Intake of Polydextrose Alters Hematology and the Profile of Short Chain Fatty Acids in Partially Gastrectomized Rats

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Conditions
2.2. Surgical Procedure
2.3. Postoperative Care
2.4. Experimental Design
2.5. Experimental Diets
2.6. Supplementation of Vitamin B12 (Cyanocobalamin)
2.7. Euthanasia
2.8. Measurement of pH of Cecal Contents
2.9. Analytical Methods
2.9.1. Percentage Composition of Formulated Diets
2.9.2. Determination of Iron Content in Diets and Liver Samples
2.9.3. Hematocrit, Hemoglobin, Mean Corpuscular Volume and Mean Corpuscular Hemoglobin Concentration
2.9.4. Collection of Blood Samples for Serum Iron Parameters
2.9.5. Protein Analysis by Western Blotting
2.9.6. Collection of Cecal Contents of Animals
2.9.7. Characterization and Quantification of Fatty Acids
2.9.8. Statistical Analyzes
3. Results
3.1. Composition of Experimental Diets
3.2. Dietary Intake, Weight Gain and Feed Efficiency Coefficient (FEC)
3.3. Fermentation Parameters
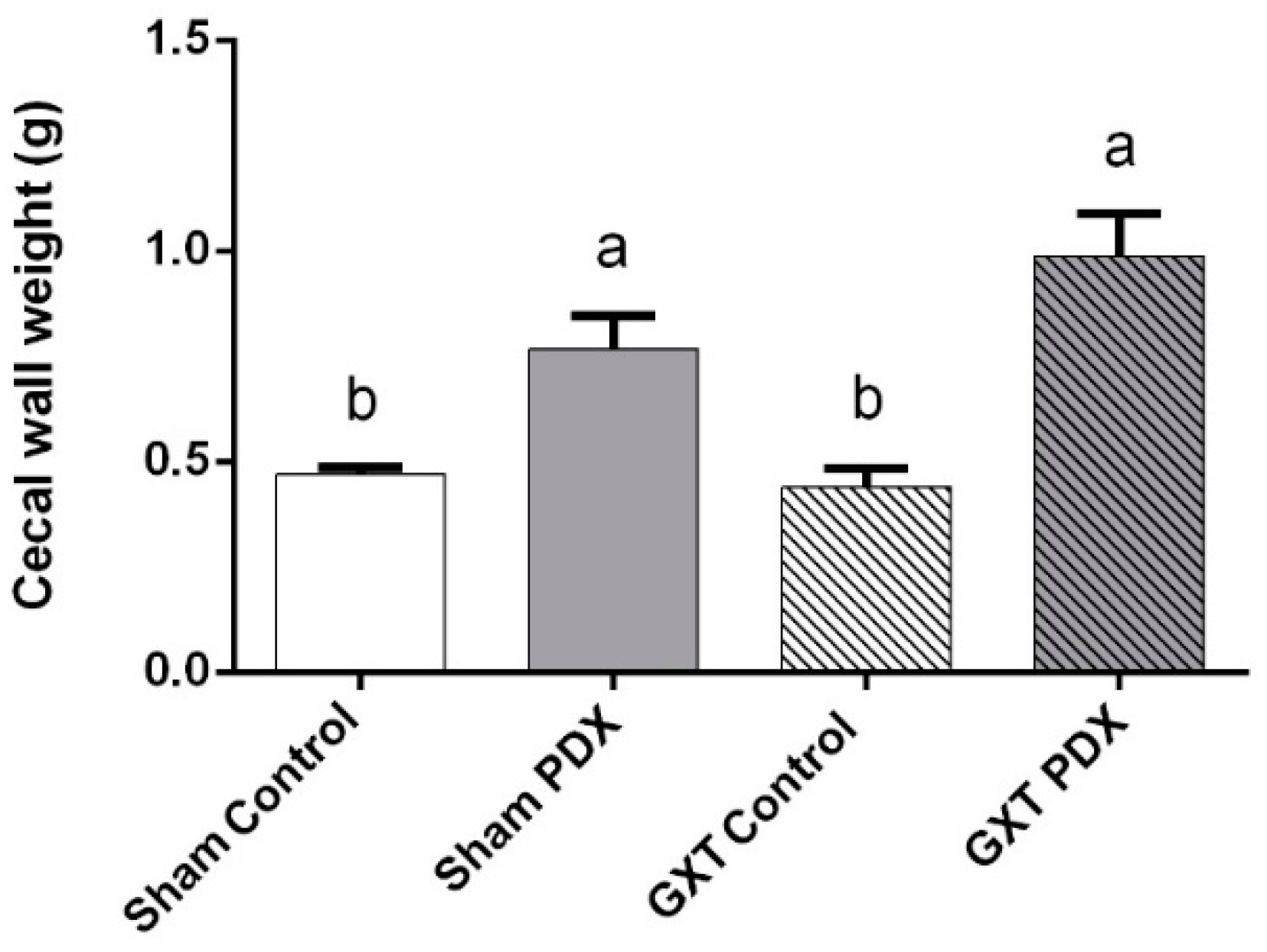
3.3.1. Cecal Wall Weight
3.3.2. pH of Cecal Contents
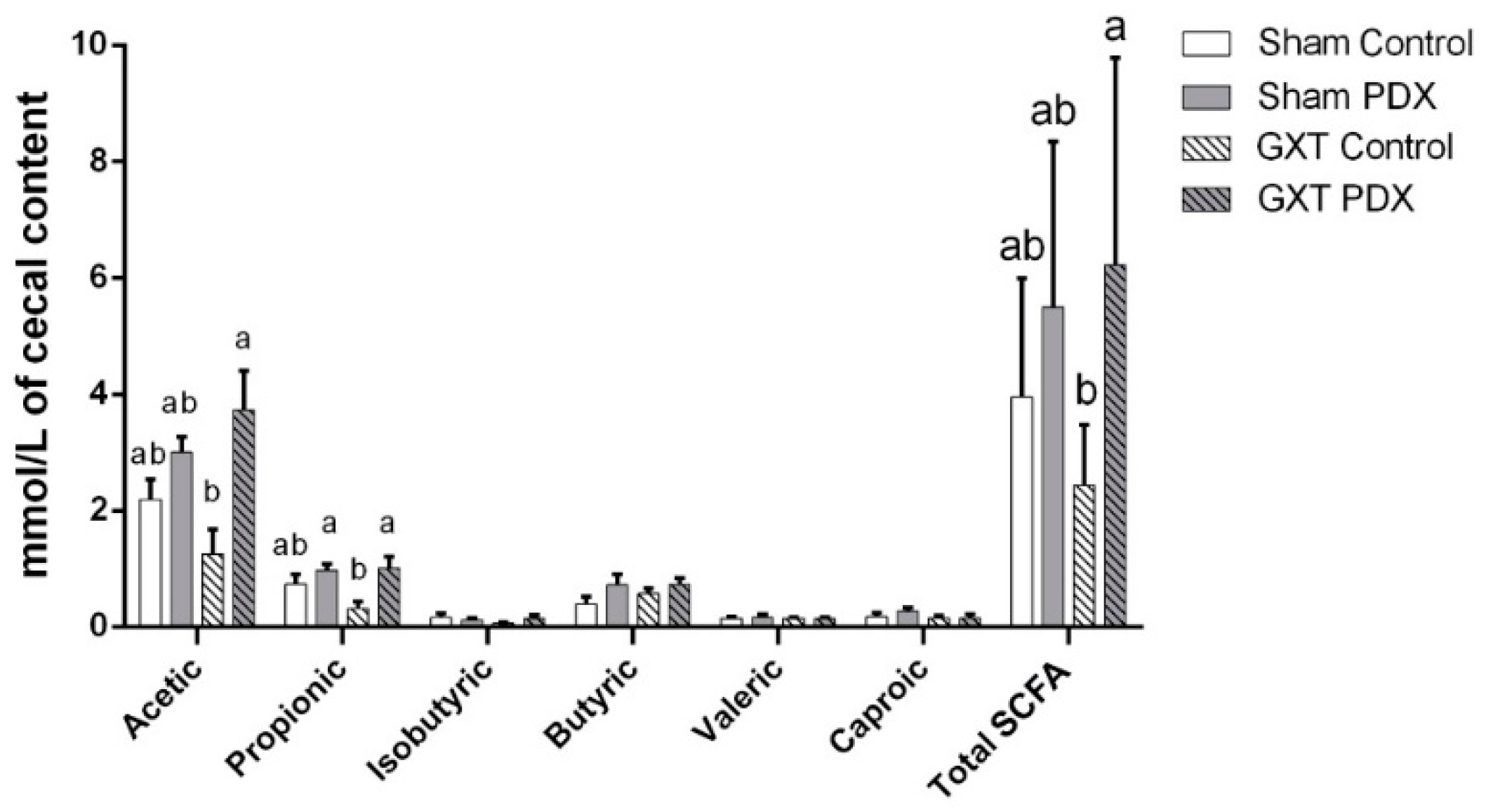
3.3.3. Characterization and Quantification of Short-Chain Fatty Acids (SCFA)
3.4. Hematologic Parameters
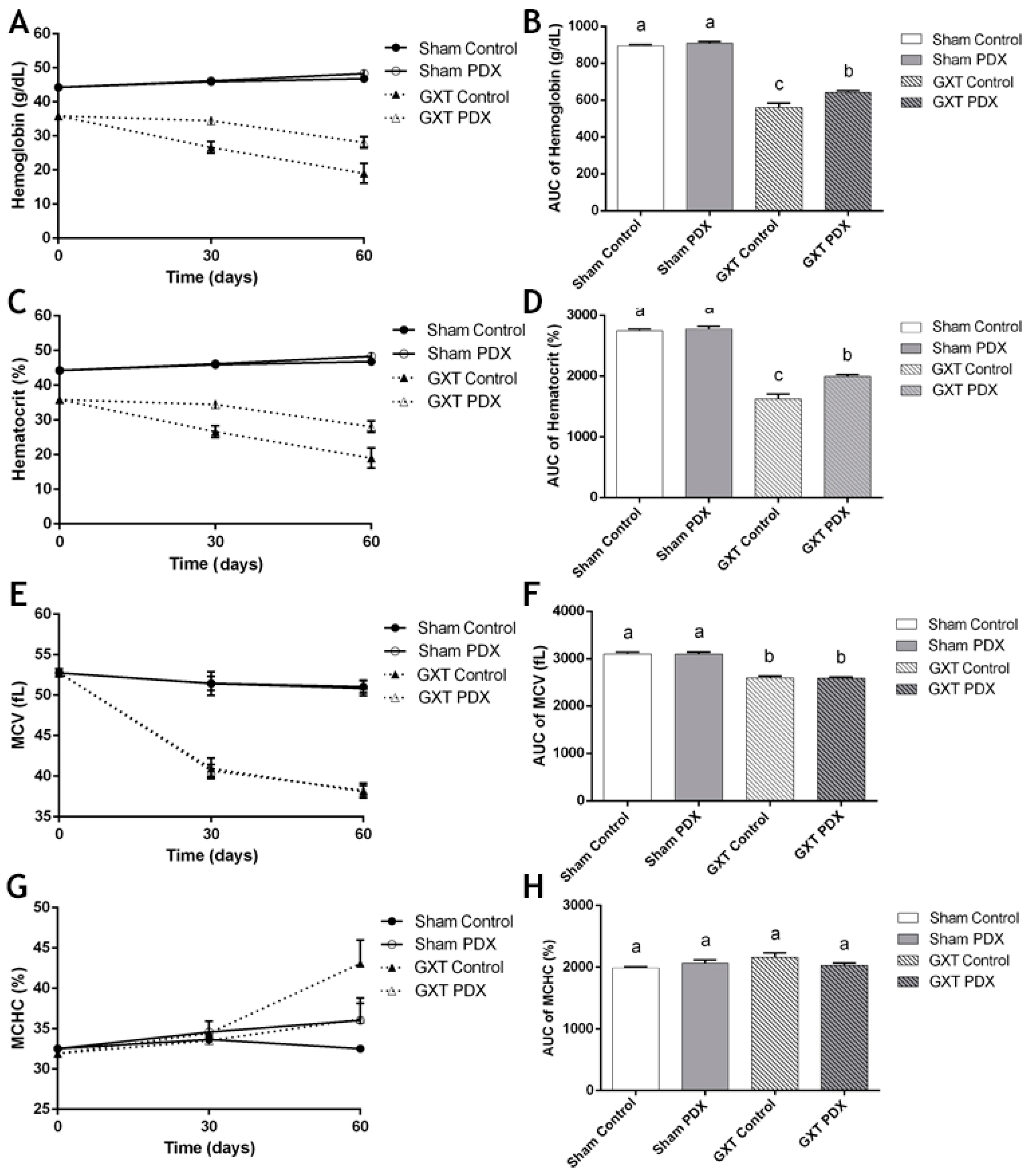
3.4.1. Hemoglobin (HGB), Hematocrit (HCT), Mean Corpuscular Volume (MCV) and Mean Corpuscular Hemoglobin Concentration (MCHC)
3.4.2. Serum Iron Parameters
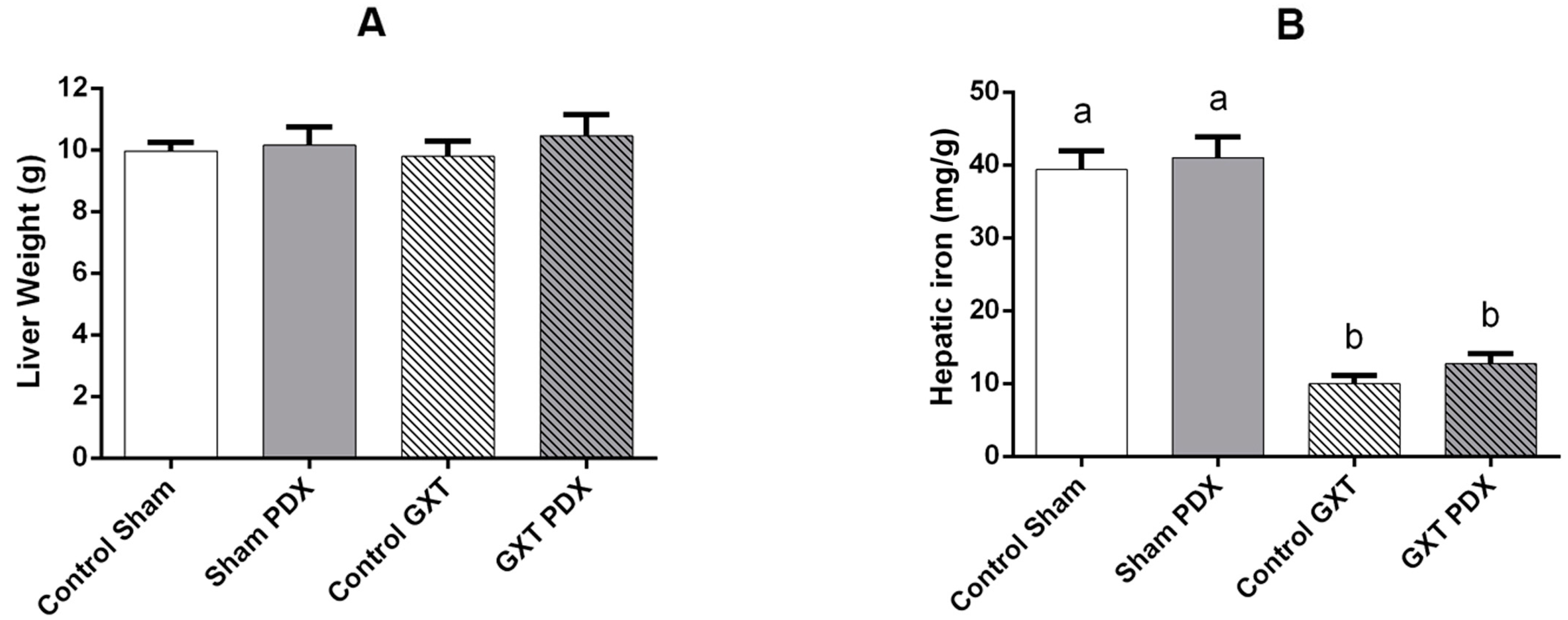
3.5. Reserve Parameters
Ferroportin Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Instituto Nacional De Câncer José Alencar Gomes Da Silva (INCA). Câncer Gástrico. Instituto Nacional de Câncer José Alencar Gomes da Silva, Rio de Janeiro. 2016. Available online: http://www2.inca.gov.br/wps/wcm/connect/tiposdecancer/site/home/estomago (accessed on 2 September 2016).
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Chung, M.C.M. The gastric fluid proteome as a potential source of gastric cancer biomarkers. J. Proteom. 2013, 90, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Yakirevich, E.; Resnick, M.B. Pathology of gastric cancer and its precursor lesions. Gastroenterol. Clin. N. Am. 2013, 42, 261–284. [Google Scholar] [CrossRef] [PubMed]
- Papini-Berto, S.J.; Burini, R.C. Causas da desnutrição pós-gastrectomia. Arq. Gastroenterol. 2001, 38, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Papini-Berto, S.J. Desnutrição protéico-energética no paciente gastrectomizado. Arq. Gastroenterol. 2002, 39, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, H.; Sawyer, M.B.; Wischmeyer, P.E.; Baracos, V.E. Nutrition modulation of gastrointestinal toxicity related to cancer chemotherapy: From preclinical findings to clinical strategy. JPEN J. Parenter. Enter. Nutr. 2011, 32, 74–90. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.S.; Gomez-Ayala, A.E.; Lopez-Aliaga, I.; Pallares, I.; Hartiti, S.; Pharm, B.; Alferez, M.J.M.; Barronuevo, M. Role of the proximal colon in mineral absorption in rats with and without ferropenic anemia. Nutr. Res. 1996, 16, 1529–1543. [Google Scholar] [CrossRef]
- Bouglé, D.; Vaghefi-Vaezzadeh, N.; Roland, N.; Roland, N.; Bouvard, G.; Arhan, P.; Bureau, F.; Neuville, D.; Maubois, J.L. Influence of short-chain fatty acids on iron absorption by proximal colon. Scand. J. Gastroenterol. 2002, 37, 1008–1011. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.F.; Tsuboi, K.H.; Araújo, M.R.; Falconi, M.A.; Ouwehand, A.C.; Andreollo, N.A.; Miyasaka, C.K. Ingestion of polydextrose increase the iron absorption in rats submitted to partial gastrectomy. Acta Cir. Bras. 2010, 25, 518–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz-Ahrens, K.E.; Schrezenmeir, J. Inulin and oligofructose and mineral metabolism: The evidence from animal trials. J. Nutr. 2007, 137, 2513S–2523S. [Google Scholar] [CrossRef] [PubMed]
- Do Carmo, M.M.R.; Walker, J.C.L.; Novello, D.; Caselato, V.M.; Sgarbieri, V.C.; Ouwehand, A.C.; Andreollo, N.A.; Hiane, P.A.; Santos, E.F. Polydextrose: Physiological Function, and Effects on Health. Nutrients 2016, 8, 553. [Google Scholar] [CrossRef] [PubMed]
- Freitas, K.C.; Amancio, O.M.; Ferreira, N.N.; Fagundes-Neto, U.; de Morais, M.B. Partially hydrolyzed guar gum increases intestinal absorption of iron in growing rats with iron deficiency anemia. Clin. Nutr. 2006, 25, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Marciano, R.; Santamarina, A.B.; De Santana, A.A.; Silva, M.L.; Amancio, O.M.; Do Nascimento, C.M.; Oyama, L.M.; De Morais, M.B. Effects of prebiotic supplementation on the expression of proteins regulating iron absorption in anaemic growing rats. Br. J. Nutr. 2015, 113, 901–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, L.; Brait, D.; Vaz, M.; Lollo, P.; Morato, P.; Oesterreich, S.; Raposo, J., Jr.; Freitas, K. Partially hydrolyzed guar gum increases ferroportin expression in the colon of anemic growing rats. Nutrients 2017, 9, 228. [Google Scholar] [CrossRef] [PubMed]
- Murphy, O. Non-polyol low-digestible carbohydrates: Food applications and functional benefits. Br. J. Nutr. 2001, 85, S47–S53. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Glahn, R.P.; Welch, R.M.; Lei, X.; Yasuda, K.; Miller, D.D. Dietary inulin affects the expression of intestinal enterocyte iron transporters, receptors and storage protein and alters the microbiota in the pig intestine. Br. J. Nutr. 2008, 99, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Pitarresi, G.; Tripodo, G.; Cavallaro, G.; Palumbo, F.S.; Giammona, G. Inulin-iron complexes: A potential treatment of iron deficiency anaemia. Eur. J. Pharm. Biopharm. 2008, 68, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Lahtinen, S.J.; Knoblock, K.; Drakoularakou, A.; Jacob, M.; Stowell, J.; Gibson, G.R.; Ouwehand, A.C. Effect of molecule branching and glycosidic linkage on the degradation of polydextrose by gut microbiota. Biosci. Biotechnol. Biochem. 2010, 74, 2016–2021. [Google Scholar] [CrossRef] [PubMed]
- Gijs, B.; Eunen, K.V.; Groen, A.K.; Venema, K.; Dirk-Jan, R.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Röytiö, H.; Ouwehand, A.C. The fermentation of polydextrose in the large intestine and its beneficial effects. Benef. Microbes 2014, 3, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Boler, B.M.V.; Serao, M.C.R.; Bauer, L.L.; Staeger, M.A.; Boileau, T.W.; Swanson, K.S.; Fahey, G.C., Jr. Digestive physiological outcomes related to polydextrose and soluble maize fibre consumption by healthy adult men. Br. J. Nutr. 2011, 106, 1864–1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dostal, A.; Chassard, C.; Hilty, F.M.; Zimmermann, M.B.; Jaeggi, T.; Rossi, S.; Lacroix, C. Iron depletion and repletion with ferrous sulfate or electrolytic iron modifies the composition and metabolic activity of the gut microbiota in rats. J. Nutr. 2012, 142, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Mugambi, M.N.; Musekiwa, A.; Lombard, M.; Young, T.; Blaauw, R. Symbiotics, probiotics or prebiotics in infant formula for full term infants: A systematic review. Nutr. J. 2012, 4, 11–18. [Google Scholar] [CrossRef]
- Tako, E.; Glahn, R.P.; Knez, M.; Stangoulis, J.C. The effect of wheat prebiotics on the gut bacterial population and iron status of iron deficient broiler chickens. Nutr. J. 2014, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.S.M.; Gibson, P.R. The trophic effect of dietary fibre is not associated with a change in total crypt number in the distal colon of rats. Carcinogenesis 2003, 24, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Kien, C.L.; Blauwiekel, R.; Bunn, J.Y.; Jetton, T.L.; Frankel, W.L.; Holst, J.J. Cecal infusion of butyrate increases intestinal cell proliferation in piglets. J. Nutr. 2007, 137, 916–922. [Google Scholar] [CrossRef] [PubMed]
- Miyazato, S.; Nakagawa, C.; Kishimoto, Y.; Tagami, H.; Hara, H. Promotive effects of resistant maltodextrin on apparent absorption of calcium, magnesium, iron and zinc in rats. Eur. J. Nutr. 2010, 49, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Lopez, H.W.; Levrat-Verny, M.A.; Coudray, C.; Besson, C.; Krespine, V.; Messager, A.; Demigné, C.; Rémésy, C. Class 2 resistant starches lower plasma and liver lipids and improve mineral retention in rats. J. Nutr. 2001, 131, 1283–1289. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Conesa, D.; López, G.; Ros, G. Effects of probiotic, prebiotic and synbiotic follow-up infant formulas on large intestine morphology and bone mineralisation in rats. J. Sci. Food Agric. 2007, 87, 1059–1068. [Google Scholar] [CrossRef]
- Lobo, A.R.; Gaievski, E.H.S.; Carli, E.D.E.; Alvares, E.P.; Colli, E. Fructo-oligosaccharides and iron bioavailability in anaemic rats: The effects on iron species distribution, ferroportin-1 expression, crypt bifurcation and crypt cell proliferation in the caecum. Br. J. Nutr. 2014, 112, 1286–1295. [Google Scholar] [CrossRef] [PubMed]
- Legette, L.L.; Lee, W.; Martin, B.R.; Story, J.A.; Campbell, J.K.; Weaver, C.M. Prebiotics enhance magnesium absorption and inulin-based fibers exert chronic effects on calcium utilization in a postmenopausal rodent model. J. Food Sci. 2012, 77, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.F.; Tsuboi, K.H.; Araújo, M.R.; Ouwehand, A.C.; Andreollo, N.A.; Miyasaka, C.K. Dietary polydextrose increases calcium absorption in normal rats. Arq. Bras. Cir. Dig. 2009, 22, 201–205. [Google Scholar] [CrossRef]
- Hara, H.; Suzuki, T.; Aoyama, Y. Ingestion of the soluble dietary fibre, polydextrose, increases calcium absorption and bone mineralization in normal and total-gastrectomized rats. Br. J. Nutr. 2000, 84, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Yeung, C.K.; Glahn, R.E.; Welch, R.M.; Miller, D.D. Prebiotics and Iron Bioavailability—Is There a Connection? J. Food Sci. 2005, 70, 88–92. [Google Scholar] [CrossRef]
- Sheridan, P.O.; Bindels, L.B.; Saulnier, D.M.; Reid, G.; Nova, E.; Holmgren, K.; O’Toole, P.W.; Bunn, J.; Delzenne, N.; Scott, K.P. Can prebiotics and probiotics improve therapeutic outcomes for undernourished individuals? Gut Microbes 2014, 1, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition Ad Hoc writing Committee on their formulation of the AIN-76A Rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.F.; Tsuboi, K.H.; Palu, B.F.; Araújo, M.R.; Andreollo, N.A.; Miyasaka, C.K. Effects of the partial gastrectomy associated to anterior truncal vagotomy in the metabolism of the iron in rats. Acta Cir. Bras. 2008, 23, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Nery, C.D.S.; Pinheiro, I.L.; Muniz, G.D.S.; Vasconcelos, D.A.A.D.; França, S.P.D.; Nascimento, E.D. Medidas murinométricas e eficiência alimentar em ratos provenientes de ninhadas reduzidas na lactação e submetidos ou não ao exercício de natação. Rev. Bras. Med. Esporte 2011, 17, 49–55, ISSN 1517-8692. [Google Scholar] [CrossRef] [Green Version]
- Ohta, A.; Ohtsuki, M.; Hosono, A.; Adachi, T.; Hara, H.; Sakata, T. Dietary fructooligosaccharides prevent osteopenia after gastrectomy in rats. J. Nutr. 1998, 128, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.F.; Tsuboi, K.H.; Araújo, M.R.; Falconi, M.A.; Oouwehand, A.C.; Andreollo, N.A.; Miyasaka, C.K. A ingestão de prebióticos previne a malabsorção de ferro e anemia induzidas pela gastrectomia? Estudo experimental em ratos. Arq. Bras. Cir. Dig. 2011, 24, 9–14. [Google Scholar] [CrossRef]
- Younes, H.; Coudray, C.; Bellanger, J.; Demigné, C.; Rayssiguier, C. Effects of two fermentable carbohydrates (inulin and resistant starch) and their combination on calcium and magnesium balance in rats. Br. J. Nutr. 2001, 86, 479–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horwitz, W. Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC), 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method for total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Petry, C.F.; Pozebon, D.; Bentlin, F.R.S. Evaluation of ICP-OES applicability for trace element determination in environmental samples. At. Spectrosc. 2006, 26, 19–27. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Zhao, G.; Nyman, M.; Jönsson, J.A. Rapid determination of short-chain fatty acids in colonic contents and faeces of humans and rats by acidified waterextraction and direct-injection gas chromatography. Biomed. Chromatogr. 2006, 20, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Zittel, T.T.; Zeeb, B.; Maier, G.W.; Kaiser, G.W.; Zwirner, M.; Liebich, H.; Starlinger, M.; Becker, H.D. High prevalence of bone disorders after gastrectomy. Am. J. Surg. 1997, 174, 431–438. [Google Scholar] [CrossRef]
- Hull, S.; Re, R.; Tiiohonen, K.; Viscione, L.; Wickham, M. Consuming polydextrose in a mid-morning snack increases acute satiety measurements and reduces subsequent energy intake at lunch in healthy human subjects. Appetite 2012, 59, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Schwab, U.; Louheranta, A.; Torronen, A.; Uusitupa, M. Impact of sugar beet pectin and polydextrose on fasting and postprandial glycemia and fasting concentrations of serum total and lipoprotein lipids in middle-aged subjects with abnormal glucose metabolism. Eur. J. Clin. Nutr. 2006, 60, 1073–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astbury, N.M.; Taylor, M.A.; Macdonald, I.A. Polydextrose results in a dose-dependent reduction in ad libitum energy intake at a subsequent test meal. Br. J. Nutr. 2013, 110, 934–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarra, A.; Astbury, N.M.; Olli, K.; Alhoniemi, E.; Tiihonen, K. Effects of polydextrose on different levels of energy intake. A systematic review and meta-analysis. Appetite 2015, 87, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, A.; Astbury, N.M.; Olli, K.; Alhoniemi, E.; Tiihonen, K. Effect of polydextrose on subjective feelings of appetite during the satiation and satiety periods: A systematic review and meta-analysis. Nutrients 2016, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Habib, N.C.; Honoré, S.M.; Geneta, S.B.; Sánchez, S.S. Hypolipidemic effect of Smallanthus sonchifolius (yacon) roots on diabetic rats: Biochemical approach. Chem. Biol. Interact. 2011, 194, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.J. The physiology of glucagon-like-peptide 1. Physiol. Rev. 2007, 87, 1409–1439. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Martin, R.J.; Tulley, R.T.; Raggio, A.M.; Mccutcheon, K.L.; Shen, L.; Danna, S.C.; Tripathy, S.; Hegsted, M.; Keenan, M.J. Dietaryresistant starch upregulates total GLP-1 and PYY in a sustained day-long manner through fermentation in rodents. Am. J. Physiol. Endocrinol. Metab. 2008, 295, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, C.; Caudwell, P.; Finlayson, G.; Webb, D.L.; Hellström, P.M.; Näslund, E.; Blundell, J.E. Comparison of postprandial profiles of ghrelin, active GLP-1, and total PYY to meals varying in fat and carbohydrate and their association with hunger and the phases of satiety. J. Clin. Endocrinol. Metab. 2013, 98, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Arumugam, V.; Haugabrooks, E.; Williamson, P.; Hendrich, S. Soluble dietary fiber (Fibersol-2) decreased hunger and increased satiety hormones in humans when ingested with a meal. Nutr. Res. 2015, 35, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Gupta, A.K. Applications of inulin and oligofructose in health and nutrition. J. Biosci. 2002, 27, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Hooda, S.; Boler, B.M.V.; Serao, M.C.R.; Brulc, J.M.; Staeger, M.A.; Boileau, T.W.; Dowd, S.E.; Fahey, G.C., Jr.; Swanson, K.S. 454 pyrosequencing reveals a shift in fecal microbiota of healthy adult men consuming polydextrose or soluble corn fiber. J. Nutr. 2012, 142, 1259–1265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Dibaise, J.K.; Zuccolo, A.; Kudma, D.; Braidotti, M.; Yu, Y.; Paramerwaran, P.; Crowell, M.D.; Wing, R.; Rittmann, B.E.; et al. Human gut microbiota in obesity and after gastric bypass. Proc. Natl. Acad. Sci. USA 2009, 106, 2365–2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, A.; Pinto, S.C.; Oliveira, R.S. Animais de Laboratório: Criação e Experimentação, 20th ed.; FIOCRUZ: Rio de Janeiro, Brazil, 2002; pp. 350–388. ISBN 85-7541-015-6. [Google Scholar]
- Schipper, L.; Harvey, L.; van der Beek, E.M.; van Dijk, G. Home alone: A systematic review and meta-analysis on the effects of individual housing on body weight, food intake and visceral fat mass in rodents. Obes. Rev. 2018, 19, 614–637. [Google Scholar] [CrossRef] [PubMed]
- Auerbach, M.H.; Craig, S.A.S.; Howllet, J.F.; Hayes, K.C. Caloric Availability of Polydextrose. Nutr. Rev. 2007, 65, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Tiihonen, K.K.; Roÿotiö, H.; Putaala, H.; Ouwehand, A.C. Polydextrose functional fibre Improving digestive health, satiety and beyond. Nutrafoods 2011, 10, 23–28. [Google Scholar] [CrossRef]
- Freitas, K.C.; Amancio, O.M.; de Morais, M.B. High-performance inulin and oligofructose prebiotics increase the intestinal absorption of iron in rats with iron deficiency anaemia during the growth phase. Br. J. Nutr. 2012, 108, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Costabile, A.; Fava, F.; Roÿotiö, H.; Forssten, S.D.; Olli, K.; Klievink, J.; Rowland, I.R.; Ouwehand, A.C.; Rastall, R.A.; Gibson, G.R.; et al. Impact of polydextrose on the faecal microbiota: A double-blind, crossover, placebo-controlled feeding study in healthy human subjects. Br. J. Nutr. 2012, 108, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Forssten, S.D.; Roÿotiö, H.; Hibberd, A.A.; Ouwehand, A.C. The effect of polydextrose and probiotic lactobacilli in a Clostridium difficile-infected human colonic model. Microb. Ecol. Health Dis. 2015, 26, 27988. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, S.; Yde, C.C.; Forssten, S.; Ouwehand, A.C.; Saarinen, M.; Jensen, H.M.; Gibson, G.R.; Rastall, R.; Fava, F.; Bertram, H.C. Impact of dietary polydextrose fiber on the human gut metabolome. J. Agric. Food Chem. 2014, 62, 9944–9951. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, M.; Shimomura, Y.; Suzuki, M. Dietary polydextrose affects the large intestine in rats. J. Nutr. 1994, 124, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Fergunson, L.R.; Ramesh, R.C.; Harris, P.J. Changing concepts of dietary fiber: Implications for carcinogenesis. Nutr. Cancer 2001, 39, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Velasquez, O.C.; Seto, R.W.; Choi, J.; Zhou, D.; Breen, F.; Fisher, J.D.; Rombeuau, J.L. Butyrate inhibits deoxycholate-induced increase of colonic mucosal DNA and protein synthesis in vivo. Dis. Colon Rectum 1997, 40, 1368–1375. [Google Scholar] [CrossRef]
- Wong, J.M.W.; Souza, R.; Kendall, C.W.C.; Eman, A.; Jenkins, A.J.A. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Jie, Z.; Bang-Yao, L.; Ming-Jie, X.; Hai-Wei, L.; Zu-Kang, Z.; Ting-Song, W.; Craig, S.A.S. Studies on the effects of polydextrose intake on physiologic functions in Chinese people. Am. J. Clin. Nutr. 2000, 72, 1503–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cançado, R.D.; Chiattone, C.S. Anemia ferropênica no adulto: Causas, diagnóstico e tratamento. Rev. Bras. Hematol. Hemoter. 2010, 32, 240–246. [Google Scholar] [CrossRef]
- Derelanko, M.J. Determination of erythrocyte life span in F-344, Wistar, and Sprague-Dawley rats using a modification of the [3H] diisopropylfluorophosphate ([3H]DFP) method. Fundam. Appl. Toxicol. 1987, 9, 271–276. [Google Scholar] [CrossRef]
- Belcher, E.H.; Harriss, E.B. Studies of red cell life span in the rat. J. Physiol. 1959, 146, 217–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, J.D.; Baynes, R.D.; Skikne, B.S. Iron deficiency and the measurement of iron status. Nutr. Res. Rev. 1992, 5, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, C.M.; Looker, A.C. Laboratory methodologies for indicators of iron status: Strengths, limitations, and analytical challenges. Am. J. Clin. Nutr. 2017, 106, 1606S–1614S. [Google Scholar] [CrossRef] [PubMed]
- Suchdev, P.S.; Williams, A.M.; Mei, Z.; Flores-Ayala, R.; Pasricha, S.R.; Rogers, L.M.; Namaste, S.M. Assessment of iron status in settings of inflammation: Challenges and potential approaches. Am. J. Clin. Nutr. 2017, 106, 1626S–1633S. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.D.C.; Baracat, E.C.E.; Sgarbieri, V.C. Anemia Ferropriva e Anemia de Doença Crônica: Distúrbios do Metabolismo de Ferro. Segur. Aliment. Nutr. 2006, 54–63. [Google Scholar] [CrossRef]
- Lynch, S. Case studies: Iron. Am. J. Clin. Nutr. 2011, 94, 673S–678S. [Google Scholar] [CrossRef] [PubMed]
- Machado, L.P.; Kohayagawa, A.; Yonezawa, L.A.; Saito, A.E. Metabolismo do ferro em equinos atletas. Cienc. Rural 2010, 40, 703–711, ISSN 0103-8478. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, K.; Bjarnason, I.; Laftah, A.H.; Latunde-dada, G.O.; Simpson, R.J.; Mckie, A.T. Expression of iron absorption genes in mouse large intestine. Scand. J. Gastroenterol. 2005, 40, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Johnston, K.L.; Johnson, D.M.; Marks, J.; Srai, S.K.; Debnem, E.S.; Sharp, P.A. Non-haem iron transport in the rat proximal colon. Eur. J. Clin. Investig. 2006, 36, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D.; Coporaso, J.G.; Hooda, S.; Brulc, J.M.; Fahey, G.C., Jr.; Swanson, K.S. Fiber supplementation influences phylogenetic structure and functional capacity of the human intestinal microbiome: Follow-up of a randomized controlled trial. Am. J. Clin. Nutr. 2015, 101, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Lemos, A.R.; Ismael, L.A.S.; Boato, C.C.M.; Borges, M.T.F.; Rondó, P.H. Hepcidin as a biochemical parameter for the assessment of iron deficiency anemia. Rev. Assoc. Med. Bras. 2010, 56, 596–599. [Google Scholar] [CrossRef]
- Oates, P.S. The role of hepcidin and ferroportin in iron absorption. Histol. Histopathol. 2007, 22, 791. [Google Scholar] [CrossRef] [PubMed]
- Mena, N.P.; Esparza, A.; Tapia, V.; Valdés, P.; Nuñez, M.T. Hepcidin inhibits apical iron uptake in intestinal cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Araujo, J.C.R.; Carvalho, J.J.; Serra, H.O. Influence of duodenogastric reflux in the gastric mucosa histological changes of rats infected with Helicobacter pylori. Rev. Col. Bras. Cir. 2016, 43, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Pietrangelo, A. The ferroportin disease. Blood Cells Mol. Dis. 2004, 32, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Pak, M.; Lopez, M.A.; Gabayan, V.; Ganz, T.; Riviera, S. Suppression of hepcidin during anemia requires erythropoietic activity. Blood 2006, 108, 3730–3735. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Control Diet * | Supplemented Diet (PDX) ** |
|---|---|---|
| Cornstarch 2 | 466 | 441 |
| Maltodextrin 3 | 155 | 155 |
| Casein 3,*** | 140 | 140 |
| Sucrose 4 | 100 | 100 |
| Soybean oil 5 | 40 | 40 |
| Cellulose 3 | 50 | 0 |
| Polydextrose 1 | 0 | 75 |
| Mineral mix 3 | 35 | 35 |
| Vitamin mix 3 | 10 | 10 |
| L-cystine 6 | 1.8 | 1.8 |
| Choline bitartrate 7 | 2.5 | 2.5 |
| Tert-butylhydroquinone 7 | 0.008 | 0.008 |
| Chemical Component | Without Polydextrose (AIN-93M Standard) | With Polydextrose (7.5%) | p-Value |
|---|---|---|---|
| Total solids | 90.12 ± 0.26 b | 90.89 ± 0.36 a | 0.0139 |
| Moisture (%) | 9.11 ± 0.36 b | 9.88 ± 0.26 a | 0.0491 |
| Total ash (g/100 g) | 3.03 ± 0.02 b | 2.81 ± 0.05 a | 0.0002 |
| Lipids (g/100 g) | 4.89 ± 0.90 a | 4.89 ± 0.71 a | 0.9975 |
| Proteins (g/100 g) | 9.55 ± 0.24 a | 9.52 ± 0.30 a | 0.8748 |
| Total carbohydrates (g/100 g) * | 73.42 ± 1.36 a | 72.91 ± 0.50 a | 0.5086 |
| Iron (mg/g) | 0.12 ± 0.02 a | 0.12 ± 0.02 a | 0.9625 |
| Caloric value (kcal/100 g) *** | 375.87 ± 3.18 a | 373.69 ± 3.72 a | 0.4076 |
| Variable | Sham Control | Sham PDX | GXT Control | GXT PDX | p-Value |
|---|---|---|---|---|---|
| Dietary intake (g) | 1149.45 ± 5.81 a | 1070.96 ± 6.69 b | 1087.84 ± 5.46 b | 1008.88 ± 5.81 c | <0.0001 |
| Body weight (g) | 148.00 ± 10.56 a | 137.70 ± 6.29 a | 94.2 ± 9.11 b | 116.60 ± 16.25 a,b | 0.0067 |
| Body length (cm) | 24.19 ± 0.62 a | 23.88 ± 069 a | 23.79 ± 0.31 a | 23.93 ± 1.14 a | 0.8091 |
| FEC | 0.13 ± 0.01 a | 0.13 ± 0.01 a | 0.09 ± 0.01 b,c | 0.12 ± 0.02 b | 0.0007 |
| Variable | Sham Control | Sham PDX | GXT Control | GXT PDX | p-Value |
|---|---|---|---|---|---|
| Serum iron (µg/dL) | 160.25 ± 44.62 a | 167.00 ± 24.86 a | 37.78 ± 20.77 b | 51.00 ± 20.67 b | <0.0001 |
| UIBC (µg/dL) | 317.00 ± 50.58 b | 304.00 ± 70.39 b | 618.67 ± 43.64 a | 618.57 ± 63.02 a | <0.0001 |
| TIBC (µg/dL) | 477.25 ± 60.19 b | 479.30 ± 14.90 b | 654.17 ± 52.25 a | 667.38 ± 66.27 a | <0.0001 |
| TSI (%) * | 32.88 ± 9.85 a | 33.70 ± 8.23 a | 4.83 ± 5.24 b | 7.25 ± 5.38 b | <0.0001 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Do Carmo, M.M.R.; Sarmento, U.C.; Cavalheiro, L.F.; Fernandes, A.; Filiú, W.F.d.O.; Gielow, K.D.C.F.; Costa, D.S.; Guercio, A.C.; Do Nascimento, V.A.; Ribeiro, C.F.A.; et al. Intake of Polydextrose Alters Hematology and the Profile of Short Chain Fatty Acids in Partially Gastrectomized Rats. Nutrients 2018, 10, 792. https://doi.org/10.3390/nu10060792
Do Carmo MMR, Sarmento UC, Cavalheiro LF, Fernandes A, Filiú WFdO, Gielow KDCF, Costa DS, Guercio AC, Do Nascimento VA, Ribeiro CFA, et al. Intake of Polydextrose Alters Hematology and the Profile of Short Chain Fatty Acids in Partially Gastrectomized Rats. Nutrients. 2018; 10(6):792. https://doi.org/10.3390/nu10060792
Chicago/Turabian StyleDo Carmo, Mariane Moreira Ramiro, Ulana Chaves Sarmento, Leandro Fontoura Cavalheiro, Anderson Fernandes, Wander Fernando de Oliveira Filiú, Karine De Cássia Freitas Gielow, Deiler Sampaio Costa, Adriana Conceiçon Guercio, Valter Aragão Do Nascimento, Camila Fontoura Acosta Ribeiro, and et al. 2018. "Intake of Polydextrose Alters Hematology and the Profile of Short Chain Fatty Acids in Partially Gastrectomized Rats" Nutrients 10, no. 6: 792. https://doi.org/10.3390/nu10060792
APA StyleDo Carmo, M. M. R., Sarmento, U. C., Cavalheiro, L. F., Fernandes, A., Filiú, W. F. d. O., Gielow, K. D. C. F., Costa, D. S., Guercio, A. C., Do Nascimento, V. A., Ribeiro, C. F. A., De Castro, A. P., Carvalho, C. M. E., Novello, D., Santos-Junior, V. D. A., Morato, P. N., Amaya-Farfan, J., Hiane, P. A., & Dos Santos, E. F. (2018). Intake of Polydextrose Alters Hematology and the Profile of Short Chain Fatty Acids in Partially Gastrectomized Rats. Nutrients, 10(6), 792. https://doi.org/10.3390/nu10060792