The Role of Omega-3 Fatty Acids in Reverse Cholesterol Transport: A Review
by
Alex Pizzini
1,†, 
Lukas Lunger
2,†,
Egon Demetz
1,
Richard Hilbe
1,
Guenter Weiss
1,
Christoph Ebenbichler
2 and
Ivan Tancevski
1,* 1
Department of Internal Medicine II, Infectious Diseases, Pneumology, Rheumatology, Medical University of Innsbruck, 6020 Innsbruck, Austria
2
Department of Internal Medicine I, Gastroenterology, Hepatology, Endocrinology and Metabolism, Medical University of Innsbruck, 6020 Innsbruck, Austria
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Nutrients 2017, 9(10), 1099; https://doi.org/10.3390/nu9101099
Submission received: 2 August 2017
/
Revised: 22 September 2017
/
Accepted: 28 September 2017
/
Published: 6 October 2017
(This article belongs to the Special Issue Omega-3 Polyunsaturated Fatty Acids and Cardiovascular Health)
{kind=link}
{kind=link}
{kind=link}
Abstract
:The beneficial effects of omega-3 polyunsaturated fatty acids (n-3 PUFAs) on cardiovascular disease have been studied extensively. However, it remains unclear to what extent n-3 PUFAs may impact Reverse Cholesterol Transport (RCT). RCT describes a mechanism by which excess cholesterol from peripheral tissues is transported to the liver for hepatobiliary excretion, thereby inhibiting foam cell formation and the development of atherosclerosis. The aim of this review is to summarize the literature and to provide an updated overview of the effects of n-3 PUFAs on key players in RCT, including apoliprotein AI (apoA-I), ATP-binding cassette transporter A1 (ABCA1), ABCG1, apoE, scavenger receptor class B type I (SR-BI), cholesteryl ester transfer protein (CETP), low-density lipoprotein receptor (LDLr), cholesterol 7 alpha-hydroxylase (CYP7A1) and ABCG5/G8. Based on current knowledge, we conclude that n-3 PUFAs may beneficially affect RCT, mainly by influencing high-density lipoprotein (HDL) remodeling and by promoting hepatobiliary sterol excretion.
1. Introduction
Ischemic heart disease is still the leading cause of death in Western societies [1]. A strong inverse correlation between plasma concentrations of high-density lipoprotein cholesterol (HDL-C) and the incidence of atherosclerotic-driven cardiovascular disease (CVD) has been shown previously [2,3,4]. This has led to the hypothesis that interventions aimed at increasing HDL-C levels might positively influence the risk of CVD [5]. However, data on the efficacy of isolated HDL-C enhancing therapies are scarce and their effects remain controversial [6,7,8,9]. A main atheroprotective function of HDL particles is believed to be translated by the promotion of so-called “Reverse Cholesterol Transport” (RCT). This process describes the clearing pathways of peripheral, subendothelial macrophage- and fibroblast-derived cholesterol, either directly via HDL (hepatic uptake via scavenger receptor B-I, SR-BI), or indirectly by shifting cholesterol from HDL particles to apoB-containing lipoproteins for subsequent uptake into hepatocytes, via low-density lipoprotein receptors (LDLr) (Figure 1) [5,9].
Omega-3 polyunsaturated fatty acids (n-3 PUFAs) may beneficially influence the risk of cardiovascular disease (CVD), especially in the secondary prevention of CVD. Recent findings from randomized controlled trials and meta-analyses led to a class IIa recommendation by the American Heart Association for dietary supplementation with n-3 PUFAs in populations at high risk of cardiovascular disease [11]. Several molecular pathways and mechanisms of action affecting CVD risk have been found to be influenced by n-3 PUFAs, including the alteration of physical and chemical properties of cellular membranes, direct interaction with, and modulation of, membrane channels and proteins, regulation of gene expression via nuclear receptors and transcription factors, changes in eicosanoid profiles, and the conversion of n-3 PUFAs to bioactive metabolites [12]. So far, effects on HDL-C plasma levels have been shown to be only moderate [13,14,15].
Cholesterol metabolism is divided into an exogenous and an endogenous pathway; in the endogenous pathway, cholesterol is synthesized by the liver and extrahepatic tissues, and enters the circulation as a component of lipoproteins, or is secreted into bile. In the exogenous pathway, cholesterol from dietary and biliary sources is absorbed in the intestine and ultimately enters the circulation [16]. Due to the absence of peripheral cholesterol catabolic mechanisms, efficient efflux of intracellular cholesterol and subsequent transport of peripheral cholesterol to the liver is crucial, to avoid the accumulation of cytotoxic cholesterol crystals and the formation of atherosclerotic plaques in arteries [17]. Four cholesterol efflux routes (two active and two passive processes) have been described: ATP-binding cassette transporters A1 (ABCA1) and G1 (ABCG1) (active), aqueous diffusion and SR-BI facilitated cholesterol desorption (passive) (Figure 2) [10,17]. Cholesterol efflux, the first step of RCT, prevents intracellular cholesterol accumulation and has been shown to correlate with protection from atherosclerotic disease [17,18,19]. HDL particles are crucial mediators of cholesterol efflux from lipid-laden macrophages [17,20]. HDL particles comprise a heterogeneous lipoprotein population that differs in size, charge, density and composition [21].
The beneficial effects of omega-3 polyunsaturated fatty acids (n-3 PUFAs) on dyslipidemia are well established in literature. It has been found previously that eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), the two most important n-3 PUFAs in human physiology, may exert their atheroprotective functions by promoting intracellular catabolism of apolipoprotein B-100 containing lipoproteins, suppressing hepatic apoB production, stimulating plasma triglyceride clearance via lipoprotein lipase (LPL), increasing the very low density lipoprotein (VLDL) to LDL conversion rate, reducing LDL synthesis and attenuating postprandial lipemia [22,23,24,25]. It appears likely that n-3 PUFAs may mediate these effects, predominately by inhibiting sterol regulatory element-binding protein-1 (SREBP-1) mediated pathways, including the activation of the nuclear transcription factors, Hepatocyte Nuclear Factor-4 Alpha (HNF4A), Farnesoid X Receptor, Liver X Receptor (LXR), and Peroxisome Proliferator-activated Receptors (PPARs) [26].
Even though previous literature investigating the effect of n-3 PUFAs and their effects on cholesterol efflux routes and RCT is still ambiguous, it has been clearly shown that n-3 PUFAs promote the RCT mechanism. Using an established RCT model, in which (3H)-labeled macrophages are injected into the peritoneal cavity of animals—where the appearance of a tracer in plasma and feces is monitored—different laboratories have demonstrated that n-3 PUFAs promote macrophage-to-feces RCT in rodents [27,28,29]. In human macrophages, in turn, n-3 PUFAs have been shown to significantly increase cholesterol efflux from macrophages [30].
This review aims to summarize current knowledge on the effects of n-3 PUFAs on RCT and related pathways.
1.1. Apolipoprotein AI (apoA-I), Paraoxonase-1 (PON1), and n-3 PUFAs
ApoA-I is the principal protein component of HDL particles. It mediates HDL maturation by promoting ABCA1-mediated cholesterol efflux from cells, and by stimulating enzymatic cholesterol esterification via lecithin cholesterol acyl transferase (LCAT) [10,17,31]. Plasma LCAT activity was increased in hamsters fed a high-fat diet enriched with n-3 PUFAs (n-3 PUFA oil mixture), which was associated with an increased macrophage-to-feces RCT [29]. Moreover, n-3 PUFAs (fish oil, perilla oil, soybean oil) were found to increase hepatic protein expression of apoA-I in obese insulin resistant rats, resulting in significantly increased apoA-I plasma levels [32]. However, clear data demonstrating increased cholesterol efflux from macrophages through induction of apoA-I are lacking.
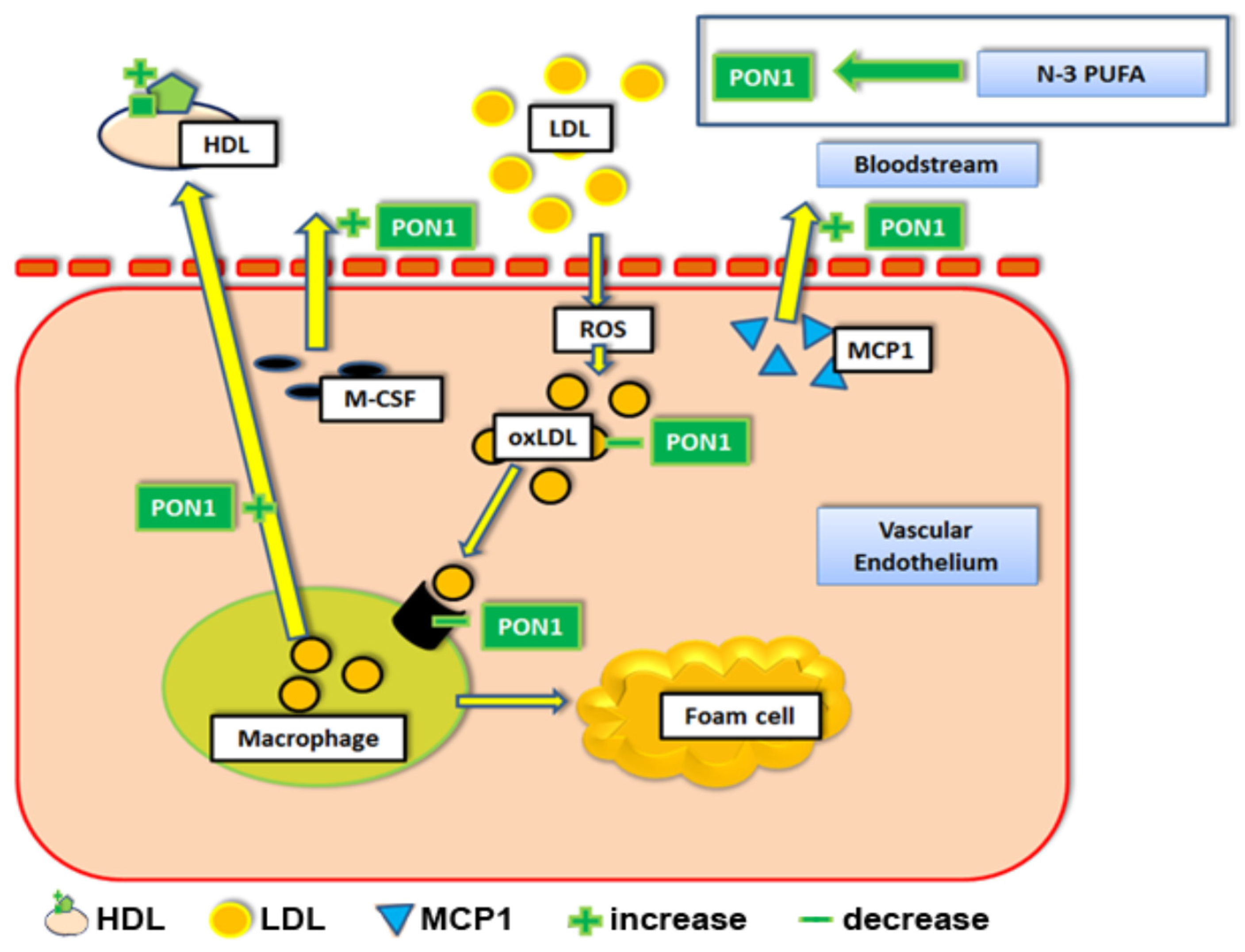
Interestingly, induction of apoA-I expression by n-3 PUFAs (EPA) may also critically influence the anti-oxidative effects of HDLs [33,34]. ApoA-I is known to stabilize the enzymatic activity of paraoxonase-1 (PON1) [35,36] which associates with HDL particles. PON1, in turn, prevents oxidative modification of LDLs, detoxifies oxidized LDLs (oxLDL), inhibits uptake of oxLDLs by macrophages, and reduces macrophage oxidative stress. Overall, PON1 transgenic apoE−/− mice have been found to have dramatically reduced levels of pro-inflammatory cytokines within aortic tissues compared to solely apoE−/− mice (e.g. macrophage colony stimulation factor, M-CSF and in monocyte chemotactic protein, MCP1, Figure 3) [36,37,38], which may result from their antioxidative effects in locally accumulating cells of myeloid origin. Notably, it was also suggested that EPA may enhance the antioxidant qualities of HDLs by activating HDL-associated PON1, independently of apoA-I [34,39]. Finally, PON1 has also been shown to increase the cholesterol efflux capacity of macrophages; both human Tamm-Horsfall protein 1 (THP-1) macrophages as well as murine J774 macrophages displayed a dose-dependent increase in cholesterol efflux when treated with increasing concentrations of PON1 [40].
Taken together, n-3 PUFAs seem not to exert direct effects on the lipoprotein cholesterol content, but rather modify HDL composition and function, resulting in increased HDL maturation and PON1-dependent antioxidative effects.
1.2. ATP-Binding Cassette Transporter A1 (ABCA1) and n-3 PUFAs
ABCA1 plays a key role in the efflux of phospholipids and free cholesterol from cells to lipid-free apoA-I [31,41], leading to the generation of discoidal nascent HDL particles [17,42]. In a subsequent step, LCAT facilitates the condensation of cholesterol into the lipoprotein core via esterification, yielding spherical mature HDL particles [10,43,44]. Mature HDL particles require supplementary, non-ABCA1 mediated lipidation, either via ABCG1, aqueous diffusion or SR-BI mediated pathways [10,17,45].
n-3 PUFAs appear to have diverging effects on macrophage-specific cholesterol efflux in mice and in humans. In vitro studies showed that EPA impairs ABCA1 mediated cholesterol efflux in both human and murine macrophages, whereas DHA stimulates ABCA1 dependent efflux in murine macrophages [46]. Chadli et al. as well as Nishimoto et al. reported n-3 PUFAs (n-3 PUFA oil mixture; EPA) to stimulate RCT, which was reflected by increased fecal sterol excretion [27,29]. However, the authors failed to prove a stimulation of HDL-mediated cholesterol efflux from macrophages. In summary, n-3 PUFAs appear not to promote ABCA1/apoA-I mediated cholesterol efflux from macrophages [29,47].
1.3. ATP-Binding Cassette Transporter G1 (ABCG1) and n-3 PUFAs
Pertaining to the family of ATP-binding cassette transporters, ABCG1 is an integral membrane protein which actively pumps intracellular cholesterol to the outer plasma membrane, leaving it to freely desorb into the extracellular milieu [17]. In contrast to ABCA1, ABCG1 does not enhance the transport of cholesterol and phospholipids to lipid-free apoA-I, rather it mediates the efflux of cellular cholesterol to mature HDL [48,49].
It appears that major emphasis has been put on investigating ABCG1 in in vitro gene expression following a n-3 PUFA challenge. Available studies suggest that EPA treatment may reduce ABCG1 expression in human THP-1 monocytic cells; however, this does not occur in fully differentiated macrophages [50]. Further in vitro studies by Uehara et al. corroborated these findings and suggested EPA as a suppressor of the transcription and protein expression of ABCG1 and ABCA1 in murine RAW 264.7 macrophages [47].
ABCA1 and ABCG1 are not only expressed in macrophages, but also in liver cells, where they critically influence HDL synthesis and maturation. As mentioned above, n-3 PUFAs have been found to increase macrophage-to-feces RCT in hamsters, which on one hand could be traced back to increased hepatobiliary flux of bile acids and cholesterol, on the other hand appears to be due to an enhanced cholesterol acceptor capacity of HDL [29].
1.4. Apolipoprotein E (apoE) and n-3 PUFAs
ApoE is a further important apolipoprotein found in HDL particles, and its role in RCT is well established—apoE enhances cholesterol efflux via ABCA1 in macrophages and delivers HDL cholesterol to the liver through interactions with SR-BI and LDLr. It also facilitates the maturation of large HDL particles through LCAT [10]. Most available studies on the interplay between n-3 PUFAs and apoE relate to Alzheimer’s disease; previous findings suggest apoE4 polymorphisms to be a genetic risk factor for late onset Alzheimer’s disease [51], and epidemiological studies have postulated a clinical benefit of dietary supplementation with n-3 PUFAs in patients with dementia [51,52,53]. Studies investigating the direct effects of n-3 PUFAs on apoE expression and function are scarce since the most commonly used atherosclerosis model is the apoE knockout mouse [54].
Yet, in obese rats fed a mixture of n-3 and n-6 PUFAs (blend of safflower and soybean oil), an increase in apoE within HDL particles has been observed [55]. However, the study did not investigate the effects of isolated n-3 PUFA supplementation on apoE plasma levels. A recent study in healthy volunteers with an n-3 PUFA (EPA:DHA ratio = 0.54) enriched diet showed a significant increase in apoE within the LDL fraction and in a subgroup with reduced triglyceride levels, an increase in total plasma apoE concentrations [56]. Interestingly, genetic polymorphisms of apoE seem to impact the metabolism and bioavailability of supplemented EPA and DHA [57,58,59].
1.5. Scavenger Receptor Class B Type I (SR-BI) and n-3 PUFAs
The scavenger receptor class B type I (SR-BI) is a transporter which, in contrast to ABCA1 and ABCG1, enables passive bidirectional flux of cholesterol between cells and HDL particles [10].
To understand the impact of n-3 PUFAs on SR-BI, it is crucial to mention that this pathway is involved not only in the desorption of peripheral cholesterol to HDL particles, but also in the hepatic clearance of HDL derived cholesterol [10]. Therefore, in peripheral macrophages, the relative contribution of SR-BI mediated cellular cholesterol efflux has been controversially discussed. Some animal studies have suggested a significant efflux function of peripheral SR-BI [60,61], whereas other results from cell cultures and animal models are ambiguous [62,63,64,65]. It therefore appears that peripheral SR-BI mediated cholesterol efflux may play a minor role, as compared to clearance via ABCG1 and ABCA1 [64].
On the other hand, the major importance of SR-BI lies in its ability to clear HDL cholesterol via hepatic cells, an important step in RCT [66]. Little is known about PUFAs and their effects on SR-BI; a study in genetically obese mutant rats fed a combination of n-3 and n-6 PUFAs (blend of safflower and soybean oil) showed an increase in hepatic SR-BI expression [55]. ApoE knock out mice, fed different ratios of n-3/n-6 PUFAs (n3-PUFA derived from perilla seed oil containing mostly alpha-linolenic acid (ALA), and minimal amounts of EPA and DHA; safflower oil containing 83% Linoleic acid (LA) was used as the primary source of n-6 fatty acids) displayed reduced hepatic SR-BI mRNA levels when fed a relatively low n-3/n-6 PUFA ratio [67]. In high n-3/n-6 PUFA ratio fed mice, hepatic SR-BI mRNA as well as HDL particle plasma levels increased, while no effect on atherogenesis was observed. Chadli et al. analysed the impact of a high-fat diet, supplemented with n-3 PUFAs in hamsters, an animal species expressing Cholesteryl Ester Transfer Protein (CETP) [29]. This study revealed, as mentioned above, an increase in RCT following n-3 PUFA treatment, probably due to increased expression of the main hepatic genes involved in RCT, including SR-BI [29].
A recent in vitro analysis by Mashurabad et al. showed that EPA leads to a downregulation in SR-BI via PPARα-dependent mechanisms in enterocytes (Caco-2/TC7 cell line), suggesting that the implications of n-3 PUFAs on the translation of RCT modifying proteins do not only occur in hepatocytes [68].
In summary, the available literature shows an effect of n-3 PUFAs on SR-BI expression in hepatocytes, which may translate into elevated plasma HDL cholesterol clearance as part of the RCT. No data is available about n-3 PUFA and cholesterol efflux, which is driven through SR-BI from peripheral macrophages.
1.6. Cholesterol Ester Transfer Protein (CETP) and n-3 PUFAs
CETP is a plasma glycoprotein secreted by the liver which enables the exchange of cholesteryl esters between HDL and other lipoproteins, such as LDL, thereby promoting hepatic clearance of macrophage-derived cholesterol, via an indirect pathway by hepatic LDLr.
CETP transgenic mice treated with fish oil containing n-3 PUFAs showed significant increases in CETP plasma levels and activity, as well as elevated CETP mRNA expression in the liver [69]. The authors hypothesized a PPARα-related induction of CETP transcription through binding of n-3 PUFAs to PPARα response elements in the CETP gene [70], since PUFAs have been described as natural ligands for PPARα [71]. In a recent study in hamsters fed a high-fat diet and administered EPA or DHA, distinct results for the CETP-LDL cholesterol pathways were observed [72]. It was found that DHA, in contrast to EPA, led to a significant increase in LDL cholesterol concentrations, an increase in plasma CETP activity as well as elevated CETP mRNA expression in adipose tissue. The effect of EPA on the same parameters, however, was less prominent and suggests that the impact of n-3 PUFAs on CETP and RCT may depend on the type of n-3 PUFA administered [72]. These findings underline the importance of distinct analyses of different PUFA subclasses, as they may exert differential effects on cholesterol metabolism [73].
1.7. LDL-Receptor (LDLr) and n-3 PUFAs
Hepatic LDLr mediated uptake of circulating LDL cholesterol constitutes a final part of RCT. However, the effects of n-3 PUFAs on this last step of RCT are not entirely clear. Animal studies in LDLr deficient mice suggest that the beneficial effect of n-3 PUFAs (EPA, DHA and docosapentaenoic acid n-3) on serum lipoprotein profiles and aortic plaque formation may be independent of LDLr [74]. Also, another study by Vasandani underlined the hypothesis that n-3 PUFAs (EPA:DHA ratio = 1.48) may decrease the plasma concentration of apoB-containing lipoproteins, independent of LDLr or lipoprotein receptor-related protein mediated pathways [75].
It was previously shown that n-3 PUFAs promote post-ER presecretory proteolysis (PERPP)/autophagy, which degrades apoB100, thereby critically reducing lipoprotein secretion from the liver. n-3 PUFA induced autophagy was shown to be dependent on lipid peroxidation, which could be blocked by antioxidants, including vitamin E [76,77]. The manipulation of oxidation in humans for the purpose of preventing CVD has received substantial attention and effort. Administration of antioxidants, such as vitamin E, has generally lessened arterial lesions in animal models of atherosclerosis but has not resulted in consistent benefits [78] and has even caused occasional harm in humans [79,80]. Results by Fisher and co-workers have implied that the potential benefits of antioxidants on human arterial lesion development might be counterbalanced by potentially harmful alterations of apoB100 metabolism [76], which raises the question of whether similar effects may also hamper the hepatic secretion of proteins involved in RCT.
1.8. Cholesterol 7alpha-Hydroxylase (CYP7A1), ABCG5/G8, NPC1L1 and n-3 PUFAs
The last steps of RCT comprise hepatobiliary excretion of neutral sterols via ABCG5/G8, or excretion of acidic sterols after conversion of cholesterol into bile acids via cholesterol 7alpha-hydroxylase 1 (CYP7A1). Bérard et al. showed that dietary supplementation with n-3 PUFAs (EPA:DHA ratio = 1.46) induces CYP7A1 transcription through the D-site binding protein and LXR in murine hepatocytes [81]. n-3 PUFAs appear to increase bile acid synthesis by inducing hepatic CYP7A1 expression and to enhance hepatobiliary excretion of neutral sterols via ABCG5 in hamsters, which together should promote macrophage-to-feces RCT [29]. Increased hepatobiliary excretion of neutral sterols has also been observed in experiments in C57BL/6 mice fed a diet supplemented with fish oil, most likely through hepatic upregulation of ABCG5/G8 and intestinal downregulation of Niemann-Pick C1-like protein 1 (NPC1L1), which drives intestinal sterol reabsorption under physiological conditions [27].
Together with the liver, the intestine serves as a homeostatic organ in cholesterol metabolism. Recent evidence has substantiated the pivotal role of intestinal RCT. This mechanism has been described as trans-intestinal excretion of plasma-derived cholesterol (TICE) [82]. Located at the brush border membrane of the small intestine, ABCG5/G8 is the main contributor to TICE; ABCB1a/b might play a role as an additional route. Whereas no influence of n-3-PUFAs on intestinal ABCB1a/b has been investigated so far, there is evidence that DHA and EPA may affect intestinal cholesterol absorption by downregulating NPC1L1 [83], which, in turn, may critically affect ABCG5/G8 mediated trans-intestinal cholesterol excretion and thereby RCT.
2. Conclusions
n-3 PUFAs have a positive impact on atherosclerosis and CVD, a major concern of today’s health care systems. It is important to emphasize that the positive effects of n-3 PUFAs on CVD are thought to be mediated by diverse mechanisms, including the alteration of physical and chemical properties of cellular membranes, direct interaction with, and modulation of, membrane channels and proteins, regulation of gene expression via nuclear receptors and transcription factors, changes in eicosanoid profiles, and conversion of n-3 PUFAs to bioactive metabolites, which may promote ischemia-induced myocyte healing.
The aim of this article was to review possible beneficial effects of n-3 PUFAs on RCT. After evaluating available literature, it appears likely that this effect is, to a certain degree, mediated by positively influencing RCT on different levels, as illustrated by the stimulation of macrophage-to-feces RCT. However, these studies may not adequately illustrate the complexity of involved mechanisms and steps. Taking a closer look at the pivotal steps of RCT, the available literature suggests that n-3 PUFAs might affect several, but not all, involved processes. Interpretation of available studies is difficult and hampered by heterogenous study designs, sample sizes, different PUFA formulations used (n-3/n-6 PUFAs, phytosterols and/or single formulations of EPA or DHA etc.), and varying experimental approaches (in vivo versus in vitro). The application of results derived from mouse studies is limited by striking differences between murine and human lipoprotein metabolism, the mouse being an ‘HDL’ animal without CETP in plasma. Available studies in hamsters, which naturally express CETP, may help to better extrapolate observed physiological effects of n-3 PUFAs on RCT to the human setting. However, the number of studies in this animal model is limited, warranting further experiments in CETP-expressing, potentially dyslipidemic animal models to verify existing hypotheses on n-3 PUFAs and RCT. In summary, n-3 PUFAs may not significantly affect the first step of RCT—i.e., ATP-dependent cholesterol efflux mechanisms within atherosclerotic plaque macrophages—rather, they appear to beneficially affect HDL remodeling through LCAT and CETP, facilitating SR-BI and LDLr mediated hepatic uptake of plaque-derived excess cholesterol. In addition, n-3 PUFAs were clearly shown to promote hepatobiliary excretion of neutral and of acidic sterols, which is a pivotal final step in RCT.
Gaining further insight into the functionality of RCT may help to determine key populations who may substantially benefit from n-3 PUFA supplementation, an easy-to-obtain and interesting dietary supplement, with potentially promising effects in CVD.
Acknowledgments
Funding by the Austrian K Project Vascage (to G.W. and I.T.) is gratefully acknowledged.
Author Contributions
The authors Alex Pizzini and Lukas Lunger did equally contribute to conceive this manuscript and are therefore listed as shared first authors. Manuscript compilation and graphical design were authored to equal shares by Alex Pizzini and Lukas Lunger. Egon Demetz, Guenter Weiss, Christoph Ebenbichler and Ivan Tancevski supervised and corrected the resulting manuscript. Richard Hilbe significantly contributed to the revision of the final manuscript.
Conflicts of interest
The authors declare no conflict of interest.
References
- World Health Organization (WHO). The Top 10 Causes of Death; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Rader, D.J. Regulation of reverse cholesterol transport and clinical implications. Am. J. Cardiol. 2003, 92, 42–49. [Google Scholar] [CrossRef]
- Kannel, W.B.; Dawber, T.R.; Friedman, G.D.; Glennon, W.E.; Mcnamara, P.M. Risk Factors in Coronary Heart Disease: An Evaluation of Several Serum Lipids as Predictors of Coronary Heart Disease: The Framingham Study. Ann. Intern. Med. 1964, 61 (5 Pt 1), 888–899. [Google Scholar] [CrossRef]
- Assmann, G.; Cullen, P.; Schulte, H. Simple Scoring Scheme for Calculating the Risk of Acute Coronary Events Based on the 10-Year Follow-Up of the Prospective Cardiovascular Münster (PROCAM) Study. Circulation 2002, 105, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, H.K.; Kiss, D.; Rader, D. HDL-cholesterol and cardiovascular disease: rethinking our approach. Curr. Opin. Cardiol. 2015, 30, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Badimon, J.J.; Badimon, L.; Fuster, V. Regression of atherosclerotic lesions by high density lipoprotein plasma fraction in the cholesterol-fed rabbit. J. Clin. Investig. 1990, 85, 1234–1241. [Google Scholar] [CrossRef] [PubMed]
- Rubin, E.M.; Krauss, R.M.; Spangler, E.A.; Verstuyft, J.G.; Clift, S.M. Inhibition of early atherogenesis in transgenic mice by human apolipoprotein AI. Nature 1991, 353, 265–267. [Google Scholar] [CrossRef] [PubMed]
- Tangirala, R.K.; Tsukamoto, K.; Chun, S.H.; Usher, D.; Puré, E.; Rader, D.J. Regression of Atherosclerosis Induced by Liver-Directed Gene Transfer of Apolipoprotein A-I in Mice. Circulation 1999, 100, 1816–1822. [Google Scholar] [CrossRef] [PubMed]
- Mahdy Ali, K.; Wonnerth, A.; Huber, K.; Wojta, J. Cardiovascular disease risk reduction by raising HDL cholesterol—Current therapies and future opportunities. Br. J. Pharmacol. 2012, 167, 1177–1194. [Google Scholar] [CrossRef] [PubMed]
- Rosenson, R.S.; Brewer, H.B.; Davidson, W.S.; Fayad, Z.A.; Fuster, V.; Goldstein, J.; Hellerstein, M.; Jiang, X.C.; Phillips, M.C.; Rader, D.J.; et al. Cholesterol efflux and atheroprotection: Advancing the concept of reverse cholesterol transport. Circulation 2012, 125, 1905–1919. [Google Scholar] [CrossRef] [PubMed]
- Siscovick, D.S.; Barringer, T.A.; Fretts, A.M.; Wu, J.H.Y.; Lichtenstein, A.H.; Costello, R.B.; Kris-Etherton, P.M.; Jacobson, T.A.; Engler, M.B.; Alger, H.M.; et al. Omega-3 Polyunsaturated Fatty Acid (Fish Oil) Supplementation and the Prevention of Clinical Cardiovascular Disease. Circulation 2017, 135, e867–e884. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Wu, J.H.Y. Omega-3 Fatty Acids and Cardiovascular Disease. J. Am. Coll. Cardiol. 2011, 58, 2047–2067. [Google Scholar] [CrossRef] [PubMed]
- Ramprasath, V.R.; Jones, P.J.H.; Buckley, D.D.; Woollett, L.A.; Heubi, J.E. Decreased plasma cholesterol concentrations after PUFA-rich diets are not due to reduced cholesterol absorption/synthesis. Lipids 2012, 47, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Vecka, M.; Dušejovská, M.; Stankova, B.; Zeman, M.; Vavrova, L.; Kodydkova, J.; Slabý, A.; Žák, A. N-3 polyunsaturated fatty acids in the treatment of atherogenic dyslipidemia. Neuro Endocrinol. Lett. 2012, 33 (Suppl. S2), 87–92. [Google Scholar] [PubMed]
- Wang, F.; Wang, Y.; Zhu, Y.; Liu, X.; Xia, H.; Yang, X.; Sun, G. Treatment for 6 months with fish oil-derived n-3 polyunsaturated fatty acids has neutral effects on glycemic control but improves dyslipidemia in type 2 diabetic patients with abdominal obesity: A randomized, double-blind, placebo-controlled trial. Eur. J. Nutr. 2016. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, J. The role of the exogenous pathway in hypercholesterolaemia. Eur. Hear. J. Suppl. 2001, 3, 2–5. [Google Scholar] [CrossRef]
- Phillips, M.C. Molecular mechanisms of cellular cholesterol efflux. J. Biol. Chem. 2014, 289, 24020–24029. [Google Scholar] [CrossRef] [PubMed]
- Cuchel, M.; Rader, D.J. Macrophage reverse cholesterol transport: Key to the regression of atherosclerosis? Circulation 2006, 113, 2548–2555. [Google Scholar] [CrossRef] [PubMed]
- Rader, D.J.; Alexander, E.T.; Weibel, G.L.; Billheimer, J.; Rothblat, G.H. The role of reverse cholesterol transport in animals and humans and relationship to atherosclerosis. J. Lipid Res. 2009, 50 (Suppl. S1), 89–94. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Van Berkel, T.J.; Van Eck, M. Relative roles of various efflux pathways in net cholesterol efflux from macrophage foam cells in atherosclerotic lesions. Curr. Opin. Lipidol. 2010, 21, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Khera, A.V.; Cuchel, M.; de la Llera-Moya, M.; Rodrigues, A.; Burke, M.F.; Jafri, K.; French, B.C.; Phillips, J.A.; Mucksavage, M.L.; Wilensky, R.L.; et al. Cholesterol efflux capacity, high-density lipoprotein function, and Atherosclerosis. N. Engl. J. Med. 2011, 364, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.C.; Watts, G.F.; Mori, T.A.; Barrett, P.H.R.; Redgrave, T.G.; Beilin, L.J. Randomized controlled trial of the effect of n-3 fatty acid supplementation on the metabolism of apolipoprotein B-100 and chylomicron remnants in men with visceral obesity. Am. J. Clin. Nutr. 2003, 77, 300–307. [Google Scholar] [PubMed]
- Park, Y.; Harris, W.S. Omega-3 fatty acid supplementation accelerates chylomicron triglyceride clearance. J. Lipid Res. 2003, 44, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.C.; Watts, G.F.; Barrett, P.H.R.; Beilin, L.J.; Redgrave, T.G.; Mori, T.A. Regulatory Effects of HMG CoA Reductase Inhibitor and Fish Oils on Apolipoprotein B-100 Kinetics in Insulin-Resistant Obese Male Subjects With Dyslipidemia. Diabetes 2002, 51, 2377–2386. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.G.; Stone, N.J. Antiatherosclerotic and antithrombotic effects of omega-3 fatty acids. Am. J. Cardiol. 2006, 98, 39i–49i. [Google Scholar] [CrossRef] [PubMed]
- Zuliani, G.; Galvani, M.; Leitersdorf, E.; Volpato, S.; Cavalieri, M.; Fellin, R. The Role of Polyunsaturated Fatty Acids (PUFA) in the Treatment of Dyslipidemias. Curr. Pharm. Des. 2009, 15, 4087–4093. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, T.; Pellizzon, M.A.; Aihara, M.; Stylianou, I.M.; Billheimer, J.T.; Rothblat, G.; Rader, D.J. Fish Oil Promotes Macrophage Reverse Cholesterol Transport in Mice. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1502–1508. [Google Scholar] [CrossRef] [PubMed]
- Balogun, K.A.; Randunu, R.S.; Cheema, S.K. The effect of dietary omega-3 polyunsaturated fatty acids on plasma lipids and lipoproteins of C57BL/6 mice is age and sex specific. Prostaglandins Leukot. Essent. Fat. Acids 2014, 91, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Kasbi Chadli, F.; Nazih, H.; Krempf, M.; Nguyen, P.; Ouguerram, K. Omega 3 Fatty Acids Promote Macrophage Reverse Cholesterol Transport in Hamster Fed High Fat Diet. PLoS ONE 2013, 8, e61109. [Google Scholar] [CrossRef] [PubMed]
- Moss, J.W.E.; Davies, T.S.; Garaiova, I.; Plummer, S.F.; Michael, D.R.; Ramji, D.P. A Unique Combination of Nutritionally Active Ingredients Can Prevent Several Key Processes Associated with Atherosclerosis In Vitro. PLoS ONE 2016, 11, e0151057. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Park, Y.; Koo, S.I. ATP-binding cassette transporter A1 and HDL metabolism: Effects of fatty acids. J. Nutr. Biochem. 2012, 23, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Zhang, T.; Zhao, S.; Li, W.; Ma, L.; Ding, M.; Liu, Y. Effects of n-3 polyunsaturated fatty acids high fat diet intervention on the synthesis of hepatic high-density lipoprotein cholesterol in obesity-insulin resistance rats. Lipids Health Dis. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Irino, Y.; Shinohara, M.; Tsuda, S.; Mori, T.; Nagao, M.; Oshita, T.; Mori, K.; Hara, T.; Toh, R.; et al. Eicosapentaenoic Acid-Enriched High-Density Lipoproteins Exhibit Anti-Atherogenic Properties. Circ. J. 2017. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Ishida, T.; Nagao, M.; Mori, T.; Monguchi, T.; Sasaki, M.; Mori, K.; Kondo, K.; Nakajima, H.; Honjo, T.; et al. Administration of high dose eicosapentaenoic acid enhances anti-inflammatory properties of high-density lipoprotein in Japanese patients with dyslipidemia. Atherosclerosis 2014, 237, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Gaidukov, L.; Viji, R.I.; Yacobson, S.; Rosenblat, M.; Aviram, M.; Tawfik, D.S. ApoE induces serum paraoxonase PON1 activity and stability similar to ApoA-I. Biochemistry 2010, 49, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Litvinov, D.; Mahini, H.; Garelnabi, M. Antioxidant and anti-inflammatory role of paraoxonase 1: Implication in arteriosclerosis diseases. N. Am. J. Med. Sci. 2012, 4, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Fülöp, P.; Harangi, M.; Seres, I.; Paragh, G. Paraoxonase-1 and adipokines: Potential links between obesity and Atherosclerosis. Chem. Biol. Interact. 2016. [Google Scholar] [CrossRef] [PubMed]
- Mackness, M.; Mackness, B. Human paraoxonase-1 (PON1): Gene structure and expression, promiscuous activities and multiple physiological roles. Gene 2015, 567, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Burillo, E.; Mateo-Gallego, R.; Cenarro, A.; Fiddyment, S.; Bea, A.M.; Jorge, I.; Vázquez, J.; Civeira, F. Beneficial effects of omega-3 fatty acids in the proteome of high-density lipoprotein proteome. Lipids Health Dis. 2012, 11, 116. [Google Scholar] [CrossRef] [PubMed]
- Berrougui, H.; Loued, S.; Khalil, A. Purified human paraoxonase-1 interacts with plasma membrane lipid rafts and mediates cholesterol efflux from macrophages. Free Radic. Biol. Med. 2012, 52, 1372–1381. [Google Scholar] [CrossRef] [PubMed]
- Landry, Y.D.; Denis, M.; Nandi, S.; Bell, S.; Vaughan, A.M.; Zha, X. ATP-binding cassette transporter A1 expression disrupts raft membrane microdomains through its ATPase-related functions. J. Biol. Chem. 2006, 281, 36091–36101. [Google Scholar] [CrossRef] [PubMed]
- Vedhachalam, C.; Duong, P.T.; Nickel, M.; Nguyen, D.; Dhanasekaran, P.; Saito, H.; Rothblat, G.H.; Lund-Katz, S.; Phillips, M.C. Mechanism of ATP-binding cassette transporter A1-mediated cellular lipid efflux to apolipoprotein A-I and formation of high density lipoprotein particles. J. Biol. Chem. 2007, 282, 25123–25130. [Google Scholar] [CrossRef] [PubMed]
- Daniels, T.F.; Killinger, K.M.; Michal, J.J.; Wright, R.W.; Jiang, Z. Lipoproteins, cholesterol homeostasis and cardiac health. Int. J. Biol. Sci. 2009, 5, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Rye, K.A.; Barter, P.J. Cardioprotective functions of HDLs. J. Lipid Res. 2014, 55, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Mulya, A.; Lee, J.Y.; Gebre, A.K.; Thomas, M.J.; Colvin, P.L.; Parks, J.S. Minimal lipidation of pre-beta HDL by ABCA1 results in reduced ability to interact with ABCA1. Arter. Thromb. Vasc. Biol. 2007, 27, 1828–1836. [Google Scholar] [CrossRef] [PubMed]
- Fournier, N.; Tardivel, S.; Benoist, J.-F.; Vedie, B.; Rousseau-Ralliard, D.; Nowak, M.; Allaoui, F.; Paul, J.L. Eicosapentaenoic acid membrane incorporation impairs ABCA1-dependent cholesterol efflux via a protein kinase A signaling pathway in primary human macrophages. Biochim. Biophys. Acta 2016, 1861, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Uehara, Y.; Miura, S.; von Eckardstein, A.; Abe, S.; Fujii, A.; Matsuo, Y.; Rust, S.; Lorkowski, S.; Assmann, G.; Yamada, T.; et al. Unsaturated fatty acids suppress the expression of the ATP-binding cassette transporter G1 (ABCG1) and ABCA1 genes via an LXR/RXR responsive element. Atherosclerosis 2007, 191, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, A.M.; Oram, J.F. ABCG1 Redistributes Cell Cholesterol to Domains Removable by High Density Lipoprotein but Not by Lipid-depleted Apolipoproteins. J. Biol. Chem. 2005, 280, 30150–30157. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan, S.; Oram, J.F.; Asztalos, B.F.; Vaughan, A.M.; Lund-Katz, S.; Adorni, M.P.; Phillips, M.C.; Rothblat, G.H. Effects of acceptor composition and mechanism of ABCG1-mediated cellular free cholesterol efflux. J. Lipid Res. 2009, 50, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Moradi Sarabi, M.; Doosti, M.; Einollahi, N.; Hesami, S.S.; Dashti, N. Effect of eicosapentaenoic acid on the expression of ABCG1 gene in the human monocyte THP-1 cells. Acta Med. Iran. 2014, 52, 176–181. [Google Scholar] [PubMed]
- Barberger-Gateau, P.; Samieri, C.; Féart, C.; Plourde, M. Dietary Omega 3 polyunsaturated fatty acids and Alzheimer’s disease: Interaction with apolipoprotein E genotype. Curr. Alzheimer Res. 2011, 8, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Evans, D.A.; Bienias, J.L.; Tangney, C.C.; Bennett, D.A.; Wilson, R.S.; Aggarwal, N.; Schneider, J. Consumption of fish and n-3 fatty acids and risk of incident Alzheimer disease. Arch. Neurol. 2003, 60, 940–946. [Google Scholar] [CrossRef] [PubMed]
- Kalmijn, S.; Launer, L.J.; Ott, A.; Witteman, J.C.; Hofman, A.; Breteler, M.M. Dietary fat intake and the risk of incident dementia in the Rotterdam Study. Ann. Neurol. 1997, 42, 776–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolovou, G.; Anagnostopoulou, K.; Mikhailidis, D.P.; Cokkinos, D.V. Apolipoprotein E knockout models. Curr. Pharm. Des. 2008, 14, 338–351. [Google Scholar] [CrossRef] [PubMed]
- Sheril, A.; Jeyakumar, S.M.; Jayashree, T.; Giridharan, N.V.; Vajreswari, A. Impact of feeding polyunsaturated fatty acids on cholesterol metabolism of dyslipidemic obese rats of WNIN/GR-Ob strain. Atherosclerosis 2009, 204, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Lambert, C.; Cubedo, J.; Padró, T.; Sánchez-Hernández, J.; Antonijoan, R.M.; Perez, A.; Badimon, L. Phytosterols and Omega 3 Supplementation Exert Novel Regulatory Effects on Metabolic and Inflammatory Pathways: A Proteomic Study. Nutrients 2017, 9, 599. [Google Scholar] [CrossRef] [PubMed]
- Minihane, A.M. Impact of Genotype on EPA and DHA Status and Responsiveness to Increased Intakes. Nutrients 2016, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chouinard-Watkins, R.; Conway, V.; Minihane, A.M.; Jackson, K.G.; Lovegrove, J.A.; Plourde, M. Interaction between BMI and APOE genotype is associated with changes in the plasma long-chain-PUFA response to a fish-oil supplement in healthy participants. Am. J. Clin. Nutr. 2015, 102, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Thifault, E.; Cormier, H.; Bouchard-Mercier, A.; Rudkowska, I.; Paradis, A.-M.; Garneau, V.; Ouellette, C.; Lemieux, S.; Couture, P.; Vohl, M.C. Effects of age, sex, body mass index and APOE genotype on cardiovascular biomarker response to an n-3 polyunsaturated fatty acid supplementation. J. Nutrigenet. Nutrigenom. 2013, 6, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Van Eck, M.; Bos, I.S.T.; Hildebrand, R.B.; Van Rij, B.T.; Van Berkel, T.J.C. Dual role for scavenger receptor class B, type I on bone marrow-derived cells in atherosclerotic lesion development. Am. J. Pathol. 2004, 165, 785–794. [Google Scholar] [CrossRef]
- Ji, Y.; Jian, B.; Wang, N.; Sun, Y.; Moya, M.L.; Phillips, M.C.; Rothblat, G.H.; Swaney, J.B.; Tall, A.R. Scavenger receptor BI promotes high density lipoprotein-mediated cellular cholesterol efflux. J. Biol. Chem. 1997, 272, 20982–20985. [Google Scholar] [CrossRef] [PubMed]
- Yancey, P.G.; de la Llera-Moya, M.; Swarnakar, S.; Monzo, P.; Klein, S.M.; Connelly, M.A.; Johnson, W.J.; Williams, D.L.; Rothblat, G.H. High density lipoprotein phospholipid composition is a major determinant of the bi-directional flux and net movement of cellular free cholesterol mediated by scavenger receptor BI. J. Biol. Chem. 2000, 275, 36596–36604. [Google Scholar] [CrossRef] [PubMed]
- Yvan-Charvet, L.; Pagler, T.A.; Wang, N.; Senokuchi, T.; Brundert, M.; Li, H.; Rinninger, F.; Tall, A.R. SR-BI inhibits ABCG1-stimulated net cholesterol efflux from cells to plasma HDL. J. Lipid Res. 2008, 49, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Collins, H.L.; Ranalletta, M.; Fuki, I.V.; Billheimer, J.T.; Rothblat, G.H.; Tall, A.R.; Rader, D.J. Macrophage ABCA1 and ABCG1, but not SR-BI, promote macrophage reverse cholesterol transport in vivo. J. Clin. Investig. 2007, 117, 2216–2224. [Google Scholar] [CrossRef] [PubMed]
- Brundert, M.; Heeren, J.; Bahar-Bayansar, M.; Ewert, A.; Moore, K.J.; Rinninger, F. Selective uptake of HDL cholesteryl esters and cholesterol efflux from mouse peritoneal macrophages independent of SR-BI. J. Lipid Res. 2006, 47, 2408–2421. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Da Silva, J.R.; Reilly, M.; Billheimer, J.T.; Rothblat, G.H.; Rader, D.J. Hepatic expression of scavenger receptor class B type I (SR-BI) is a positive regulator of macrophage reverse cholesterol transport in vivo. J. Clin. Investig. 2005, 115, 2870–2874. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Geng, Y.; Xiao, N.; Yin, M.; Mao, L.; Ren, G.; Zhang, C.; Liu, P.; Lu, N.; An, L.; Pan, J. High dietary n-6/n-3 PUFA ratio promotes HDL cholesterol level, but does not suppress atherogenesis in apolipoprotein E-null mice 1. J. Atheroscler. Thromb. 2009, 16, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Mashurabad, P.C.; Kondaiah, P.; Palika, R.; Ghosh, S.; Nair, M.K.; Raghu, P. Eicosapentaenoic acid inhibits intestinal β-carotene absorption by downregulation of lipid transporter expression via PPAR-α dependent mechanism. Arch. Biochem. Biophys. 2016, 590, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Raposo, H.F.; Patrício, P.R.; Simões, M.C.; Oliveira, H.C.F. Fibrates and fish oil, but not corn oil, up-regulate the expression of the cholesteryl ester transfer protein (CETP) gene. J. Nutr. Biochem. 2014, 25, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Cheema, S.K.; Agarwal-Mawal, A.; Murray, C.M.; Tucker, S. Lack of stimulation of cholesteryl ester transfer protein by cholesterol in the presence of a high-fat diet. J. Lipid Res. 2005, 46, 2356–2366. [Google Scholar] [CrossRef] [PubMed]
- Michalik, L.; Wahli, W. Peroxisome proliferator-activated receptors: Three isotypes for a multitude of functions. Curr. Opin. Biotechnol. 1999, 10, 564–570. [Google Scholar] [CrossRef]
- Ishida, T.; Ohta, M.; Nakakuki, M.; Kami, H.; Uchiyama, R.; Kawano, H.; Notsu, T.; Imada, K.; Shimano, H. Distinct regulation of plasma LDL cholesterol by eicosapentaenoic acid and docosahexaenoic acid in high fat diet-fed hamsters: Participation of cholesterol ester transfer protein and LDL receptor. Prostaglandins Leukot. Essent. Fat. Acids 2013, 88, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Leigh-Firbank, E.C.; Minihane, A.M.; Leake, D.S.; Wright, J.W.; Murphy, M.C.; Griffin, B.A.; Williams, C.M. Eicosapentaenoic acid and docosahexaenoic acid from fish oils: Differential associations with lipid responses. Br. J. Nutr. 2002, 87, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Amanda Pepper-Yowell, M.S.; Ballou, M.; Trojan, S.; Thompson, L.; Moustaid-Moussa, N.; Wang, S.; Sheridan, M. The Role Docosapentaenoic Acid Omega-3 Plays in Controlling Lipid Metabolism and Inflammation Associated with Obesity, Atherosclerosis, and Neurodegenerative Disease. Ph.D. Thesis, Texas Tech University, Lubbock, TX, USA, 2014. [Google Scholar]
- Vasandani, C.; Kafrouni, A.I.; Caronna, A.; Bashmakov, Y.; Gotthardt, M.; Horton, J.D.; Spady, D.K. Upregulation of hepatic LDL transport by n-3 fatty acids in LDL receptor knockout mice. J. Lipid Res. 2002, 43, 772–784. [Google Scholar] [PubMed]
- Pan, M.; Cederbaum, A.I.; Zhang, Y.-L.; Ginsberg, H.N.; Williams, K.J.; Fisher, E.A. Lipid peroxidation and oxidant stress regulate hepatic apolipoprotein B degradation and VLDL production. J. Clin. Investig. 2004, 113, 1277–1287. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.; Maitin, V.; Parathath, S.; Andreo, U.; Lin, S.X.; St. Germain, C.; Yao, Z.; Maxfield, F.R.; Williams, K.J.; Fisher, E.A. Presecretory oxidation, aggregation, and autophagic destruction of apoprotein-B: A pathway for late-stage quality control. Proc. Natl. Acad. Sci. USA 2008, 105, 5862–5867. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, D.; Witztum, J.L. Is the Oxidative Modification Hypothesis Relevant to Human Atherosclerosis? Circulation 2002, 105, 2107–2111. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.G.; Zhao, X.-Q.; Chait, A.; Fisher, L.D.; Cheung, M.C.; Morse, J.S.; Dowdy, A.A.; Marino, E.K.; Bolson, E.L.; Alaupovic, P.; et al. Simvastatin and Niacin, Antioxidant Vitamins, or the Combination for the Prevention of Coronary Disease. N. Engl. J. Med. 2001, 345, 1583–1592. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.G.; Cheung, M.C.; Lee, A.C.; Zhao, X.-Q.; Chait, A. Antioxidant Vitamins and Lipid Therapy. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Bérard, A.M.; Dumon, M.-F.; Darmon, M. Dietary fish oil up-regulates cholesterol 7α-hydroxylase mRNA in mouse liver leading to an increase in bile acid and cholesterol excretion. FEBS Lett. 2004, 559, 125–128. [Google Scholar] [CrossRef]
- Jakulj, L.; Besseling, J.; Stroes, E.S.G.; Groen, A.K. Intestinal cholesterol secretion: Future clinical implications. Neth. J. Med. 2013, 71, 459–465. [Google Scholar] [PubMed]
- Alvaro, A.; Rosales, R.; Masana, L.; Vallvé, J.-C. Polyunsaturated fatty acids down-regulate in vitro expression of the key intestinal cholesterol absorption protein NPC1L1: No effect of monounsaturated nor saturated fatty acids. J. Nutr. Biochem. 2010, 21, 518–525. [Google Scholar] [CrossRef] [PubMed]
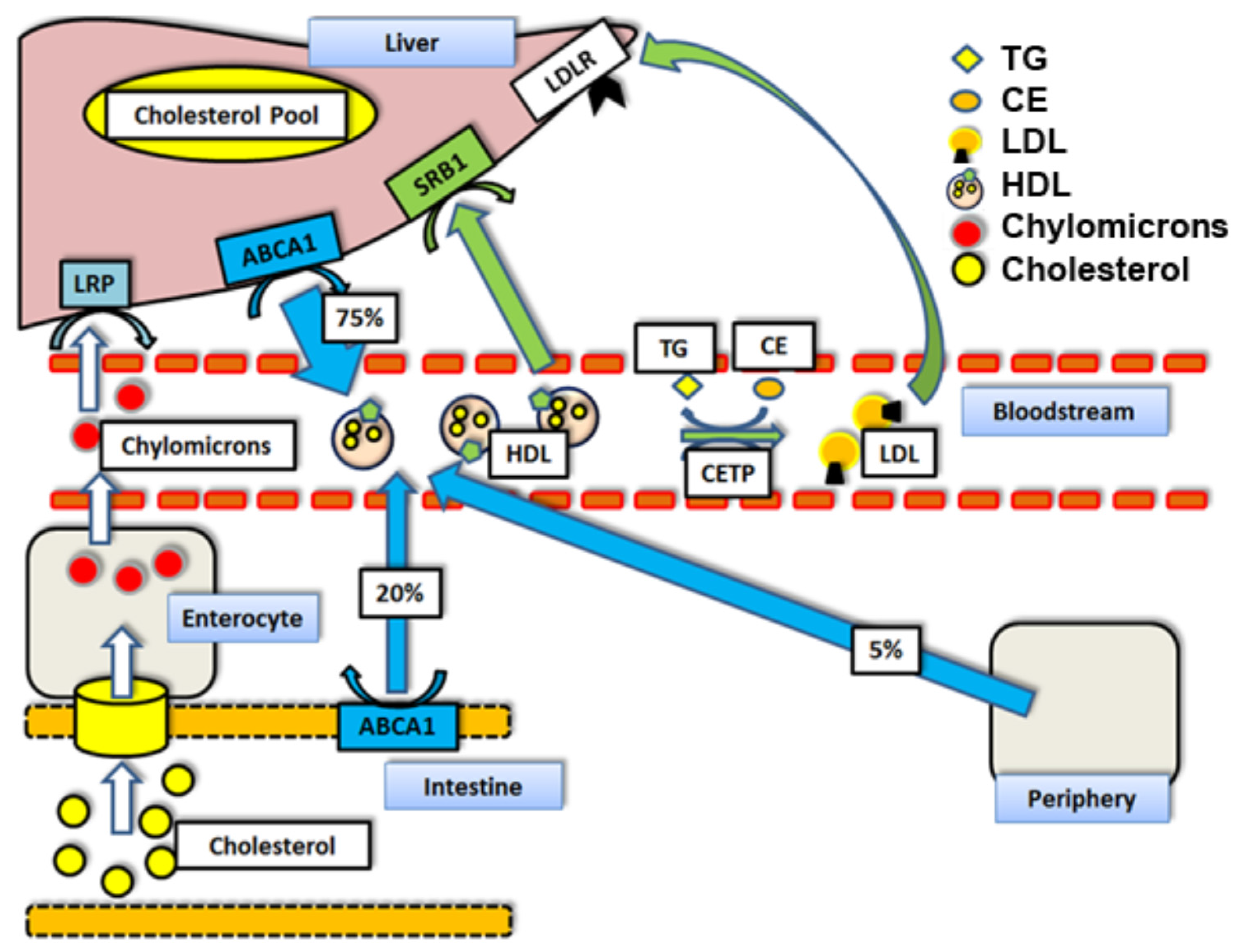
Figure 1.
Reverse Cholesterol Transport mechanisms and respective contributions to HDL cholesterol loads by percent [10]: Abbreviations: TG = triglycerides. HDL = high density lipoprotein particle. CETP = cholesteryl ester transfer protein. LDL = low density lipoprotein particle. ABCA1 = ATP binding cassette transporter A1. LDLr = Low density lipoprotein receptor. LRP = LDLr receptor-related protein. SR-BI = scavenger receptor class B type I. CE = cholesteryl ester.
Figure 1.
Reverse Cholesterol Transport mechanisms and respective contributions to HDL cholesterol loads by percent [10]: Abbreviations: TG = triglycerides. HDL = high density lipoprotein particle. CETP = cholesteryl ester transfer protein. LDL = low density lipoprotein particle. ABCA1 = ATP binding cassette transporter A1. LDLr = Low density lipoprotein receptor. LRP = LDLr receptor-related protein. SR-BI = scavenger receptor class B type I. CE = cholesteryl ester.
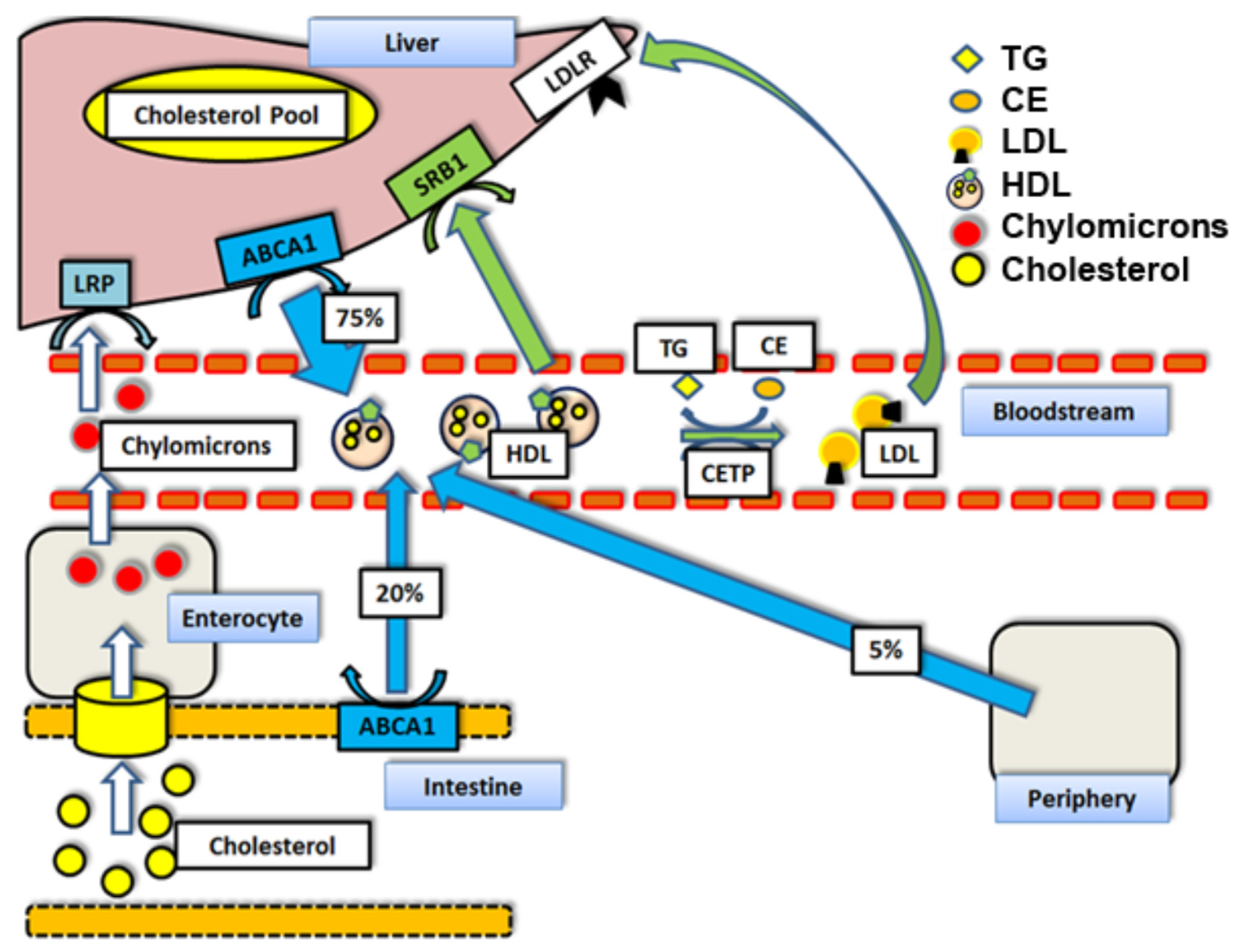
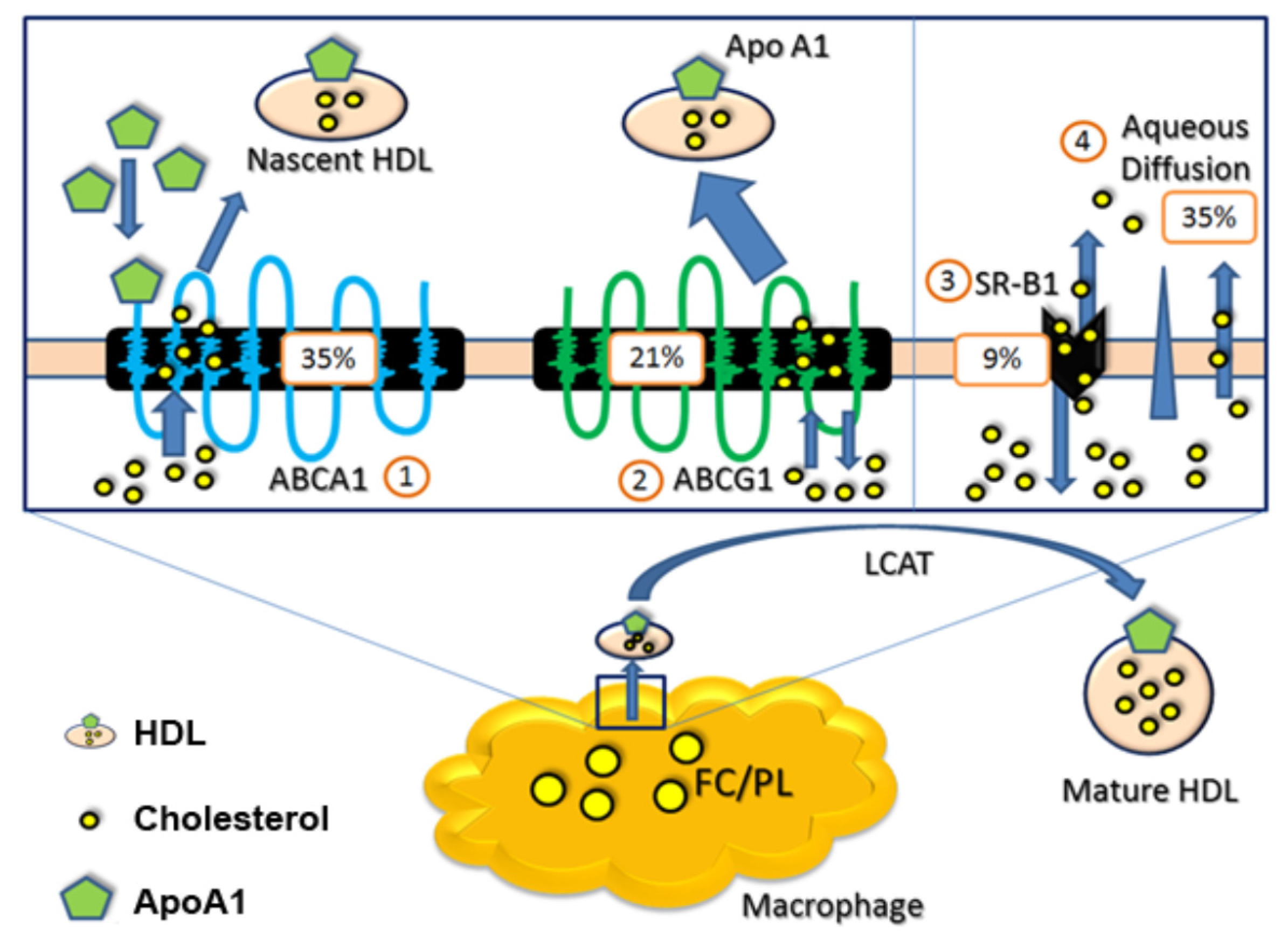
Figure 2.
Macrophage cholesterol efflux routes and relative contributions to cholesterol efflux by percent [10]. Abbreviations: ABCA1 = ATP-binding cassette transporter A1, ABCG1 = ATP-binding cassette transporter G1, apo A1 = apolipoprotein A1, FC/PL = free cholesterol/phospholipids, HDL = high density lipoprotein, SR-BI = scavenger receptor class B type I. LCAT = lecithin cholesterol acyl transferase.
Figure 2.
Macrophage cholesterol efflux routes and relative contributions to cholesterol efflux by percent [10]. Abbreviations: ABCA1 = ATP-binding cassette transporter A1, ABCG1 = ATP-binding cassette transporter G1, apo A1 = apolipoprotein A1, FC/PL = free cholesterol/phospholipids, HDL = high density lipoprotein, SR-BI = scavenger receptor class B type I. LCAT = lecithin cholesterol acyl transferase.

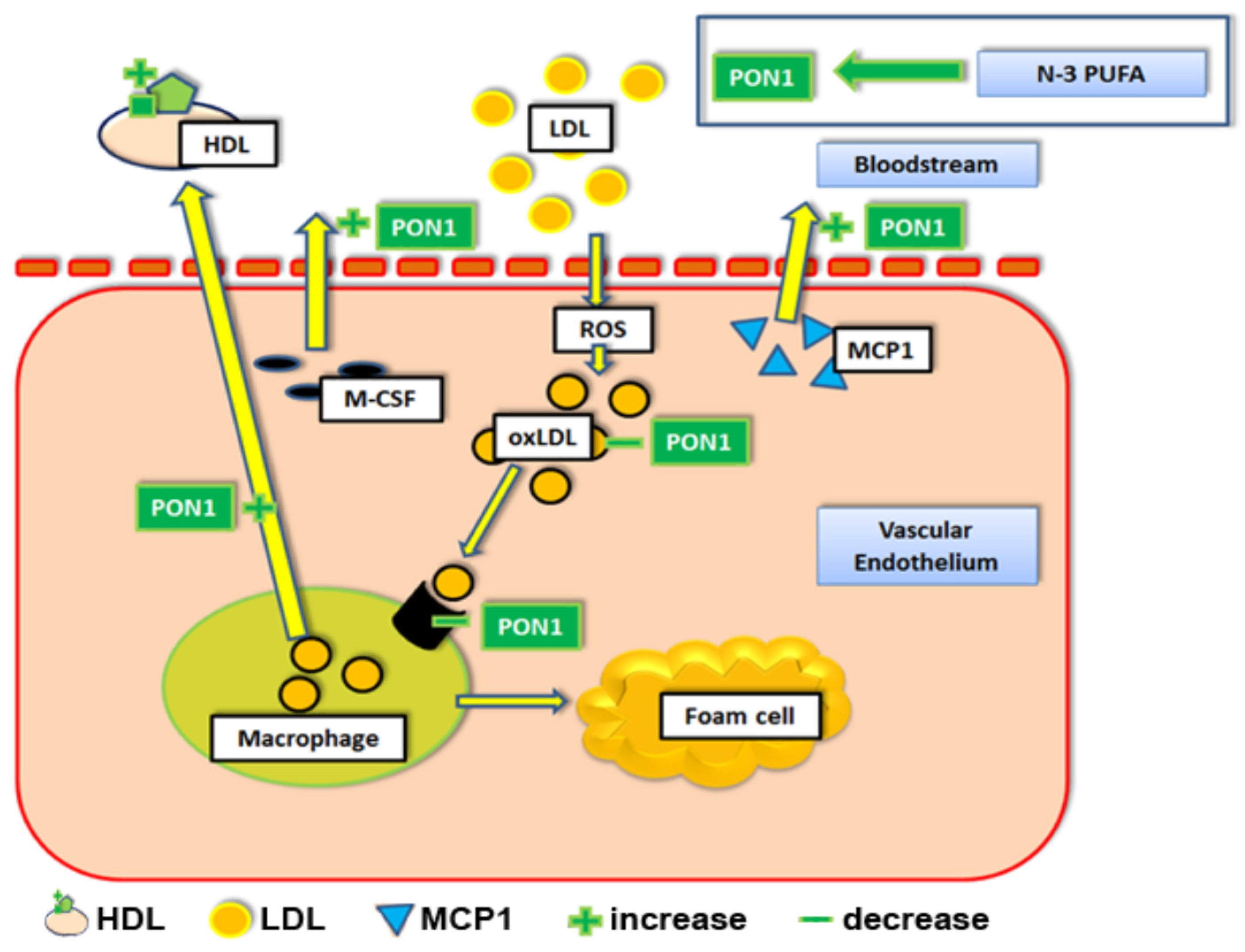
Figure 3.
The effects of n-3 PUFAs on paraoxonase 1 and cholesterol efflux. Abbreviations: HDL = high density lipoprotein particle. LDL = low density lipoprotein. PON1 = paraoxonase 1. MCP1 = Monocyte chemotactic protein 1. ROS = reactive oxygen species. M-CSF = macrophage colony stimulating factor. oxLDL = oxidized LDL. n-3 PUFA = Omega 3 polyunsaturated fatty acid.
Figure 3.
The effects of n-3 PUFAs on paraoxonase 1 and cholesterol efflux. Abbreviations: HDL = high density lipoprotein particle. LDL = low density lipoprotein. PON1 = paraoxonase 1. MCP1 = Monocyte chemotactic protein 1. ROS = reactive oxygen species. M-CSF = macrophage colony stimulating factor. oxLDL = oxidized LDL. n-3 PUFA = Omega 3 polyunsaturated fatty acid.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pizzini, A.; Lunger, L.; Demetz, E.; Hilbe, R.; Weiss, G.; Ebenbichler, C.; Tancevski, I. The Role of Omega-3 Fatty Acids in Reverse Cholesterol Transport: A Review. Nutrients 2017, 9, 1099. https://doi.org/10.3390/nu9101099
AMA Style
Pizzini A, Lunger L, Demetz E, Hilbe R, Weiss G, Ebenbichler C, Tancevski I. The Role of Omega-3 Fatty Acids in Reverse Cholesterol Transport: A Review. Nutrients. 2017; 9(10):1099. https://doi.org/10.3390/nu9101099
Chicago/Turabian StylePizzini, Alex, Lukas Lunger, Egon Demetz, Richard Hilbe, Guenter Weiss, Christoph Ebenbichler, and Ivan Tancevski. 2017. "The Role of Omega-3 Fatty Acids in Reverse Cholesterol Transport: A Review" Nutrients 9, no. 10: 1099. https://doi.org/10.3390/nu9101099
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.