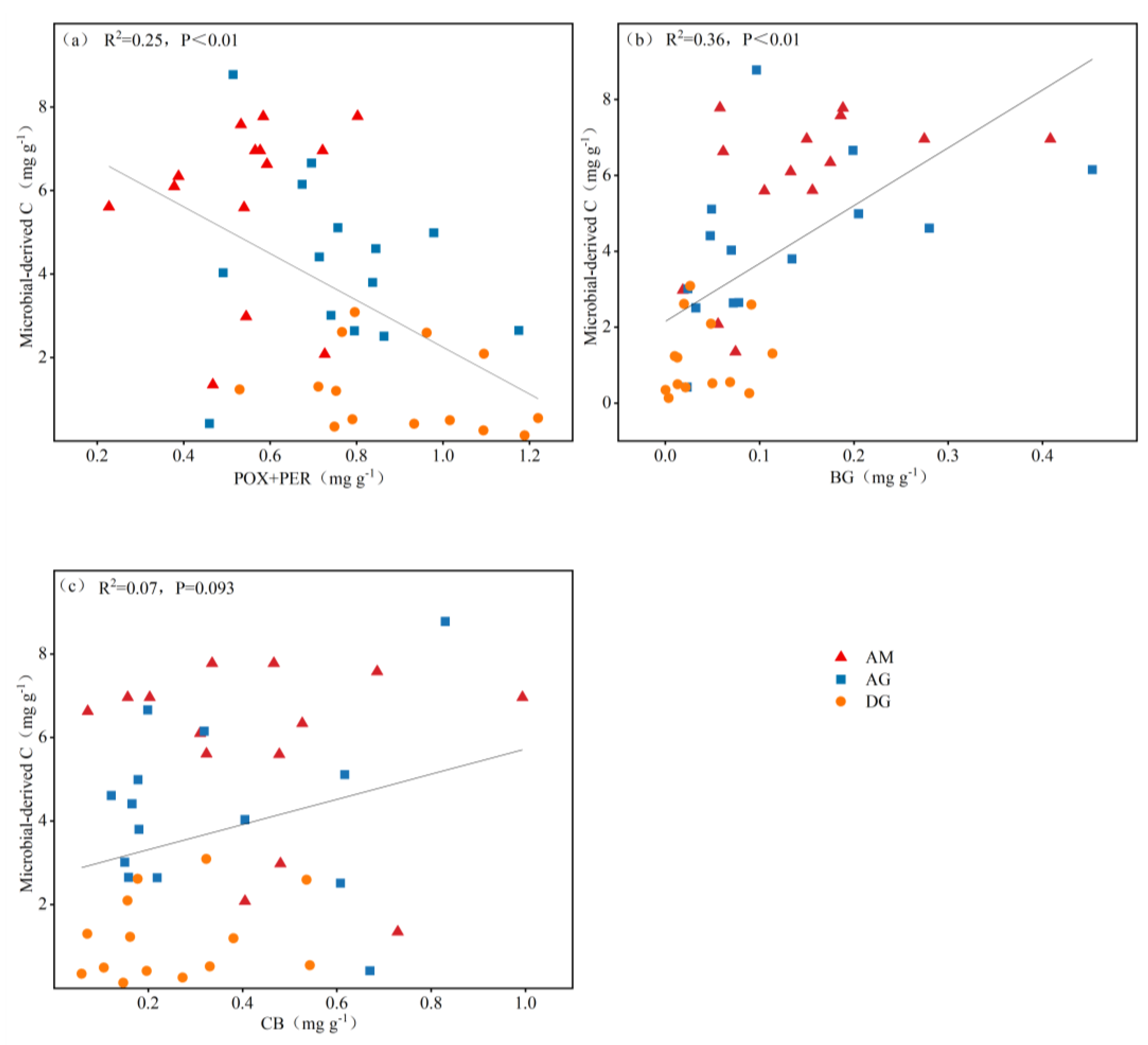
Figure 5.
Correlation of soil microbial sources of carbon, oxidative enzymes, BG, and CB with environmental parameters. Positive correlations are indicated in red, negative correlations in blue, and the strength of the correlation is indicated by the color shade; the stronger the correlation, the darker the color. An asterisk (*) indicates the significance of the correlation, with 1, 2, and 3 asterisks denoting p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001, respectively. POX+PER: polyphenol oxidase+peroxidase; BG: β-glucosidase; CB: cellulase; TN: total nitrogen; TP: total phosphorus; EC: electrical conductivity; Clay: clay granule; Silt: sand granule; Sand: powder granule; CEC: cation exchange capacity; MC: moisture content.
Figure 5.
Correlation of soil microbial sources of carbon, oxidative enzymes, BG, and CB with environmental parameters. Positive correlations are indicated in red, negative correlations in blue, and the strength of the correlation is indicated by the color shade; the stronger the correlation, the darker the color. An asterisk (*) indicates the significance of the correlation, with 1, 2, and 3 asterisks denoting p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001, respectively. POX+PER: polyphenol oxidase+peroxidase; BG: β-glucosidase; CB: cellulase; TN: total nitrogen; TP: total phosphorus; EC: electrical conductivity; Clay: clay granule; Silt: sand granule; Sand: powder granule; CEC: cation exchange capacity; MC: moisture content.
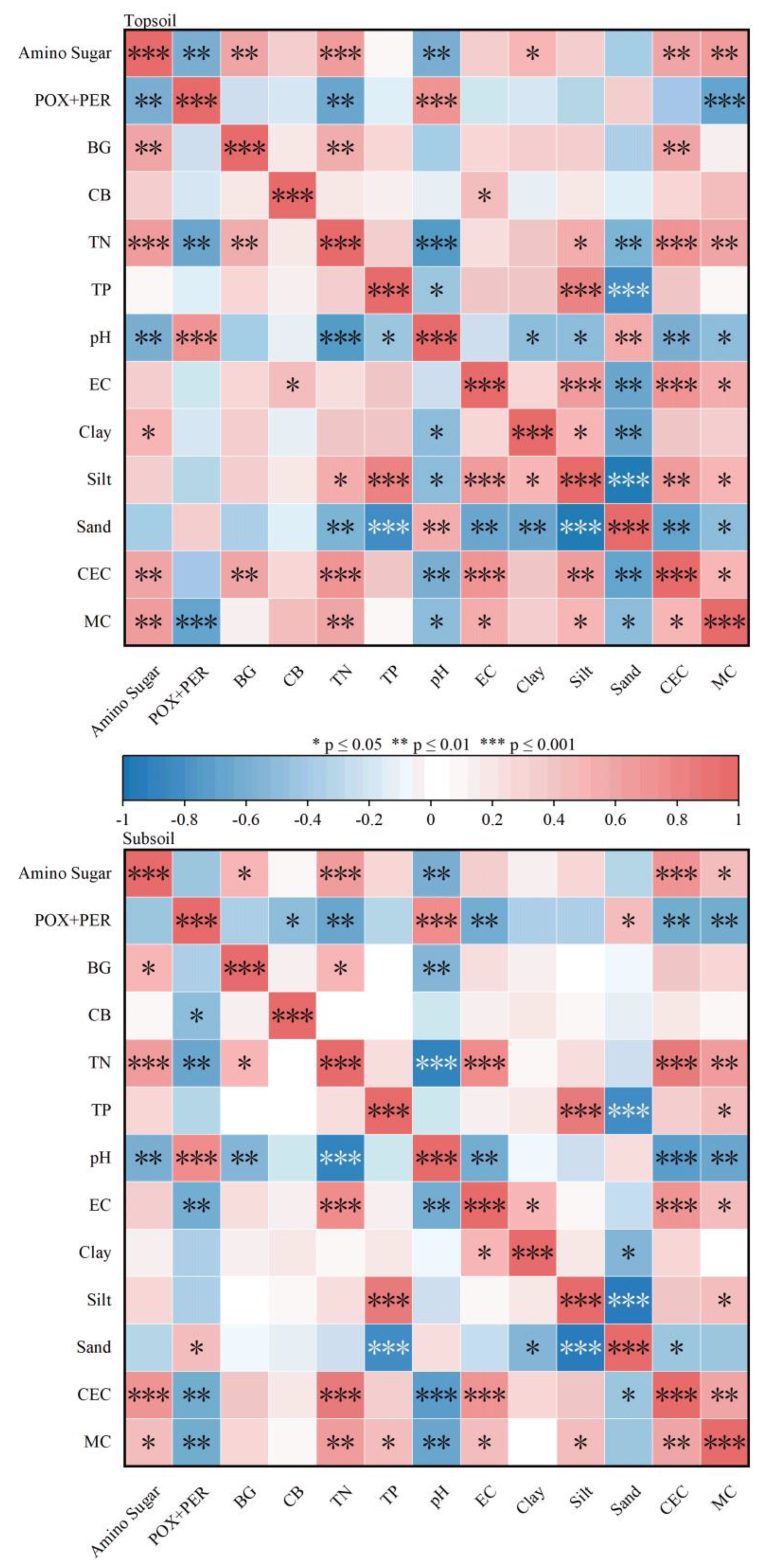
Figure 6.
Relative importance of environmental parameters. (a) topsoil amino sugar principal component analysis; (b) subsoil amino sugar principal component analysis. Principal component analysis (PCA) was used to analyze the significance of environmental parameters for amino sugars, BG, CB, and POX+PER in topsoil and subsoil. AM: alpine meadow; AG: alpine grassland; DG: desert grassland; POX+PER: polyphenol oxidase+peroxidase; BG: β-glucosidase; CB: cellulase; TN: total nitrogen; TP: total phosphorus; EC: electrical conductivity; Clay: clay particles; Silt: powder particles; Sand: sand particles; CEC: cation exchange capacity; MC: moisture content.
Figure 6.
Relative importance of environmental parameters. (a) topsoil amino sugar principal component analysis; (b) subsoil amino sugar principal component analysis. Principal component analysis (PCA) was used to analyze the significance of environmental parameters for amino sugars, BG, CB, and POX+PER in topsoil and subsoil. AM: alpine meadow; AG: alpine grassland; DG: desert grassland; POX+PER: polyphenol oxidase+peroxidase; BG: β-glucosidase; CB: cellulase; TN: total nitrogen; TP: total phosphorus; EC: electrical conductivity; Clay: clay particles; Silt: powder particles; Sand: sand particles; CEC: cation exchange capacity; MC: moisture content.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, Y.; Wang, X.; Sun, Y.; Wu, J.; Deng, T.; Yuan, M.; Duan, W.; Zhao, Y. Hydrolases Control Soil Carbon Sequestration in Alpine Grasslands in the Tibetan Plateau. Sustainability 2024, 16, 3508. https://doi.org/10.3390/su16093508
AMA Style
Zhang Y, Wang X, Sun Y, Wu J, Deng T, Yuan M, Duan W, Zhao Y. Hydrolases Control Soil Carbon Sequestration in Alpine Grasslands in the Tibetan Plateau. Sustainability. 2024; 16(9):3508. https://doi.org/10.3390/su16093508
Chicago/Turabian StyleZhang, Yuanye, Xia Wang, Yuxin Sun, Jinhong Wu, Tao Deng, Menghan Yuan, Wenhui Duan, and Yunfei Zhao. 2024. "Hydrolases Control Soil Carbon Sequestration in Alpine Grasslands in the Tibetan Plateau" Sustainability 16, no. 9: 3508. https://doi.org/10.3390/su16093508
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.