Egg Intake during Carbohydrate Restriction Alters Peripheral Blood Mononuclear Cell Inflammation and Cholesterol Homeostasis in Metabolic Syndrome

 ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Study Design and Dietary Intervention
2.2. Blood Collection and Body Weight
2.3. Peripheral Blood Mononuclear Cell Isolation
2.4. Quantitative Real-Time RT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| ABCA1 | 5′-TTTCTCAGACAACACTTGACCAAGTA-3′ | 5′-GGTTTTTGTGTAATGAGAGGTCTTTTAA-3′ |
| ABCG1 | 5′-CGGAGGGCAGCTGTGAAC-3′ | 5′-GGGTCCTTCAGGAACCGAAT-3′ |
| HMGCR | 5′-CCCAGTTGTGCGTCTTCCA-3′ | 5′-TTCGAGCCAGGCTTTCACTT-3′ |
| IL-1β | 5′-ACGATGCACCTGTACGATCACT-3′ | 5′-CACCAAGCTTTTTTGCTGTGAGT-3′ |
| IL-6 | 5′-GCTGCAGGCACAGAACCA-3′ | 5′-GCTGCGCAGAATGAGATGAG-3′ |
| LDLR | 5′-ACTGGGTTGACTCCAAACTTCAC-3′ | 5′-GGTTGCCCCCGTTGACA-3′ |
| TLR4 | 5′-GCAGGTGCTGGATTTATC-3′ | 5′-GTAGAGAGGTGGCTTAGG-3′ |
| TNFα | 5′-GGGACCTCTCTCTAATCA-3′ | 5′-CTACAACATGGGCTACAG-3′ |
| 18S RNA | 5′-CGGCTACCACATCCAAGGAA-3′ | 5′-GCTGGAATTACCGCGGCT-3′ |
2.5. NF-κB p65 DNA Binding Activity
2.6. PBMC Stimulation Assays
2.7. Quantification of PBMC Cholesterol Content
2.8. Quantification of PBMC ABCA1 Protein
2.9. Lipid Raft Staining, Microscopy, and Quantification
2.10. Statistical Analysis
3. Results
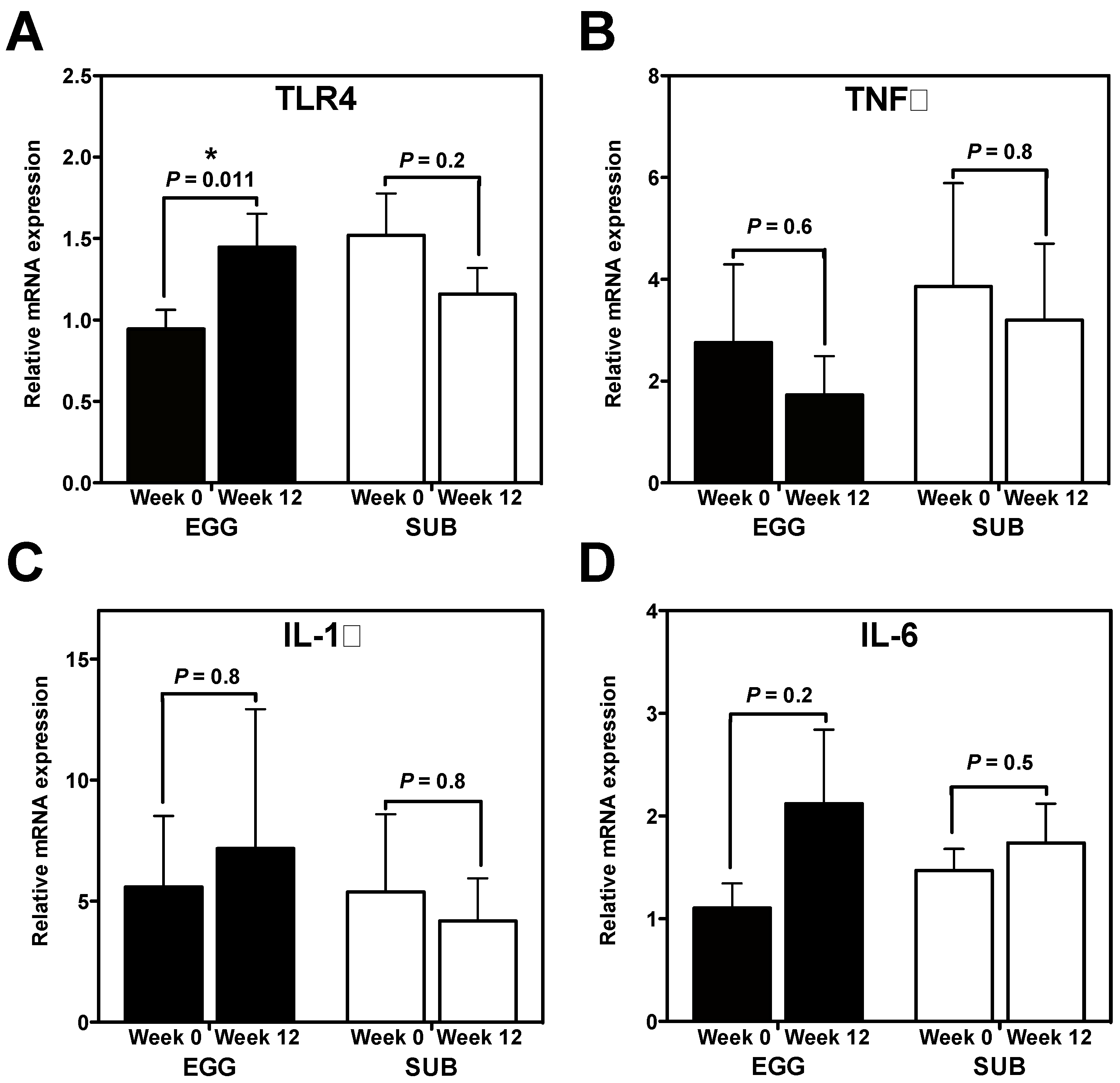
3.1. Effects of Egg Intake during Moderate Carbohydrate Restriction on Inflammatory Gene Expression

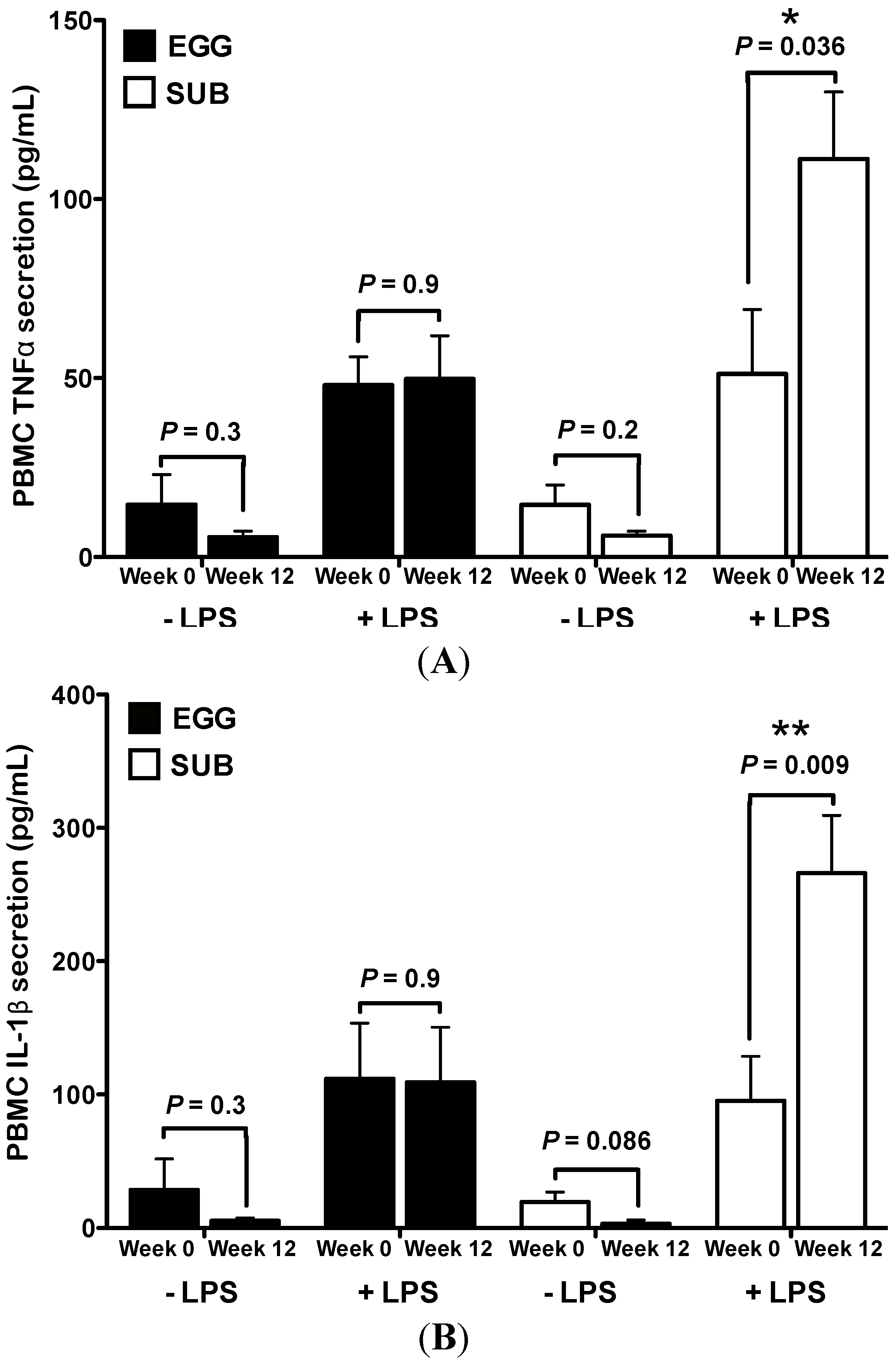
3.2. Inflammatory Challenge

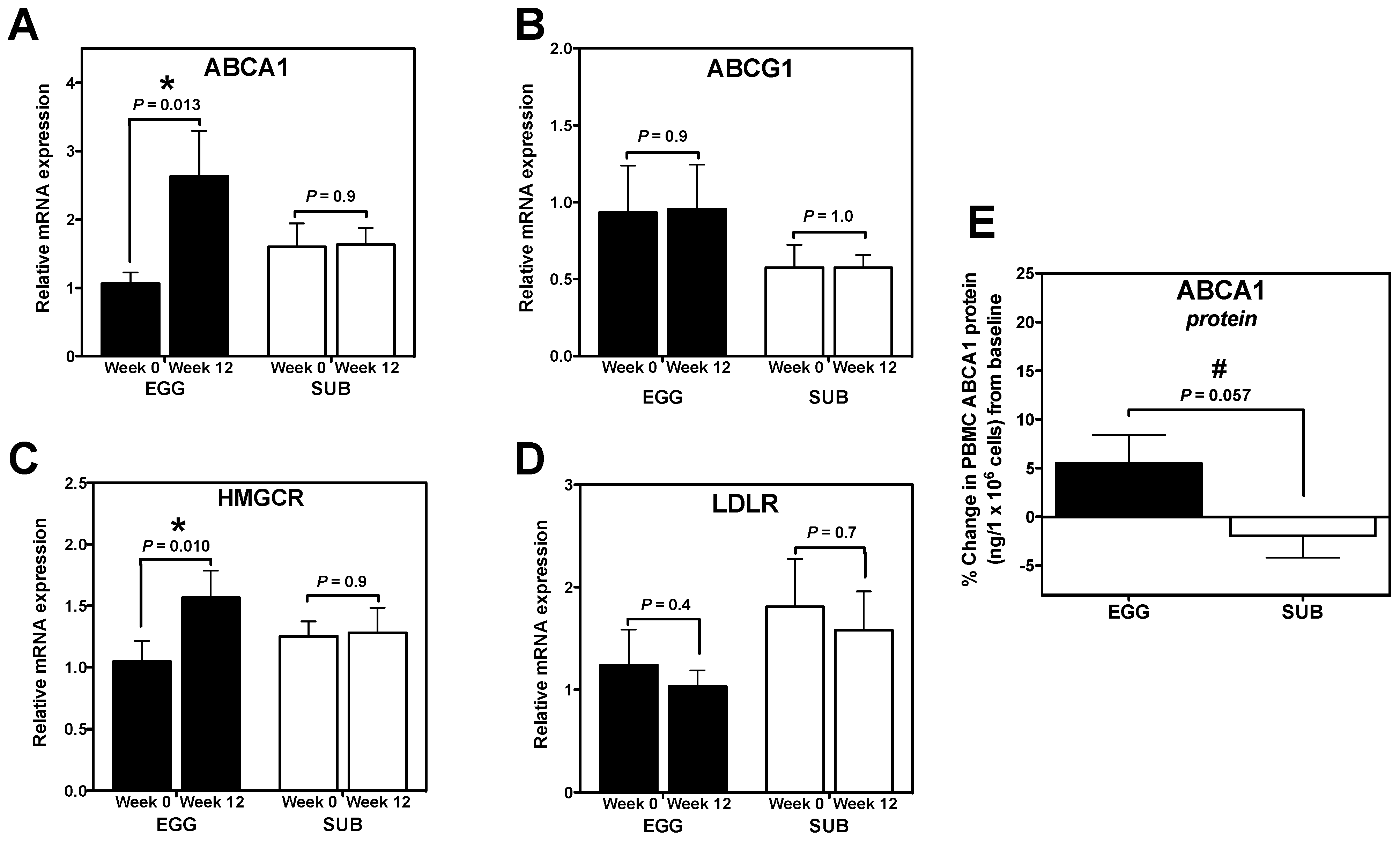
3.3. Cholesterol Gene Expression

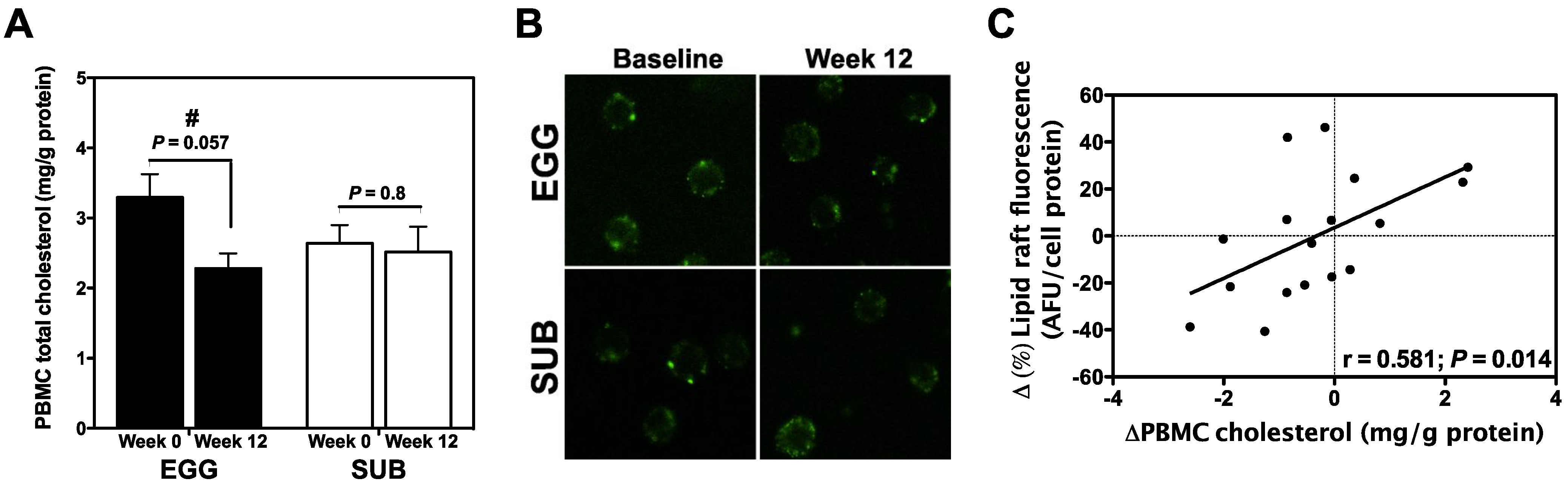
3.4. Effects of Egg Intake on PBMC Cholesterol and Lipid Raft Content

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Berg, A.H.; Scherer, P.E. Adipose tissue, inflammation, and cardiovascular disease. Circ. Res. 2005, 96, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Ndumele, C.E.; Nasir, K.; Conceicao, R.D.; Carvalho, J.A.; Blumenthal, R.S.; Santos, R.D. Hepatic steatosis, obesity, and the metabolic syndrome are independently and additively associated with increased systemic inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1927–1932. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M. Inflammation, metabolic syndrome, and diet responsiveness. Circulation 2003, 108, 126–128. [Google Scholar] [CrossRef] [PubMed]
- Haffner, S.M. The metabolic syndrome: Inflammation, diabetes mellitus, and cardiovascular disease. Am. J. Cardiol. 2006, 97, 3A–11A. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.; Tang, L.; Wang, Y.; Charnigo, R.; de Villiers, W.; Eckhardt, E. T-lymphocyte responses to intestinally absorbed antigens can contribute to adipose tissue inflammation and glucose intolerance during high fat feeding. PLoS One 2010, 5, e13951. [Google Scholar] [CrossRef] [PubMed]
- Winer, S.; Chan, Y.; Paltser, G.; Truong, D.; Tsui, H.; Bahrami, J.; Dorfman, R.; Wang, Y.; Zielenski, J.; Mastronardi, F.; et al. Normalization of obesity-associated insulin resistance through immunotherapy. Nat. Med. 2009, 15, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.A.; Sheridan, P.A.; Beck, M.A. Diet-induced obesity in mice reduces the maintenance of influenza-specific cd8+ memory t cells. J. Nutr. 2010, 140, 1691–1697. [Google Scholar] [CrossRef] [PubMed]
- Sheu, W.H.; Chang, T.M.; Lee, W.J.; Ou, H.C.; Wu, C.M.; Tseng, L.N.; Lang, H.F.; Wu, C.S.; Wan, C.J.; Lee, I.T. Effect of weight loss on proinflammatory state of mononuclear cells in obese women. Obesity 2008, 16, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, H.; Aljada, A.; Hofmeyer, D.; Syed, T.; Mohanty, P.; Dandona, P. Circulating mononuclear cells in the obese are in a proinflammatory state. Circulation 2004, 110, 1564–1571. [Google Scholar] [CrossRef] [PubMed]
- Strissel, K.J.; DeFuria, J.; Shaul, M.E.; Bennett, G.; Greenberg, A.S.; Obin, M.S. T-cell recruitment and th1 polarization in adipose tissue during diet-induced obesity in c57bl/6 mice. Obesity 2010, 18, 1918–1925. [Google Scholar] [CrossRef] [PubMed]
- Matarese, G.; Procaccini, C.; de Rosa, V.; Horvath, T.L.; la Cava, A. Regulatory t cells in obesity: The leptin connection. Trends Mol. Med. 2010, 16, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Blesso, C.N.; Andersen, C.J.; Barona, J.; Volk, B.; Volek, J.S.; Fernandez, M.L. Effects of carbohydrate restriction and dietary cholesterol provided by eggs on clinical risk factors in metabolic syndrome. J. Clin. Lipidol. 2013, 7, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Ratliff, J.C.; Mutungi, G.; Puglisi, M.J.; Volek, J.S.; Fernandez, M.L. Eggs modulate the inflammatory response to carbohydrate restricted diets in overweight men. Nutr. Metab. 2008, 5, 6. [Google Scholar] [CrossRef]
- Forsythe, C.E.; Phinney, S.D.; Fernandez, M.L.; Quann, E.E.; Wood, R.J.; Bibus, D.M.; Kraemer, W.J.; Feinman, R.D.; Volek, J.S. Comparison of low fat and low carbohydrate diets on circulating fatty acid composition and markers of inflammation. Lipids 2008, 43, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Teno, C.; Perez-Martinez, P.; Delgado-Lista, J.; Yubero-Serrano, E.M.; Garcia-Rios, A.; Marin, C.; Gomez, P.; Jimenez-Gomez, Y.; Camargo, A.; Rodriguez-Cantalejo, F.; et al. Dietary fat modifies the postprandial inflammatory state in subjects with metabolic syndrome: The lipgene study. Mol. Nutr. Food Res. 2012, 56, 854–865. [Google Scholar] [CrossRef] [PubMed]
- Kolehmainen, M.; Mykkanen, O.; Kirjavainen, P.V.; Leppanen, T.; Moilanen, E.; Adriaens, M.; Laaksonen, D.E.; Hallikainen, M.; Puupponen-Pimia, R.; Pulkkinen, L.; et al. Bilberries reduce low-grade inflammation in individuals with features of metabolic syndrome. Mol. Nutr. Food Res. 2012, 56, 1501–1510. [Google Scholar]
- Bensinger, S.J.; Bradley, M.N.; Joseph, S.B.; Zelcer, N.; Janssen, E.M.; Hausner, M.A.; Shih, R.; Parks, J.S.; Edwards, P.A.; Jamieson, B.D.; et al. Lxr signaling couples sterol metabolism to proliferation in the acquired immune response. Cell 2008, 134, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Yvan-Charvet, L.; Welch, C.; Pagler, T.A.; Ranalletta, M.; Lamkanfi, M.; Han, S.; Ishibashi, M.; Li, R.; Wang, N.; Tall, A.R. Increased inflammatory gene expression in abc transporter-deficient macrophages: Free cholesterol accumulation, increased signaling via toll-like receptors, and neutrophil infiltration of atherosclerotic lesions. Circulation 2008, 118, 1837–1847. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Westcott, M.M.; Bi, X.; Liu, M.; Gowdy, K.M.; Seo, J.; Cao, Q.; Gebre, A.K.; Fessler, M.B.; Hiltbold, E.M.; et al. Myeloid cell-specific abca1 deletion protects mice from bacterial infection. Circ. Res. 2012, 111, 1398–1409. [Google Scholar] [CrossRef] [PubMed]
- Surls, J.; Nazarov-Stoica, C.; Kehl, M.; Olsen, C.; Casares, S.; Brumeanu, T.D. Increased membrane cholesterol in lymphocytes diverts t-cells toward an inflammatory response. PLoS One 2012, 7, e38733. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Lee, J.Y.; Timmins, J.M.; Brown, J.M.; Boudyguina, E.; Mulya, A.; Gebre, A.K.; Willingham, M.C.; Hiltbold, E.M.; Mishra, N.; et al. Increased cellular free cholesterol in macrophage-specific abca1 knock-out mice enhances pro-inflammatory response of macrophages. J. Biol. Chem. 2008, 283, 22930–22941. [Google Scholar] [CrossRef] [PubMed]
- Out, R.; Hoekstra, M.; Hildebrand, R.B.; Kruit, J.K.; Meurs, I.; Li, Z.; Kuipers, F.; van Berkel, T.J.; van Eck, M. Macrophage abcg1 deletion disrupts lipid homeostasis in alveolar macrophages and moderately influences atherosclerotic lesion development in LDL receptor-deficient mice. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2295–2300. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Pennings, M.; Hildebrand, R.B.; Ye, D.; Calpe-Berdiel, L.; Out, R.; Kjerrulf, M.; Hurt-Camejo, E.; Groen, A.K.; Hoekstra, M.; et al. Enhanced foam cell formation, atherosclerotic lesion development, and inflammation by combined deletion of ABCA1 and SR-BI in bone marrow-derived cells in LDL receptor knockout mice on western-type diet. Circ. Res. 2010, 107, e20–e31. [Google Scholar] [CrossRef] [PubMed]
- Brunham, L.R.; Singaraja, R.R.; Duong, M.; Timmins, J.M.; Fievet, C.; Bissada, N.; Kang, M.H.; Samra, A.; Fruchart, J.C.; McManus, B.; et al. Tissue-specific roles of ABCA1 influence susceptibility to atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.J.; Gebre, A.K.; Parks, J.S.; Hedrick, C.C. Atp-binding cassette transporter G1 negatively regulates thymocyte and peripheral lymphocyte proliferation. J. Immunol. 2010, 184, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Owen, J.S.; Wilson, M.D.; Li, H.; Griffiths, G.L.; Thomas, M.J.; Hiltbold, E.M.; Fessler, M.B.; Parks, J.S. Macrophage ABCA1 reduces MyD88-dependent toll-like receptor trafficking to lipid rafts by reduction of lipid raft cholesterol. J. Lipid Res. 2010, 51, 3196–3206. [Google Scholar] [CrossRef] [PubMed]
- Yaqoob, P. The nutritional significance of lipid rafts. Ann. Rev. Nutr. 2009, 29, 257–282. [Google Scholar] [CrossRef]
- Van Eck, M.; Bos, I.S.; Kaminski, W.E.; Orso, E.; Rothe, G.; Twisk, J.; Bottcher, A.; van Amersfoort, E.S.; Christiansen-Weber, T.A.; Fung-Leung, W.P.; et al. Leukocyte ABCA1 controls susceptibility to atherosclerosis and macrophage recruitment into tissues. Proc. Natl. Acad. Sci. USA 2002, 99, 6298–6303. [Google Scholar] [CrossRef] [PubMed]
- Levine, D.M.; Parker, T.S.; Donnelly, T.M.; Walsh, A.; Rubin, A.L. In vivo protection against endotoxin by plasma high density lipoprotein. Proc. Natl. Acad. Sci. USA 1993, 90, 12040–12044. [Google Scholar] [CrossRef] [PubMed]
- Read, T.E.; Harris, H.W.; Grunfeld, C.; Feingold, K.R.; Calhoun, M.C.; Kane, J.P.; Rapp, J.H. Chylomicrons enhance endotoxin excretion in bile. Infect. Immun. 1993, 61, 3496–3502. [Google Scholar] [PubMed]
- Munford, R.S.; Andersen, J.M.; Dietschy, J.M. Sites of tissue binding and uptake in vivo of bacterial lipopolysaccharide-high density lipoprotein complexes: Studies in the rat and squirrel monkey. J. Clin. Investig. 1981, 68, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults. Executive Summary of the Third Report of the National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III). J. Am. Med. Assoc. 2001, 285, 2486–2497. [Google Scholar] [CrossRef]
- Andersen, C.J.; Blesso, C.N.; Lee, J.; Barona, J.; Shah, D.; Thomas, M.J.; Fernandez, M.L. Egg consumption modulates HDL lipid composition and increases the cholesterol-accepting capacity of serum in metabolic syndrome. Lipids 2013, 48, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Blesso, C.N.; Andersen, C.J.; Bolling, B.W.; Fernandez, M.L. Egg intake improves carotenoid status by increasing plasma hdl cholesterol in adults with metabolic syndrome. Food Funct. 2013, 4, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Kovacs-Nolan, J.; Phillips, M.; Mine, Y. Advances in the value of eggs and egg components for human health. J. Agric. Food Chem. 2005, 53, 8421–8431. [Google Scholar] [CrossRef] [PubMed]
- Blesso, C.N.; Andersen, C.J.; Barona, J.; Volek, J.S.; Fernandez, M.L. Whole egg consumption improves lipoprotein profiles and insulin sensitivity to a greater extent than yolk-free egg substitute in individuals with metabolic syndrome. Metabolism 2013, 62, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.K.; Rasmussen, H.E.; Ehlers, S.J.; Blobaum, K.R.; Lu, F.; Schlegal, V.L.; Carr, T.P.; Lee, J.Y. Repression of proinflammatory gene expression by lipid extract of nostoc commune var sphaeroides kutzing, a blue-green alga, via inhibition of nuclear factor-kappab in raw 264.7 macrophages. Nutr. Res. 2008, 28, 83–91. [Google Scholar] [CrossRef]
- Rasmussen, H.E.; Blobaum, K.R.; Park, Y.K.; Ehlers, S.J.; Lu, F.; Lee, J.Y. Lipid extract of nostoc commune var. Sphaeroides kutzing, a blue-green alga, inhibits the activation of sterol regulatory element binding proteins in hepg2 cells. J. Nutr. 2008, 138, 476–481. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Clark, R.M.; Park, Y.; Lee, J.; Fernandez, M.L. Lutein decreases oxidative stress and inflammation in liver and eyes of guinea pigs fed a hypercholesterolemic diet. Nutr. Res. Pract. 2012, 6, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Allsup, D.J.; Kamiguti, A.S.; Lin, K.; Sherrington, P.D.; Matrai, Z.; Slupsky, J.R.; Cawley, J.C.; Zuzel, M. B-cell receptor translocation to lipid rafts and associated signaling differ between prognostically important subgroups of chronic lymphocytic leukemia. Cancer Res. 2005, 65, 7328–7337. [Google Scholar] [CrossRef] [PubMed]
- Rentoukas, E.; Tsarouhas, K.; Kaplanis, I.; Korou, E.; Nikolaou, M.; Marathonitis, G.; Kokkinou, S.; Haliassos, A.; Mamalaki, A.; Kouretas, D.; et al. Connection between telomerase activity in pbmc and markers of inflammation and endothelial dysfunction in patients with metabolic syndrome. PLoS One 2012, 7, e35739. [Google Scholar] [CrossRef] [PubMed]
- Stoll, L.L.; Denning, G.M.; Weintraub, N.L. Endotoxin, TLR4 signaling and vascular inflammation: Potential therapeutic targets in cardiovascular disease. Curr. Pharm. Des. 2006, 12, 4229–4245. [Google Scholar] [CrossRef] [PubMed]
- Esser, D.; Oosterink, E.; op’t Roodt, J.; Henry, R.M.; Stehouwer, C.D.; Muller, M.; Afman, L.A. Vascular and inflammatory high fat meal responses in young healthy men; a discriminative role of IL-8 observed in a randomized trial. PLoS One 2013, 8, e53474. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wang, H.; Yao, H.; Wei, Q.; Mao, X.M.; Jiang, T.; Xiang, J.; Dila, N. Expression and activity of the TLR4/NF-kappaB signaling pathway in mouse intestine following administration of a short-term high-fat diet. Exp. Ther. Med. 2013, 6, 635–640. [Google Scholar] [PubMed]
- Shi, H.; Kokoeva, M.V.; Inouye, K.; Tzameli, I.; Yin, H.; Flier, J.S. TLR4 links innate immunity and fatty acid-induced insulin resistance. J. Clin. Investig. 2006, 116, 3015–3025. [Google Scholar] [CrossRef] [PubMed]
- Rossmeisl, M.; Jilkova, Z.M.; Kuda, O.; Jelenik, T.; Medrikova, D.; Stankova, B.; Kristinsson, B.; Haraldsson, G.G.; Svensen, H.; Stoknes, I.; et al. Metabolic effects of n-3 pufa as phospholipids are superior to triglycerides in mice fed a high-fat diet: Possible role of endocannabinoids. PLoS One 2012, 7, e38834. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, J.P.; Schneider, I.; Meyer, H.; Neubronner, J.; von Schacky, C.; Hahn, A. Incorporation of EPA and DHA into plasma phospholipids in response to different omega-3 fatty acid formulations—A comparative bioavailability study of fish oil vs. Krill oil. Lipids Health Dis. 2011, 10, 145. [Google Scholar] [CrossRef]
- Kim, J.J.; Sears, D.D. Tlr4 and insulin resistance. Gastroenterol. Res. Pract. 2010, 2010, 212563. [Google Scholar]
- Wong, S.W.; Kwon, M.J.; Choi, A.M.; Kim, H.P.; Nakahira, K.; Hwang, D.H. Fatty acids modulate toll-like receptor 4 activation through regulation of receptor dimerization and recruitment into lipid rafts in a reactive oxygen species-dependent manner. J. Biol. Chem. 2009, 284, 27384–27392. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Molina, A.; Castro, G.; Martin-Escalante, D.; Bravo, D.; Lopez-Miranda, J.; Castro, P.; Lopez-Segura, F.; Fruchart, J.C.; Ordovas, J.M.; Perez-Jimenez, F. Effects of different dietary cholesterol concentrations on lipoprotein plasma concentrations and on cholesterol efflux from fu5ah cells. Am. J. Clin. Nutr. 1998, 68, 1028–1033. [Google Scholar] [PubMed]
- Walton, K.A.; Cole, A.L.; Yeh, M.; Subbanagounder, G.; Krutzik, S.R.; Modlin, R.L.; Lucas, R.M.; Nakai, J.; Smart, E.J.; Vora, D.K.; et al. Specific phospholipid oxidation products inhibit ligand activation of toll-like receptors 4 and 2. Arterioscler. Thromb. Vasc.Biol. 2003, 23, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Ikonen, E. Cellular cholesterol trafficking and compartmentalization. Nat. Rev. Mol. Cell Biol. 2008, 9, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zhou, H.; Gu, Q.; Li, C. The expression of atp-binding cassette transporters in hypertensive patients. Hypertens. Res. 2009, 32, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.C.; Albrecht, C.; Pavitt, D.; Paul, V.; Pourreyron, C.; Newman, S.P.; Godsland, I.F.; Valabhji, J.; Johnston, D.G. Type 2 diabetes is associated with reduced atp-binding cassette transporter a1 gene expression, protein and function. PLoS One 2011, 6, e22142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, C.; Kanter, J.E.; Bornfeldt, K.E.; Leboeuf, R.C.; Oram, J.F. Diabetes reduces the cholesterol exporter abca1 in mouse macrophages and kidneys. J. Lipid Res. 2010, 51, 1719–1728. [Google Scholar] [CrossRef] [PubMed]
- Mei, C.L.; Chen, Z.J.; Liao, Y.H.; Wang, Y.F.; Peng, H.Y.; Chen, Y. Interleukin-10 inhibits the down-regulation of ATP binding cassette transporter A1 by tumour necrosis factor-alpha in THP-1 macrophage-derived foam cells. Cell Biol. Int. 2007, 31, 1456–1461. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liao, D.; Bharadwaj, U.; Li, M.; Yao, Q.; Chen, C. C-reactive protein inhibits cholesterol efflux from human macrophage-derived foam cells. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Landry, Y.D.; Denis, M.; Nandi, S.; Bell, S.; Vaughan, A.M.; Zha, X. ATP-binding cassette transporter A1 expression disrupts raft membrane microdomains through its ATPase-related functions. J. Biol. Chem. 2006, 281, 36091–36101. [Google Scholar] [CrossRef] [PubMed]
- Tellier, E.; Canault, M.; Poggi, M.; Bonardo, B.; Nicolay, A.; Alessi, M.C.; Nalbone, G.; Peiretti, F. HDLs activate ADAM17-dependent shedding. J. Cell. Physiol. 2008, 214, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Liao, D.F.; Tang, C.K. ATP-binding membrane cassette transporter A1 (ABCA1): A possible link between inflammation and reverse cholesterol transport. Mol. Med. 2010, 16, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Liu, Y.; Kessler, P.S.; Vaughan, A.M.; Oram, J.F. The macrophage cholesterol exporter abca1 functions as an anti-inflammatory receptor. J. Biol. Chem. 2009, 284, 32336–32343. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Andersen, C.J.; Lee, J.-Y.; Blesso, C.N.; Carr, T.P.; Fernandez, M.L. Egg Intake during Carbohydrate Restriction Alters Peripheral Blood Mononuclear Cell Inflammation and Cholesterol Homeostasis in Metabolic Syndrome. Nutrients 2014, 6, 2650-2667. https://doi.org/10.3390/nu6072650
Andersen CJ, Lee J-Y, Blesso CN, Carr TP, Fernandez ML. Egg Intake during Carbohydrate Restriction Alters Peripheral Blood Mononuclear Cell Inflammation and Cholesterol Homeostasis in Metabolic Syndrome. Nutrients. 2014; 6(7):2650-2667. https://doi.org/10.3390/nu6072650
Chicago/Turabian StyleAndersen, Catherine J., Ji-Young Lee, Christopher N. Blesso, Timothy P. Carr, and Maria Luz Fernandez. 2014. "Egg Intake during Carbohydrate Restriction Alters Peripheral Blood Mononuclear Cell Inflammation and Cholesterol Homeostasis in Metabolic Syndrome" Nutrients 6, no. 7: 2650-2667. https://doi.org/10.3390/nu6072650