Effects of a High-Protein Diet Including Whole Eggs on Muscle Composition and Indices of Cardiometabolic Health and Systemic Inflammation in Older Adults with Overweight or Obesity: A Randomized Controlled Trial

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
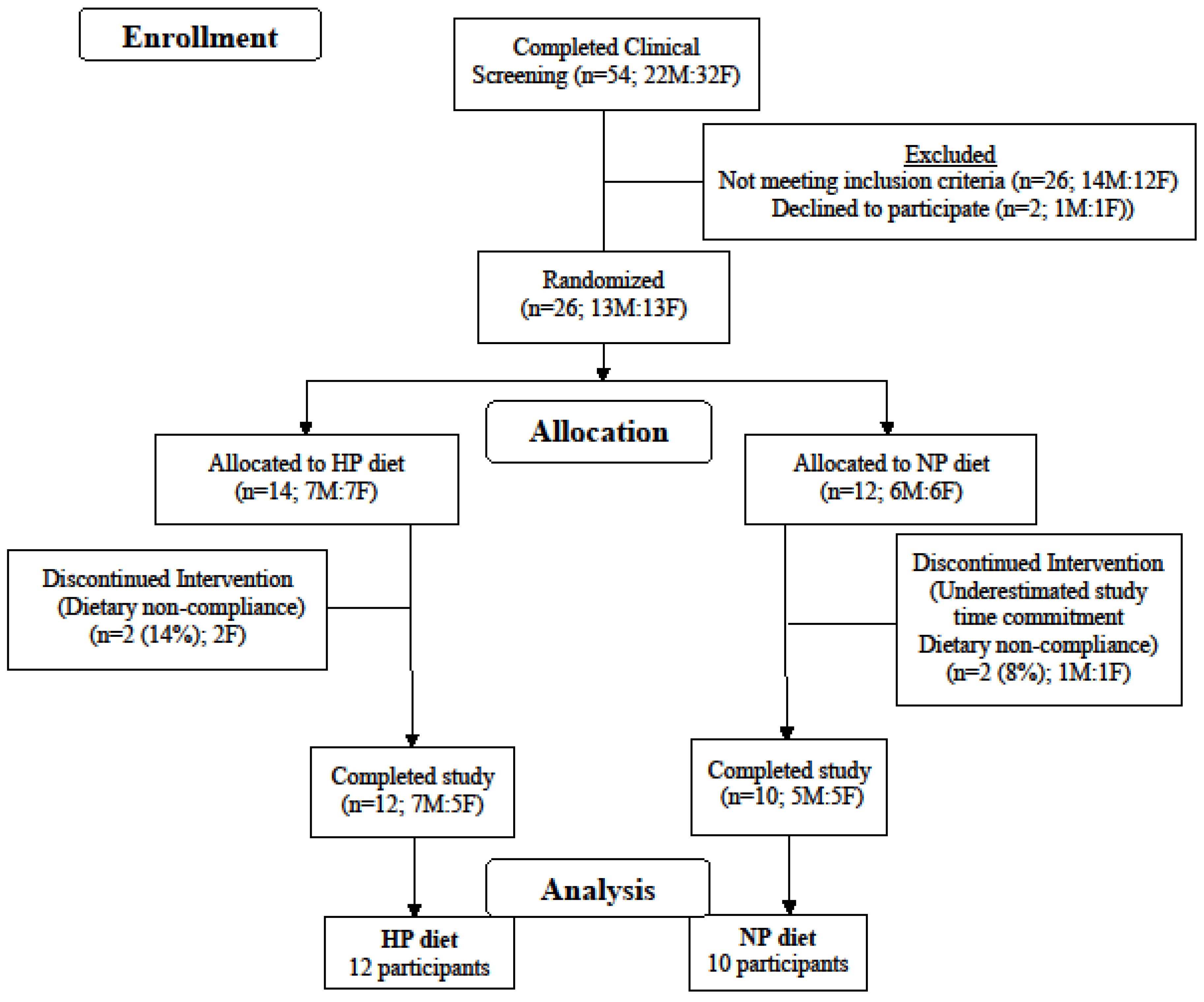
2.1. Participant Characteristics and Design
2.2. Dietary Intervention
2.3. Muscle Composition
2.4. Whole Body Composition
2.5. Clinical Health Assessments
2.6. Blood Analyses
2.7. Sample Size Estimation
2.8. Statistical Analysis
3. Results
3.1. Participant Characteristics
3.2. Muscle Composition
3.3. Whole Body Composition
3.4. Indices of Cardiometabolic Health
3.5. Systemic Inflammation
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Goodpaster, B.H.; Thaete, F.L.; Simoneau, J.A.; Kelley, D.E. Subcutaneous abdominal fat and thigh muscle composition predict insulin sensitivity independently of visceral fat. Diabetes 1997, 46, 1579–1585. [Google Scholar] [CrossRef] [PubMed]
- Goodpaster, B.H.; Thaete, F.L.; Kelley, D.E. Thigh adipose tissue distribution is associated with insulin resistance in obesity and in type 2 diabetes mellitus. Am. J. Clin. Nutr. 2000, 71, 885–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodpaster, B.H.; Krishnaswami, S.; Resnick, H.; Kelley, D.E.; Haggerty, C.; Harris, T.B.; Schwartz, A.V.; Kritchevsky, S.; Newman, A.B. Association between regional adipose tissue distribution and both type 2 diabetes and impaired glucose tolerance in elderly men and women. Diabetes Care 2003, 26, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, D.; Kuznia, P.; Heshka, S.; Albu, J.; Heymsfield, S.B.; Goodpaster, B.; Visser, M.; Harris, T.B. Adipose tissue in muscle: A novel depot similar in size to visceral adipose tissue. Am. J. Clin. Nutr. 2005, 81, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Yim, J.E.; Heshka, S.; Albu, J.; Heymsfield, S.; Kuznia, P.; Harris, T.; Gallagher, D. Intermuscular adipose tissue rivals visceral adipose tissue in independent associations with cardiovascular risk. Int. J. Obesity 2007, 31, 1400–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wronska, A.; Kmiec, Z. Structural and biochemical characteristics of various white adipose tissue depots. Acta. Physiol. 2012, 205, 194–208. [Google Scholar] [CrossRef] [PubMed]
- Beasley, L.E.; Koster, A.; Newman, A.B.; Javaid, M.K.; Ferrucci, L.; Kritchevsky, S.B.; Kuller, L.H.; Pahor, M.; Schaap, L.A.; Visser, M.; et al. Inflammation and race and gender differences in computerized tomography-measured adipose depots. Obesity 2009, 17, 1062–1069. [Google Scholar] [CrossRef] [PubMed]
- Malavazos, A.E.; Corsi, M.M.; Ermetici, F.; Coman, C.; Sardanelli, F.; Rossi, A.; Morricone, L.; Ambrosi, B. Proinflammatory cytokines and cardiac abnormalities in uncomplicated obesity: Relationship with abdominal fat deposition. Nutr. Metab. Cardiovas. 2007, 17, 294–302. [Google Scholar] [CrossRef] [PubMed]
- MohamedAli, V.; Goodrick, S.; Rawesh, A.; Yudkin, J.; Coppack, S. Subcutaneous adipose tissue releases interleukin-6 but not tumour necrosis factor-alpha in vivo. Diabetes 1997, 46, 215. [Google Scholar]
- Ryan, A.S.; Nicklas, B.J. Age-related changes in fat deposition in mid-thigh muscle in women: Relationships with metabolic cardiovascular disease risk factors. Int. J. Obesity 1999, 23, 126–132. [Google Scholar] [CrossRef]
- Yim, J.E.; Heshka, S.; Albu, J.B.; Heymsfield, S.; Gallagher, D. Femoral-gluteal subcutaneous and intermuscular adipose tissues have independent and opposing relationships with CVD risk. J. Appl. Physiol. 2008, 104, 700–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koster, A.; Stenholm, S.; Alley, D.E.; Kim, L.J.; Simonsick, E.M.; Kanaya, A.M.; Visser, M.; Houston, D.K.; Nicklas, B.J.; Tylavsky, F.A.; et al. Body fat distribution and inflammation among obese older adults with and without metabolic syndrome. Obesity 2010, 18, 2354–2361. [Google Scholar] [CrossRef] [PubMed]
- Dube, M.C.; Lemieux, S.; Piche, M.E.; Corneau, L.; Bergeron, J.; Riou, M.E.; Weisnagel, S.J. The contribution of visceral adiposity and mid-thigh fat-rich muscle to the metabolic profile in postmenopausal women. Obesity 2011, 19, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Durheim, M.T.; Slentz, C.A.; Bateman, L.A.; Mabe, S.K.; Kraus, W.E. Relationships between exercise-induced reductions in thigh intermuscular adipose tissue, changes in lipoprotein particle size, and visceral adiposity. Am. J. Physiol-Endoc. Metab. 2008, 295, E407–E412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz-Peiffer, C. Signalling aspects of insulin resistance in skeletal muscle: Mechanisms induced by lipid oversupply. Cell Signal. 2000, 12, 583–594. [Google Scholar] [CrossRef]
- Kelley, D.E.; Goodpaster, B.H. Skeletal muscle triglyceride—An aspect of regional adiposity and insulin resistance. Diabetes Care 2001, 24, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Miljkovic-Gacic, I.; Gordon, C.L.; Goodpaster, B.H.; Bunker, C.H.; Patrick, A.L.; Kuller, L.H.; Wheeler, V.W.; Evans, R.W.; Zmuda, J.M. Adipose tissue infiltration in skeletal muscle: Age patterns and association with diabetes among men of african ancestry. Am. J. Clin. Nutr. 2008, 87, 1590–1595. [Google Scholar] [CrossRef] [PubMed]
- Zoico, E.; Rossi, A.; Di Francesco, V.; Sepe, A.; Olioso, D.; Pizzini, F.; Fantin, F.; Bosello, O.; Cominacini, L.; Harris, T.B.; et al. Adipose tissue infiltration in skeletal muscle of healthy elderly men: Relationships with body composition, insulin resistance, and inflammation at the systemic and tissue level. J. Gerontol. A Biol. Sci. Med. Sci. 2009, 65, 265–299. [Google Scholar] [CrossRef] [PubMed]
- Delmonico, M.J.; Harris, T.B.; Visser, M.; Park, S.W.; Conroy, M.B.; Velasquez-Mieyer, P.; Boudreau, R.; Manini, T.M.; Nevitt, M.; Newman, A.B.; et al. Longitudinal study of muscle strength, quality, and adipose tissue infiltration. Am. J. Clin. Nutr. 2009, 90, 1579–1585. [Google Scholar] [PubMed] [Green Version]
- Goodpaster, B.H.; Carlson, C.L.; Visser, M.; Kelley, D.E.; Scherzinger, A.; Harris, T.B.; Stamm, E.; Newman, A.B. Attenuation of skeletal muscle and strength in the elderly: The health abc study. J. Appl. Physiol. (1985) 2001, 90, 2157–2165. [Google Scholar] [CrossRef] [PubMed]
- Goodpaster, B.H.; Chomentowski, P.; Ward, B.K.; Rossi, A.; Glynn, N.W.; Delmonico, M.J.; Kritchevsky, S.B.; Pahor, M.; Newman, A.B. Effects of physical activity on strength and skeletal muscle fat infiltration in older adults: A randomized controlled trial. J. Appl. Physiol. 2008, 105, 1498–1503. [Google Scholar] [CrossRef] [PubMed]
- Hilton, T.N.; Tuttle, L.J.; Bohnert, K.L.; Mueller, M.J.; Sinacore, D.R. Excessive adipose tissue infiltration in skeletal muscle in individuals with obesity, diabetes mellitus, and peripheral neuropathy: Association with performance and function. Phys. Ther. 2008, 88, 1336–1344. [Google Scholar] [CrossRef] [PubMed]
- Manini, T.M.; Clark, B.C.; Nalls, M.A.; Goodpaster, B.H.; Ploutz-Snyder, L.L.; Harris, T.B. Reduced physical activity increases intermuscular adipose tissue in healthy young adults. Am. J. Clin. Nutr. 2007, 85, 377–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, Y.; Marcus, R.L.; Lastayo, P.C. Intramuscular adipose tissue and central activation in older adults. Muscle Nerve 2012, 46, 813–816. [Google Scholar] [CrossRef] [PubMed]
- Goodpaster, B.H.; Park, S.W.; Harris, T.B.; Kritchevsky, S.B.; Nevitt, M.; Schwartz, A.V.; Simonsick, E.M.; Tylavsky, F.A.; Visser, M.; Newman, A.B. The loss of skeletal muscle strength, mass, and quality in older adults: The health, aging and body composition study. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2006, 61, 1059–1064. [Google Scholar] [CrossRef]
- Visser, M.; Goodpaster, B.H.; Kritchevsky, S.B.; Newman, A.B.; Nevitt, M.; Rubin, S.M.; Simonsick, E.M.; Harris, T.B.; Study, H.A. Muscle mass, muscle strength, and muscle fat infiltration as predictors of incident mobility limitations in well-functioning older persons. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2005, 60, 324–333. [Google Scholar] [CrossRef]
- Visser, M.; Kritchevsky, S.B.; Goodpaster, B.H.; Newman, A.B.; Nevitt, M.; Stamm, E.; Harris, T.B. Leg muscle mass and composition in relation to lower extremity performance in men and women aged 70 to 79: The health, aging and body composition study. J. Am. Geriatr. Soc. 2002, 50, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, L.J.S.; Sinacore, D.R.; Mueller, M.J. Intermuscular adipose tissue is muscle specific and associated with poor functional performance. J. Aging Res. 2012, 2012, 172957. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, T.; Paulsen, S.K.; Bruun, J.M.; Pedersen, S.B.; Richelsen, B. Exercise training versus diet-induced weight-loss on metabolic risk factors and inflammatory markers in obese subjects: A 12-week randomized intervention study. Am. J. Physiol. Endocrinol. Metabol. 2010, 298, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Goodpaster, B.H.; Kelley, D.E.; Wing, R.R.; Meier, A.; Thaete, F.L. Effects of weight loss on regional fat distribution and insulin sensitivity in obesity. Diabetes 1999, 48, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Janssen, I.; Hudson, R.; Fortier, A.; Ross, R. Effects of an energy-restrictive diet with or without exercise on abdominal fat, intermuscular fat, and metabolic risk factors in obese women. Diabetes Care 2002, 25, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Mazzali, G.; Di Francesco, V.; Zoico, E.; Fantin, F.; Zamboni, G.; Benati, C.; Bambara, V.; Negri, M.; Bosello, O.; Zamboni, M. Interrelations between fat distribution, muscle lipid content, adipocytokines, and insulin resistance: Effect of moderate weight loss in older women. Am. J. Clin. Nutr. 2006, 84, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.S.; Ortmeyer, H.K.; Sorkin, J.D. Exercise with calorie restriction improves insulin sensitivity and glycogen synthase activity in obese postmenopausal women with impaired glucose tolerance. Am. J. Physiol.-Endoc. Metab. 2012, 302, E145–E152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addison, O.; Marcus, R.L.; Lastayo, P.C.; Ryan, A.S. Intermuscular fat: A review of the consequences and causes. Int. J. Endocrinol. 2014, 2014, 309570. [Google Scholar] [CrossRef] [PubMed]
- Gregorio, L.; Brindisi, J.; Kleppinger, A.; Sullivan, R.; Mangano, K.M.; Bihuniak, J.D.; Kenny, A.M.; Kerstetter, J.E.; Insogna, K.L. Adequate dietary protein is associated with better physical performance among post-menopausal women 60–90 years. J. Nutr. Health Aging 2014, 18, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Houston, D.K.; Nicklas, B.J.; Ding, J.Z.; Harris, T.B.; Tylavsky, F.A.; Newman, A.B.; Lee, J.S.; Sahyoun, N.R.; Visser, M.; Kritchevsky, S.B.; et al. Dietary protein intake is associated with lean mass change in older, community-dwelling adults: The health, aging, and body composition (Health ABC) study. Am. J. Clin. Nutr. 2008, 87, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Iglay, H.B.; Thyfault, J.P.; Apolzan, J.W.; Campbell, W.W. Resistance training and dietary protein: Effects on glucose tolerance and contents of skeletal muscle insulin signaling proteins in older persons. Am. J. Clin. Nutr. 2007, 85, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Solerte, S.B.; Gazzaruso, C.; Bonacasa, R.; Rondanelli, M.; Zamboni, M.; Basso, C.; Locatelli, E.; Schifino, N.; Giustina, A.; Fioravanti, M. Nutritional supplements with oral amino acid mixtures increases whole-body lean mass and insulin sensitivity in elderly subjects with sarcopenia. Am. J. Cardiol. 2008, 101, 69E–77E. [Google Scholar] [CrossRef] [PubMed]
- Ratliff, J.C.; Mutungi, G.; Puglisi, M.J.; Volek, J.S.; Fernandez, M.L. Eggs modulate the inflammatory response to carbohydrate restricted diets in overweight men. Nutr. Metab. 2008, 5, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westphal, S.; Kastner, S.; Taneva, E.; Leodolter, A.; Dierkes, J.; Luley, C. Postprandial lipid and carbohydrate responses after the ingestion of a casein-enriched mixed meal. Am. J. Clin. Nutr. 2004, 80, 284–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortensen, L.S.; Hartvigsen, M.L.; Brader, L.J.; Astrup, A.; Schrezenmeir, J.; Holst, J.J.; Thomsen, C.; Hermansen, K. Differential effects of protein quality on postprandial lipemia in response to a fat-rich meal in type 2 diabetes: Comparison of whey, casein, gluten, and cod protein. Am. J. Clin. Nutr. 2009, 90, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.R.; Robinson, M.J.; Fry, J.L.; Tang, J.E.; Glover, E.I.; Wilkinson, S.B.; Prior, T.; Tarnopolsky, M.A.; Phillips, S.M. Ingested protein dose response of muscle and albumin protein synthesis after resistance exercise in young men. Am. J. Clin. Nutri. 2009, 89, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.C.; Newcomer, B.R.; Buchthal, S.D.; Sirikul, B.; Oster, R.A.; Hunter, G.R.; Gower, B.A. Relationship of intramyocellular lipid to insulin sensitivity may differ with ethnicity in healthy girls and women. Obesity (Silver Spring) 2011, 19, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Haffner, S.M. Insulin resistance, inflammation, and the prediabetic state. Am. J. Cardiol. 2003, 92, 18J–26J. [Google Scholar] [CrossRef]
- Ouchi, N.; Kihara, S.; Funahashi, T.; Nakamura, T.; Nishida, M.; Kumada, M.; Okamoto, Y.; Ohashi, K.; Nagaretani, H.; Kishida, K.; et al. Reciprocal association of c-reactive protein with adiponectin in blood stream and adipose tissue. Circulation 2003, 107, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Benedict, H.A. A Biometric Study of Basal Metabolism in Man; Carnegie Institute of Washington: Washington, DC, USA, 1919. [Google Scholar]
- Song, M.Y.; Ruts, E.; Kim, J.; Janumala, I.; Heymsfield, S.; Gallagher, D. Sarcopenia and increased adipose tissue infiltration of muscle in elderly african american women. Am. J. Clin. Nutr. 2004, 79, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Berg, H.E.; Tedner, B.; Tesch, P.A. Changes in lower limb muscle cross-sectional area and tissue fluid volume after transition from standing to supine. Acta. Physiol. Scand. 1993, 148, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Schrauwen-Hinderling, V.B.; Kooi, M.E.; Hesselink, M.K.; Moonen-Kornips, E.; Schaart, G.; Mustard, K.J.; Hardie, D.G.; Saris, W.H.; Nicolay, K.; Schrauwen, P. Intramyocellular lipid content and molecular adaptations in response to a 1-week high-fat diet. Obes. Res. 2005, 13, 2088–2094. [Google Scholar] [CrossRef] [PubMed]
- Borsheim, E.; Bui, Q.U.; Tissier, S.; Cree, M.G.; Ronsen, O.; Morio, B.; Ferrando, A.A.; Kobayashi, H.; Newcomer, B.R.; Wolfe, R.R. Amino acid supplementation decreases plasma and liver triacylglycerols in elderly. Nutrition 2009, 25, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoaglin, D.C.; Iglewicz, B.; Tukey, J.W. Performance of some resistant rules for outlier labeling. J. Am. Stat. Assoc. 1986, 81, 991–999. [Google Scholar] [CrossRef]
- Strasser, B.; Volaklis, K.; Fuchs, D.; Burtscher, M. Role of dietary protein and muscular fitness on longevity and aging. Aging Dis. 2018, 9, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, R.; Sakai, A.; Murayama, M.; Ochi, A.; Abe, T.; Hirasaka, K.; Ohno, A.; Teshima-Kondo, S.; Yanagawa, H.; Yasui, N.; et al. Effects of dietary soy protein on skeletal muscle volume and strength in humans with various physical activities. J. Med. Investig. 2015, 62, 177–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, J.E. Diet, exercise or diet with exercise: Comparing the effectiveness of treatment options for weight-loss and changes in fitness for adults (18–65 years old) who are overfat, or obese; systematic review and meta–analysis. J. Diabetes Metab. Disord. 2015, 14, 31. [Google Scholar] [CrossRef] [PubMed]
- Dhurandhar, E.J.; Kaiser, K.A.; Dawson, J.A.; Alcorn, A.S.; Keating, K.D.; Allison, D.B. Predicting adult weight change in the real world: A systematic review and meta-analysis accounting for compensatory changes in energy intake or expenditure. Int. J. Obes. 2015, 39, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Raynor, H.A.; Champagne, C.M. Position of the academy of nutrition and dietetics: Interventions for the treatment of overweight and obesity in adults. J. Acad. Nutr. Diet. 2016, 116, 129–147. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; O’Connor, L.E.; Sands, L.P.; Slebodnik, M.B.; Campbell, W.W. Effects of dietary protein intake on body composition changes after weight loss in older adults: A systematic review and meta-analysis. Nutr. Rev. 2016, 74, 210–224. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.J.; Campbell, W.W. Sarcopenia and age-related changes in body composition and functional capacity. J. Nutr. 1993, 123, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Rantanen, T.; Avlund, K.; Suominen, H.; Schroll, M.; Frandin, K.; Pertti, E. Muscle strength as a predictor of onset of adl dependence in people aged 75 years. Aging Clin. Exp. Res. 2002, 14, 10–15. [Google Scholar] [PubMed]
- Rantanen, T.; Harris, T.; Leveille, S.G.; Visser, M.; Foley, D.; Masaki, K.; Guralnik, J.M. Muscle strength and body mass index as long-term predictors of mortality in initially healthy men. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2000, 55, M168–M173. [Google Scholar] [CrossRef]
- Katz, D.L.; Gnanaraj, J.; Treu, J.A.; Ma, Y.; Kavak, Y.; Njike, V.Y. Effects of egg ingestion on endothelial function in adults with coronary artery disease: A randomized, controlled, crossover trial. Am. Heart J. 2015, 169, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Fuller, N.R.; Caterson, I.D.; Sainsbury, A.; Denyer, G.; Fong, M.; Gerofi, J.; Baqleh, K.; Williams, K.H.; Lau, N.S.; Markovic, T.P. The effect of a high-egg diet on cardiovascular risk factors in people with type 2 diabetes: The diabetes and egg (diabegg) study-a 3-mo randomized controlled trial. Am. J. Clin. Nutr. 2015, 101, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, S.; Gardener, H.; Tiozzo, E.; Ying Kuen, C.; Elkind, M.S.; Sacco, R.L.; Rundek, T. Egg consumption and carotid atherosclerosis in the northern manhattan study. Atherosclerosis 2014, 235, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Arad, Y.; Goodman, K.J.; Roth, M.; Newstein, D.; Guerci, A.D. Coronary calcification, coronary disease risk factors, c-reactive protein, and atherosclerotic cardiovascular disease events—The St. Francis heart study. J. Am. Coll. Cardiol. 2005, 46, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Superko, H.R.; King, S. Lipid management to reduce cardiovascular risk—A new strategy is required. Circulation 2008, 117, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Superko, H.R. Beyond LDL cholesterol reduction. Circulation 1996, 94, 2351–2354. [Google Scholar] [CrossRef] [PubMed]
- Millan, J.; Pinto, X.; Munoz, A.; Zuniga, M.; Rubies-Prat, J.; Pallardo, L.F.; Masana, L.; Mangas, A.; Hernandez-Mijares, A.; Gonzalez-Santos, P.; et al. Lipoprotein ratios: Physiological significance and clinical usefulness in cardiovascular prevention. Vasc. Health Risk Manag. 2009, 5, 757–765. [Google Scholar] [PubMed]
- Budinger, T.F.; Fischer, H.; Hentschel, D.; Reinfelder, H.E.; Schmitt, F. Physiological effects of fast oscillating magnetic field gradients. J. Comput. Assist. Tomogr. 1991, 15, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Ham, C.L.; Engels, J.M.; van de Wiel, G.T.; Machielsen, A. Peripheral nerve stimulation during MRI: Effects of high gradient amplitudes and switching rates. J. Magn. Reson. Imaging 1997, 7, 933–937. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.D.; Noseworthy, M.D. Motion and distortion correction of skeletal muscle echo planar images. Magn. Reson. Imaging 2016, 34, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Roujol, S.; Basha, T.A.; Weingartner, S.; Akcakaya, M.; Berg, S.; Manning, W.J.; Nezafat, R. Impact of motion correction on reproducibility and spatial variability of quantitative myocardial t2 mapping. J. Cardiovasc. Magn. Reson. 2015, 17, 46. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Total | Egg | Meat | Plant | Grain | Dairy | Residual | |
|---|---|---|---|---|---|---|---|
| NP | |||||||
| Breakfast, g | 12.7 | 0.0 | 3.0 | 0.6 | 7.0 | 1.1 | 0.9 |
| Lunch, g | 12.6 | 0.0 | 2.7 | 0.6 | 4.5 | 2.8 | 1.9 |
| Dinner, g | 31.7 | 0.0 | 15.0 | 1.6 | 6.5 | 6.2 | 2.4 |
| Snack, g | 12.5 | 0.0 | 0.0 | 3.5 | 0.0 | 7.7 | 1.3 |
| Total Intake, g | 69.5 | 0.0 | 20.7 | 6.3 | 18.1 | 17.8 | 6.5 |
| Percentage, % | 100% | 0% | 30% | 9% | 26% | 26% | 9% |
| HP | |||||||
| Breakfast, g | 30.0 | 12.5 | 6.2 | 1.3 | 7.6 | 1.8 | 0.6 |
| Lunch, g | 31.2 | 6.3 | 8.1 | 2.3 | 5.6 | 8.6 | 0.3 |
| Dinner, g | 32.4 | 0.0 | 16.2 | 1.5 | 4.6 | 7.9 | 2.2 |
| Snack, g | 25.8 | 10.6 | 0.0 | 2.2 | 0.0 | 12.5 | 0.5 |
| Total Intake, g | 119.4 | 29.4 | 30.5 | 7.4 | 17.8 | 30.8 | 3.5 |
| Percentage, % | 100% | 25% | 26% | 6% | 15% | 26% | 3% |
| Added Protein (HP-NP) | |||||||
| Breakfast, g | 17.3 | 12.5 | 3.2 | 0.7 | 0.6 | 0.7 | −0.3 |
| Lunch, g | 18.6 | 6.3 | 5.4 | 1.7 | 1.1 | 5.8 | −1.6 |
| Dinner, g | 0.7 | 0.0 | 1.2 | −0.1 | −1.9 | 1.7 | −0.2 |
| Snack, g | 13.3 | 10.6 | 0.0 | −1.3 | 0.0 | 4.8 | −0.8 |
| Total Intake, g | 49.9 | 29.4 | 9.8 | 1.1 | −0.3 | 13.0 | −3.0 |
| Percentage, % | 100% | 59% | 20% | 2% | −1% | 26% | −6% |
| Variable | All | HP | NP |
|---|---|---|---|
| n | 22 | 12 | 10 |
| Age, years | 70 ± 5 | 70 ± 6 | 71 ± 3 |
| Male:Female | 12:10 | 7:5 | 5:5 |
| Body mass, kg | 92.1 ± 15.4 | 90.1 ± 16.4 | 91.0 ± 16.1 |
| Height, cm | 171 ± 11 | 167 ± 12 | 172 ± 9 |
| BMI, kg/m 2 | 31.3 ± 3.2 | 32.2 ± 3.4 | 30.5 ± 3.3 |
| Waist, cm | 110 ± 9 | 110 ± 11 | 110 ± 9 |
| Hip, cm | 115 ± 9 | 114 ± 11 | 115 ± 9 |
| Lean mass, kg | 51.3 ± 10.9 | 49.0 ± 11.6 | 50.7 ± 10.3 |
| Fat mass, kg | 37.2 ± 8.4 | 37.6 ± 9.2 | 36.9 ± 8.9 |
| Percentage body fat | 42.1 ± 7.3 | 43.5 ± 8.3 | 42.0 ± 6.7 |
| Fasting glucose, mg/dL | 99.2 ± 9.7 | 98.1 ± 11.1 | 98.3 ± 10.0 |
| Fasting insulin, µU/mL | 13.1 ± 7.5 | 11.4 ± 7.1 | 13.1 ± 7.2 |
| HOMA-IR | 3.3 ± 2.1 | 2.8 ± 2.1 | 3.3 ± 2.0 |
| Total cholesterol, mg/dL | 190.4 ± 37.0 | 201.4 ± 33.5 | 185.5 ± 26.7 |
| LDL cholesterol, mg/dL | 115.7 ± 32.4 | 123.5 ± 38.0 | 111.5 ± 18.2 |
| HDL cholesterol, mg/dL | 48.7 ± 14.6 | 54.0 ± 11.2 | 47.6 ± 18.1 |
| Total Cholesterol:HDL | 4.2 ± 1.2 | 3.9 ± 1.1 | 4.3 ± 1.5 |
| Triglycerides, mg/dL | 130.1 ± 58.6 | 119.9 ± 64.0 | 131.6 ± 64.7 |
| Manual SBP 3, mm Hg | 132 ± 14 | 129 ± 12 | 134 ± 15 |
| Manual DBP 3, mm Hg | 84 ± 7 | 81 ± 7 | 86 ± 8 |
| BUN, mg/dL | 16.5 ± 3.3 | 17.0 ± 4.6 | 16.2 ± 2.6 |
| Total Energy, kcal/day | 2011 ± 537 | 2061 ± 597 | 1957 ± 597 |
| Total Fat, g/day | 79 ± 31 | 82 ± 35 | 75 ± 35 |
| Total Fat, % | 34 ± 7 | 34 ± 6 | 33 ± 6 |
| Total Carbohydrates, g/day | 245 ± 69 | 249 ± 71 | 241 ± 71 |
| Total Carbohydrates, % | 48 ± 8 | 48 ± 6 | 48 ± 6 |
| Total Protein, g/day | 82 ± 24 | 84 ± 15 | 79 ± 15 |
| Total Protein, % | 17 ± 5 | 16 ± 4 | 17 ± 4 |
| Animal Protein, g/day | 54 ± 25 | 55 ± 12 | 53 ± 12 |
| Vegetable Protein, g/day | 28 ± 8 | 29 ± 6 | 27 ± 6 |
| Baseline | Post | ∆ | ||
|---|---|---|---|---|
| Thigh | (n = 18) | (n = 19) | (n = 18) | |
| MCSA, mm2 × 104 | NP | 258.4 ± 54.0 | 218.9 ± 47.0 | −29.2 ± 65.1 |
| HP | 260.2 ± 69.4 | 254.4 ± 98.50 | 9.3 ± 114.0 | |
| All | 259.3 ± 60.3 | 237.6 ± 78.5 | −8.8 ± 93.5 | |
| SubQ, mm2 × 104 | NP | 104.8 ± 49.0 | 68.3 ± 36.4 | −28.6 ± 33.8 |
| HP | 82.1 ± 42.0 | 82.4 ± 35.9 | 1.8 ± 53.8 | |
| All | 93.5 ± 45.7 | 75.7 ± 45.7 | −12.5 ± 46.8 | |
| IMAT, mm3 × 104 | NP | 16.7 ± 11.3 | 12.5 ± 4.4 | −4.9 ± 10.2 |
| HP | 17.9 ± 7.8 | 15.1 ± 7.4 | −2.0 ± 6.1 | |
| All | 17.3 ± 9.4 | 13.9 ± 6.1 | −3.3 ± 8.2 | |
| Muscle Volume, mm3 × 104 | NP | 119.5 ± 38.7 | 110.5 ± 45.3 | −7.4 ± 28.3 |
| HP | 151.4 ± 68.2 | 125.5 ± 61.1 | −16.2 ± 60.5 | |
| All | 135.4 ± 56.3 | 118.4 ± 53.2 | −12.1 ± 46.9 | |
| Total Fat:CSA | NP | 0.47 ± 0.16 | 0.38 ± 0.16 | −0.07 ± 0.06 |
| HP | 0.40 ± 0.18 | 0.40 ± 0.15 | −0.02 ± 0.10 | |
| All | 0.43 ± 0.17 | 0.39 ± 0.15 | −0.04 ± 0.09 | |
| IMAT:Muscle Volume | NP | 0.12 ± 0.04 | 0.12 ± 0.03 | −0.03 ± 0.06 |
| HP | 0.11 ± 0.03 | 0.13 ± 0.05 | −0.01 ± 0.01 | |
| All | 0.12 ± 0.03 | 0.13 ± 0.04 | −0.02 ± 0.04 | |
| SubQ:Muscle Volume | NP | 1.03 ± 0.64 | 0.82 ± 0.72 | −0.10 ± 0.28 |
| HP | 0.78 ± 0.73 | 0.87 ± 0.64 | −0.01 ± 0.31 | |
| All | 0.90 ± 0.68 | 0.85 ± 0.66 | −0.05 ± 0.29 | |
| Calf | (n = 15) | (n = 17) | (n = 15) | |
| MCSA, mm2 × 104 | NP | 134.9 ± 28.8 | 143.6 ± 41.6 | 2.0 ± 15.5 |
| HP | 142.0 ± 30.7 | 141.0 ± 27.2 | −1.9 ± 4.8 | |
| All | 138.6 ± 29.0 | 142.2 ± 33.6 | 0.2 ± 11.6 | |
| SubQ, mm2 × 104 | NP | 48.3 ± 41.6 | 49.8 ± 25.2 | 0.6 ± 2.7 |
| HP | 48.8 ± 33.7 | 49.8 ± 31.9 | −0.9 ± 2.0 | |
| All | 48.6 ± 30.3 | 49.8 ± 28.0 | −0.1 ± 2.4 | |
| IMAT, mm3 × 104 | NP | 9.3 ± 3.3 | 9.9 ± 4.6 | −0.6 ± 1.3 |
| HP | 10.0 ± 3.6 | 10.0 ± 3.6 | −1.1 ± 1.0 | |
| All | 9.7 ± 3.4 | 9.9 ± 3.3 | −0.9 ± 1.1 | |
| Muscle Volume, mm3 × 104 | NP | 77.6 ± 21.7 | 80.5 ± 22.3 | 1.2 ± 6.9 |
| HP | 79.6 ± 18.3 | 79.2 ± 18.3 | −0.5 ± 3.0 | |
| All | 78.6 ± 19.3 | 79.8 ± 20.1 | −0.2 ± 5.3 | |
| Total Fat: CSA | NP | 0.44 ± 0.23 | 0.44 ± 0.22 | 0.01 ± 0.06 |
| HP | 0.42 ± 0.25 | 0.43 ± 0.24 | −0.01 ± 0.01 | |
| All | 0.43 ± 0.23 | 0.43 ± 0.22 | −0.004 ± 0.05 | |
| IMAT:Muscle Volume | NP | 0.12 ± 0.04 | 0.12 ± 0.05 | −0.01 ± 0.02 |
| HP | 0.12 ± 0.03 | 0.13 ± 0.03 | −0.01 ± 0.01 | |
| All | 0.12 ± 0.03 | 0.13 ± 0.04 | −0.01 ± 0.01 | |
| SubQ:Muscle Volume | NP | 0.67 ± 0.39 | 0.66± 0.36 | 0.01 ± 0.02 A |
| HP | 0.64 ± 0.43 | 0.68 ± 0.44 | −0.01 ± 0.02 A,B | |
| All | 0.66 ± 0.40 | 0.67 ± 0.40 | −0.002 ± 0.02 | |
| Baseline | Post | ∆ | |||
|---|---|---|---|---|---|
| (n = 22) | (n = 22) | (n = 22) | |||
| Appendicular fat mass, kg | NP | 15.72 ± 4.23 | 14.68 ± 4.05 | −1.04 ± 0.76 A | |
| HP | 14.84 ± 4.99 | 14.41 ± 4.94 | −0.43 ± 0.52 A | ||
| All | 15.24 ± 4.58 | 14.53 ± 4.45 | −0.71 ± 0.70 | ||
| Appendicular lean mass, kg | NP | 23.34 ± 5.49 | 22.76 ± 5.56 | −0.58 ± 0.66 | |
| HP | 23.61 ± 5.93 | 23.22 ± 5.90 | −0.39 ± 0.71 | ||
| All | 23.49 ± 5.60 | 23.01 ± 5.62 | −0.48 ± 0.68 | ||
| Trunk fat mass, kg | NP | 20.13 ± 6.27 | 18.51 ± 6.32 | −1.62 ± 1.21 | |
| HP | 21.64 ± 4.89 | 20.18 ± 5.05 | −1.46 ± 1.16 | ||
| All | 20.95 ± 5.47 | 19.42 ± 5.59 | −1.53 ± 1.16 | ||
| Trunk lean mass, kg | NP | 23.98 ± 4.67 | 23.30 ± 4.58 | −0.68 ± 0.72 A | |
| HP | 24.69 ± 5.49 | 24.81 ± 5.56 | 0.12 ± 0.63 A | ||
| All | 24.37 ± 5.03 | 24.13 ± 5.08 | −0.24 ± 0.77 | ||
| Percent body fat | NP | 41.99 ± 6.65 | 40.75 ± 6.77 | −1.24 ± 1.26 | |
| HP | 42.20 ± 8.14 | 41.00 ± 8.26 | −1.20 ± 1.04 | ||
| All | 42.11 ± 7.33 | 40.88 ± 7.44 | −1.22 ± 1.11 | ||
| Total fat mass, kg | NP | 36.83 ± 8.85 | 34.15 ± 8.98 | −2.68 ± 1.81 | |
| HP | 37.50 ± 8.32 | 35.60 ± 8.65 | −1.90 ± 1.52 | ||
| All | 37.20 ± 8.37 | 34.94 ± 8.62 | −2.25 ± 1.67 B | ||
| Total lean mass, kg | NP | 50.71 ± 10.33 | 49.42 ± 10.37 | −1.29 ± 0.97 A,B | |
| HP | 51.80 ± 11.77 | 51.52 ± 11.76 | −0.28 ± 1.20 A | ||
| All | 51.30 ± 10.89 | 50.56 ± 10.94 | −0.74 ± 1.19 | ||
| Total mass, kg | NP | 90.5 ± 15.8 | 86.49 ± 16.23 | −3.96 ± 2.12 | |
| HP | 92.1 ± 15.4 | 89.89 ± 16.07 | −2.20 ± 2.46 | ||
| All | 91.3 ± 15.2 | 88.3 ± 15.8 | −3.00 ± 2.43 B | ||
| Baseline | Post | ∆ | |||
|---|---|---|---|---|---|
| (n = 22) | (n = 22) | (n = 22) | |||
| Waist, cm | NP | 110 ± 9 | 102 ± 13 | −8 ± 9 | |
| HP | 110 ± 11 | 107 ± 13 | −3 ± 8 | ||
| All | 110 ± 9 | 105 ± 13 | −5 ± 9 B | ||
| (n = 22) | (n = 22) | (n = 22) | |||
| Hip, cm | NP | 115 ± 9 | 111 ± 12 | −5 ± 3 A,B | |
| HP | 114 ± 11 | 116 ± 9 | 1 ± 4 A | ||
| All | 115 ± 9 | 114 ± 10 | −2 ± 5 | ||
| (n = 22) | (n = 22) | (n = 22) | |||
| Fasting glucose, mmol/L | NP | 5.5 ± 0.6 | 5.5 ± 0.4 | 0.0 ± 0.6 | |
| HP | 5.6 ± 0.5 | 5.4 ± 0.5 | −0.1 ± 0.4 | ||
| All | 5.5 ± 0.5 | 5.5 ± 0.5 | −0.1 ± 0.5 | ||
| (n = 19) | (n = 22) | (n = 19) | |||
| Fasting insulin, mmol/L | NP | 91.0 ± 50.0 | 82.6 ± 36.1 | −8.3 ± 38.9 | |
| HP | 90.3 ± 57.6 | 62.5 ± 37.5 | −35.4 ± 34.7 | ||
| All | 91 ± 52.1 | 69.5 ± 36.8 | −21.5 ± 38.2 | ||
| HOMA-IR | NP | 3.26 ± 1.98 | 2.92 ± 1.33 | −0.35 ± 1.59 | |
| HP | 3.32 ± 2.43 | 2.23 ± 1.43 | −1.37 ± 1.51 | ||
| All | 3.29 ± 2.15 | 2.55 ± 1.40 | −0.83 ± 1.60 | ||
| (n = 22) | (n = 22) | (n = 22) | |||
| Total cholesterol, mmol/L | NP | 4.8 ± 0.7 | 4.6 ± 0.8 | −0.2 ± 0.4 | |
| HP | 5.0 ± 1.2 | 5.0 ± 1.1 | <−0.1 ± 0.6 | ||
| All | 4.9 ± 1.0 | 4.8 ± 1.0 | −0.1 ± 0.5 | ||
| LDL cholesterol, mmol/L | NP | 2.9 ± 0.5 | 2.6 ± 0.5 | −0.3 ± 0.3 A,B | |
| HP | 3.1 ± 1.1 | 3.2 ± 1.0 | 0.1 ± 0.4 A | ||
| All | 3.0 ± 0.8 | 3.0 ± 0.9 | <−0.1 ± 0.4 | ||
| HDL cholesterol, mmol/L | NP | 1.2 ± 0.5 | 1.2 ± 0.5 | <−0.1 ± 0.2 | |
| HP | 1.3 ± 0.3 | 1.2 ± 0.3 | −0.1 ± 0.2 | ||
| All | 1.3 ± 0.4 | 1.2 ± 0.4 | −0.1 ± 0.2 | ||
| Cholesterol: HDL | NP | 4.33 ± 1.50 | 4.12 ± 1.27 | −0.21 ± 0.56 | |
| HP | 4.04 ± 1.02 | 4.18 ± 0.95 | 0.14 ± 0.36 | ||
| All | 4.17 ± 1.24 | 4.15 ± 1.08 | −0.02 ± 0.48 | ||
| Triglycerides, mmol/L | NP | 1.5 ± 0.7 | 1.6 ± 0.8 | 0.1 ± 0.4 | |
| HP | 1.5 ± 0.6 | 1.2 ± 0.4 | −0.3 ± 0.6 | ||
| All | 1.5 ± 0.7 | 1.4 ± 0.6 | −0.1 ± 0.5 | ||
| Manual SBP 3, mm HG | NP | 134 ± 15 | 132 ± 11 | −2 ± 12 | |
| HP | 130 ± 14 | 123 ± 17 | −7 ± 13 | ||
| All | 132 ± 14 | 127 ± 14 | −4 ± 13 | ||
| Manual DBP 3, mm HG | NP | 86 ± 8 | 84 ± 7 | −2 ± 6 | |
| HP | 83 ± 6 | 78 ± 10 | −5 ± 5 | ||
| All | 84 ± 7 | 81 ± 9 | −4 ± 6 | ||
| BUN 2, mmol/L | NP | 5.8 ± 0.9 | 5.6 ± 1.2 C | −0.2 ± 1.2 A | |
| HP | 6.0 ± 1.4 | 7.6 ± 2.6 C | 1.6 ± 1.8 A,B | ||
| All | 5.9 ± 1.2 | 6.7 ± 2.2 | 0.8 ± 1.8 | ||
| (n = 17) | (n = 17) | (n = 17) | |||
| IGF-1, pg/ml | NP | 1.52 ± 0.99 | 1.37 ± 0.69 | −0.15 ± 0.43 | |
| HP | 1.95 ± 1.71 | 2.82 ± 2.47 | 0.99 ± 1.81 | ||
| All | 1.80 ± 1.47 | 2.31 ± 2.11 | 0.56 ± 1.53 B | ||
| Baseline | Post | ∆ | ||
|---|---|---|---|---|
| (n = 21) | (n = 20) | (n = 20) | ||
| HsCRP, mg/L | NP | 2.25 ± 1.50 | 2.10 ± 1.77 | −0.14 ± 0.97 |
| HP | 1.32 ± 1.08 | 1.62 ± 1.49 | 0.30 ± 0.77 | |
| All | 1.72 ± 1.35 | 1.83 ± 1.64 | 0.11 ± 0.89 B | |
| (n = 21) | (n = 20) | (n = 20) | ||
| TNFα, pg/mL | NP | 2.93 ± 1.47 | 3.37 ± 1.31 | 1.06 ± 1.49 |
| HP | 2.97 ± 1.06 | 4.52 ± 2.11 | 0.65 ± 1.99 | |
| All | 2.95 ± 1.24 | 4.00 ± 1.84 | 0.84 ± 1.74 | |
| (n = 19) | (n = 19) | (n = 19) | ||
| IL-6, pg/mL | NP | 3.55 ± 2.01 | 4.04 ± 2.50 | 1.04 ± 2.82 |
| HP | 2.62 ± 1.04 | 3.36 ± 1.68 | 0.75 ± 1.56 | |
| All | 3.06 ± 1.60 | 3.38 ± 2.07 | 0.89 ± 2.20 | |
| (n = 20) | (n = 20) | (n = 20) | ||
| MCP-1, pg/mL | NP | 154.99 ± 42.42 | 175.45 ± 28.92 | 20.11 ± 45.26 |
| HP | 164.22 ± 36.47 | 167.79 ± 49.30 | 3.57 ± 27.40 | |
| All | 160.07 ± 38.47 | 171.24 ± 40.58 | 11.44 ± 36.99 B | |
| (n = 19) | (n = 20) | (n = 19) | ||
| Adiponectin, μg/mL | NP | 9.48 ± 2.39 | 8.07 ± 3.44 | −0.92 ± 3.20 |
| HP | 11.76 ± 4.31 | 11.37 ± 5.63 | −0.76 ± 4.62 | |
| All | 10.69 ± 9.72 | 9.72 ± 4.85 | −0.83 ± 3.94 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wright, C.S.; Zhou, J.; Sayer, R.D.; Kim, J.E.; Campbell, W.W. Effects of a High-Protein Diet Including Whole Eggs on Muscle Composition and Indices of Cardiometabolic Health and Systemic Inflammation in Older Adults with Overweight or Obesity: A Randomized Controlled Trial. Nutrients 2018, 10, 946. https://doi.org/10.3390/nu10070946
Wright CS, Zhou J, Sayer RD, Kim JE, Campbell WW. Effects of a High-Protein Diet Including Whole Eggs on Muscle Composition and Indices of Cardiometabolic Health and Systemic Inflammation in Older Adults with Overweight or Obesity: A Randomized Controlled Trial. Nutrients. 2018; 10(7):946. https://doi.org/10.3390/nu10070946
Chicago/Turabian StyleWright, Christian S., Jing Zhou, R. Drew Sayer, Jung Eun Kim, and Wayne W. Campbell. 2018. "Effects of a High-Protein Diet Including Whole Eggs on Muscle Composition and Indices of Cardiometabolic Health and Systemic Inflammation in Older Adults with Overweight or Obesity: A Randomized Controlled Trial" Nutrients 10, no. 7: 946. https://doi.org/10.3390/nu10070946
APA StyleWright, C. S., Zhou, J., Sayer, R. D., Kim, J. E., & Campbell, W. W. (2018). Effects of a High-Protein Diet Including Whole Eggs on Muscle Composition and Indices of Cardiometabolic Health and Systemic Inflammation in Older Adults with Overweight or Obesity: A Randomized Controlled Trial. Nutrients, 10(7), 946. https://doi.org/10.3390/nu10070946