Resveratrol and Amyloid-Beta: Mechanistic Insights


1
Department of Neuropharmacology, Research Institute of Medicine and Pharmacy, Qiqihar Medical University, Qiqihar 161006, China
2
Department of Pathophysiology, Qiqihar Medical University, Qiqihar 161006, China
*
Author to whom correspondence should be addressed.
Nutrients 2017, 9(10), 1122; https://doi.org/10.3390/nu9101122
Submission received: 19 August 2017
/
Revised: 25 September 2017
/
Accepted: 2 October 2017
/
Published: 14 October 2017
(This article belongs to the Special Issue Effects of Resveratrol Supplementation on Human Health and Disease)
Abstract
:The amyloid-beta (Aβ) hypothesis that dyshomeostasis between Aβ production and clearance is a very early, key molecular factor in the etiology of Alzheimer’s disease (AD) has been proposed and examined in the AD research field. Scientists have focused on seeking natural products or drugs to influence the dynamic equilibrium of Aβ, targeting production and clearance of Aβ. There is emerging evidence that resveratrol (Res), a naturally occurring polyphenol mainly found in grapes and red wine, acts on AD in numerous in vivo and in vitro models. Res decreases the amyloidogenic cleavage of the amyloid precursor protein (APP), enhances clearance of amyloid beta-peptides, and reduces Aβ aggregation. Moreover, Res also protects neuronal functions through its antioxidant properties. This review discusses the action of Res on Aβ production, clearance and aggregation and multiple potential mechanisms, providing evidence of the useful of Res for AD treatment.
1. Introduction
Alzheimer’s disease (AD) is considered the most frequent form of dementia, affecting more than 35.6 million people worldwide and becoming an emerging burden on society [1,2]. Although researchers have delved into all aspects of this complex, multifactorial syndrome in recent years, the exact molecular events underlying AD remain to be exhaustively elucidated. The amyloid-beta (Aβ) hypothesis which synthesizes histopathological and genetic information has become the dominant model of AD pathogenesis and is guiding the development of a novel strategy in the treatment of AD [3]. The pathological accumulation of Aβ in the brain leads to impairment of synaptic function and structure, progressive tau deposition, neuronal destruction and finally the clinical symptoms of AD [3]. Aβ homeostasis in the brain is governed by its production and clearance mechanisms [4]. Aβ was produced by sequential scission of amyloid precursor protein (APP) by the β-APP cleaving enzyme (BACE) and γ-secretase [5]. Numerous pathways have been reported to play roles in Aβ clearance and degradation, including autophagic pathway and receptor-mediated endocytosis [6]. Some proteins with a crucial role in Aβ clearance are Aβ-degrading enzymes (ADEs), which could degrade or cleave Aβ into smaller fragments, and lipoprotein receptor-related protein 1 (LRP1), which regulates metabolism of Aβ and brain homeostasis through multiple pathways and transporters such as P-glycoprotein (P-gp), localized in astrocytes and on the abluminal side of the cerebral endothelium, where they transport Aβ from brain to blood [7,8,9]. So some researchers focused on screening drugs which can influence the dynamic equilibrium of Aβ, targeting production and clearance of Aβ.
Resveratrol (3,5,4’-trihydroxy-trans-stilbene, Res), a naturally polyphenolic phytoalexin, abundantly found in grapes, berries, red wine and many other plant species, shows diverse biological activities such as antioxidant, anticarcinogenic and anti-inflammatory properties, and so on [10]. Recent evidence found that, in various in vitro and in vivo models of AD, Res plays a prominent role in the prevention and treatment of AD [11,12]. Res attenuated Aβ-induced cytotoxicity, apoptosis and intracellular reactive oxygen intermediate (ROI) accumulation in PC12 cells [13]. Similarly, Res inhibits Aβ-induced neuronal apoptosis in PC12 cells through regulation of silent information regulator 1 (SIRT1)-Rho-associated kinase 1 (ROCK1) signaling pathway [14]. In Res-treated APP/PS1 mice which are a model of cerebral Aβ deposition, there was a significant reduction in the number of activated microglia, providing anti-inflammatory effects of Res against Aβ-triggered microglial activation [15]. It is interesting that Aβ accumulation in the brain has been shown to induce cytotoxicity, apoptosis and activation of astrocytes in cell and animal models [16,17,18]. So we explore whether Res can affect Aβ accumulation and aggregation in AD model (Table 1 and Table 2). In this review, we discuss the effects of Res on Aβ production and clearance and clarify those multiple mechanisms in various models and explore Res as a new drug candidate in AD treatment.
2. The Source and Pharmacological Profile of Res
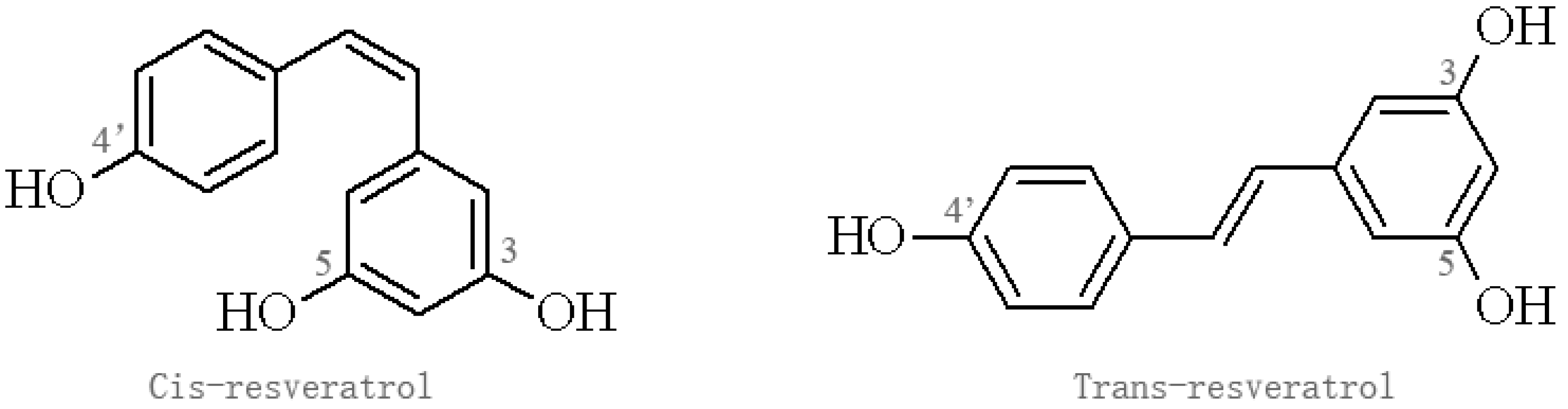
Res belongs to the stilbene family of phytoalexins produced by various plants in response to environmental stress, which have been detected in at least 72 plant species, especially grape vines, pine trees, jackfruit, blueberry, cranberry, and mulberry [19,20]. Res containing two aromatic rings in its structure exists in two isomers: trans-Res, which has been most widely studied, and cis-Res; the trans form is more stable and potent than the cis form, which may have different biological effects (Figure 1) [21]. Res has diverse biological activities and is not known to cause significant adverse effects in experimental animals and humans [22,23,24]. A clinical trial found that oral Res at a single dose of 5 g in 10 healthy volunteers resulted in no apparent adverse effects, and the peak level of Res in plasma was 539 ng/mL, while plasma concentration of two Res monoglucuronides and Res 3-sulfate were 1285 ng/mL, 1735 ng/mL, 4294 ng/mL, respectively [22]. Similarly, a double-blind, randomized, placebo-controlled study showed that Res is safe in healthy individuals, but produced a relatively low plasma concentration. Res exhibits low oral bioavailability due to its rapid metabolism and elimination, which is rapidly metabolized by conjugation to glucuronic acid and/or sulfate, forming Res glucuronides, sulfates, and/or sulfoglucuronides [25]. Thus, Res analogues and inhibitors affecting Res metabolism were developed to enhance its bioavailability.
3. Effect of Res on Aβ Production
3.1. Inhibitory Activity against BACE1
BACE1, mainly expressed in neurons of the brain, exhibited all the known characteristics of the β-secretase, which cleave extracellular of APP important in the pathogenesis of AD [36]. Tamagno et al. reported that Aβ induces production of oxidative stress markers 4-hydroxynonenal and upregulates BACE-1 expression in NT2 neuronal cells, further fostering amyloidogenic processing of APP, thereby increasing accumulation of Aβ [37]. Scientists hope to find some active compounds or develop new drugs to block BACE1 (BACE inhibitors), further decreasing Aβ accumulation, regarding BACE as a key target for AD treatment. Studies showed that Res and Res oligomers significantly inhibited BACE activity in a dose dependent manner, which was assessed by fluorescence resonance energy transfer (FRET) assay [30]. Similarly, Koukoulitsa et al. found that Res and its derivatives bearing one (tert-butyl, 1-ethylpopyl) or two bulky electron donating groups ortho to 4’-OH displayed different potencies against BACE1 using time-resolved fluorescence (TRF) assay, further suggesting that Res can inhibit BACE1 function [31,38]. In contrast, Marambaud et al. found that Res does not inhibit Aβ production through neither affecting β- nor γ-secretases [39]. The paradoxical results may be caused by different methods and cells, which need further studies. Some potent BACE1 inhibitors is now undergoing clinical trials such as E2609, AZD3293 and LY2886721, and some adverse effects also reported [40]. Finding active components such as Res in herbs targeting β-secretases could be a strategy in AD treatment.
3.2. Inhibitory Activity against γ-Secretase
γ-Secretases are a family of intramembrane cleaving aspartyl proteases, consisting of four subunits presenilin (PSEN), anterior pharynx defective-1, nicastrin, and presenilin enhancer-2 [41]. When the expression and activity of those four subunits were changed, the catalytic function of γ-secretases could also be changed [42]. Recent evidence showed that rats fed with Res, a SIRT1 inducer, exhibit a significant increase in PSEN1 expression, which is one of SIRT1-specific DNA targets [26]. Unfortunately, the results did not show whether the activity of γ-secretase was changed and Aβ production could be suppressed by SIRT1 [26]. Choi et al. found that Res (5–20 μM) as well as its analogues inhibit γ-secretase activity and increase α-secretase activity in Neuro2a neuroblastoma cells, which may be associated with a decrease in Aβ levels, without causing cell death [32]. However, another study showed that Res has no effect on γ-secretases, without affecting Aβ production and APP metabolism [39].
3.3. Autophagy Induction
Emerging evidence indicated that autophagy pathways could affect developmental and neurodegenerative processes, including AD. AD etiology is also associated with damaged mitochondria in the neurons. Misfolded proteins translocated and accumulated into the mitochondrial membrane, leading to the disruption of oxidative phosphorylation and then autophagy activation [43]. Accordingly, neuronal autophagy is known to play key roles in synaptic plasticity, anti-inflammatory function in glial cells, oligodendrocyte development, and myelination process [44]. Studies found that a decline in autophagy efficiency during aging led to accumulation of Aβ and cytochrome c release in the mitochondrial membrane, resulting cell death and neurodegeneration [45,46]. Moreover, accumulation of pathological autophagic vacuoles can be observed in PS1/APP mouse AD model [47]. Imbalance between autophagic flux and degradation result in decreased proteolysis of Aβ [48]. In AD, autophagy impairment stimulates PSEN1 expression and then increases γ-secretase activity, leading to augment of Aβ synthesis [49]. Res, an autophagy inducer, decreased PSEN1 expression, consistent with a suppression of Aβ production [33]. Moreover, Res can activate tyrosyl transferRNA (tRNA) synthetase (TyrRS)-auto-poly-ADP-ribosylation of poly (ADP-ribose) polymerase 1 (PARP1)-SIRT1 signaling pathway, inducing autophagy in PC12 cells, thereby attenuating neurotoxicity caused by Aβ [50]. Res have the therapeutic potential of AD through activating autophagy.
4. Effect of Res on Aβ Clearance
Metabolic pathway of Aβ in AD pathogenesis, especially in late-onset sporadic AD (LOAD) has been raised, and the main mechanisms of Aβ clearance have been considered as new therapeutic targets [51,52]. Res could modulate Aβ clearance in various ways such as activation of ADEs, transport across the blood-brain barrier (BBB) into the circulation, uptake by microglial phagocytosis and inhibition of Aβ aggregation [53,54].
4.1. Activation of ADEs
Disturbances in the activity of ADEs, including neprilysin (NEP), insulin-degrading enzyme (IDE), angiotensin-converting enzyme (ACE), endothelin-converting enzyme (ECE) and plasmin, could induce Aβ accumulation, resulting in AD pathology [55,56]. Activity of ADEs could be affected by Res in different AD models. In vitro studies, NEP and ACE activities were induced effectively in SK-N-SH cells with low concentrations of Res for 4 days [34]. The authors found that Res enhances the differentiation state of proliferating cells, which correlated with up-regulation of the cellular enzymatic activity to enhance NEP and ACE activity [34]. Similarly, Marambaud found that NEP activity was significantly increased after Res treatment on intact HEK293 cells. However, proteasomal degradation pathway might involve in Res-mediated decrease of Aβ but not the increased NEP activity induced by Res [16]. Moreover, study found that in human endothelial cells, Res decreased ECE-1 mRNA expression and reduced ECE-1 activity, further affecting endothelin synthesis and conversion; however, incubation concentration of Res (30 μM) cannot achieve in rat and/or human plasma [57]. Although ECE-1 activity can be inhibited by Res, it cannot induce Aβ accumulation in brain, leading to AD. In vivo study, Res could dramatically increase both the estradiol level and NEP level in LPS-treated rats and control rats using NEP ELISA kit [27]. Those results indicated that Res can involve in activation of ADEs, but whether Res promotes clearance of Aβ through ADEs needs further studies.
4.2. Plasminogen System
The plasminogen system is a complex enzymatic cascade [58]. Plasmin (PL), degrading many plasma proteins, is an important serine protease released from the inactive zymogen plasminogen (PLG) by two physiological activators tissue-type plasminogen activator (t-PA) and urokinase-type plasminogen activator (u-PA) [59]. Function of t-PA and u-PA were controlled by PA inhibitor type 1 (PAI-1) and type 2 (PAI-2) [60]. PL is involved in many pathophysiological processes through its ability to cleave fibrin, fibronectin, thrombospondin, laminin, and von Willebrand factor. The plasminogen system has been implicated in neuronal plasticity and long-term potentiation (LTP) in the brain [61]. Studies showed that tPA-plasmin system contributes to the clearance of Aβ in mouse brain and the prevention of Aβ-induced neurotoxicity, suggesting that the plasminogen system is associated with late-onset AD [62]. Similarly, polymorphisms in the u-PA gene have been associated with AD susceptibility [63]. Res increased plasminogen activators t-PA and u-PA expression in cultured human umbilical vein endothelial cells (HUVECs), leading to plasminogen endoproteolysis and plasmin activation.
4.3. Neurovascular Pathways
The neurovascular unit (NVU) consists of different cell types, including brain endothelial cells, glial cells and neurons, which controls BBB permeability, neurovascular coupling and clearance of toxins such as Aβ from brain [64]. BBB permeability controls entry from blood and promotes clearance of macromolecules from the brain [64]. Moreover, transporters and receptors in the brain endothelium regulate delivery of drugs and endogenous substances in the brain. Studies have shown that Aβ is a substrate of LRP1, the receptor for advanced glycation end products (RAGE) and several ABC transporters such as P-gp and Breast cancer resistance protein (BCRP) [65,66,67,68]. Under physiological conditions, vascular damage and changes in the expression of several BBB transporters and receptors could affect Aβ transport from blood-to-brain and/or brain-to-blood. Res, a novel transporter modulator, defends BBB integrity and changes transporter and receptors expressions in BBB, thereby regulating Aβ homeostasis [28,65]. We will focus on effect of Res on: (1) integrity of BBB, which regulates delivery of energy metabolites and essential nutrients; (2) ABC transporters in brain that mediate Aβ efflux into circulation from brain; (3) LRP1 mediating Aβ efflux from brain; and (4) RAGE, which mediates Aβ reentry into the brain from circulation and the neurovascular inflammatory response.
4.3.1. BBB Integrity
Several studies have suggested that injured BBB integrity and cerebrovascular dysfunction lead to faulty Aβ clearance from the brain [69]. Res could protect BBB integrity in different animal models [70]. In ovariectomized + D-galactose induced rat model of AD, Res decreases the insoluble Aβ42 level and protects the BBB integrity through regulating the expressions of RAGE, matrix metalloprotein-9 (MMP-9) and Claudin-5 [29]. In clinic, Res appears to restore BBB integrity in AD patients, reducing the ability of harmful immune molecules secreted by immune cells to infiltrate from the body [71]. Similarly, Res has exhibited restoration of BBB integrity via reduction of MMP-9 and induce adaptive immune responses which may promote brain resilience to Aβ deposition in AD patients [72]. Those studies suggested that Res could protect BBB integrity, changing Aβ homeostasis.
4.3.2. P-gp
In vitro and in vivo studies showed that eliminated P-gp, first ABC transporter detected in endothelial cells of the human BBB lead to Aβ accumulation [9,73]. Furthermore, P-gp activity at the BBB is reduced in AD individuals [74], suggesting that impaired P-gp activity may mediate cerebral Aβ accumulation. Our previous studies indicated that Res enhanced bestatin absorption by downregulating P-gp expression in Caco-2 cells [10]. Moreover, Res and its major metabolites could penetrate BBB and be measured in Cerebrospinal fluid (CSF) in AD patients [75], at least indicating that Res has the opportunity to have CNS effects. Unfortunately, no effects of Res treatment on Aβ42 level in plasma and CSF has been detected [75], and no research has yet investigated whether P-gp expression at the BBB is changed by Res. A larger study is required to determine whether Res can affect Aβ clearance through regulating P-gp.
4.3.3. LRP1
LRP1, one receptor in the LDL receptor family, is not only a multifunctional scavenger and cargo transporter but also has signal transduction activity [76]. As a cargo transporter, some studies have reported that LRP1 transported several ligands from brain to blood including Aβ [77,78]. Moreover, several genetic studies have indicated that LRP1 expression at the BBB is reduced in AD and have been considered as a therapeutic target in AD [79,80]. After treatment with Res in AD transgenic female mice, the brain LRP1 protein expression was increased, while its mRNA was not changed [28]. The author further found that Res could upregulate and stabilize transthyretin (TTR) which binds Aβ peptide avoiding its aggregation and toxicity, resulting increased LRP1 [28]. In vitro studies, LRP1 mediated the cytotoxicity of the fibrillar Aβ oligomers following their binding to prion protein (PrPC), and Res could reduce PrPC and LRP1-mediated binding in SH-SY5Y cells, remodeling the fibrillar Aβ oligomer conformation [81].
4.3.4. RAGE
RAGE, a multiligand receptor of the immunoglobulin superfamily of cell surface molecules, binds distinct classes of ligands such as AGE proteins and Aβ [82]. RAGE expressed at the luminal side of the BBB transport Aβ from the blood into the brain. In addition, intraneuronal transport of Aβ via neuronal RAGE leads to mitochondrial dysfunction [83]. RAGE expression is increased in the AD capillary under pathological conditions. Res has been shown to reduce RAGE expression in vascular cells [84]. Scientists are now comfortable referring to AD as type 3 diabetes, which results from insulin resistance in the brain. Similarly, Res can also downregulate RAGE expression in the kidney and liver of rats with type 2 diabetes [85,86]. However, whether RAGE expression triggered by Res could affect Aβ uptake, leading to Aβ accumulation in brain needs further research. Given the important role of RAGE in Aβ accumulation in AD, finding drugs which can block RAGE may contribute to control of Aβ-mediated brain disorder.
5. Aβ Plaque Disruption
Aggregation of Aβ leads to activation of microglia and astrocytes, and loss of cholinergic neurons, which is a constant feature of AD [87]. Res not only plays an important role in affecting Aβ homeostasis, but can also inhibit Aβ aggregation from lower molecular weight oligomers into higher molecular weight oligomers and disrupt preformed Aβ aggregation [54,88]. One study found that Res could bind directly to Aβ in different states including monomer and fibril Aβ [35]. Additionally, from indirect inhibitory effect, Res could enhance the binding of Aβ oligomers and TTR which can stabilize Aβ oligomers structure, preventing plaque aggregation, which provides new perspective into the protective properties of Res against AD [28,89].
6. Res Analogs in AD Treatment
Res analogs which show better bioavailability, efficacy and stability compared to Res are being tested for their activity in relation to many degenerative conditions. Some studies showed that Res derivatives exhibit neuroprotective effects in vivo and in vitro models [90]. Compounds 5d, a3, 5-dimethoxyl derivatives of Res, can cross BBB in vitro and are the potent inhibitor of Aβ42 aggregation, disintegration of highly structured with low neurotoxicity [91]. Similarly, Piceatannol, a metabolite of Res found in red wine, protects Aβ-induced neural cell death through affecting the accumulation of reactive oxygen species (ROS) induced by Res in PC12 cells [90]. Pterostilbene is a stilbenoid chemically related to Res and is a potent modulator of cognition and cellular stress, associated with regulating peroxisome proliferator-activated receptor alpha (PPARα) protein expression [92]. Various Res analogs and derivatives developed with improved bioavailability possess neuroprotective activities and could be promising drug candidates in the treatment of AD.
7. Conclusions and Challenges
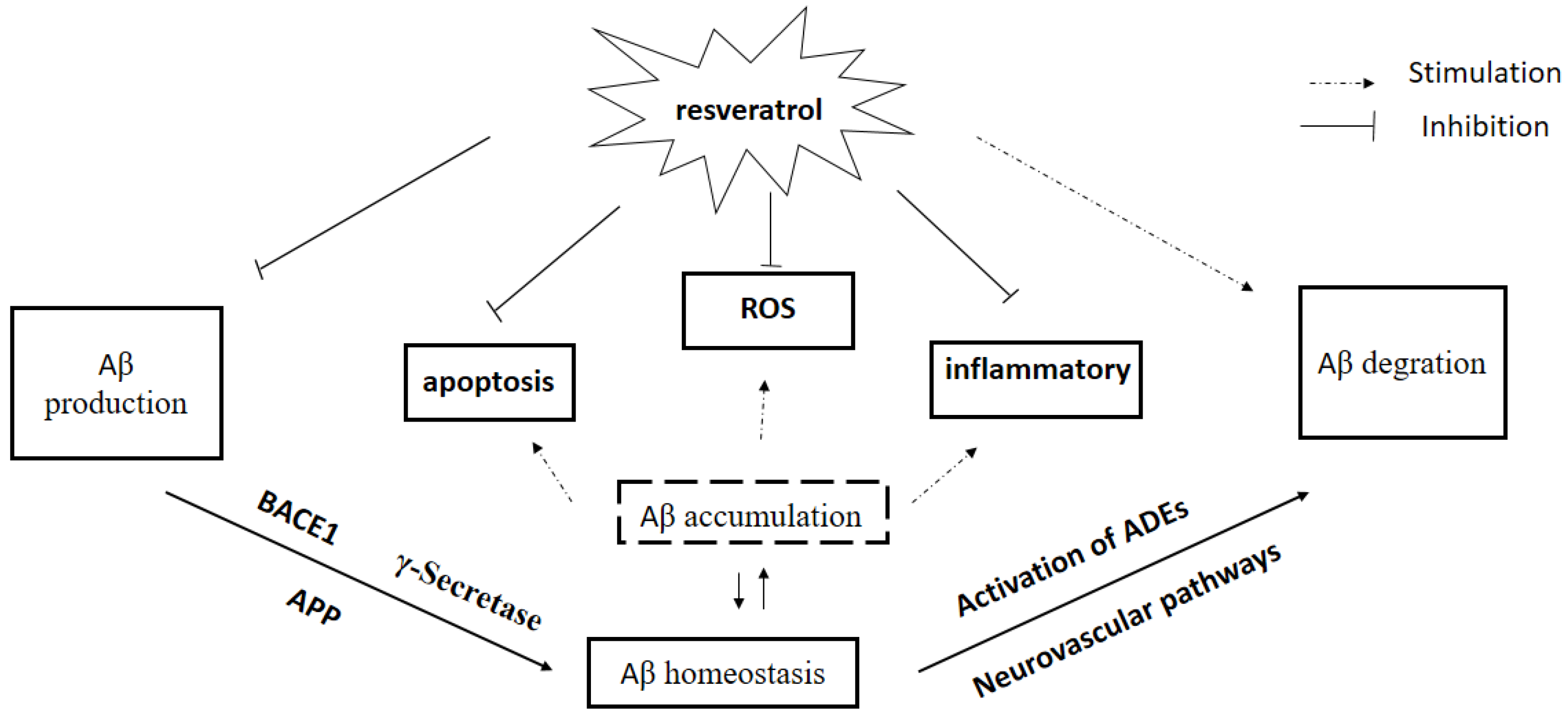
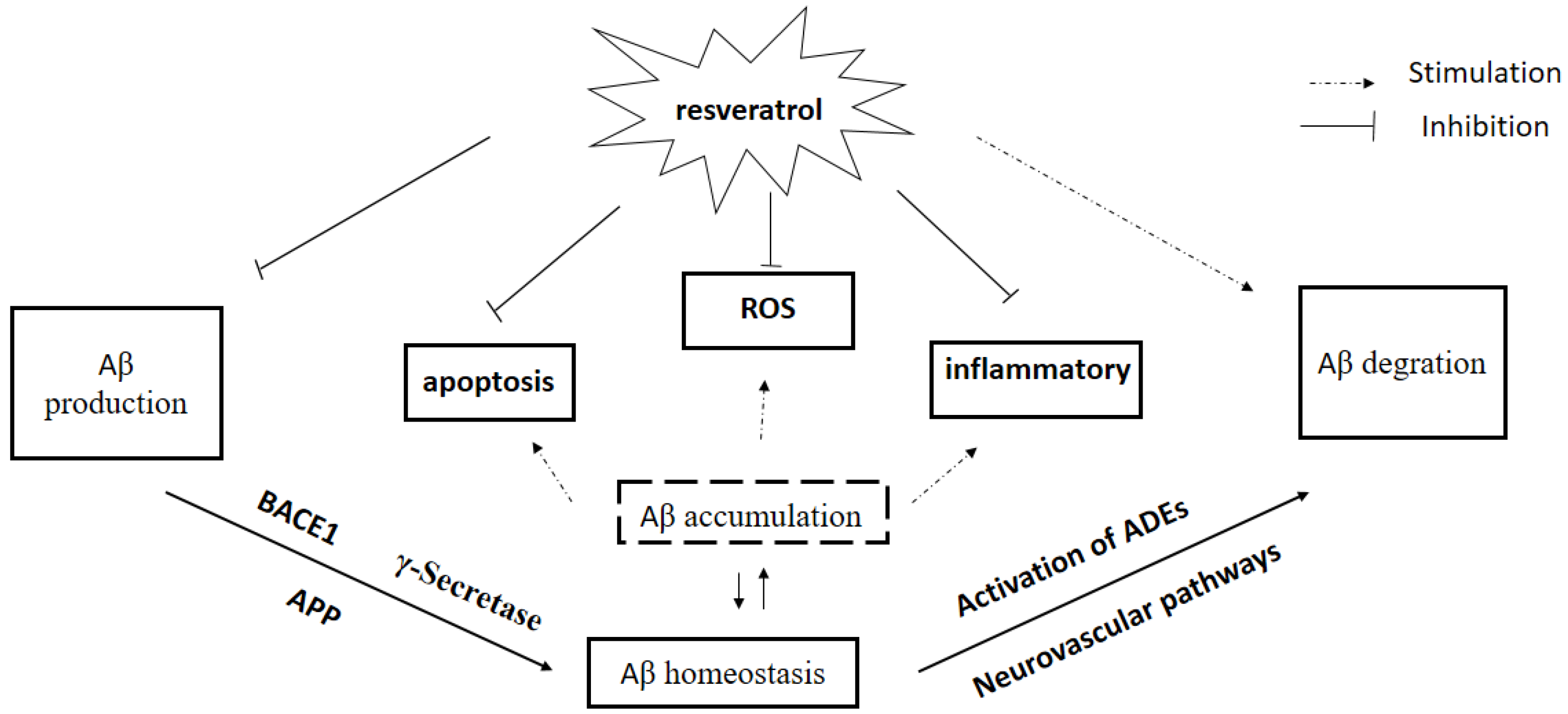
Recent observations provide strong evidence for the link between Aβ accumulation in the brain and AD, and the role of Aβ clearance pathway in AD. There is an urgent need to search for and develop disease-modifying natural products and drugs to treat the major neurodegenerative disorders [93]. In this chapter, we have briefly reviewed the literature on effects of Res, a new rising star in AD treatment, on Aβ production and Aβ clearance, BBB dysfunction, and Aβ plaque disruption, supporting an essential role of Res on Aβ homeostasis in AD pathogenesis (Figure 2). In addition to directly regulating Aβ homeostasis in brain, several in vitro studies showed that Res is believed to afford strong antioxidative and anti-inflammatory properties induced by Aβ, indirectly. Res attenuated Aβ-induced oxidative stress in vivo and in vitro [94,95], suggesting that Res may be of benefit in AD treatment. Clinical trials indicated that Res seems to be well tolerated with less toxicity and can penetrate BBB easily. However, low oral bioavailability of Res limited its clinical efficacy because of rapid excretion and extensive metabolism [96]. Numerous clinical trials tried to investigate the effects of Res on neurodegenerative diseases including AD, although there are many difficulties such as bioavailability and side effects [97]. Addressing these questions will lead to a better understanding of mechanisms of Res on AD treatment, which will contribute to searching for, developing and designing new drugs for AD.
Acknowledgments
We thank the financial support of the Nursing Program for Young Scholars of Heilongjiang Province of China (No. UNPYSCT-2016116), Natural Science Foundation of China (No. 81303248) and the Natural Science Foundation of Heilongjiang Province of China (No. H2015028).This research was in part supported by Item of Scientific Research Fund for Doctor of Qiqihar Medical University (QY2016B-09).
Author contributions
Yongming Jia designed research, searched literatures, wrote the paper and revised the paper. Na Wang searched literatures. Xue-wei Liu revised the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Van Wijngaarden, P.; Hadoux, X.; Alwan, M.; Keel, S.; Dirani, M. Emerging ocular biomarkers of Alzheimer disease. Clin. Exp. Ophthalmol. 2017, 45, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Tarawneh, R.; Holtzman, D.M. The clinical problem of symptomatic Alzheimer disease and mild cognitive impairment. Cold Spring Harb. Perspect. Med. 2012, 2, a006148. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, A.; Nelson, A.R.; Sagare, A.P.; Zlokovic, B.V. Impaired vascular-mediated clearance of brain amyloid β in Alzheimer’s disease: The role, regulation and restoration of LRP1. Front. Aging Neurosci. 2015, 7, 136. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, R.J.; Wong, P.C. Amyloid precursor protein processing and Alzheimer’s disease. Annu. Rev. Neurosci. 2011, 34, 185–204. [Google Scholar] [CrossRef] [PubMed]
- Ries, M.; Sastre, M. Mechanisms of aβ clearance and degradation by glial cells. Front. Aging Neurosci. 2016, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, M.; Tachibana, M.; Kanekiyo, T.; Bu, G. Role of LRP1 in the pathogenesis of Alzheimer’s disease: Evidence from clinical and preclinical studies. J. Lipid Res. 2017, 58, 1267–1281. [Google Scholar] [CrossRef] [PubMed]
- Jha, N.K.; Jha, S.K.; Kumar, D.; Kejriwal, N.; Sharma, R.; Ambasta, R.K.; Kumar, P. Impact of insulin degrading enzyme and neprilysin in Alzheimer’s disease biology: Characterization of putative cognates for therapeutic applications. J. Alzheimers Dis. 2015, 48, 891–917. [Google Scholar] [CrossRef] [PubMed]
- Van Assema, D.M.; van Berckel, B.N. Blood-brain barrier ABC-transporter P-glycoprotein in Alzheimer’s disease: Still a suspect? Curr. Pharm. Des. 2016, 22, 5808–5816. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Liu, Z.; Huo, X.; Wang, C.; Meng, Q.; Liu, Q.; Sun, H.; Sun, P.; Yang, X.; Shu, X.; et al. Enhancement effect of resveratrol on the intestinal absorption of bestatin by regulating PEPT1, MDR1 and MRP2 in vivo and in vitro. Int. J. Pharm. 2015, 495, 588–598. [Google Scholar] [CrossRef] [PubMed]
- Rege, S.D.; Geetha, T.; Griffin, G.D.; Broderick, T.L.; Babu, J.R. Neuroprotective effects of resveratrol in Alzheimer disease pathology. Front. Aging Neurosci. 2014, 6, 218. [Google Scholar] [CrossRef] [PubMed]
- Braidy, N.; Jugder, B.E.; Poljak, A.; Jayasena, T.; Mansour, H.; Nabavi, S.M.; Sachdev, P.; Grant, R. Resveratrol as a potential therapeutic candidate for the treatment and management of Alzheimer’s disease. Curr. Top. Med. Chem. 2016, 16, 1951–1960. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Surh, Y.J. Protective effect of resveratrol on β-amyloid-induced oxidative PC12 cell death. Free Radic. Biol. Med. 2003, 34, 1100–1110. [Google Scholar] [CrossRef]
- Feng, X.; Liang, N.; Zhu, D.; Gao, Q.; Peng, L.; Dong, H.; Yue, Q.; Liu, H.; Bao, L.; Zhang, J.; et al. Resveratrol inhibits β-amyloid-induced neuronal apoptosis through regulation of SIRT1-ROCK1 signaling pathway. PLoS ONE 2013, 8, e59888. [Google Scholar] [CrossRef] [PubMed]
- Capiralla, H.; Vingtdeux, V.; Zhao, H.; Sankowski, R.; Al-Abed, Y.; Davies, P.; Marambaud, P. Resveratrol mitigates lipopolysaccharide- and Aβ-mediated microglial inflammation by inhibiting the TLR4/NF-KAPPAB/STAT signaling cascade. J. Neurochem. 2012, 120, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Garwood, C.J.; Pooler, A.M.; Atherton, J.; Hanger, D.P.; Noble, W. Astrocytes are important mediators of Aβ-induced neurotoxicity and tau phosphorylation in primary culture. Cell Death Dis. 2011, 2, e167. [Google Scholar] [CrossRef] [PubMed]
- Sollvander, S.; Nikitidou, E.; Brolin, R.; Soderberg, L.; Sehlin, D.; Lannfelt, L.; Erlandsson, A. Accumulation of amyloid-β by astrocytes result in enlarged endosomes and microvesicle-induced apoptosis of neurons. Mol. Neurodegener. 2016, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Amin, F.U.; Shah, S.A.; Kim, M.O. Vanillic acid attenuates Aβ1-42-induced oxidative stress and cognitive impairment in mice. Sci. Rep. 2017, 7, 40753. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; He, M.; Cao, J.; Wang, H.; Ding, J.; Jiao, Y.; Li, R.; He, J.; Wang, D.; Wang, Y. The comparative analysis of the potential relationship between resveratrol and stilbene synthase gene family in the development stages of grapes (vitis quinquangularis and vitis vinifera). Plant Physiol. Biochem. 2014, 74, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Harikumar, K.B.; Aggarwal, B.B. Resveratrol: A multitargeted agent for age-associated chronic diseases. Cell Cycle 2008, 7, 1020–1035. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.M.; Ha, Y.M.; Kim, J.A.; Chung, K.W.; Uehara, Y.; Lee, K.J.; Chun, P.; Byun, Y.; Chung, H.Y.; Moon, H.R. Synthesis of novel azo-resveratrol, azo-oxyresveratrol and their derivatives as potent tyrosinase inhibitors. Bioorg. Med. Chem. Lett. 2012, 22, 7451–7455. [Google Scholar] [CrossRef] [PubMed]
- Boocock, D.J.; Faust, G.E.; Patel, K.R.; Schinas, A.M.; Brown, V.A.; Ducharme, M.P.; Booth, T.D.; Crowell, J.A.; Perloff, M.; Gescher, A.J.; et al. Phase I dose escalation pharmacokinetic study in healthy volunteers of resveratrol, a potential cancer chemopreventive agent. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
- Markus, M.A.; Morris, B.J. Resveratrol in prevention and treatment of common clinical conditions of aging. Clin. Interv. Aging 2008, 3, 331–339. [Google Scholar] [PubMed]
- Williams, L.D.; Burdock, G.A.; Edwards, J.A.; Beck, M.; Bausch, J. Safety studies conducted on high-purity trans-resveratrol in experimental animals. Food Chem. Toxicol. 2009, 47, 2170–2182. [Google Scholar] [CrossRef] [PubMed]
- Burkon, A.; Somoza, V. Quantification of free and protein-bound trans-resveratrol metabolites and identification of trans-resveratrol-c/o-conjugated diglucuronides—Two novel resveratrol metabolites in human plasma. Mol. Nutr. Food Res. 2008, 52, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Torres, G.; Dileo, J.N.; Hallas, B.H.; Horowitz, J.M.; Leheste, J.R. Silent information regulator 1 mediates hippocampal plasticity through presenilin1. Neuroscience 2011, 179, 32–40. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, N.S.; Bayan, Y. Possible role of resveratrol targeting estradiol and neprilysin pathways in lipopolysaccharide model of Alzheimer disease. Adv. Exp. Med. Biol. 2015, 822, 107–118. [Google Scholar] [PubMed]
- Santos, L.M.; Rodrigues, D.; Alemi, M.; Silva, S.C.; Ribeiro, C.A.; Cardoso, I. Resveratrol administration increases transthyretin protein levels ameliorating AD features—Importance of transthyretin tetrameric stability. Mol. Med. 2016, 22, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.F.; Li, N.; Wang, Q.; Cheng, X.J.; Li, X.M.; Liu, T.T. Resveratrol decreases the insoluble Aβ1–42 level in hippocampus and protects the integrity of the blood-brain barrier in ad rats. Neuroscience 2015, 310, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.W.; Choi, Y.H.; Cha, M.R.; Kim, Y.S.; Yon, G.H.; Hong, K.S.; Park, W.K.; Kim, Y.H.; Ryu, S.Y. In vitro BACE-1 inhibitory activity of resveratrol oligomers from the seed extract of paeonia lactiflora. Planta Med. 2011, 77, 374–376. [Google Scholar] [CrossRef] [PubMed]
- Koukoulitsa, C.; Villalonga-Barber, C.; Csonka, R.; Alexi, X.; Leonis, G.; Dellis, D.; Hamelink, E.; Belda, O.; Steele, B.R.; Micha-Screttas, M.; et al. Biological and computational evaluation of resveratrol inhibitors against Alzheimer’s disease. J. Enzyme Inhib. Med. Chem. 2016, 31, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.; Kim, S.; Jang, B.-G.; Kim, M.-J. Piceatannol, a natural analogue of resveratrol, effectively reduces β-amyloid levels via activation of alpha-secretase and matrix metalloproteinase-9. J. Funct. Foods 2009, 23, 124–134. [Google Scholar]
- Ohta, K.; Mizuno, A.; Ueda, M.; Li, S.; Suzuki, Y.; Hida, Y.; Hayakawa-Yano, Y.; Itoh, M.; Ohta, E.; Kobori, M.; et al. Autophagy impairment stimulates PS1 expression and gamma-secretase activity. Autophagy 2010, 6, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Melzig, M.F.; Escher, F. Induction of neutral endopeptidase and angiotensin-converting enzyme activity of SK-N-SH cells in vitro by quercetin and resveratrol. Pharmazie 2002, 57, 556–558. [Google Scholar] [PubMed]
- Ge, J.F.; Qiao, J.P.; Qi, C.C.; Wang, C.W.; Zhou, J.N. The binding of resveratrol to monomer and fibril amyloid β. Neurochem. Int. 2012, 61, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Willem, M.; Garratt, A.N.; Novak, B.; Citron, M.; Kaufmann, S.; Rittger, A.; DeStrooper, B.; Saftig, P.; Birchmeier, C.; Haass, C. Control of peripheral nerve myelination by the β-secretase BACE1. Science 2006, 314, 664–666. [Google Scholar] [CrossRef] [PubMed]
- Tamagno, E.; Bardini, P.; Guglielmotto, M.; Danni, O.; Tabaton, M. The various aggregation states of β-amyloid 1–42 mediate different effects on oxidative stress, neurodegeneration, and BACE-1 expression. Free Radic. Biol. Med. 2006, 41, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Skretas, G.; Meligova, A.K.; Villalonga-Barber, C.; Mitsiou, D.J.; Alexis, M.N.; Micha-Screttas, M.; Steele, B.R.; Screttas, C.G.; Wood, D.W. Engineered chimeric enzymes as tools for drug discovery: Generating reliable bacterial screens for the detection, discovery, and assessment of estrogen receptor modulators. J. Am. Chem. Soc. 2007, 129, 8443–8457. [Google Scholar] [CrossRef] [PubMed]
- Marambaud, P.; Zhao, H.; Davies, P. Resveratrol promotes clearance of Alzheimer’s disease amyloid-β peptides. J. Biol. Chem. 2005, 280, 37377–37382. [Google Scholar] [CrossRef] [PubMed]
- Chiang, K.; Koo, E.H. Emerging therapeutics for Alzheimer’s disease. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 381–405. [Google Scholar] [CrossRef] [PubMed]
- De Strooper, B. APH-1, PEN-2, and nicastrin with presenilin generate an active gamma-secretase complex. Neuron 2003, 38, 9–12. [Google Scholar] [CrossRef]
- Acx, H.; Serneels, L.; Radaelli, E.; Muyldermans, S.; Vincke, C.; Pepermans, E.; Muller, U.; Chavez-Gutierrez, L.; De Strooper, B. Inactivation of gamma-secretases leads to accumulation of substrates and non-Alzheimer neurodegeneration. EMBO Mol. Med. 2017, 9, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Rhein, V.; Song, X.; Wiesner, A.; Ittner, L.M.; Baysang, G.; Meier, F.; Ozmen, L.; Bluethmann, H.; Drose, S.; Brandt, U.; et al. Amyloid-β and tau synergistically impair the oxidative phosphorylation system in triple transgenic Alzheimer’s disease mice. Proc. Natl. Acad. Sci. USA 2009, 106, 20057–20062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesidou, E.; Lagoudaki, R.; Touloumi, O.; Poulatsidou, K.N.; Simeonidou, C. Autophagy and neurodegenerative disorders. Neural Regen. Res. 2013, 8, 2275–2283. [Google Scholar] [PubMed]
- Rubinsztein, D.C.; Marino, G.; Kroemer, G. Autophagy and aging. Cell 2011, 146, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Rockenstein, E.; Crews, L.; Masliah, E. Role of protein aggregation in mitochondrial dysfunction and neurodegeneration in Alzheimer’s and Parkinson’s diseases. Neuromol. Med. 2003, 4, 21–36. [Google Scholar] [CrossRef]
- Duff, K.; Eckman, C.; Zehr, C.; Yu, X.; Prada, C.M.; Perez-tur, J.; Hutton, M.; Buee, L.; Harigaya, Y.; Yager, D.; et al. Increased amyloid-β 42 (43) in brains of mice expressing mutant presenilin 1. Nature 1996, 383, 710–713. [Google Scholar] [CrossRef] [PubMed]
- Nixon, R.A.; Yang, D.S. Autophagy failure in Alzheimer’s disease—Locating the primary defect. Neurobiol. Dis. 2011, 43, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.; Javed, S.; Javed, S.; Tariq, A.; Samec, D.; Tejada, S.; Nabavi, S.F.; Braidy, N.; Nabavi, S.M. Resveratrol and Alzheimer’s disease: Mechanistic insights. Mol. Neurobiol. 2017, 54, 2622–2635. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Mi, M.T. Resveratrol attenuates Aβ25–35 caused neurotoxicity by inducing autophagy through the TYrRS-PARP1-SIRT1 signaling pathway. Neurochem. Res. 2016, 41, 2367–2379. [Google Scholar] [CrossRef] [PubMed]
- Nalivaeva, N.N.; Beckett, C.; Belyaev, N.D.; Turner, A.J. Are amyloid-degrading enzymes viable therapeutic targets in Alzheimer’s disease? J. Neurochem. 2012, 120 (Suppl. 1), 167–185. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.S.; Jo, S.A. Mechanisms of amyloid-β peptide clearance: Potential therapeutic targets for Alzheimer’s disease. Biomol. Ther. 2012, 20, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xiong, H.; Callaghan, D.; Liu, H.; Jones, A.; Pei, K.; Fatehi, D.; Brunette, E.; Stanimirovic, D. Blood-brain barrier transport of amyloid β peptides in efflux pump knock-out animals evaluated by in vivo optical imaging. Fluids Barriers CNS 2013, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Ghobeh, M.; Ahmadian, S.; Meratan, A.A.; Ebrahim-Habibi, A.; Ghasemi, A.; Shafizadeh, M.; Nemat-Gorgani, M. Interaction of Aβ (25–35) fibrillation products with mitochondria: Effect of small-molecule natural products. Biopolymers 2014, 102, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Miners, J.S.; Barua, N.; Kehoe, P.G.; Gill, S.; Love, S. Aβ-degrading enzymes: Potential for treatment of Alzheimer disease. J. Neuropathol. Exp. Neurol. 2011, 70, 944–959. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Quinto, J.; Herdt, A.; Eckman, C.B.; Eckman, E.A. Endothelin-converting enzymes and related metalloproteases in Alzheimer’s disease. J. Alzheimers Dis. 2013, 33 (Suppl. 1), 101–110. [Google Scholar]
- Coppa, T.; Lazze, M.C.; Cazzalini, O.; Perucca, P.; Pizzala, R.; Bianchi, L.; Stivala, L.A.; Forti, L.; Maccario, C.; Vannini, V.; et al. Structure-activity relationship of resveratrol and its analogue, 4,4’-dihydroxy-trans-stilbene, toward the endothelin axis in human endothelial cells. J. Med. Food 2011, 14, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Wyganowska-Swiatkowska, M.; Surdacka, A.; Skrzypczak-Jankun, E.; Jankun, J. The plasminogen activation system in periodontal tissue. Int. J. Mol. Med. 2014, 33, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.L.; Zhan, D.M.; Xi, S.H.; He, X.L. Roles of tissue plasminogen activator and its inhibitor in proliferative diabetic retinopathy. Int. J. Ophthalmol. 2014, 7, 764–767. [Google Scholar] [PubMed]
- Rakic, J.M.; Maillard, C.; Jost, M.; Bajou, K.; Masson, V.; Devy, L.; Lambert, V.; Foidart, J.M.; Noel, A. Role of plasminogen activator-plasmin system in tumor angiogenesis. Cell Mol. Life Sci. 2003, 60, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Salles, F.J.; Strickland, S. Localization and regulation of the tissue plasminogen activator-plasmin system in the hippocampus. J. Neurosci. 2002, 22, 2125–2134. [Google Scholar] [PubMed]
- Melchor, J.P.; Pawlak, R.; Strickland, S. The tissue plasminogen activator-plasminogen proteolytic cascade accelerates amyloid-β (Aβ) degradation and inhibits Aβ-induced neurodegeneration. J. Neurosci. 2003, 23, 8867–8871. [Google Scholar] [PubMed]
- Ertekin-Taner, N.; Ronald, J.; Feuk, L.; Prince, J.; Tucker, M.; Younkin, L.; Hella, M.; Jain, S.; Hackett, A.; Scanlin, L.; et al. Elevated amyloid β protein (Aβ42) and late onset Alzheimer’s disease are associated with single nucleotide polymorphisms in the urokinase-type plasminogen activator gene. Hum. Mol. Genet. 2005, 14, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Zacchigna, S.; Lambrechts, D.; Carmeliet, P. Neurovascular signalling defects in neurodegeneration. Nat. Rev. Neurosci. 2008, 9, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Lam, F.C.; Liu, R.; Lu, P.; Shapiro, A.B.; Renoir, J.M.; Sharom, F.J.; Reiner, P.B. β-Amyloid efflux mediated by p-glycoprotein. J. Neurochem. 2001, 76, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Candela, P.; Gosselet, F.; Saint-Pol, J.; Sevin, E.; Boucau, M.C.; Boulanger, E.; Cecchelli, R.; Fenart, L. Apical-to-basolateral transport of Amyloid-β peptides through blood-brain barrier cells is mediated by the receptor for advanced glycation end-products and is restricted by p-glycoprotein. J. Alzheimers Dis. 2010, 22, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.D.; Bierhaus, A.; Nawroth, P.P.; Stern, D.M. RAGE and Alzheimer’s disease: A progression factor for amyloid-β-induced cellular perturbation? J. Alzheimers Dis. 2009, 16, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Sagare, A.; Deane, R.; Bell, R.D.; Johnson, B.; Hamm, K.; Pendu, R.; Marky, A.; Lenting, P.J.; Wu, Z.; Zarcone, T.; et al. Clearance of amyloid-β by circulating lipoprotein receptors. Nat. Med. 2007, 13, 1029–1031. [Google Scholar] [CrossRef] [PubMed]
- Deane, R.; Wu, Z.; Sagare, A.; Davis, J.; Du Yan, S.; Hamm, K.; Xu, F.; Parisi, M.; LaRue, B.; Hu, H.W.; et al. LRP/amyloid β-peptide interaction mediates differential brain efflux of Aβ isoforms. Neuron 2004, 43, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.; Sarkar, C.; Singh, S.P.; Zhang, Z.; Munasinghe, J.; Peng, S.; Chandra, G.; Kong, E.; Mukherjee, A.B. The blood-brain barrier is disrupted in a mouse model of infantile neuronal ceroid lipofuscinosis: Amelioration by resveratrol. Hum. Mol. Genet. 2012, 21, 2233–2244. [Google Scholar] [CrossRef] [PubMed]
- Georgetown University Medical Center. Resveratrol Appears to Restore Blood-Brain Barrier Integrity in Alzheimer’s Disease. Available online: http://www.sciencedaily.com/releases/2016/07/160727140041.htm (accessed on 27 July 2016).
- Moussa, C.; Hebron, M.; Huang, X.; Ahn, J.; Rissman, R.A.; Aisen, P.S.; Turner, R.S. Resveratrol regulates neuro-inflammation and induces adaptive immunity in Alzheimer’s disease. J. Neuroinflamm. 2017, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Cordon-Cardo, C.; O′Brien, J.P.; Casals, D.; Rittman-Grauer, L.; Biedler, J.L.; Melamed, M.R.; Bertino, J.R. Multidrug-resistance gene (P-glycoprotein) is expressed by endothelial cells at blood-brain barrier sites. Proc. Natl. Acad. Sci. USA 1989, 86, 695–698. [Google Scholar] [CrossRef] [PubMed]
- Deo, A.K.; Borson, S.; Link, J.M.; Domino, K.; Eary, J.F.; Ke, B.; Richards, T.L.; Mankoff, D.A.; Minoshima, S.; O’Sullivan, F.; et al. Activity of P-glycoprotein, a β-amyloid transporter at the blood-brain barrier, is compromised in patients with mild Alzheimer disease. J. Nucl. Med. 2014, 55, 1106–1111. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.S.; Thomas, R.G.; Craft, S.; van Dyck, C.H.; Mintzer, J.; Reynolds, B.A.; Brewer, J.B.; Rissman, R.A.; Raman, R.; Aisen, P.S.; et al. A randomized, double-blind, placebo-controlled trial of resveratrol for Alzheimer disease. Neurology 2015, 85, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Dieckmann, M.; Dietrich, M.F.; Herz, J. Lipoprotein receptors—An evolutionarily ancient multifunctional receptor family. Biol. Chem. 2010, 391, 1341–1363. [Google Scholar] [CrossRef] [PubMed]
- Sagare, A.P.; Deane, R.; Zlokovic, B.V. Low-density lipoprotein receptor-related protein 1: A physiological Aβ homeostatic mechanism with multiple therapeutic opportunities. Pharmacol. Ther. 2012, 136, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.D.; Deane, R.; Chow, N.; Long, X.; Sagare, A.; Singh, I.; Streb, J.W.; Guo, H.; Rubio, A.; Van Nostrand, W.; et al. SRF and myocardin regulate LRP-mediated amyloid-β clearance in brain vascular cells. Nat. Cell Biol. 2009, 11, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Christoforidis, M.; Schober, R.; Krohn, K. Genetic-morphologic association study: Association between the low density lipoprotein-receptor related protein (LRP) and cerebral amyloid angiopathy. Neuropathol. Appl. Neurobiol. 2005, 31, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Silverberg, G.D.; Messier, A.A.; Miller, M.C.; Machan, J.T.; Majmudar, S.S.; Stopa, E.G.; Donahue, J.E.; Johanson, C.E. Amyloid efflux transporter expression at the blood-brain barrier declines in normal aging. J. Neuropathol. Exp. Neurol. 2010, 69, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Rushworth, J.V.; Griffiths, H.H.; Watt, N.T.; Hooper, N.M. Prion protein-mediated toxicity of amyloid-β oligomers requires lipid rafts and the transmembrane LRP1. J. Biol. Chem. 2013, 288, 8935–8951. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.F.; Ramasamy, R.; Schmidt, A.M. The RAGE axis: A fundamental mechanism signaling danger to the vulnerable vasculature. Circ. Res. 2010, 106, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Takuma, K.; Fang, F.; Zhang, W.; Yan, S.; Fukuzaki, E.; Du, H.; Sosunov, A.; McKhann, G.; Funatsu, Y.; Nakamichi, N.; et al. RAGE-mediated signaling contributes to intraneuronal transport of amyloid-β and neuronal dysfunction. Proc. Natl. Acad. Sci. USA 2009, 106, 20021–20026. [Google Scholar] [CrossRef] [PubMed]
- Slevin, M.; Ahmed, N.; Wang, Q.; McDowell, G.; Badimon, L. Unique vascular protective properties of natural products: Supplements or future main-line drugs with significant anti-atherosclerotic potential? Vasc. Cell 2012, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Khazaei, M.; Karimi, J.; Sheikh, N.; Goodarzi, M.T.; Saidijam, M.; Khodadadi, I.; Moridi, H. Effects of resveratrol on receptor for advanced glycation end products (RAGE) expression and oxidative stress in the liver of rats with type 2 diabetes. Phytother. Res. 2016, 30, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Moridi, H.; Karimi, J.; Sheikh, N.; Goodarzi, M.T.; Saidijam, M.; Yadegarazari, R.; Khazaei, M.; Khodadadi, I.; Tavilani, H.; Piri, H.; et al. Resveratrol-dependent down-regulation of receptor for advanced glycation end-products and oxidative stress in kidney of rats with diabetes. Int. J. Endocrinol. Metab. 2015, 13, e23542. [Google Scholar] [CrossRef] [PubMed]
- Malm, T.M.; Jay, T.R.; Landreth, G.E. The evolving biology of microglia in Alzheimer’s disease. Neurotherapeutics 2015, 12, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Aucoin, D.; Ahmed, M.; Ziliox, M.; Van Nostrand, W.E.; Smith, S.O. Capping of Aβ42 oligomers by small molecule inhibitors. Biochemistry 2014, 53, 7893–7903. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, C.A.; Saraiva, M.J.; Cardoso, I. Stability of the transthyretin molecule as a key factor in the interaction with a-β peptide-relevance in Alzheimer’s disease. PLoS ONE 2012, 7, e45368. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lee, K.W.; Lee, H.J. Protective effects of piceatannol against β-amyloid-induced neuronal cell death. Ann. N. Y. Acad. Sci. 2007, 1095, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Guo, Y.; Yan, J.; Luo, Z.; Luo, H.B.; Yan, M.; Huang, L.; Li, X. Design, synthesis, and evaluation of multitarget-directed resveratrol derivatives for the treatment of Alzheimer’s disease. J. Med. Chem. 2013, 56, 5843–5859. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Rimando, A.; Pallas, M.; Camins, A.; Porquet, D.; Reeves, J.; Shukitt-Hale, B.; Smith, M.A.; Joseph, J.A.; Casadesus, G. Low-dose pterostilbene, but not resveratrol, is a potent neuromodulator in aging and Alzheimer’s disease. Neurobiol. Aging 2012, 33, 2062–2071. [Google Scholar] [CrossRef] [PubMed]
- Aprahamian, I.; Stella, F.; Forlenza, O.V. New treatment strategies for Alzheimer’s disease: Is there a hope? Indian J. Med. Res. 2013, 138, 449–460. [Google Scholar] [PubMed]
- Murakami, K.; Irie, K.; Ohigashi, H.; Hara, H.; Nagao, M.; Shimizu, T.; Shirasawa, T. Formation and stabilization model of the 42-mer Aβ radical: Implications for the long-lasting oxidative stress in Alzheimer’s disease. J. Am. Chem. Soc. 2005, 127, 15168–15174. [Google Scholar] [CrossRef] [PubMed]
- Tabner, B.J.; El-Agnaf, O.M.; Turnbull, S.; German, M.J.; Paleologou, K.E.; Hayashi, Y.; Cooper, L.J.; Fullwood, N.J.; Allsop, D. Hydrogen peroxide is generated during the very early stages of aggregation of the amyloid peptides implicated in Alzheimer disease and familial British dementia. J. Biol. Chem. 2005, 280, 35789–35792. [Google Scholar] [CrossRef] [PubMed]
- Cottart, C.H.; Nivet-Antoine, V.; Laguillier-Morizot, C.; Beaudeux, J.L. Resveratrol bioavailability and toxicity in humans. Mol. Nutr. Food Res. 2010, 54, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Pasinetti, G.M.; Wang, J.; Marambaud, P.; Ferruzzi, M.; Gregor, P.; Knable, L.A.; Ho, L. Neuroprotective and metabolic effects of resveratrol: Therapeutic implications for Huntington’s disease and other neurodegenerative disorders. Exp. Neurol. 2011, 232, 1–6. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Isomers of resveratrol.
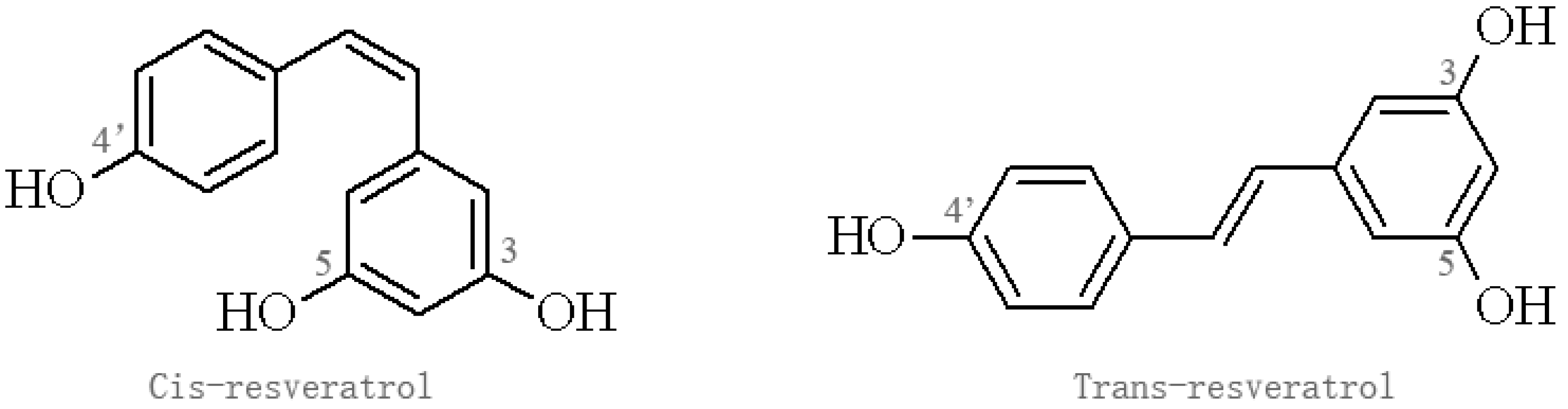
Figure 2.
Effect of resveratrol on Aβ homeostasis. Aβ: amyloid-beta; APP: amyloid precursor protein; ROS: reactive oxygen species; BACE: β-APP cleaving enzyme. ADEs: Aβ-degrading enzymes.
Figure 2.
Effect of resveratrol on Aβ homeostasis. Aβ: amyloid-beta; APP: amyloid precursor protein; ROS: reactive oxygen species; BACE: β-APP cleaving enzyme. ADEs: Aβ-degrading enzymes.

{kind=link}
{kind=link}
Table 1.
Neuroprotective effects of resveratrol in Alzheimer’s disease (AD) models in vivo.
| Experimental Animal | Model/Method | Action of Resveratrol | Dosage | Duration of Treatment | Reference |
|---|---|---|---|---|---|
| APP/PS1 mice | AD | Inhibits Aβ-mediated microglial activation | AIN-93G diet supplemented with 0.35% resveratrol | 15 weeks | [15] |
| Sprague-Dawley rats | - | increases PSEN1 expression in the rat brain | dietary resveratrol | 28 days | [26] |
| Mice | Injected with lipopolysaccharide | increases both the estradiol level and NEP level | injected with resveratrol 4 mg/kg | 7 days | [27] |
| APP/transthyretin (TTR) mice | AD | Increased LRP1 expression upregulated and stabilizes TTR | 174 mg/kg/day | 2 months | [28] |
| Wistar female rats | AD | decreases level of insoluble Aβ in the hippocampus | 80 mg/kg | 12 weeks | [29] |
| reduces the expression of RAGE in the hippocampus | |||||
| inhabits the expression of MMP-9 |
Aβ: amyloid-beta; NEP: neprilysin; APP: amyloid precursor protein; LRP1: lipoprotein receptor-related protein 1; RAGE: the receptor for advanced glycation end products; MMP-9: matrix metalloprotein-9.
Table 2.
Mechanism of resveratrol on amyloid-beta (Aβ) production and clearance in vitro.
| Experimental Model/Method | Exposure | Mechanism of Resveratrol | Dosage of Resveratrol | Duration of Resveratrol Treatment | Reference |
|---|---|---|---|---|---|
| PC12 cells | Aβ25–35 induced | attenuated Aβ-induced cytotoxicity, apoptotic features, and intracellular ROI accumulation. | 25 μM | 24 h | [13] |
| PC12 cells | Aβ25–35 induced | inhibited the cell apoptosis | 12.5–100 μM | 24–48 h | [14] |
| prevented the LDH leakage | |||||
| maintained the intracellular Ca2+ homeostasis | |||||
| Purified baculovirus-expressed BACE-1 | - | inhibition of BACE-1 | 11.9 μM (IC50) | - | [30] |
| TRF assay | - | inhibition of BACE-1 | 28 μM (IC50) | 30 min | [31] |
| Neuro2a cells HEK293 | transfected with a plasmid containing APPsw | reduced γ-secretase activity | 2.5–20 μM | 24 h | [32] |
| induced MMP-9 activation | |||||
| autophagy-related 5 knockdown HEK293 | - | induced conversion of LC3-I to LC3-II | 60 μM | 24 h | [33] |
| suppression of Presenilin-1 induction | |||||
| suppressed Aβ production | |||||
| SK-N-SH cells | - | induction of NEP and ACE activity | 10 μM | 4 days | [34] |
| Hippocampal samples from AD patients | AD | binds to both fibril and monomer Aβ | 1.56–100 μM | - | [35] |
ROI: reactive oxygen intermediate; BACE: β-APP cleaving enzyme.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jia, Y.; Wang, N.; Liu, X. Resveratrol and Amyloid-Beta: Mechanistic Insights. Nutrients 2017, 9, 1122. https://doi.org/10.3390/nu9101122
AMA Style
Jia Y, Wang N, Liu X. Resveratrol and Amyloid-Beta: Mechanistic Insights. Nutrients. 2017; 9(10):1122. https://doi.org/10.3390/nu9101122
Chicago/Turabian StyleJia, Yongming, Na Wang, and Xuewei Liu. 2017. "Resveratrol and Amyloid-Beta: Mechanistic Insights" Nutrients 9, no. 10: 1122. https://doi.org/10.3390/nu9101122
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.