Effects of ω3- and ω6-Polyunsaturated Fatty Acids on RANKL-Induced Osteoclast Differentiation of RAW264.7 Cells: A Comparative in Vitro Study


 , and
, and 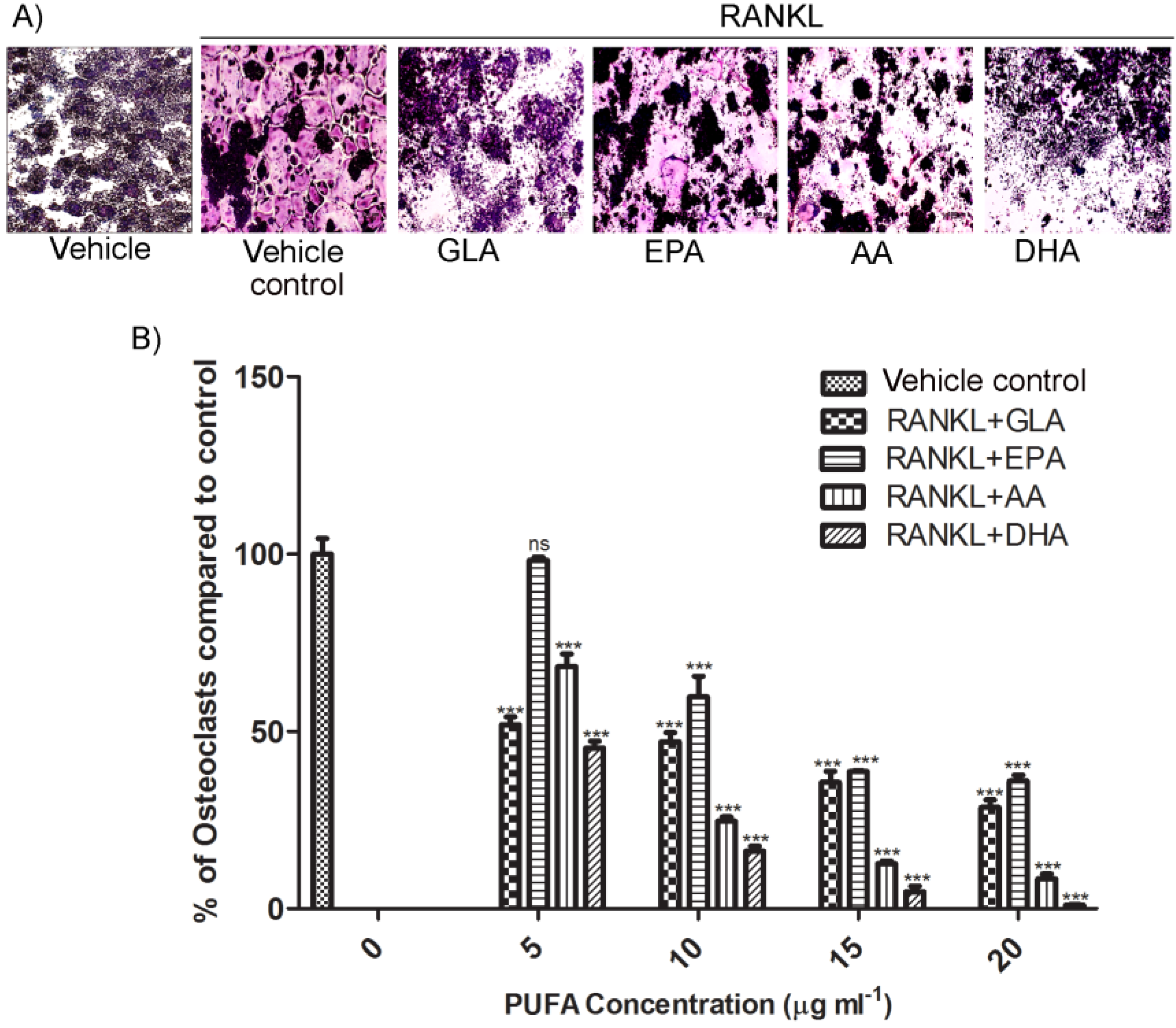{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Reagents and Materials
2.2. Cell Culture and Maintenance
2.3. Tartrate Resistant Acid Phosphatase (TRAP)-Positive Cell Staining
2.4. Fluorescent Microscopy
2.5. Scanning Electron Microscopy
2.6. Resorption Assay on Bone Mimetic Plates
2.7. Western Blot Analysis
2.8. cDNA Synthesis and RT-PCR Analysis
2.9. Statistics
3. Results
3.1. Effects of PUFAs on Osteoclastogenesis

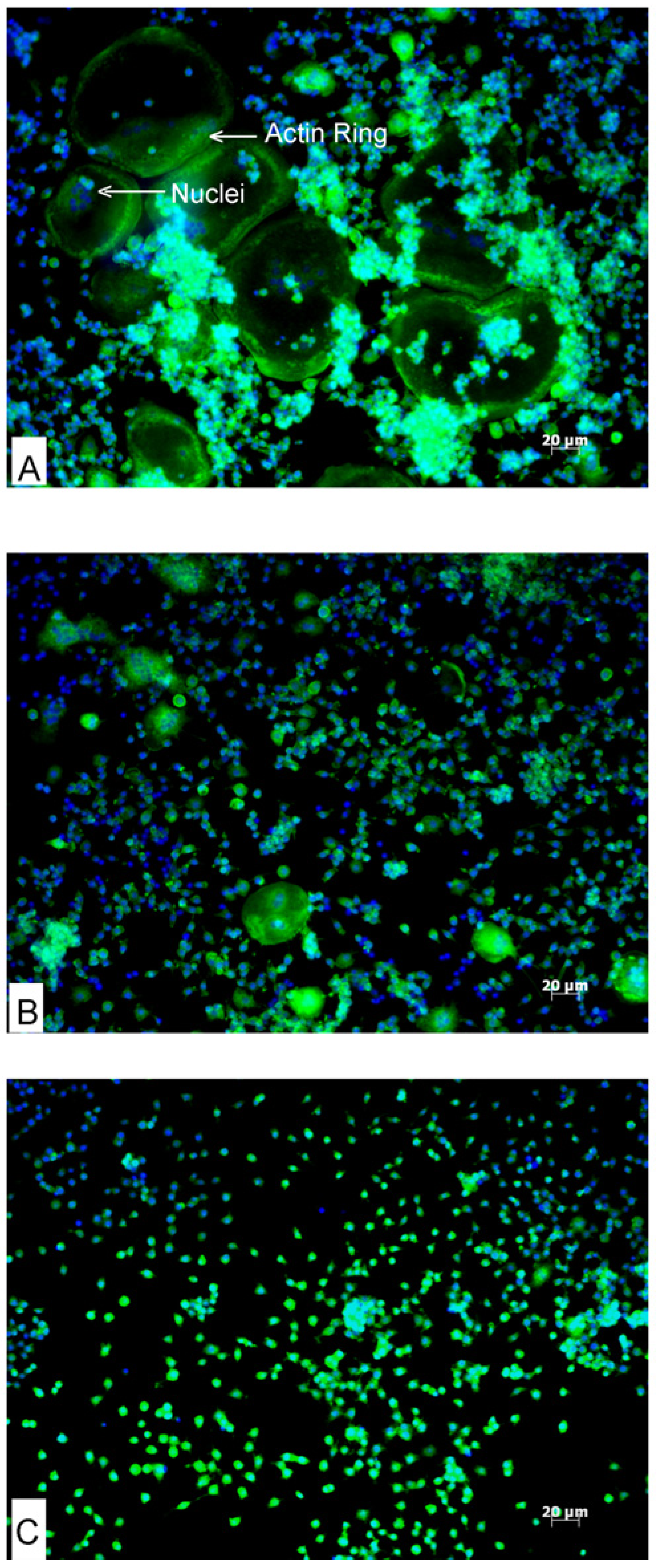
3.2. Effects of AA and DHA on Actin Ring Formation

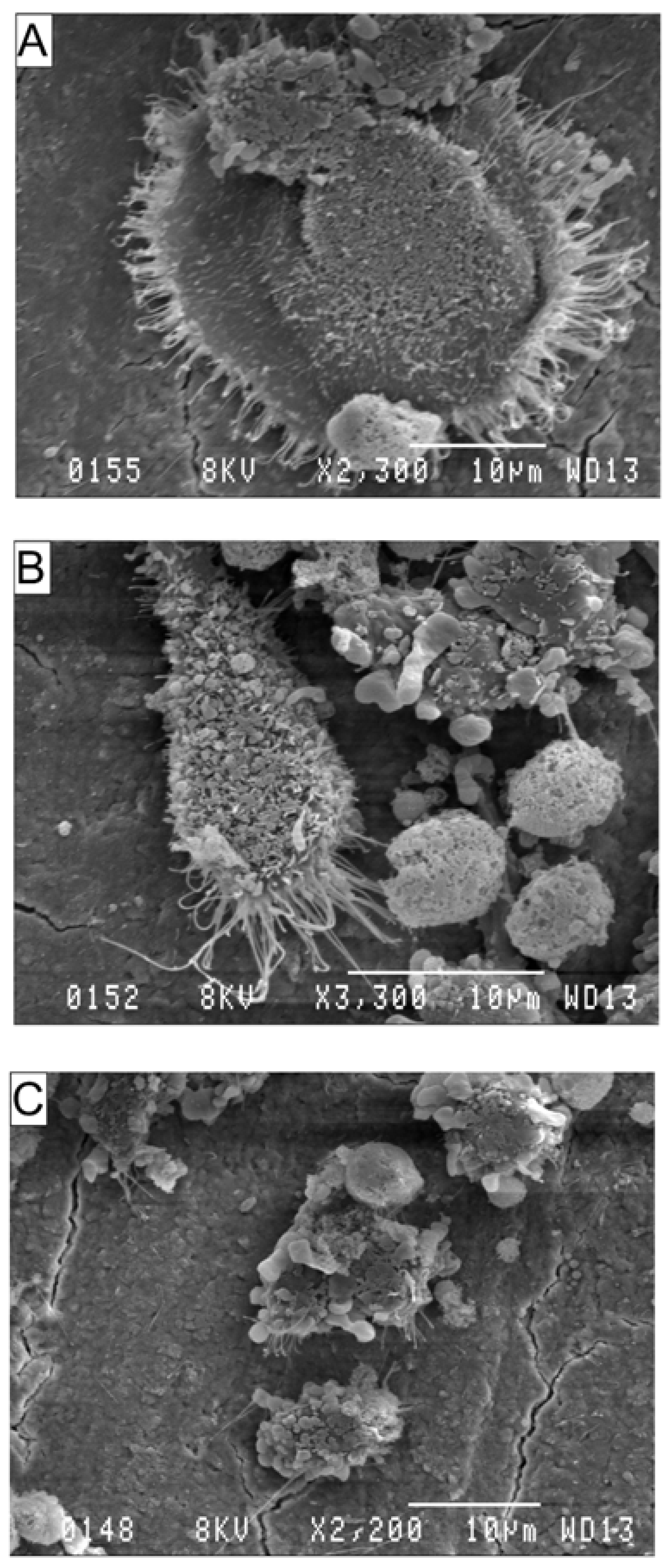
3.3. Effects of AA and DHA on the Ultrastructure of Osteoclasts

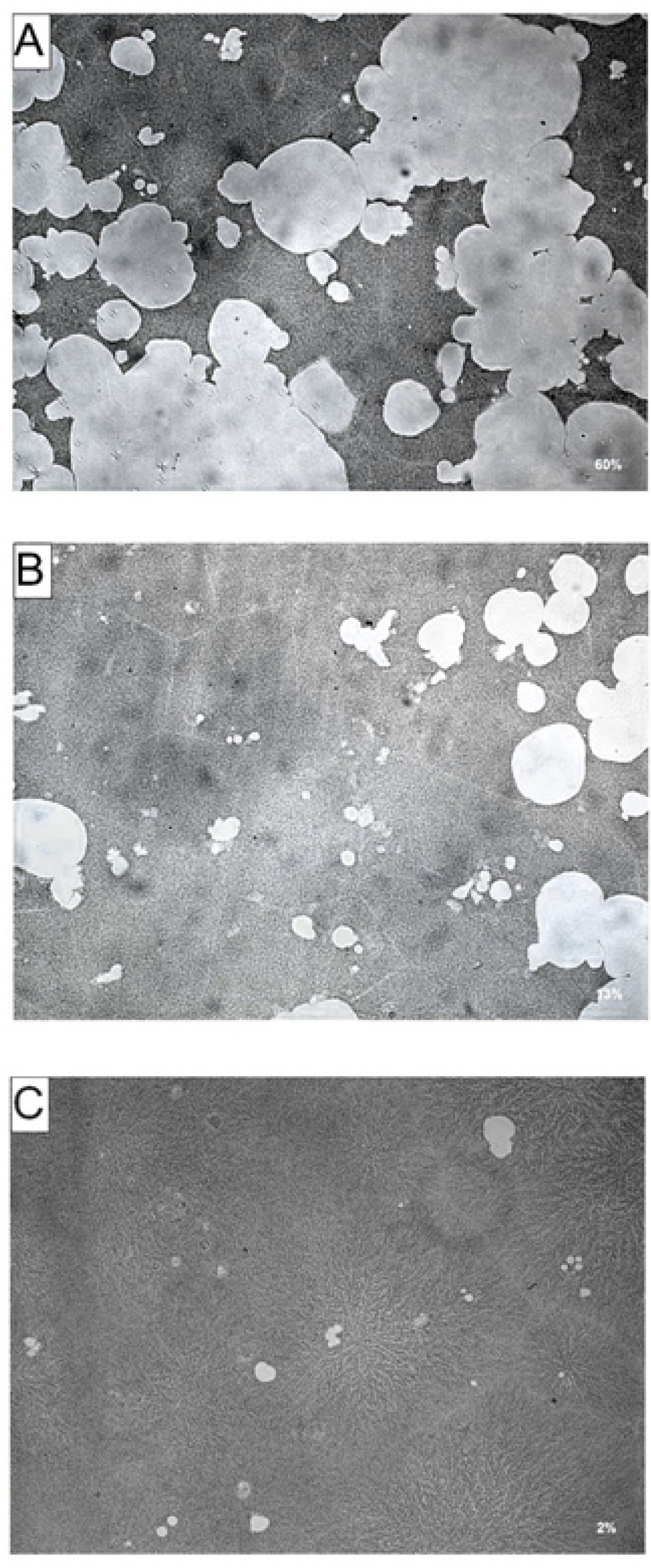
3.4. Effects of AA and DHA on Bone Resorption

3.5. Effects of AA and DHA on Osteoclast Specific Gene Expression
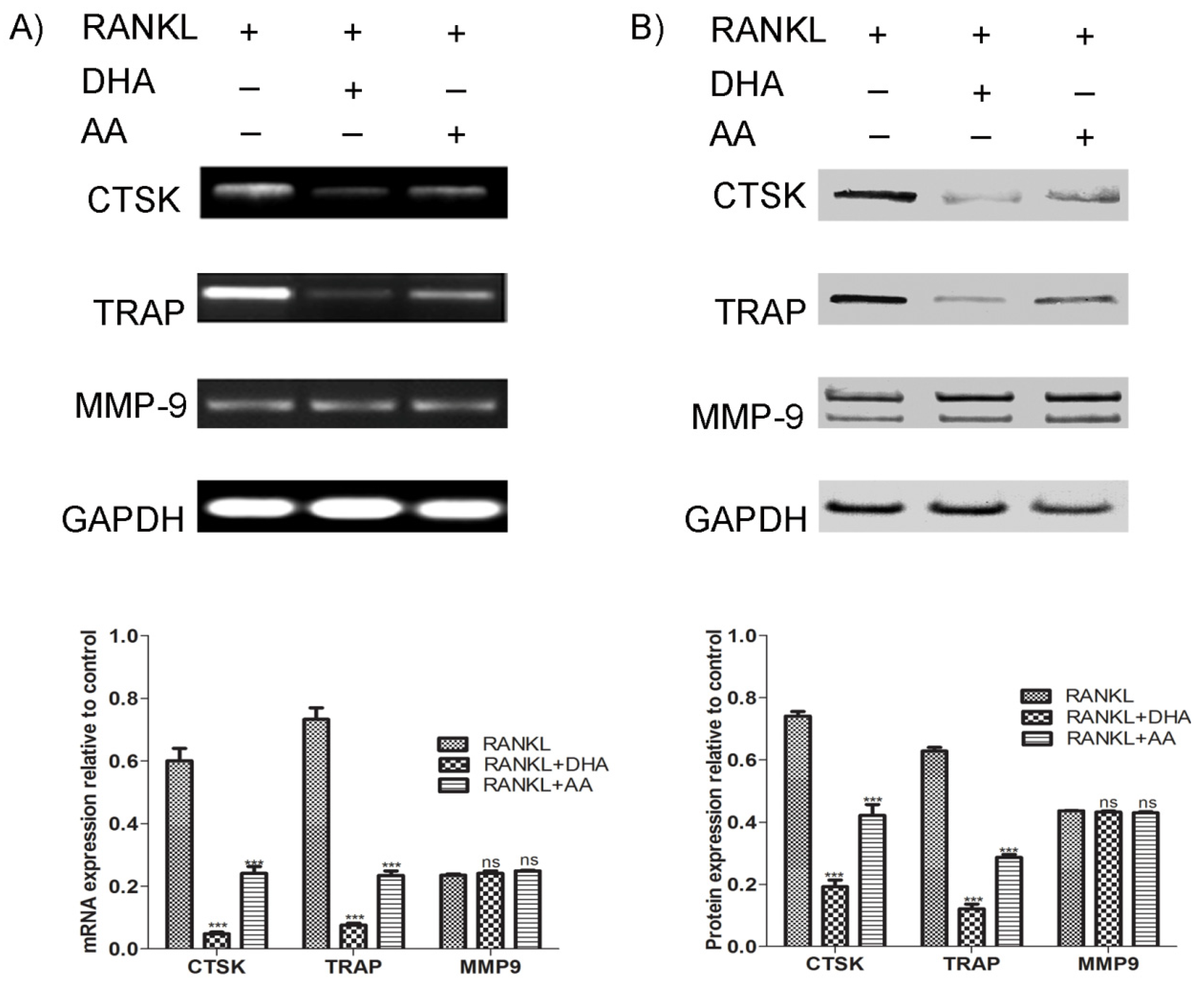
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nishida, C.; Uauy, R.; Kumanyika, S.; Shetty, P. The joint who/fao expert consultation on diet, nutrition and the prevention of chronic diseases: Process, product and policy implications. Public Health Nutr. 2004, 7, 245–250. [Google Scholar]
- Hadjidakis, D.J.; Androulakis, I.I. Bone remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef]
- Nakashima, T.; Takayanagi, H. New regulation mechanisms of osteoclast differentiation. Ann. N. Y. Acad. Sci. 2011, 1240, E13–E18. [Google Scholar] [CrossRef]
- Halleen, J.M.; Tiitinen, S.L.; Ylipahkala, H.; Fagerlund, K.M.; Vaananen, H.K. Tartrate-resistant acid phosphatase 5b (TRACP 5b) as a marker of bone resorption. Clin. Lab. 2006, 52, 499–509. [Google Scholar]
- Saftig, P.; Hunziker, E.; Everts, V.; Jones, S.; Boyde, A.; Wehmeyer, O.; Suter, A.; von Figura, K. Functions of cathepsin K in bone resorption. Lessons from cathepsin K deficient mice. Adv. Exp. Med. Biol. 2000, 477, 293–303. [Google Scholar]
- Mangano, K.M.; Sahni, S.; Kerstetter, J.E.; Kenny, A.M.; Hannan, M.T. Polyunsaturated fatty acids and their relation with bone and muscle health in adults. Curr. Osteoporos. Rep. 2013, 11, 203–212. [Google Scholar] [CrossRef]
- Napier, J.A. Plumbing the depths of pufa biosynthesis: A novel polyketide synthase-like pathway from marine organisms. Trends Plant Sci. 2002, 7, 51–54. [Google Scholar] [CrossRef]
- Kruger, M.C.; Horrobin, D.F. Calcium metabolism, osteoporsis and essential fatty acids: A review. Prog. Lipid Res. 1997, 36, 131–151. [Google Scholar] [CrossRef]
- Gogus, U.; Smith, C. n-3 Omega fatty acids: A review of current knowledge. Int. J. Food Sci. Technol. 2010, 45, 417–436. [Google Scholar] [CrossRef]
- Burdge, G.C. Metabolism of alpha-linolenic acid in humans. Prostaglandins Leukot. Essent. Fat. Acids 2006, 75, 161–168. [Google Scholar] [CrossRef]
- Calder, P.C. Fatty acids and inflammation: The cutting edge between food and pharma. Eur. J. Pharmacol. 2011, 668 (Suppl. 1), S50–S58. [Google Scholar] [CrossRef]
- Kris-Etherton, P.; Taylor, D.S.; Yu-Poth, S.; Huth, P.; Moriarty, K.; Fishell, V.; Hargrove, R.L.; Zhao, G.; Etherton, T.D. Polyunsaturated fatty acids in the food chain in the united states. Am. J. Clin. Nutr. 2000, 71, 179S–188S. [Google Scholar]
- Kruger, M.C.; Coetzee, M.; Haag, M.; Weiler, H. Long-chain polyunsaturated fatty acids: Selected mechanisms of action on bone. Prog. Lipid Res. 2010, 49, 438–449. [Google Scholar] [CrossRef]
- Grey, A. Fatty acids and bone. Clin. Rev. Bone Miner. Metab. 2009, 7, 210–215. [Google Scholar] [CrossRef]
- Collin-Osdoby, P.; Osdoby, P. Rankl-mediated osteoclast formation from murine raw 264.7 cells. Methods Mol. Biol. 2012, 816, 187–202. [Google Scholar] [CrossRef]
- Mouline, C.C.; Quincey, D.; Laugier, J.P.; Carle, G.F.; Bouler, J.M.; Rochet, N.; Scimeca, J.C. Osteoclastic differentiation of mouse and human monocytes in a plasma clot/biphasic calcium phosphate microparticles composite. Eur. Cell. Mater. 2010, 20, 379–392. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Gomez Candela, C.; Bermejo Lopez, L.M.; Loria Kohen, V. Importance of a balanced omega 6/omega 3 ratio for the maintenance of health: Nutritional recommendations. Nutr. Hosp. 2011, 26, 323–329. [Google Scholar]
- Reinwald, S.; Li, Y.; Moriguchi, T.; Salem, N., Jr.; Watkins, B.A. Repletion with (n-3) fatty acids reverses bone structural deficits in (n-3)-deficient rats. J. Nutr. 2004, 134, 388–394. [Google Scholar]
- Watkins, B.A.; Li, Y.; Seifert, M.F. Dietary ratio of n-6/n-3 pufas and docosahexaenoic acid: Actions on bone mineral and serum biomarkers in ovariectomized rats. J. Nutr. Biochem. 2006, 17, 282–289. [Google Scholar] [CrossRef]
- Novack, D.V.; Teitelbaum, S.L. The osteoclast: Friend or foe? Annu. Rev. Pathol. 2008, 3, 457–484. [Google Scholar] [CrossRef]
- Nunez, E.A. Free fatty acids as modulators of the steroid hormone message. Prostaglandins Leukot. Essent. Fat. Acids 1993, 48, 63–70. [Google Scholar]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef]
- Rahman, M.M.; Bhattacharya, A.; Fernandes, G. Docosahexaenoic acid is more potent inhibitor of osteoclast differentiation in raw 264.7 cells than eicosapentaenoic acid. J. Cell. Physiol. 2008, 214, 201–209. [Google Scholar] [CrossRef]
- Wilson, S.R.; Peters, C.; Saftig, P.; Bromme, D. Cathepsin K activity-dependent regulation of osteoclast actin ring formation and bone resorption. J. Biol. Chem. 2009, 284, 2584–2592. [Google Scholar] [CrossRef]
- Hollberg, K.; Hultenby, K.; Hayman, A.; Cox, T.; Andersson, G. Osteoclasts from mice deficient in tartrate-resistant acid phosphatase have altered ruffled borders and disturbed intracellular vesicular transport. Exp. Cell. Res. 2002, 279, 227–238. [Google Scholar] [CrossRef]
- Gelb, B.D.; Shi, G.P.; Chapman, H.A.; Desnick, R.J. Pycnodysostosis, a lysosomal disease caused by cathepsin K deficiency. Science 1996, 273, 1236–1238. [Google Scholar]
- Kumta, S.M.; Huang, L.; Cheng, Y.Y.; Chow, L.T.; Lee, K.M.; Zheng, M.H. Expression of VEGF and MMP-9 in giant cell tumor of bone and other osteolytic lesions. Life Sci. 2003, 73, 1427–1436. [Google Scholar] [CrossRef]
- Vu, T.H.; Shipley, J.M.; Bergers, G.; Berger, J.E.; Helms, J.A.; Hanahan, D.; Shapiro, S.D.; Senior, R.M.; Werb, Z. MMP-9/gelatinase B is a key regulator of growth plate angiogenesis and apoptosis of hypertrophic chondrocytes. Cell 1998, 93, 411–422. [Google Scholar] [CrossRef]
- Hill, P.A.; Murphy, G.; Docherty, A.J.; Hembry, R.M.; Millican, T.A.; Reynolds, J.J.; Meikle, M.C. The effects of selective inhibitors of matrix metalloproteinases (MMPs) on bone resorption and the identification of MMPs and TIMP-1 in isolated osteoclasts. J. Cell. Sci. 1994, 107, 3055–3064. [Google Scholar]
- Everts, V.; Korper, W.; Jansen, D.C.; Steinfort, J.; Lammerse, I.; Heera, S.; Docherty, A.J.; Beertsen, W. Functional heterogeneity of osteoclasts: Matrix metalloproteinases participate in osteoclastic resorption of calvarial bone but not in resorption of long bone. FASEB J. 1999, 13, 1219–1230. [Google Scholar]
- Rahman, M.M.; Bhattacharya, A.; Fernandes, G. Conjugated linoleic acid inhibits osteoclast differentiation of raw 264.7 cells by modulating rankl signaling. J. Lipid Res. 2006, 47, 1739–1748. [Google Scholar] [CrossRef]
- Sun, D.; Krishnan, A.; Zaman, K.; Lawrence, R.; Bhattacharya, A.; Fernandes, G. Dietary n-3 fatty acids decrease osteoclastogenesis and loss of bone mass in ovariectomized mice. J. Bone Miner. Res. 2003, 18, 1206–1216. [Google Scholar] [CrossRef]
- Hutchins-Wiese, H.L.; Picho, K.; Watkins, B.A.; Li, Y.; Tannenbaum, S.; Claffey, K.; Kenny, A.M. High-dose eicosapentaenoic acid and docosahexaenoic acid supplementation reduces bone resorption in postmenopausal breast cancer survivors on aromatase inhibitors: A pilot study. Nutr. Cancer 2014, 66, 68–76. [Google Scholar] [CrossRef]
- Farina, E.K.; Kiel, D.P.; Roubenoff, R.; Schaefer, E.J.; Cupples, L.A.; Tucker, K.L. Dietary intakes of arachidonic acid and alpha-linolenic acid are associated with reduced risk of hip fracture in older adults. J. Nutr. 2011, 141, 1146–1153. [Google Scholar] [CrossRef]
- Rahman, M.M.; Bhattacharya, A.; Banu, J.; Kang, J.X.; Fernandes, G. Endogenous n-3 fatty acids protect ovariectomy induced bone loss by attenuating osteoclastogenesis. J. Cell. Mol. Med. 2009, 13, 1833–1844. [Google Scholar] [CrossRef]
- Akiyama, M.; Nakahama, K.-I.; Morita, I. Impact of docosahexaenoic acid on gene expression during osteoclastogenesis in vitro—A comprehensive analysis. Nutrients 2013, 5, 3151–3162. [Google Scholar] [CrossRef]
- Das, U.N. Essential fatty acids: Biochemistry, physiology and pathology. Biotechnol. J. 2006, 1, 420–439. [Google Scholar] [CrossRef]
- Wahli, W.; Michalik, L. Ppars at the crossroads of lipid signaling and inflammation. Trends Endocrinol. Metab. 2012, 23, 351–363. [Google Scholar] [CrossRef]
- Marks, F. Arachidonic acid and companions: An abundant source of biological signals. In Prostaglandins, Leukotrienes and Other Eicosanoids; Wiley-VCH Verlag GmbH: Weinheim, Germany, 2007; pp. 1–46. [Google Scholar]
- Serhan, C.N.; Arita, M.; Hong, S.; Gotlinger, K. Resolvins, docosatrienes, and neuroprotectins, novel omega-3-derived mediators, and their endogenous aspirin-triggered epimers. Lipids 2004, 39, 1125–1132. [Google Scholar] [CrossRef]
- Herrera, B.S.; Ohira, T.; Gao, L.; Omori, K.; Yang, R.; Zhu, M.; Muscara, M.N.; Serhan, C.N.; van Dyke, T.E.; Gyurko, R. An endogenous regulator of inflammation, resolvin E1, modulates osteoclast differentiation and bone resorption. Br. J. Pharmacol. 2008, 155, 1214–1223. [Google Scholar] [CrossRef]
- Zhu, M.; van Dyke, T.E.; Gyurko, R. Resolvin E1 regulates osteoclast fusion via DC-STAMP and NFATc1. FASEB J. 2013, 27, 3344–3353. [Google Scholar] [CrossRef]
- Yuan, J.; Akiyama, M.; Nakahama, K.; Sato, T.; Uematsu, H.; Morita, I. The effects of polyunsaturated fatty acids and their metabolites on osteoclastogenesis in vitro. Prostaglandins Other Lipid Mediat. 2010, 92, 85–90. [Google Scholar]
- Krieger, N.S.; Parker, W.R.; Alexander, K.M.; Bushinsky, D.A. Prostaglandins regulate acid-induced cell-mediated bone resorption. Am. J. Physiol. Ren. Physiol. 2000, 279, F1077–F1082. [Google Scholar]
- Watkins, B.A.; Li, Y.; Allen, K.G.; Hoffmann, W.E.; Seifert, M.F. Dietary ratio of (n-6)/(n-3) polyunsaturated fatty acids alters the fatty acid composition of bone compartments and biomarkers of bone formation in rats. J. Nutr. 2000, 130, 2274–2284. [Google Scholar]
- Li, G.; Wu, P.; Xu, Y.; Yu, Y.; Sun, L.; Zhu, L.; Ye, D. The effect of lipoxin a4 on the interaction between macrophage and osteoblast: Possible role in the treatment of aseptic loosening. BMC Musculoskelet. Disord. 2009, 10, 57. [Google Scholar] [CrossRef]
- Nilforoushan, D.; Gramoun, A.; Glogauer, M.; Manolson, M.F. Nitric oxide enhances osteoclastogenesis possibly by mediating cell fusion. Nitric Oxide 2009, 21, 27–36. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Boeyens, J.C.A.; Deepak, V.; Chua, W.-H.; Kruger, M.C.; Joubert, A.M.; Coetzee, M. Effects of ω3- and ω6-Polyunsaturated Fatty Acids on RANKL-Induced Osteoclast Differentiation of RAW264.7 Cells: A Comparative in Vitro Study. Nutrients 2014, 6, 2584-2601. https://doi.org/10.3390/nu6072584
Boeyens JCA, Deepak V, Chua W-H, Kruger MC, Joubert AM, Coetzee M. Effects of ω3- and ω6-Polyunsaturated Fatty Acids on RANKL-Induced Osteoclast Differentiation of RAW264.7 Cells: A Comparative in Vitro Study. Nutrients. 2014; 6(7):2584-2601. https://doi.org/10.3390/nu6072584
Chicago/Turabian StyleBoeyens, Jan C. A., Vishwa Deepak, Wei-Hang Chua, Marlena C. Kruger, Annie M. Joubert, and Magdalena Coetzee. 2014. "Effects of ω3- and ω6-Polyunsaturated Fatty Acids on RANKL-Induced Osteoclast Differentiation of RAW264.7 Cells: A Comparative in Vitro Study" Nutrients 6, no. 7: 2584-2601. https://doi.org/10.3390/nu6072584
APA StyleBoeyens, J. C. A., Deepak, V., Chua, W.-H., Kruger, M. C., Joubert, A. M., & Coetzee, M. (2014). Effects of ω3- and ω6-Polyunsaturated Fatty Acids on RANKL-Induced Osteoclast Differentiation of RAW264.7 Cells: A Comparative in Vitro Study. Nutrients, 6(7), 2584-2601. https://doi.org/10.3390/nu6072584