
A Naturally Derived Watercress Flower-Based Phenethyl Isothiocyanate-Enriched Extract Induces the Activation of Intrinsic Apoptosis via Subcellular Ultrastructural and Ca2+ Efflux Alterations in an In Vitro Model of Human Malignant Melanoma
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
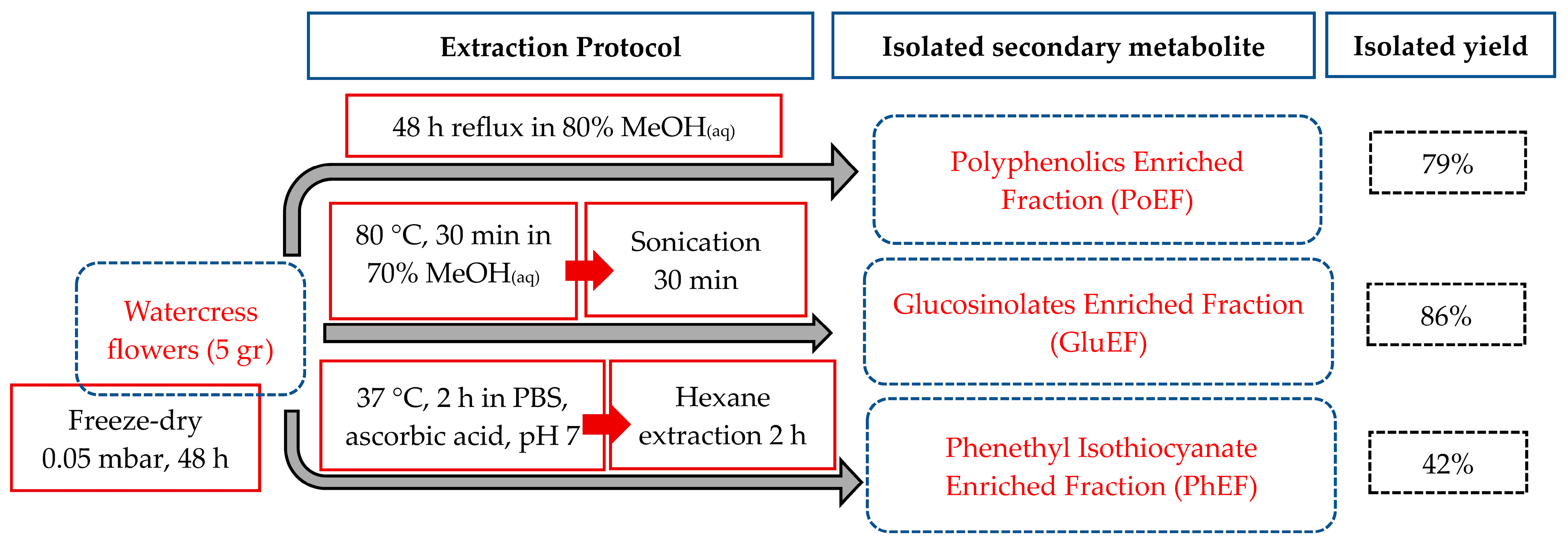
2.1. Plant Material Cultivation, Processing and Storage
2.2. Extraction of Phenethyl Isothiocyanate Content
2.3. Extraction of Polyphenols’ Content
2.4. Extraction of Glucosinolate Content
2.5. Cell Lines
2.6. Determination of Cytotoxicity
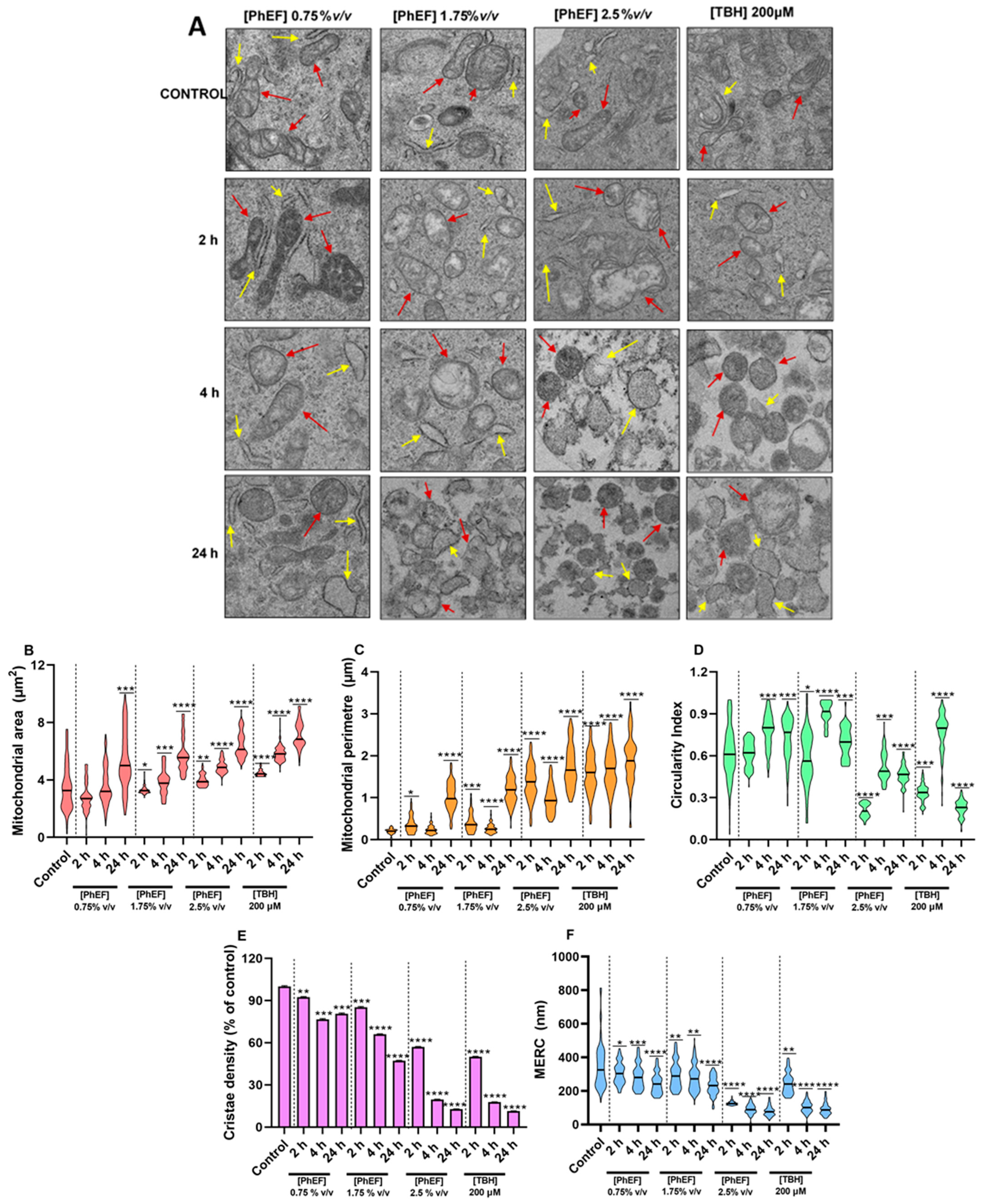
2.7. Transmission Electron Microscope (TEM) Imaging and Quantification
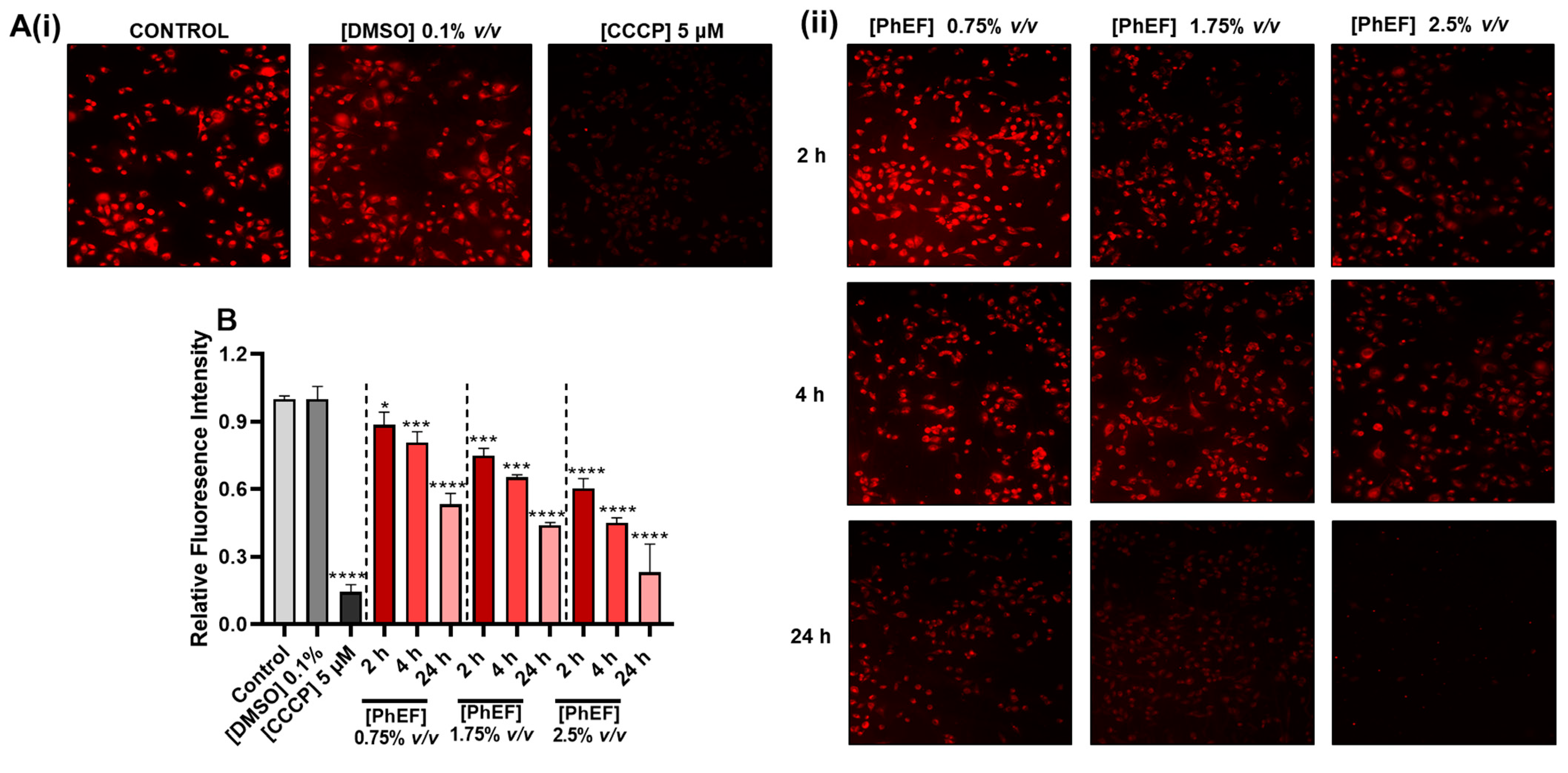
2.8. Determination of Mitochondrial Membrane Depolarization
2.9. Determination of Caspase Activity
2.10. Statistical Analyses
3. Results
3.1. Plant Processing, Extraction and Isolation of Polyphenolics, Glucosinolates and Phenethyl Isothiocyanate-Enriched Fractions
3.2. Cytotoxic Profile of Isolated Polyphenol, Phenethyl Isothiocyanate and Glucosinolate-Enriched Extracts
3.3. Characterization of PhEF-Induced Ultrastructural Subcellular Alterations
3.4. Determination of PhEF-Induced Mitochondrial Membrane Depolarization
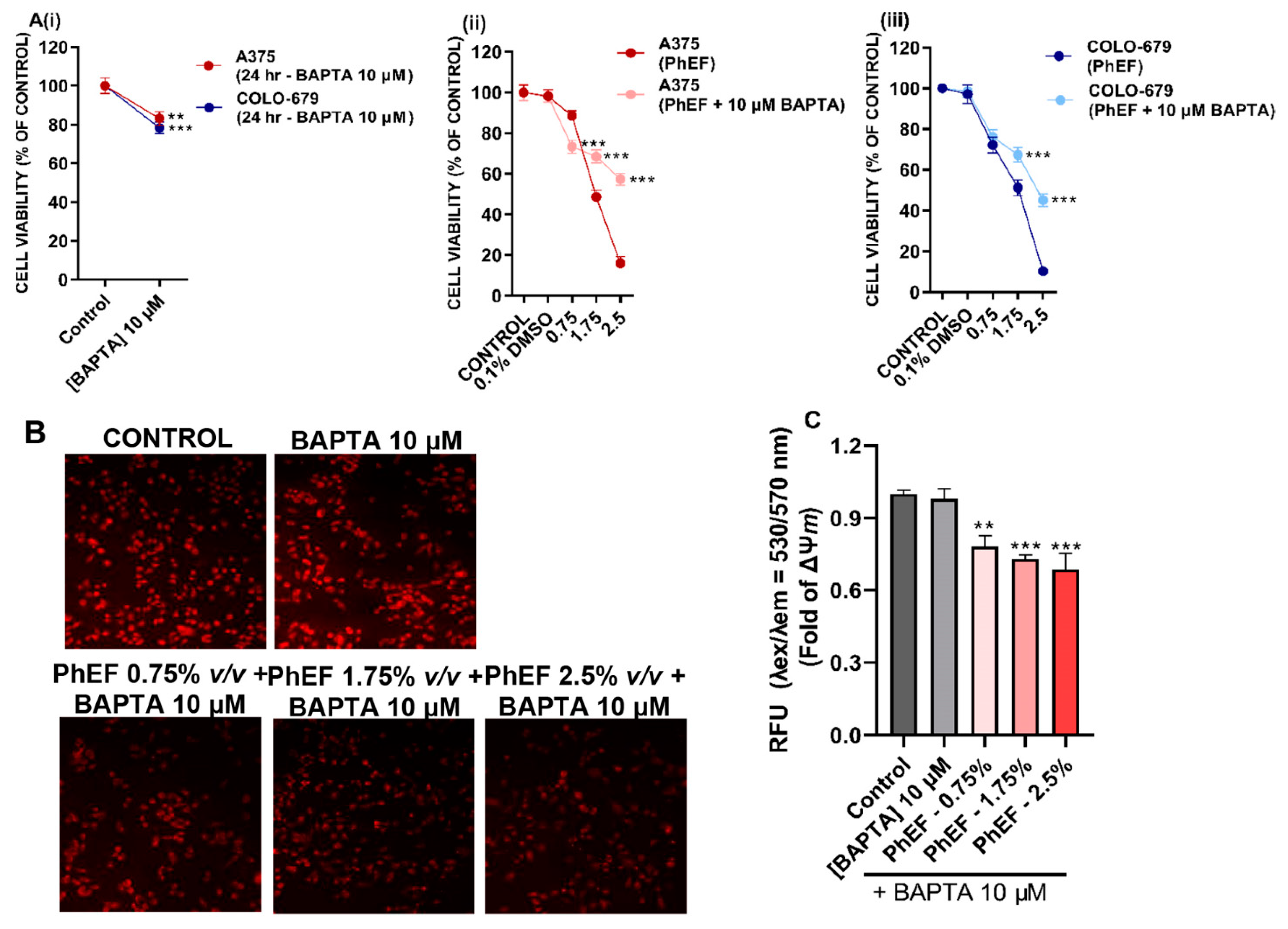
3.5. Determination of PhEF-Induced Ca2+ Efflux Perturbations
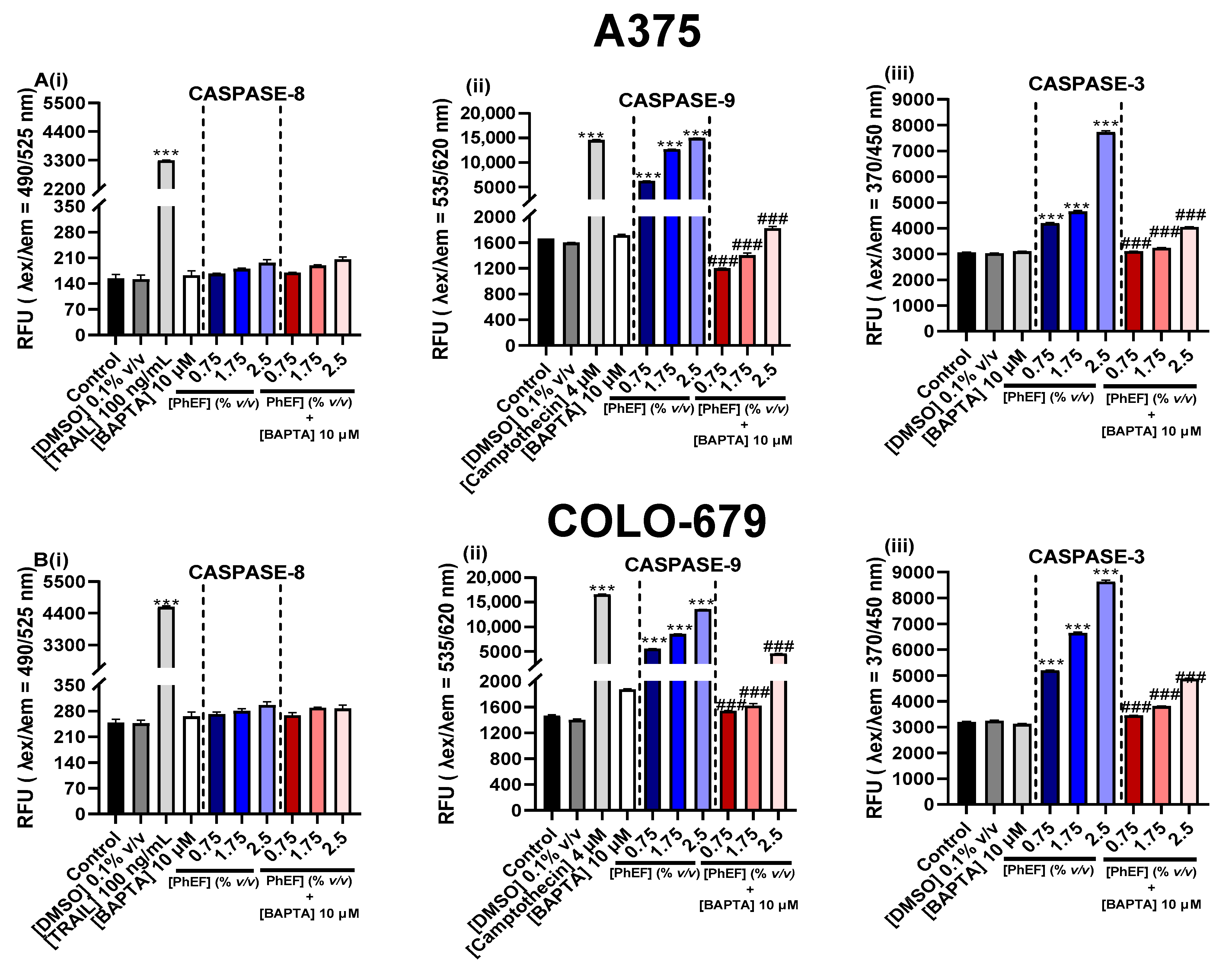
3.6. Determination of PhEF-Induced Activation of Caspases-8, -9 and -3
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kristal, A.R.; Lampe, J.W. Brassica Vegetables and Prostate Cancer Risk: A Review of the Epidemiological Evidence. Nutr. Cancer 2002, 42, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kolonel, L.N.; Hankin, J.H.; Whittemore, A.S.; Wu, A.H.; Gallagher, R.P.; Wilkens, L.R.; John, E.M.; Howe, G.R.; Dreon, D.M.; West, D.W.; et al. Vegetables, Fruits, Legumes and Prostate Cancer: A Multiethnic Case-Control Study1. Cancer Epidemiol. Biomark. Prev. 2000, 9, 795–804. [Google Scholar]
- Hadjzadeh, M.-A.-R.; Rajaei, Z.; Moradi, R.; Ghorbani, A. Effects of Hydroalcoholic Extract of Watercress (Nasturtium officinale) Leaves on Serum Glucose and Lipid Levels in Diabetic Rats. Indian J. Physiol. Pharmacol. 2015, 59, 223–230. [Google Scholar] [PubMed]
- Shahani, S.; Behzadfar, F.; Jahani, D.; Ghasemi, M.; Shaki, F. Antioxidant and Anti-Inflammatory Effects of Nasturtium officinale Involved in Attenuation of Gentamicin-Induced Nephrotoxicity. Toxicol. Mech. Methods 2017, 27, 107–114. [Google Scholar] [CrossRef]
- Klimek-Szczykutowicz, M.; Szopa, A.; Ekiert, H. Chemical Composition, Traditional and Professional Use in Medicine, Application in Environmental Protection, Position in Food and Cosmetics Industries, and Biotechnological Studies of Nasturtium officinale (Watercress)—A Review. Fitoterapia 2018, 129, 283–292. [Google Scholar] [CrossRef]
- Fenton-Navarro, B.; Martínez, M.U.; Castro, B.F.; Castillo, O.M.; López-Rodríguez, M.; Arellanes, S.P.; Hernadez, A.V. Antioxidant and Hypoglycemic Effects of Watercress (Nasturtium officinale) Extracts in Diabetic Rats. Afr. J. Tradit. Complement. Altern. Med. 2018, 15, 68–79. [Google Scholar] [CrossRef]
- Qian, Y.; Hibbert, L.E.; Milner, S.; Katz, E.; Kliebenstein, D.J.; Taylor, G. Improved Yield and Health Benefits of Watercress Grown in an Indoor Vertical Farm. Sci. Hortic. 2022, 300, 111068. [Google Scholar] [CrossRef]
- Panahi Kokhdan, E.; Khodabandehloo, H.; Ghahremani, H.; Doustimotlagh, A.H. A Narrative Review on Therapeutic Potentials of Watercress in Human Disorders. Evid.-Based Complement. Altern. Med. 2021, 2021, 5516450. [Google Scholar] [CrossRef]
- Singh, S.V.; Singh, K. Cancer Chemoprevention with Dietary Isothiocyanates Mature for Clinical Translational Research. Carcinogenesis 2012, 33, 1833–1842. [Google Scholar] [CrossRef]
- Giallourou, N.S.; Rowland, I.R.; Rothwell, S.D.; Packham, G.; Commane, D.M.; Swann, J.R. Metabolic Targets of Watercress and PEITC in MCF-7 and MCF-10A Cells Explain Differential Sensitisation Responses to Ionising Radiation. Eur. J. Nutr. 2019, 58, 2377–2391. [Google Scholar] [CrossRef]
- Syed Alwi, S.S.; Cavell, B.E.; Telang, U.; Morris, M.E.; Parry, B.M.; Packham, G. In Vivo Modulation of 4E Binding Protein 1 (4E-BP1) Phosphorylation by Watercress: A Pilot Study. Br. J. Nutr. 2010, 104, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- Mitsiogianni, M.; Koutsidis, G.; Mavroudis, N.; Trafalis, D.T.; Botaitis, S.; Franco, R.; Zoumpourlis, V.; Amery, T.; Galanis, A.; Pappa, A.; et al. The Role of Isothiocyanates as Cancer Chemo-Preventive, Chemo-Therapeutic and Anti-Melanoma Agents. Antioxidants 2019, 8, 106. [Google Scholar] [CrossRef] [PubMed]
- Coscueta, E.R.; Reis, C.A.; Pintado, M. Phenylethyl Isothiocyanate Extracted from Watercress By-Products with Aqueous Micellar Systems: Development and Optimisation. Antioxidants 2020, 9, 698. [Google Scholar] [CrossRef]
- Choi, M.; Lee, C. Immortalization of Primary Keratinocytes and Its Application to Skin Research. Biomol. Ther. 2015, 23, 391–399. [Google Scholar] [CrossRef]
- Araújo-Rodrigues, H.; Santos, D.; Campos, D.A.; Guerreiro, S.; Ratinho, M.; Rodrigues, I.M.; Pintado, M.E. Impact of Processing Approach and Storage Time on Bioactive and Biological Properties of Rocket, Spinach and Watercress Byproducts. Foods 2021, 10, 2301. [Google Scholar] [CrossRef] [PubMed]
- Zeb, A. Phenolic Profile and Antioxidant Potential of Wild Watercress (Nasturtium officinale L.). Springerplus 2015, 4, 714. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.; Faulkner, K.; Williamson, G.; Mithen, R. 7-Methylsulfinylheptyl and 8-Methylsulfinyloctyl Isothiocyanates from Watercress Are Potent Inducers of Phase II Enzymes. Carcinogenesis 2000, 21, 1983–1988. [Google Scholar] [CrossRef]
- Kyriakou, S.; Tragkola, V.; Alghol, H.; Anestopoulos, I.; Amery, T.; Stewart, K.; Winyard, P.G.; Trafalis, D.T.; Franco, R.; Pappa, A.; et al. Evaluation of Bioactive Properties of Lipophilic Fractions of Edible and Non-Edible Parts of Nasturtium officinale (Watercress) in a Model of Human Malignant Melanoma Cells. Pharmaceuticals 2022, 15, 141. [Google Scholar] [CrossRef]
- Kyriakou, S.; Michailidou, K.; Amery, T.; Stewart, K.; Winyard, P.G.; Trafalis, D.T.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Polyphenolics, Glucosinolates and Isothiocyanates Profiling of Aerial Parts of Nasturtium officinale (Watercress). Front. Plant Sci. 2022, 13, 998755. [Google Scholar] [CrossRef] [PubMed]
- Schiestl, F.P. Correlation Analyses between Volatiles and Glucosinolates Show No Evidence for Chemical Defense Signaling in Brassica Rapa. Front. Ecol. Evol. 2014, 2, 10. [Google Scholar] [CrossRef]
- Strauss, S.Y.; Irwin, R.E.; Lambrix, V.M. Optimal Defence Theory and Flower Petal Colour Predict Variation in the Secondary Chemistry of Wild Radish. J. Ecol. 2004, 92, 132–141. [Google Scholar] [CrossRef]
- Bouwmeester, H.; Schuurink, R.C.; Bleeker, P.M.; Schiestl, F. The Role of Volatiles in Plant Communication. Plant J. 2019, 100, 892–907. [Google Scholar] [CrossRef] [PubMed]
- Boyd, L.A.; McCann, M.J.; Hashim, Y.; Bennett, R.N.; Gill, C.I.R.; Rowland, I.R. Assessment of the Anti-Genotoxic, Anti-Proliferative, and Anti-Metastatic Potential of Crude Watercress Extract in Human Colon Cancer Cells. Nutr. Cancer 2006, 55, 232–241. [Google Scholar] [CrossRef]
- Gill, C.I.R.; Haldar, S.; Boyd, L.A.; Bennett, R.; Whiteford, J.; Butler, M.; Pearson, J.R.; Bradbury, I.; Rowland, I.R. Watercress Supplementation in Diet Reduces Lymphocyte DNA Damage and Alters Blood Antioxidant Status in Healthy Adults. Am. J. Clin. Nutr. 2007, 85, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Fallah, N.; Ebrahimi, S. The Anti-Cancer Effect of Watercress (Rorripa Nasturtium Aquaticum) Extract on Breast Cancer Cells. Zahedan J. Res. Med. Sci. 2016, 18, e2725. [Google Scholar] [CrossRef]
- Yazdanparast, R.; Bahramikia, S.; Ardestani, A. Nasturtium officinale Reduces Oxidative Stress and Enhances Antioxidant Capacity in Hypercholesterolaemic Rats. Chem. Biol. Interact. 2008, 172, 176–184. [Google Scholar] [CrossRef]
- Aires, A.; Carvalho, R.; Rosa, E.A.S.; Saavedra, M.J. Phytochemical Characterization and Antioxidant Properties of Baby-Leaf Watercress Produced under Organic Production System. CyTA—J. Food 2013, 11, 343–351. [Google Scholar] [CrossRef]
- Gao, N.; Budhraja, A.; Cheng, S.; Liu, E.-H.; Chen, J.; Yang, Z.; Chen, D.; Zhang, Z.; Shi, X. Phenethyl Isothiocyanate Exhibits Antileukemic Activity In Vitro and In Vivo by Inactivation of Akt and Activation of JNK Pathways. Cell Death Dis. 2011, 2, e140. [Google Scholar] [CrossRef]
- Mitsiogianni, M.; Anestopoulos, I.; Kyriakou, S.; Trafalis, D.T.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Benzyl and Phenethyl Isothiocyanates as Promising Epigenetic Drug Compounds by Modulating Histone Acetylation and Methylation Marks in Malignant Melanoma. Investig. New Drugs 2021, 39, 1460–1468. [Google Scholar] [CrossRef]
- Mi, L.; Gan, N.; Chung, F.-L. Isothiocyanates Inhibit Proteasome Activity and Proliferation of Multiple Myeloma Cells. Carcinogenesis 2011, 32, 216–223. [Google Scholar] [CrossRef]
- Tusskorn, O.; Senggunprai, L.; Prawan, A.; Kukongviriyapan, U.; Kukongviriyapan, V. Phenethyl Isothiocyanate Induces Calcium Mobilization and Mitochondrial Cell Death Pathway in Cholangiocarcinoma KKU-M214 Cells. BMC Cancer 2013, 13, 571. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Shi, Y.; Dang, U.J.; Di Pasqua, A.J. Phenethyl Isothiocyanate and Cisplatin Co-Encapsulated in a Liposomal Nanoparticle for Treatment of Non-Small Cell Lung Cancer. Molecules 2019, 24, 801. [Google Scholar] [CrossRef] [PubMed]
- Kyriakou, S.; Trafalis, D.T.; Deligiorgi, M.V.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Assessment of Methodological Pipelines for the Determination of Isothiocyanates Derived from Natural Sources. Antioxidants 2022, 11, 642. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Ma, W.Y.; Li, J.; Hecht, S.S.; Dong, Z. Essential Role of P53 in Phenethyl Isothiocyanate-Induced Apoptosis. Cancer Res. 1998, 58, 4102–4106. [Google Scholar]
- Yu, R.; Jiao, J.J.; Duh, J.L.; Tan, T.H.; Kong, A.N. Phenethyl Isothiocyanate, a Natural Chemopreventive Agent, Activates c-Jun N-Terminal Kinase 1. Cancer Res. 1996, 56, 2954–2959. [Google Scholar]
- Gupta, P.; Wright, S.E.; Kim, S.-H.; Srivastava, S.K. Phenethyl Isothiocyanate: A Comprehensive Review of Anti-Cancer Mechanisms. Biochim. Biophys. Acta 2014, 1846, 405–424. [Google Scholar] [CrossRef]
- Acuna, U.M.; Ezzone, N.; Rakotondraibe, L.H.; Carcache DE Blanco, E.J. Activity in MCF-7 Estrogen-Sensitive Breast Cancer Cells of Capsicodendrin from Cinnamosma fragrans. Anticancer Res. 2021, 41, 5935–5944. [Google Scholar] [CrossRef]
- Tseng, E.; Scott-Ramsay, E.A.; Morris, M.E. Dietary Organic Isothiocyanates Are Cytotoxic in Human Breast Cancer MCF-7 and Mammary Epithelial MCF-12A Cell Lines. Exp. Biol. Med. 2004, 229, 835–842. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and Antagonism in Natural Product Extracts: When 1 + 1 Does Not Equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef]
- Yilmaz, V.; Louca, P.; Potamiti, L.; Panayiotidis, M.; Strati, K. A Novel Lineage-Tracing Mouse Model for Studying Early MmuPV1 Infections. eLife 2022, 11, e72638. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Sharma, K.; Ishaq, M.; Sharma, G.; Khan, M.A.; Dutta, R.K.; Majumdar, S. Pentoxifylline Triggers Autophagy via ER Stress Response That Interferes with Pentoxifylline Induced Apoptosis in Human Melanoma Cells. Biochem. Pharmacol. 2016, 103, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Lam, J.; Katti, P.; Biete, M.; Mungai, M.; AshShareef, S.; Neikirk, K.; Garza Lopez, E.; Vue, Z.; Christensen, T.A.; Beasley, H.K.; et al. A Universal Approach to Analyzing Transmission Electron Microscopy with ImageJ. Cells 2021, 10, 2177. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.; Bong, S.J.; Park, J.S.; Park, Y.-K.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.U. De Novo Transcriptome Analysis and Glucosinolate Profiling in Watercress (Nasturtium officinale R. Br.). BMC Genom. 2017, 18, 401. [Google Scholar] [CrossRef] [PubMed]
- Sander, C.S.; Hamm, F.; Elsner, P.; Thiele, J.J. Oxidative Stress in Malignant Melanoma and Non-Melanoma Skin Cancer. Br. J. Dermatol. 2003, 148, 913–922. [Google Scholar] [CrossRef]
- Meierjohann, S. Oxidative Stress in Melanocyte Senescence and Melanoma Transformation. Eur. J. Cell Biol. 2014, 93, 36–41. [Google Scholar] [CrossRef]
- Jenkins, N.C.; Grossman, D. Role of Melanin in Melanocyte Dysregulation of Reactive Oxygen Species. BioMed Res. Int. 2013, 2013, 908797. [Google Scholar] [CrossRef]
- Becker, T.M.; Rizos, H.; Kefford, R.F.; Mann, G.J. Functional Impairment of Melanoma-Associated P16INK4a Mutants in Melanoma Cells despite Retention of Cyclin-Dependent Kinase 4 Binding1. Clin. Cancer Res. 2001, 7, 3282–3288. [Google Scholar]
- Xian, D.; Lai, R.; Song, J.; Xiong, X.; Zhong, J. Emerging Perspective: Role of Increased ROS and Redox Imbalance in Skin Carcinogenesis. Oxid. Med. Cell. Longev. 2019, 2019, 8127362. [Google Scholar] [CrossRef]
- Cesi, G.; Walbrecq, G.; Zimmer, A.; Kreis, S.; Haan, C. ROS Production Induced by BRAF Inhibitor Treatment Rewires Metabolic Processes Affecting Cell Growth of Melanoma Cells. Mol. Cancer 2017, 16, 102. [Google Scholar] [CrossRef]
- Venza, I.; Venza, M.; Visalli, M.; Lentini, G.; Teti, D.; d’Alcontres, F.S. ROS as Regulators of Cellular Processes in Melanoma. Oxid. Med. Cell. Longev. 2021, 2021, 1208690. [Google Scholar] [CrossRef] [PubMed]
- Pizzimenti, S.; Ribero, S.; Cucci, M.A.; Grattarola, M.; Monge, C.; Dianzani, C.; Barrera, G.; Muzio, G. Oxidative Stress-Related Mechanisms in Melanoma and in the Acquired Resistance to Targeted Therapies. Antioxidants 2021, 10, 1942. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, N.C.; Liu, T.; Cassidy, P.; Leachman, S.A.; Boucher, K.M.; Goodson, A.G.; Samadashwily, G.; Grossman, D. The P16INK4A Tumor Suppressor Regulates Cellular Oxidative Stress. Oncogene 2011, 30, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Nastruzzi, C.; Cortesi, R.; Esposito, E.; Menegatti, E.; Leoni, O.; Iori, R.; Palmieri, S. In Vitro Cytotoxic Activity of Some Glucosinolate-Derived Products Generated by Myrosinase Hydrolysis. J. Agric. Food Chem. 1996, 44, 1014–1021. [Google Scholar] [CrossRef]
- Đulović, A.; Burčul, F.; Čikeš Čulić, V.; Rollin, P.; Blažević, I. Glucosinolates and Cytotoxic Activity of Collard Volatiles Obtained Using Microwave-Assisted Extraction. Molecules 2023, 28, 1657. [Google Scholar] [CrossRef]
- Merinas-Amo, T.; Lozano-Baena, M.D.; Obregón-Cano, S.; Alonso-Moraga, Á.; de Haro-Bailón, A. Role of Glucosinolates in the Nutraceutical Potential of Selected Cultivars of Brassica Rapa. Foods 2021, 10, 2720. [Google Scholar] [CrossRef]
- Gamet-Payrastre, L.; Lumeau, S.; Gasc, N.; Cassar, G.; Rollin, P.; Tulliez, J. Selective Cytostatic and Cytotoxic Effects of Glucosinolates Hydrolysis Products on Human Colon Cancer Cells in Vitro. Anticancer Drugs 1998, 9, 141–148. [Google Scholar] [CrossRef]
- Kołodziejski, D.; Piekarska, A.; Hanschen, F.S.; Pilipczuk, T.; Tietz, F.; Kusznierewicz, B.; Bartoszek, A. Relationship between Conversion Rate of Glucosinolates to Isothiocyanates/Indoles and Genotoxicity of Individual Parts of Brassica Vegetables. Eur. Food Res. Technol. 2019, 245, 383–400. [Google Scholar] [CrossRef]
- Hanschen, F.S.; Kaufmann, M.; Kupke, F.; Hackl, T.; Kroh, L.W.; Rohn, S.; Schreiner, M. Brassica Vegetables as Sources of Epithionitriles: Novel Secondary Products Formed during Cooking. Food Chem. 2018, 245, 564–569. [Google Scholar] [CrossRef]
- Latimer, I.; Chand, R.; Cridge, B. The in Vitro Toxicity of Nitrile and Epithionitrile Derivatives of Glucosinolates from Swedes (Brassica napus) in Human and Bovine Liver Cells. N. Z. Vet. J. 2021, 69, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.-Y.; Huang, Y.-T.; Yu, C.-S.; Ko, Y.-C.; Wu, S.-H.; Ji, B.-C.; Yang, J.-S.; Yang, J.-L.; Hsia, T.-C.; Chen, Y.-Y.; et al. Phenethyl Isothiocyanate (PEITC) Promotes G2/M Phase Arrest via P53 Expression and Induces Apoptosis through Caspase- and Mitochondria-Dependent Signaling Pathways in Human Prostate Cancer DU 145 Cells. Anticancer Res. 2011, 31, 1691–1702. [Google Scholar]
- Dai, M.Y.; Wang, Y.; Chen, C.; Li, F.; Xiao, B.K.; Chen, S.M.; Tao, Z.Z. Phenethyl Isothiocyanate Induces Apoptosis and Inhibits Cell Proliferation and Invasion in Hep-2 Laryngeal Cancer Cells. Oncol. Rep. 2016, 35, 2657–2664. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-Y.; Lin, K.-C.; Lin, J.-P.; Tang, N.-Y.; Yang, J.-S.; Lu, K.-W.; Chung, J.-G. Phenethyl Isothiocyanate (PEITC) Inhibits the Growth of Human Oral Squamous Carcinoma HSC-3 Cells through G 0/G 1 Phase Arrest and Mitochondria-Mediated Apoptotic Cell Death. Evid.-Based Complement. Altern. Med. 2012, 2012, 718320. [Google Scholar] [CrossRef] [PubMed]
- Eisa, N.H.; Said, H.S.; ElSherbiny, N.M.; Eissa, L.A.; El-Shishtawy, M.M. Phenethyl Isothiocyanate Triggers Apoptosis, Combats Oxidative Stress and Inhibits Growth of Ehrlich Ascites Carcinoma Mouse Model. Iran. J. Pharm. Res. 2018, 17, 1328–1338. [Google Scholar]
- Thomson, S.J.; Brown, K.K.; Pullar, J.M.; Hampton, M.B. Phenethyl Isothiocyanate Triggers Apoptosis in Jurkat Cells Made Resistant by the Overexpression of Bcl-2. Cancer Res. 2006, 66, 6772–6777. [Google Scholar] [CrossRef] [PubMed]
- Hsia, T.-C.; Huang, Y.-P.; Jiang, Y.-W.; Chen, H.-Y.; Cheng, Z.-Y.; Hsiao, Y.-T.; Chen, C.-Y.; Peng, S.-F.; Chueh, F.-S.; Chou, Y.-C.; et al. Phenethyl Isothiocyanate Induces Apoptotic Cell Death Through the Mitochondria-Dependent Pathway in Gefitinib-Resistant NCI-H460 Human Lung Cancer Cells In Vitro. Anticancer Res. 2018, 38, 2137–2147. [Google Scholar]
- Sun, M.G.; Williams, J.; Munoz-Pinedo, C.; Perkins, G.A.; Brown, J.M.; Ellisman, M.H.; Green, D.R.; Frey, T.G. Correlated Three-Dimensional Light and Electron Microscopy Reveals Transformation of Mitochondria during Apoptosis. Nature 2007, 9, 1057–1065. [Google Scholar] [CrossRef]
- Bernardi, P.; Di Lisa, F. The Mitochondrial Permeability Transition Pore: Molecular Nature and Role as a Target in Cardioprotection. J. Mol. Cell. Cardiol. 2015, 78, 100–106. [Google Scholar] [CrossRef]
- Hirsch, T.; Marzo, I.; Kroemer, G. Role of the Mitochondrial Permeability Transition Pore in Apoptosis. Biosci. Rep. 1997, 17, 67–76. [Google Scholar] [CrossRef]
- Bonora, M.; Pinton, P. The Mitochondrial Permeability Transition Pore and Cancer: Molecular Mechanisms Involved in Cell Death. Front. Oncol. 2014, 4, 302. [Google Scholar] [CrossRef]
- Scorrano, L.; Ashiya, M.; Buttle, K.; Weiler, S.; Oakes, S.A.; Mannella, C.A.; Korsmeyer, S.J. A Distinct Pathway Remodels Mitochondrial Cristae and Mobilizes Cytochrome c during Apoptosis. Dev. Cell 2002, 2, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Garrido, C.; Galluzzi, L.; Brunet, M.; Puig, P.E.; Didelot, C.; Kroemer, G. Mechanisms of Cytochrome c Release from Mitochondria. Cell Death Differ. 2006, 13, 1423–1433. [Google Scholar] [CrossRef]
- Yang, R.-F.; Zhao, G.-W.; Liang, S.-T.; Zhang, Y.; Sun, L.-H.; Chen, H.-Z.; Liu, D.-P. Mitofilin Regulates Cytochrome c Release during Apoptosis by Controlling Mitochondrial Cristae Remodeling. Biochem. Biophys. Res. Commun. 2012, 428, 93–98. [Google Scholar] [CrossRef]
- Chapa-Dubocq, X.; Makarov, V.; Javadov, S. Simple Kinetic Model of Mitochondrial Swelling in Cardiac Cells. J. Cell. Physiol. 2018, 233, 5310–5321. [Google Scholar] [CrossRef]
- Javadov, S.; Chapa-Dubocq, X.; Makarov, V. Different Approaches to Modeling Analysis of Mitochondrial Swelling. Mitochondrion 2018, 38, 58–70. [Google Scholar] [CrossRef]
- Lizák, B.; Birk, J.; Zana, M.; Kosztyi, G.; Kratschmar, D.V.; Odermatt, A.; Zimmermann, R.; Geiszt, M.; Appenzeller-Herzog, C.; Bánhegyi, G. Ca2+ Mobilization-Dependent Reduction of the Endoplasmic Reticulum Lumen Is Due to Influx of Cytosolic Glutathione. BMC Biol. 2020, 18, 19. [Google Scholar] [CrossRef]
- Wilkinson, S. ER-Phagy: Shaping up and Destressing the Endoplasmic Reticulum. FEBS J. 2019, 286, 2645–2663. [Google Scholar] [CrossRef]
- Pinton, P.; Giorgi, C.; Siviero, R.; Zecchini, E.; Rizzuto, R. Calcium and Apoptosis: ER-Mitochondria Ca2+ Transfer in the Control of Apoptosis. Oncogene 2008, 27, 6407–6418. [Google Scholar] [CrossRef]
- Sukumaran, P.; Da Conceicao, V.N.; Sun, Y.; Ahamad, N.; Saraiva, L.R.; Selvaraj, S.; Singh, B.B. Calcium Signaling Regulates Autophagy and Apoptosis. Cells 2021, 10, 2125. [Google Scholar] [CrossRef]
- Ajiro, K.; Bortner, C.D.; Westmoreland, J.; Cidlowski, J.A. An Endogenous Calcium-Dependent, Caspase-Independent Intranuclear Degradation Pathway in Thymocyte Nuclei: Antagonism by Physiological Concentrations of K+ Ions. Exp. Cell Res. 2008, 314, 1237–1249. [Google Scholar] [CrossRef]
- Lv, Z.; Song, X.; Xu, J.; Jia, Z.; Yang, B.; Jia, Y.; Qiu, L.; Wang, L.; Song, L. The Modulation of Smac/DIABLO on Mitochondrial Apoptosis Induced by LPS in Crassostrea gigas. Fish Shellfish Immunol. 2019, 84, 587–598. [Google Scholar] [CrossRef]
- Chai, J.; Du, C.; Wu, J.-W.; Kyin, S.; Wang, X.; Shi, Y. Structural and Biochemical Basis of Apoptotic Activation by Smac/DIABLO. Nature 2000, 406, 855–862. [Google Scholar] [CrossRef]
- Scorrano, L.; Oakes, S.A.; Opferman, J.T.; Cheng, E.H.; Sorcinelli, M.D.; Pozzan, T.; Korsmeyer, S.J. BAX and BAK Regulation of Endoplasmic Reticulum Ca2+: A Control Point for Apoptosis. Science 2003, 300, 135–139. [Google Scholar] [CrossRef]
- Nutt, L.K.; Pataer, A.; Pahler, J.; Fang, B.; Roth, J.; McConkey, D.J.; Swisher, S.G. Bax and Bak Promote Apoptosis by Modulating Endoplasmic Reticular and Mitochondrial Ca2+ Stores. J. Biol. Chem. 2002, 277, 9219–9225. [Google Scholar] [CrossRef]
- Jones, R.G.; Bui, T.; White, C.; Madesh, M.; Krawczyk, C.M.; Lindsten, T.; Hawkins, B.J.; Kubek, S.; Frauwirth, K.A.; Wang, Y.L.; et al. The Proapoptotic Factors Bax and Bak Regulate T Cell Proliferation through Control of Endoplasmic Reticulum Ca2+ Homeostasis. Immunity 2007, 27, 268–280. [Google Scholar] [CrossRef]
- Karch, J.; Kwong, J.Q.; Burr, A.R.; Sargent, M.A.; Elrod, J.W.; Peixoto, P.M.; Martinez-Caballero, S.; Osinska, H.; Cheng, E.H.-Y.; Robbins, J.; et al. Bax and Bak Function as the Outer Membrane Component of the Mitochondrial Permeability Pore in Regulating Necrotic Cell Death in Mice. eLife 2013, 2, e00772. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EC50 (% v/v) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| PhEF | PoEF | GluEF | |||||||
| Time(h) | A375 | COLO-679 | HaCaT | A375 | COLO-679 | HaCaT | A375 | COLO-679 | HaCaT |
| 24 | 1.70 ± 0.10 | 1.80 ± 0.40 | n.d. | 1.8 ± 0.02 | 2.00 ± 0.10 | n.d. | n.d. | n.d. | n.d. |
| 48 | 1.25 ± 0.01 | 1.20 ± 0.20 | 2.10 ± 0.20 | 1.6 ± 0.20 | 1.30 ± 0.01 | 1.80 ± 0.10 | n.d. | n.d. | n.d. |
| 72 | 1.05 ± 0.01 | 1.00 ± 0.10 | 2.00 ± 0.10 | 1.1 ± 0.20 | 1.10 ± 0.02 | 1.20 ± 0.20 | 2.20 ± 0.02 | n.d. | n.d. |
| Fold of Change (Relative to Control) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| [PhEF] 0.75% v/v | [PhEF] 1.75% v/v | [PhEF] 2.50% v/v | [TBH] 200 μM | ||||||||||
| [DMSO] 0.1% v/v | 2 h | 4 h | 24 h | 2 h | 4 h | 24 h | 2 h | 4 h | 24 h | 2 h | 4 h | 24 h | |
| Mitochondrial area (μm2) | 1 | 0.8 ± 0.10 | 1.00 ± 0.10 | 1.50 ± 0.20 | 0.90 ± 0.02 | 1.10 ± 0.08 | 1.60 ± 0.10 | 1.10 ± 0.04 | 1.40 ± 0.04 | 1.80 ± 0.08 | 1.3 ± 0.02 | 1.70 ± 0.05 | 2.06 ± 0.08 |
| Mitochondrial perimeter (μm) | 1 | 1.77 ± 0.02 | 1.15 ± 0.008 | 5.00 ± 0.03 | 1.97 ± 0.02 | 1.30 ± 0.01 | 5.70 ± 0.03 | 6.59 ± 0.04 | 4.72 ± 0.03 | 8.52 ± 0.04 | 7.6 ± 0.04 | 8.06 ± 0.04 | 9.23 ± 0.04 |
| Cristae density (% control) | 1 | 0.92 ± 0.06 | 0.77 ± 0.02 | 0.81 ± 0.03 | 0.85 ± 0.03 | 0.66 ± 0.02 | 0.47 ± 0.02 | 0.57 ± 0.02 | 0.19 ± 0.001 | 0.13 ± 0.01 | 0.50 ± 0.02 | 0.18 ± 0.001 | 0.12 ± 0.001 |
| Circularity Index | 1 | 1.00 ± 0.09 | 1.28 ± 0.11 | 1.17 ± 0.11 | 0.94 ± 0.07 | 1.45 ± 0.08 | 1.12 ± 0.04 | 0.34 ± 0.02 | 0.83 ± 0.05 | 0.74 ± 0.04 | 0.54 ± 0.02 | 1.24 ± 0.10 | 0.36 ± 0.002 |
| MERC length(nm) | 1 | 0.88 ± 0.06 | 0.82 ± 0.04 | 0.71 ± 0.05 | 0.85 ± 0.04 | 0.80 ± 0.02 | 0.66 ± 0.02 | 0.36 ± 0.02 | 0.26 ± 0.01 | 0.23 ± 0.002 | 0.71 ± 0.07 | 0.28 ± 0.01 | 0.26 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyriakou, S.; Potamiti, L.; Demosthenous, N.; Amery, T.; Stewart, K.; Winyard, P.G.; Franco, R.; Pappa, A.; Panayiotidis, M.I. A Naturally Derived Watercress Flower-Based Phenethyl Isothiocyanate-Enriched Extract Induces the Activation of Intrinsic Apoptosis via Subcellular Ultrastructural and Ca2+ Efflux Alterations in an In Vitro Model of Human Malignant Melanoma. Nutrients 2023, 15, 4044. https://doi.org/10.3390/nu15184044
Kyriakou S, Potamiti L, Demosthenous N, Amery T, Stewart K, Winyard PG, Franco R, Pappa A, Panayiotidis MI. A Naturally Derived Watercress Flower-Based Phenethyl Isothiocyanate-Enriched Extract Induces the Activation of Intrinsic Apoptosis via Subcellular Ultrastructural and Ca2+ Efflux Alterations in an In Vitro Model of Human Malignant Melanoma. Nutrients. 2023; 15(18):4044. https://doi.org/10.3390/nu15184044
Chicago/Turabian StyleKyriakou, Sotiris, Louiza Potamiti, Nikoletta Demosthenous, Tom Amery, Kyle Stewart, Paul G. Winyard, Rodrigo Franco, Aglaia Pappa, and Mihalis I. Panayiotidis. 2023. "A Naturally Derived Watercress Flower-Based Phenethyl Isothiocyanate-Enriched Extract Induces the Activation of Intrinsic Apoptosis via Subcellular Ultrastructural and Ca2+ Efflux Alterations in an In Vitro Model of Human Malignant Melanoma" Nutrients 15, no. 18: 4044. https://doi.org/10.3390/nu15184044