Different Dietary Protein and PUFA Interventions Alter the Fatty Acid Concentrations, but Not the Meat Quality, of Porcine Muscle


Abstract
:1. Introduction
2. Experimental Section
2.1. Experimental Design

{kind=link}
| Group 1 (HPD-SO) | Group 2 (HPD-LO) | Group 3 (RPD-SO) | Group 4 (RPD-LO) | Group 5 (CON) | |
|---|---|---|---|---|---|
| Number (n) | 8 | 8 | 8 | 8 | 8 |
| Feeding | High protein diet with sunflower seed oil | High protein diet with linseed oil | Reduced protein diet with sunflower seed oil | Reduced protein diet with linseed oil | Control |
| Diet Composition * | |||||
| Barley | 35.0 | 35.0 | 38.4 | 38.4 | 15.5 |
| Wheat | 26.5 | 26.5 | 36.0 | 36.0 | 26.5 |
| Soybean meal | 26.0 | 26.0 | 12.0 | 12.0 | 19.5 |
| Rye | - | - | - | - | 19.0 |
| Maize | - | - | - | - | 9.0 |
| Fiber mixture | 5.0 | 5.0 | 6.0 | 6.0 | 3.0 |
| Fattening Supplement | 3.0 | 3.0 | 3.0 | 3.0 | 7.5 |
| Linseed oil | - | 4.5 | - | 4.5 | - |
| Sunflower oil | 4.5 | - | 4.5 | - | - |
| Lysine | - | - | 0.1 | 0.1 | - |
| Group 1 (HPD-SO) | Group 2 (HPD-LO) | Group 3 (RPD-SO) | Group 4 (RPD-LO) | Group 5 (CON) | |
|---|---|---|---|---|---|
| Chemical Composition * | |||||
| Dry matter | 90.0 | 88.7 | 89.8 | 89.6 | 89.3 |
| Crude protein | 19.6 | 19.4 | 15.7 | 15.0 | 17.4 |
| Crude fat | 5.2 | 5.7 | 5.9 | 6.3 | 3.4 |
| Crude fiber | 4.7 | 4.6 | 5.3 | 5.5 | 4.2 |
| Crude ash | 4.9 | 4.9 | 4.8 | 5.0 | 4.8 |
| Starch | 36.9 | 36.2 | 40.0 | 40.7 | 40.1 |
| ME (MJ/kg) | 13.7 | 13.7 | 13.5 | 13.5 | 13.2 |
| Amino Acids * | |||||
| Lysine | 0.93 | 0.95 | 0.82 | 0.82 | 0.96 |
| Methionine | 0.26 | 0.27 | 0.22 | 0.22 | 0.24 |
| Cysteine | 0.31 | 0.31 | 0.26 | 0.27 | 0.30 |
| Threonine | 0.60 | 0.61 | 0.48 | 0.45 | 0.56 |
| Fatty Acids a | |||||
| 14:0 | 0.12 | 0.11 | 0.12 | 0.11 | 0.35 |
| 16:0 | 10.30 | 10.03 | 9.89 | 9.52 | 12.44 |
| 18:0 | 3.38 | 3.84 | 3.43 | 3.75 | 1.84 |
| 18:1cis-9 | 22.81 | 17.29 | 22.96 | 18.01 | 32.46 |
| 18:2n-6 | 52.94 | 29.43 | 51.69 | 31.77 | 39.07 |
| 18:3n-3 | 6.66 | 36.11 | 8.10 | 33.80 | 6.74 |
2.2. Methods
2.2.1. Meat Quality Parameters
2.2.2. Fatty Acid Analysis
2.2.3. Statistical Analysis
3. Results and Discussion
| Group 1 (HPD-SO) | Group 2 (HPD-LO) | Group 3 (RPD-SO) | Group 4 (RPD-LO) | Group 5 (CON) | Signific. | |
|---|---|---|---|---|---|---|
| LSMSEM | LSMSEM | LSMSEM | LSMSEM | LSMSEM | ||
| ( n = 8) | ( n = 8) | ( n = 8) | ( n = 8) | ( n = 8) | ||
| Start weight (kg) | 68.81.60 | 69.71.60 | 69.41.60 | 69.01.60 | 69.21.60 | 0.997 |
| Live weight at slaughter (kg) | 120.60.75 a | 121.40.75 a | 123.20.75 a | 120.60.75 a | 117.80.75 b | <0.001 |
| ADG (g) | 831.418.38 a | 835.218.30 a | 874.418.38 a | 840.618.38 a | 761.018.38 b | <0.001 |
| ADFI (g) | 3135.649.25 | 3195.649.25 | 3191.449.25 | 3195.049.25 | 3214.249.25 | 0.832 |
| Hot carcass weight, left (kg) | 49.00.58 a,b | 49.10.58 a,b | 50.50.58 a | 49.30.58 a,b | 47.70.58 b | 0.039 |
| Hot carcass weight, right (kg) | 47.90.38 a | 48.20.38 a | 48.90.38 a | 47.80.38 a | 46.40.38 b | <0.001 |
| Liver weight (kg) | 1.70.04 | 1.80.04 | 1.70.04 | 1.70.04 | 1.80.04 | 0.119 |
| Belly weight (kg) | 1.50.14 | 1.70.14 | 1.50.14 | 1.40.14 | 1.50.14 | 0.544 |
| Back fat (mm) | 18.41.03 | 19.31.03 | 18.21.03 | 19.21.03 | 20.11.03 | 0.710 |
| Muscle area (cm2) | 52.12.01 | 50.32.01 | 54.32.01 | 51.12.01 | 52.22.01 | 0.692 |
| Group 1 (HPD-SO) | Group 2 (HPD-LO) | Group 3 (RPD-SO) | Group 4 (RPD-LO) | Group 5 (CON) | Signific. | |
|---|---|---|---|---|---|---|
| LSMSEM | LSMSEM | LSMSEM | LSMSEM | LSMSEM | ||
| ( n = 8) | ( n = 8) | ( n = 8) | ( n = 8) | ( n = 8) | ||
| IMF (%) | 1.30.17 | 1.40.17 | 1.40.17 | 1.30.17 | 1.20.17 | 0.789 |
| pH (45 min) | 6.30.06 | 6.40.06 | 6.40.06 | 6.40.06 | 6.20.06 | 0.078 |
| pH (24 h) | 5.40.02 | 5.40.02 | 5.40.03 | 5.40.02 | 5.40.03 | 0.789 |
| Color L* | 47.70.56 | 48.30.56 | 47.10.56 | 48.50.56 | 47.60.56 | 0.380 |
| a* | 7.60.32 | 7.40.32 | 7.40.32 | 7.30.32 | 7.70.32 | 0.886 |
| b* | 1.60.23 | 1.60.23 | 1.20.23 | 1.40.23 | 1.10.23 | 0.440 |
| Drip loss (g/100 g) | 3.50.41 a | 2.60.41 b | 2.80.41 a | 3.00.41 a | 4.60.41 b | 0.012 |
| Cooking loss (g/100 g) | 23.60.80 | 21.00.80 | 21.20.80 | 21.90.80 | 23.80.80 | 0.440 |
| Shear force(kg/cm2) | 5.20.27 | 4.70.27 | 5.50.27 | 4.90.27 | 4.80.27 | 0.189 |
| Group 1 (HPD-SO) | Group 2 (HPD-LO) | Group 3 (RPD-SO) | Group 4 (RPD-LO) | Group 5 (CON) | Signific. | |
|---|---|---|---|---|---|---|
| LSMSEM | LSMSEM | LSMSEM | LSMSEM | LSMSEM | ||
| ( n = 8) | ( n = 8) | ( n = 8) | ( n = 8) | ( n = 8) | ||
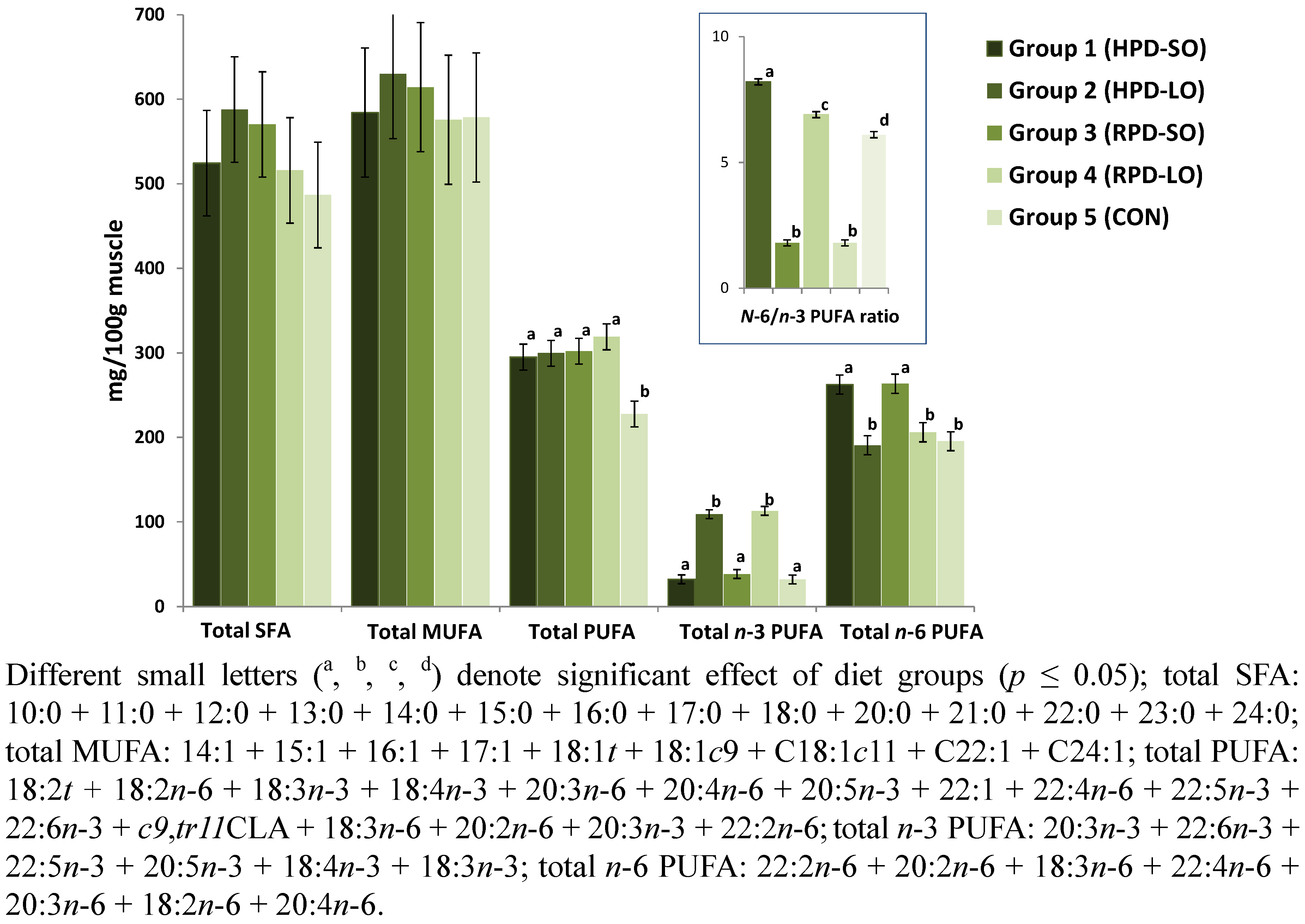
| Sum FA | 1403.4150.4 | 1517.3150.4 | 1486.3150.4 | 1410.6150.4 | 1292.9150.4 | 0.852 |
| 12:0 | 1.20.18 | 1.40.18 | 1.30.18 | 1.20.18 | 1.00.18 | 0.633 |
| 14:0 | 16.82.76 | 20.52.76 | 19.22.76 | 16.92.76 | 15.12.76 | 0.667 |
| 16:0 | 331.840.15 | 368.240.15 | 354.240.15 | 321.040.15 | 302.940.15 | 0.792 |
| 16:1 | 38.76.45 | 44.26.45 | 41.46.45 | 37.96.45 | 37.66.45 | 0.941 |
| 18:0 | 162.919.13 | 183.519.13 | 183.319.13 | 164.319.13 | 154.519.13 | 0.761 |
| 18:1cis-9 | 475.361.02 | 508.461.02 | 500.661.02 | 469.461.02 | 467.461.02 | 0.980 |
| 18:1cis-11 | 55.87.70 | 60.77.70 | 57.47.70 | 53.27.70 | 57.57.70 | 0.975 |
| 18:2n-6 | 201.310.70 a | 148.210.70 b | 203.910.70 a | 163.010.70 a,b | 139.310.70 b | <0.001 |
| 18:3n-3 | 13.14.05 a | 65.24.05 b | 16.84.05 a | 66.84.05 b | 10.34.05 a | <0.001 |
| 20:2n-6 | 5.10.40 a | 3.80.40 a,b | 5.70.40 a,c | 4.20.40 a,b | 3.30.40 b | <0.001 |
| 20:3n-6 | 6.30.22 a | 5.50.22 a,b | 6.40.22 a | 5.60.22 a,b | 7.00.22 a | <0.001 |
| 20:3n-3 | 2.50.63 a | 8.20.63 b | 3.30.63 a | 8.80.63 b | 1.80.63 a | <0.001 |
| 20:4n-6 | 43.41.01 a | 29.41.01 b | 41.41.01 a | 29.91.01 b | 39.91.01 a | <0.001 |
| 20:5n-3 | 4.40.54 a | 17.90.54 b | 5.10.54 a | 19.70.54 b | 6.30.54 a | <0.001 |
| 22:4n-6 | 4.80.14 a | 2.30.14 b | 4.60.14 a | 2.00.14 b | 4.30.14 a | <0.001 |
| 22:5n-3 | 8.50.29 a | 13.80.29 b | 9.40.29 a | 14.00.29 b | 8.9 0.29 a | <0.001 |
| 22:6n-3 | 3.70.23 a | 3.50.23 a | 3.60.23 a | 3.60.23 a | 4.70.23 b | 0.005 |

4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar]
- Corpet, D.E. Red meat and colon cancer: Should we become vegetarians, or can we make meat safer? Meat Sci. 2011, 89, 310–316. [Google Scholar] [CrossRef]
- Fretts, A.M.; Howard, B.V.; McKnight, B.; Duncan, G.E.; Beresford, S.A.A.; Mete, M.; Eilat-Adar, S.; Zhang, Y.; Siscovick, D.S. Associations of processed meat and unprocessed red meat intake with incident diabetes: The strong heart family study. Am. J. Clin. Nutr. 2012, 95, 752–758. [Google Scholar]
- World Cancer Research FundAmerican Institute for Cancer ResearchPolicy and Action for Cancer Prevention; WCRF and AICR: Washington, DC, USA, 2009.
- Wyness, L.; Weichselbaum, E.; O’Connor, A.; Williams, E.B.; Rilex, H.; Stanner, S. Red meat in the diet: An update. Nutr. Bull. 2010, 36, 34–77. [Google Scholar]
- Duran-Montge, P.; Realini, C.E.; Barroeta, A.C.; Lizardo, R.G.; Esteve-Garcia, E. De novo fatty acid synthesis and balance of fatty acids of pigs fed different fat sources. Livest. Sci. 2010, 132, 157–164. [Google Scholar] [CrossRef]
- Nuernberg, K.; Nuernberg, G.; Dannenberger, D. Effect of extruded linseed on growth and lipids of muscle and back fat in pigs. Fleischwirtschaft 2011, 91, 88–92. [Google Scholar]
- Sobotka, W.; Flis, M.; Antoszkiewicz, Z.; Lipinski, K.; Zdunczyk, Z. Effect of oat by-product antioxidants and vitamin E on the oxidative stability of pork from pigs fed diets supplemented with linseed oil. Arch. Amin. Nutr. 2012, 66, 27–38. [Google Scholar] [CrossRef]
- Alonso, A.; Campo, M.M.; Provincial, L.; Roncales, P.; Beltran, J.A. Effect of protein level in commercial diets on pork meat quality. Meat Sci. 2010, 85, 7–14. [Google Scholar]
- Calder, P.C.; Dangour, A.D.; Diekman, C.; Eilander, A.; Koletzko, B.; Meijer, G.W.; Mozaffarian, D.; Niinikoski, H.; Osendarp, S.J.; Pietinen, P.; et al. Essential fats for future health. Proceedings of the 9th Unilever Nutrition Symposium, 26-27 May 2010. Eur. J. Clin. Nutr. 2010, 64, S1–S13. [Google Scholar]
- Realini, C.E.; Duran-Montge, P.; Lizardo, R.; Gispert, M.; Oliver, M.A.; Esteve-Garcia, E. Effect of source of dietary fat on pig performance, carcass characteristics and carcass fat content, distribution and fatty acid composition. Meat Sci. 2010, 85, 606–612. [Google Scholar] [CrossRef]
- Juarez, M.; Dugan, M.E.R.; Aldai, N.; Aalhus, J.L.; Patience, J.F.; Zijlstra, R.T.; Beaulieu, A.D. Feeding co-extruded flaxseed to pigs: Effect of duration and feeding level on growth performance and backfat fatty acid composition of grower-finisher pigs. Meat Sci. 2010, 84, 578–584. [Google Scholar]
- Doran, O.; Moule, S.K.; Teye, G.A.; Whittington, F.M.; Hallett, K.G.; Wood, J.D. A reduced protein diet induces stearoyl-CoA desaturase protein expression in pig muscle but not in subcutaneous adipose tissue: relationship with intramuscular lipid formation. Br. J. Nutr. 2006, 95, 609–617. [Google Scholar] [CrossRef]
- Guo, X.; Tang, R.; Wang, W.; Liu, D.; Wang, K. Effects of dietary protein/carbohydrate ratio on fat deposition and gene expression of peroxisome proliferator activated receptor γ and heart fatty acid-binding protein of finishing pigs. Livest. Sci. 2011, 140, 111–116. [Google Scholar] [CrossRef]
- Guillevic, M.; Kouba, M.; Mourot, J. Effect of a linseed diet or sunflower diet on performances, fatty acid composition, lipogenic enzyme activity and stearoyl-CoA-desaturase activity in the pig. Livest. Sci. 2009, 124, 288–294. [Google Scholar] [CrossRef]
- Kuechenmeister, U.; Kuhn, G.; Ender, K. Seasonal effects on Ca2+ transport of sarcoplasmatic reticulum and meat quality of pigs with different malignant hyperthermia status. Meat Sci. 2000, 55, 239–245. [Google Scholar] [CrossRef]
- Dannenberger, D.; Nuernberg, K.; Nuernberg, G.; Ender, K. Carcass- and meat quality of pasture vs. concentrate fed German Simmental and German Holstein bulls. Arch. Tierz. 2006, 49, 315–328. [Google Scholar]
- Shen, X.; Dannenberger, D.; Nuernberg, K.; Nuernberg, G.; Zhao, R. Trans-18:1 and CLA isomers in rumen and duodenal digesta of bulls fed n-3 and n-6 PUFA-based diets. Lipids 2011, 46, 831–841. [Google Scholar] [CrossRef]
- Teye, G.A.; Sheard, P.R.; Whittington, F.M.; Nute, G.R.; Stewart, A.; Wood, J.D. Influence of dietary oils and protein level on pork quality. 1. Effects on muscle fatty acid composition, carcass, meat and eating quality. Meat Sci. 2006, 73, 157–165. [Google Scholar]
- Kouba, M.; Mourot, J. A review of nutritional effects on fat composition of animal products with special emphasis on n-3 polyunsaturated fatty acids. Biochimie 2011, 93, 13–17. [Google Scholar] [CrossRef]
- Raes, K.; de Smet, S.; Demeyer, D. Effect of dietary fatty acids on incorporation of long chain polyunsaturated fatty acids and conjugated linoleic acid in lamb, beef and pork meat: A review. Amin. Feed Sci. Technol. 2004, 113, 199–221. [Google Scholar]
- Cherfaoui, M.; Durand, D.; Bonnet, M.; Cassar-Malek, I.; Bauchart, D.; Thomas, A.; Gruffat, D. Expression of enzymes and transcription factors involved in the n-3 long chain PUFA biosynthesis in Limousin bull tissues. Lipids 2012, 47, 391–401. [Google Scholar] [CrossRef]
- German Nutrition Society (DGE), D-A-CH Referenzwerte für die Nährstoffzufuhr; Umschau-Verlag: Frankfurt am Main, Germany, 2012.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dannenberger, D.; Nuernberg, K.; Nuernberg, G.; Priepke, A. Different Dietary Protein and PUFA Interventions Alter the Fatty Acid Concentrations, but Not the Meat Quality, of Porcine Muscle. Nutrients 2012, 4, 1237-1246. https://doi.org/10.3390/nu4091237
Dannenberger D, Nuernberg K, Nuernberg G, Priepke A. Different Dietary Protein and PUFA Interventions Alter the Fatty Acid Concentrations, but Not the Meat Quality, of Porcine Muscle. Nutrients. 2012; 4(9):1237-1246. https://doi.org/10.3390/nu4091237
Chicago/Turabian StyleDannenberger, Dirk, Karin Nuernberg, Gerd Nuernberg, and Antje Priepke. 2012. "Different Dietary Protein and PUFA Interventions Alter the Fatty Acid Concentrations, but Not the Meat Quality, of Porcine Muscle" Nutrients 4, no. 9: 1237-1246. https://doi.org/10.3390/nu4091237
APA StyleDannenberger, D., Nuernberg, K., Nuernberg, G., & Priepke, A. (2012). Different Dietary Protein and PUFA Interventions Alter the Fatty Acid Concentrations, but Not the Meat Quality, of Porcine Muscle. Nutrients, 4(9), 1237-1246. https://doi.org/10.3390/nu4091237