CYP1A2 Genotype Variations Do Not Modify the Benefits and Drawbacks of Caffeine during Exercise: A Pilot Study

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Pre-Experimental Procedures
2.3. Experimental Design
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Goldstein, E.R.; Ziegenfuss, T.; Kalman, D.; Kreider, R.; Campbell, B.; Wilborn, C.; Taylor, L.; Willoughby, D.; Stout, J.; Graves, B.S.; et al. International society of sports nutrition position stand: Caffeine and performance. J. Int. Soc. Sports Nutr. 2010, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Lara, B.; Ruiz-Vicente, D.; Areces, F.; Abian-Vicen, J.; Salinero, J.J.; Gonzalez-Millan, C.; Gallo-Salazar, C.; Del Coso, J. Acute consumption of a caffeinated energy drink enhances aspects of performance in sprint swimmers. Br. J. Nutr. 2015, 114, 908–914. [Google Scholar] [CrossRef] [PubMed]
- Del Coso, J.; Ramirez, J.A.; Munoz, G.; Portillo, J.; Gonzalez-Millan, C.; Munoz, V.; Barbero-Alvarez, J.C.; Munoz-Guerra, J. Caffeine-containing energy drink improves physical performance of elite rugby players during a simulated match. Appl. Physiol. Nutr. Metab. 2013, 38, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Lara, B.; Gonzalez-Millan, C.; Salinero, J.J.; Abian-Vicen, J.; Areces, F.; Barbero-Alvarez, J.C.; Munoz, V.; Portillo, L.J.; Gonzalez-Rave, J.M.; Del Coso, J. Caffeine-containing energy drink improves physical performance in female soccer players. Amino Acids 2014, 46, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Perez-Lopez, A.; Salinero, J.J.; Abian-Vicen, J.; Valades, D.; Lara, B.; Hernandez, C.; Areces, F.; Gonzalez, C.; Del Coso, J. Caffeinated energy drinks improve volleyball performance in elite female players. Med. Sci. Sports Exerc. 2015, 47, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Pitchford, N.W.; Fell, J.W.; Leveritt, M.D.; Desbrow, B.; Shing, C.M. Effect of caffeine on cycling time-trial performance in the heat. J. Sci. Med. Sport 2014, 17, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, A.B.; Randell, R.K.; Jeukendrup, A.E. The metabolic and performance effects of caffeine compared to coffee during endurance exercise. PLoS ONE 2013, 8, e59561. [Google Scholar] [CrossRef] [PubMed]
- Del Coso, J.; Salinero, J.J.; Gonzalez-Millan, C.; Abian-Vicen, J.; Perez-Gonzalez, B. Dose response effects of a caffeine-containing energy drink on muscle performance: A repeated measures design. J. Int. Soc. Sports Nutr. 2012, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Duncan, M.J.; Stanley, M.; Parkhouse, N.; Cook, K.; Smith, M. Acute caffeine ingestion enhances strength performance and reduces perceived exertion and muscle pain perception during resistance exercise. Eur. J. Sport Sci. 2013, 13, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Giles, G.E.; Mahoney, C.R.; Brunye, T.T.; Gardony, A.L.; Taylor, H.A.; Kanarek, R.B. Differential cognitive effects of energy drink ingredients: Caffeine, taurine, and glucose. Pharmacol. Biochem. Behav. 2012, 102, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Haskell, C.F.; Kennedy, D.O.; Wesnes, K.A.; Scholey, A.B. Cognitive and mood improvements of caffeine in habitual consumers and habitual non-consumers of caffeine. Psychopharmacology 2005, 179, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Renda, G.; Committeri, G.; Zimarino, M.; Di Nicola, M.; Tatasciore, A.; Ruggieri, B.; Ambrosini, E.; Viola, V.; Antonucci, I.; Stuppia, L.; et al. Genetic determinants of cognitive responses to caffeine drinking identified from a double-blind, randomized, controlled trial. Eur. Neuropsychopharmacol. 2015, 25, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, H.R.; Tharion, W.J.; Shukitt-Hale, B.; Speckman, K.L.; Tulley, R. Effects of caffeine, sleep loss, and stress on cognitive performance and mood during US Navy seal training. Sea-air-land. Psychopharmacology 2002, 164, 250–261. [Google Scholar] [CrossRef] [PubMed]
- McLellan, T.M.; Kamimori, G.H.; Voss, D.M.; Bell, D.G.; Cole, K.G.; Johnson, D. Caffeine maintains vigilance and improves run times during night operations for special forces. Aviat Space Environ. Med. 2005, 76, 647–654. [Google Scholar] [PubMed]
- McLellan, T.M.; Kamimori, G.H.; Voss, D.M.; Tate, C.; Smith, S.J. Caffeine effects on physical and cognitive performance during sustained operations. Aviat Space Environ. Med. 2007, 78, 871–877. [Google Scholar] [PubMed]
- Hogervorst, E.; Bandelow, S.; Schmitt, J.; Jentjens, R.; Oliveira, M.; Allgrove, J.; Carter, T.; Gleeson, M. Caffeine improves physical and cognitive performance during exhaustive exercise. Med. Sci. Sports Exerc. 2008, 40, 1841–1851. [Google Scholar] [CrossRef] [PubMed]
- Salinero, J.J.; Lara, B.; Abian-Vicen, J.; Gonzalez-Millan, C.; Areces, F.; Gallo-Salazar, C.; Ruiz-Vicente, D.; Del Coso, J. The use of energy drinks in sport: Perceived ergogenicity and side effects in male and female athletes. Br. J. Nutr. 2014, 112, 1494–1502. [Google Scholar] [CrossRef] [PubMed]
- Tarnopolsky, M.A. Caffeine and creatine use in sport. Ann. Nutr. Metab. 2010, 57, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.K.; Green, J.M. Caffeine and anaerobic performance: Ergogenic value and mechanisms of action. Sports Med. 2009, 39, 813–832. [Google Scholar] [CrossRef] [PubMed]
- Magkos, F.; Kavouras, S.A. Caffeine use in sports, pharmacokinetics in man, and cellular mechanisms of action. Crit. Rev. Food Sci. Nutr. 2005, 45, 535–562. [Google Scholar] [CrossRef] [PubMed]
- Skinner, T.L.; Jenkins, D.G.; Coombes, J.S.; Taaffe, D.R.; Leveritt, M.D. Dose response of caffeine on 2000-m rowing performance. Med. Sci. Sports Exerc. 2010, 42, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Meyers, B.M.; Cafarelli, E. Caffeine increases time to fatigue by maintaining force and not by altering firing rates during submaximal isometric contractions. J. Appl. Physiol. 2005, 99, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Doherty, M.; Smith, P.M.; Davison, R.C.; Hughes, M.G. Caffeine is ergogenic after supplementation of oral creatine monohydrate. Med. Sci. Sports Exerc. 2002, 34, 1785–1792. [Google Scholar] [CrossRef] [PubMed]
- Perera, V.; Gross, A.S.; McLachlan, A.J. Measurement of CYP1A2 activity: A focus on caffeine as a probe. Curr. Drug Metab. 2012, 13, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Ghotbi, R.; Christensen, M.; Roh, H.K.; Ingelman-Sundberg, M.; Aklillu, E.; Bertilsson, L. Comparisons of CYP1A2 genetic polymorphisms, enzyme activity and the genotype-phenotype relationship in Swedes and Koreans. Eur. J. Clin. Pharmacol. 2007, 63, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Djordjevic, N.; Ghotbi, R.; Jankovic, S.; Aklillu, E. Induction of CYP1A2 by heavy coffee consumption is associated with the CYP1A2 -163C>A polymorphism. Eur. J. Clin. Pharmacol. 2010, 66, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Womack, C.J.; Saunders, M.J.; Bechtel, M.K.; Bolton, D.J.; Martin, M.; Luden, N.D.; Dunham, W.; Hancock, M. The influence of a CYP1A2 polymorphism on the ergogenic effects of caffeine. J. Int. Soc. Sports Nutr. 2012, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Pataky, M.W.; Womack, C.J.; Saunders, M.J.; Goffe, J.L.; D’Lugos, A.C.; El-Sohemy, A.; Luden, N.D. Caffeine and 3-km cycling performance: Effects of mouth rinsing, genotype, and time of day. Scand. J. Med. Sci. Sports 2016, 26, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Algrain, H.; Thomas, R.; Carrillo, A.; Ryan, E.; Kim, C.; Lettan, R.; Ryan, E. The effects of a polymorphism in the cytochrome p450 CYP1A2 gene on performance enhancement with caffeine in recreational cyclists. J. Caffeine Res. 2015, 6, 1–6. [Google Scholar]
- Armstrong, L.E. Caffeine, body fluid-electrolyte balance, and exercise performance. Int. J. Sport Nutr. Exerc. Metab. 2002, 12, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Attia, A.; Hachana, Y.; Chaabene, H.; Gaddour, A.; Neji, Z.; Shephard, R.J.; Chelly, M.S. Reliability and validity of a 20-s alternative to the wingate anaerobic test in team sport male athletes. PLoS ONE 2014, 9, e114444. [Google Scholar] [CrossRef] [PubMed]
- Bar-Or, O. The wingate anaerobic test. An update on methodology, reliability and validity. Sports Med. 1987, 4, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Collomp, K.; Ahmaidi, S.; Audran, M.; Chanal, J.L.; Prefaut, C. Effects of caffeine ingestion on performance and anaerobic metabolism during the wingate test. Int. J. Sports Med. 1991, 12, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Greer, F.; McLean, C.; Graham, T.E. Caffeine, performance, and metabolism during repeated wingate exercise tests. J. Appl. Physiol. 1998, 85, 1502–1508. [Google Scholar] [PubMed]
- Lieberman, H.R.; Wurtman, R.J.; Emde, G.G.; Roberts, C.; Coviella, I.L. The effects of low doses of caffeine on human performance and mood. Psychopharmacology 1987, 92, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Santos, V.G.; Santos, V.R.; Felippe, L.J.; Almeida, J.W., Jr.; Bertuzzi, R.; Kiss, M.A.; Lima-Silva, A.E. Caffeine reduces reaction time and improves performance in simulated-contest of taekwondo. Nutrients 2014, 6, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, B.H.; Edgley, B.M. Effects of caffeine on simple reaction time and movement time. Aviat Space Environ. Med. 1987, 58, 1153–1156. [Google Scholar] [PubMed]
- Crowe, M.J.; Leicht, A.S.; Spinks, W.L. Physiological and cognitive responses to caffeine during repeated, high-intensity exercise. Int. J. Sport Nutr. Exerc. Metab. 2006, 16, 528–544. [Google Scholar] [CrossRef] [PubMed]
- Del Coso, J.; Portillo, J.; Munoz, G.; Abian-Vicen, J.; Gonzalez-Millan, C.; Munoz-Guerra, J. Caffeine-containing energy drink improves sprint performance during an international rugby sevens competition. Amino Acids 2013, 44, 1511–1519. [Google Scholar] [CrossRef] [PubMed]
- Retey, J.V.; Adam, M.; Khatami, R.; Luhmann, U.F.; Jung, H.H.; Berger, W.; Landolt, H.P. A genetic variation in the adenosine A2A receptor gene (adora2a) contributes to individual sensitivity to caffeine effects on sleep. Clin. Pharmacol. Ther. 2007, 81, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Alsene, K.; Deckert, J.; Sand, P.; de Wit, H. Association between A2A receptor gene polymorphisms and caffeine-induced anxiety. Neuropsychopharmacology 2003, 28, 1694–1702. [Google Scholar] [CrossRef] [PubMed]
- Kashuba, A.D.; Bertino, J.S., Jr.; Kearns, G.L.; Leeder, J.S.; James, A.W.; Gotschall, R.; Nafziger, A.N. Quantitation of three-month intraindividual variability and influence of sex and menstrual cycle phase on CYP1A2, N-acetyltransferase-2, and xanthine oxidase activity determined with caffeine phenotyping. Clin. Pharmacol. Ther. 1998, 63, 540–551. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Variable | Placebo | Caffeine | Mean Difference | 95% Confidence Interval of the Difference | p |
|---|---|---|---|---|---|
| Reaction time stage 1 (ms) | 275 ± 50 | 268 ± 39 | –7 | (–32; 18) | 0.580 |
| AA homozygotes | 256 ± 58 | 284 ± 58 | 28 | (–50; 107) | 0.337 |
| C allele carriers | 280 ± 49 | 264 ± 34 | –16 | (–44; 13) | 0.259 |
| Reaction time stage 2 (ms) | 280 ± 42 | 276 ± 35 | –4 | (–21; 13) | 0.582 |
| AA homozygotes | 296 ± 76 | 287 ± 54 | –9 | (–104; 86) | 0.780 |
| C allele carriers | 276 ± 32 | 273 ± 31 | –3 | (–19; 14) | 0.725 |
| Reaction time stage 3 (ms) | 291 ± 59 | 278 ± 38 | –13 | (–36; 9) | 0.226 |
| AA homozygotes | 304 ± 118 | 288 ± 66 | –16 | (–159; 127) | 0.745 |
| C allele carriers | 288 ± 39 | 276 ± 30 | –12 | (–32; 6) | 0.178 |
| Reaction time stage 4 (ms) | 283 ± 43 | 276 ± 34 | –7 | (–20; 7) | 0.203 |
| AA homozygotes | 286 ± 59 | 288 ± 51 | 2 | (–55; 59) | 0.914 |
| C allele carriers | 282 ± 40 | 274 ± 30 | –8 | (–23; 6) | 0.232 |
| Reaction time Mean (ms) | 282 ± 42 | 275 ± 31 | –7.5 | (–23; 8) | 0.309 |
| AA homozygotes | 285 ± 75 | 287 ± 54 | 2 | (–84; 87) | 0.965 |
| C allele carriers | 281 ± 33 | 272 ± 23 | –9 | (–24; 4) | 0.165 |
| Variable | Placebo | Caffeine | Mean Difference | 95% IC for the Difference | p |
|---|---|---|---|---|---|
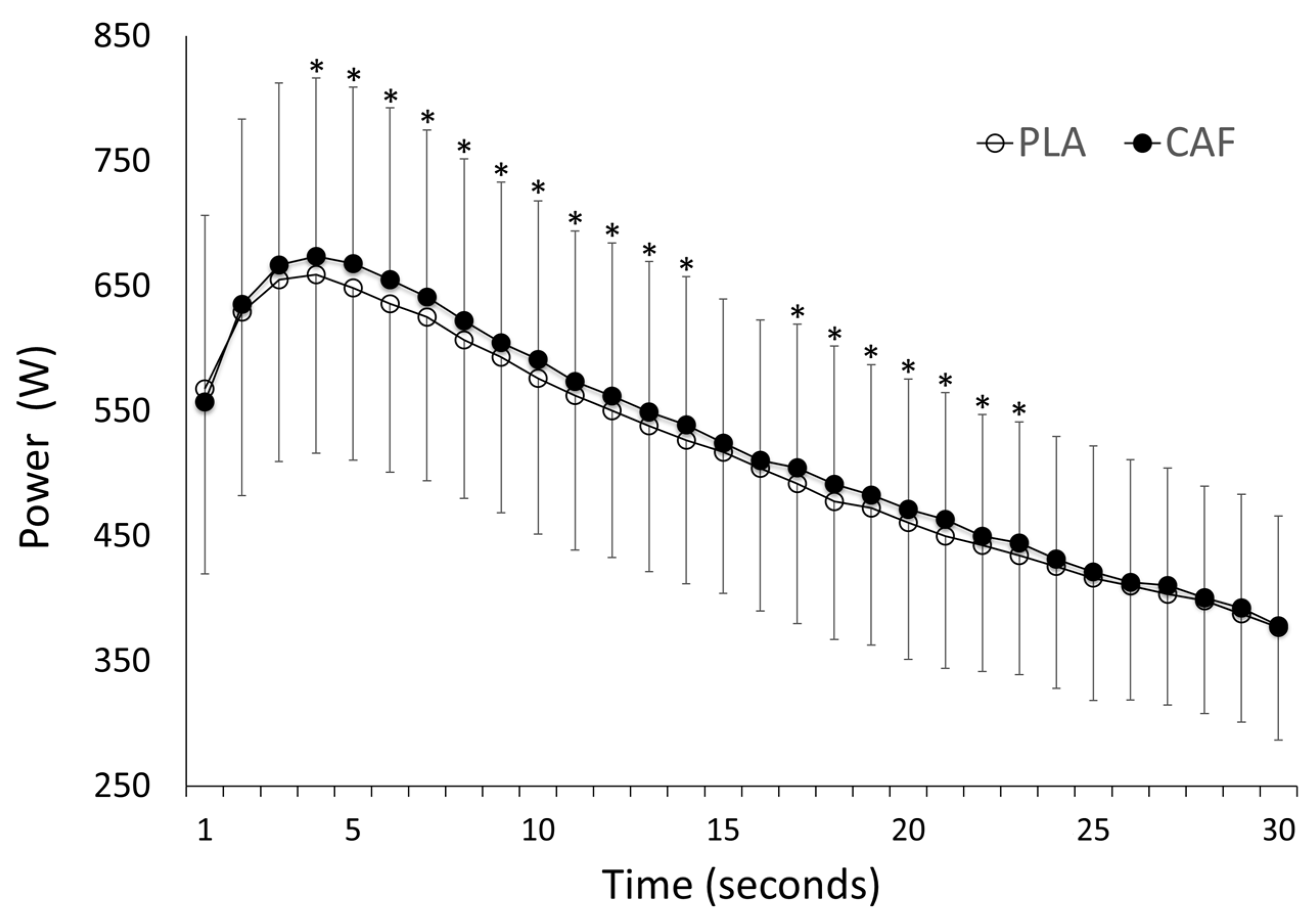
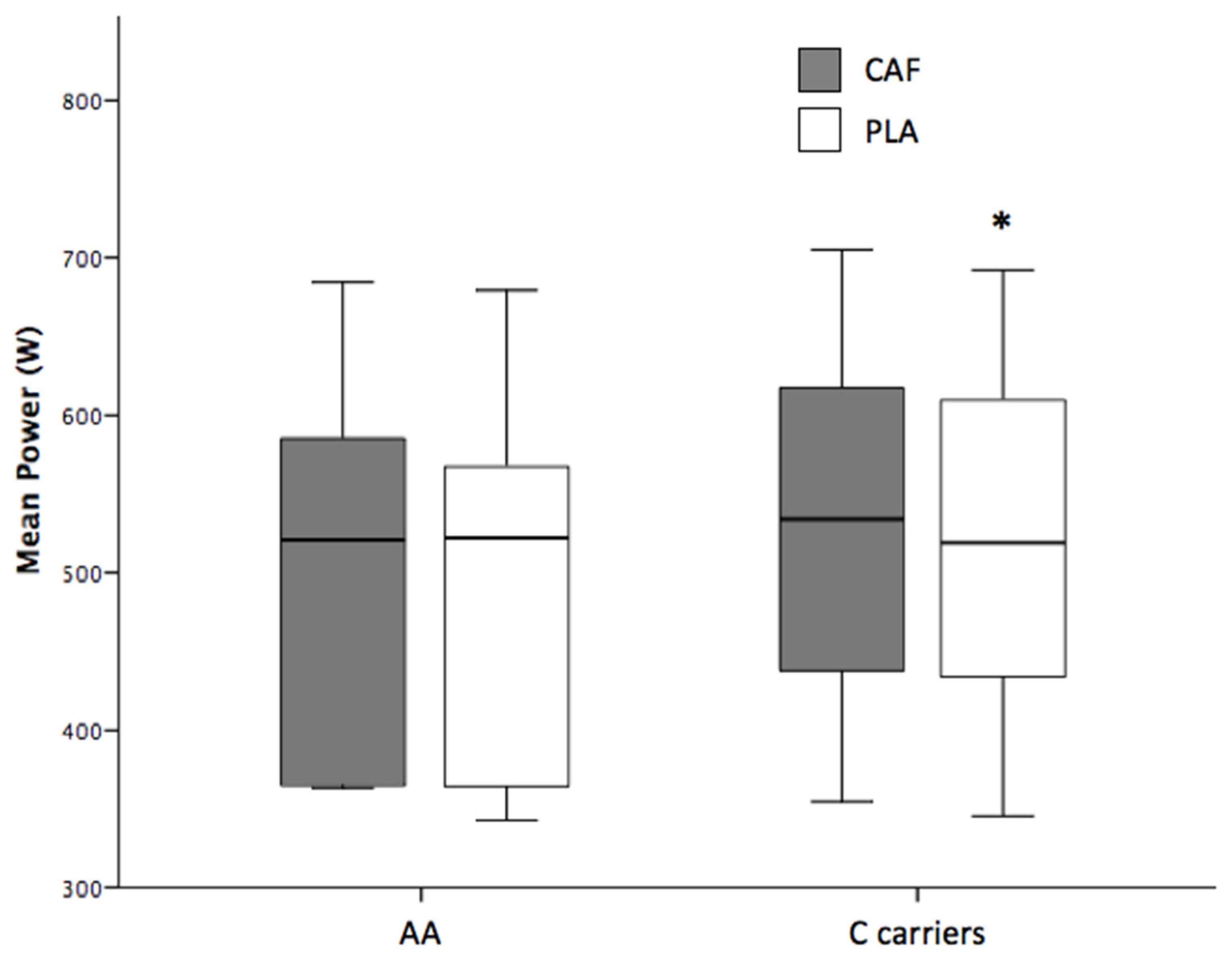
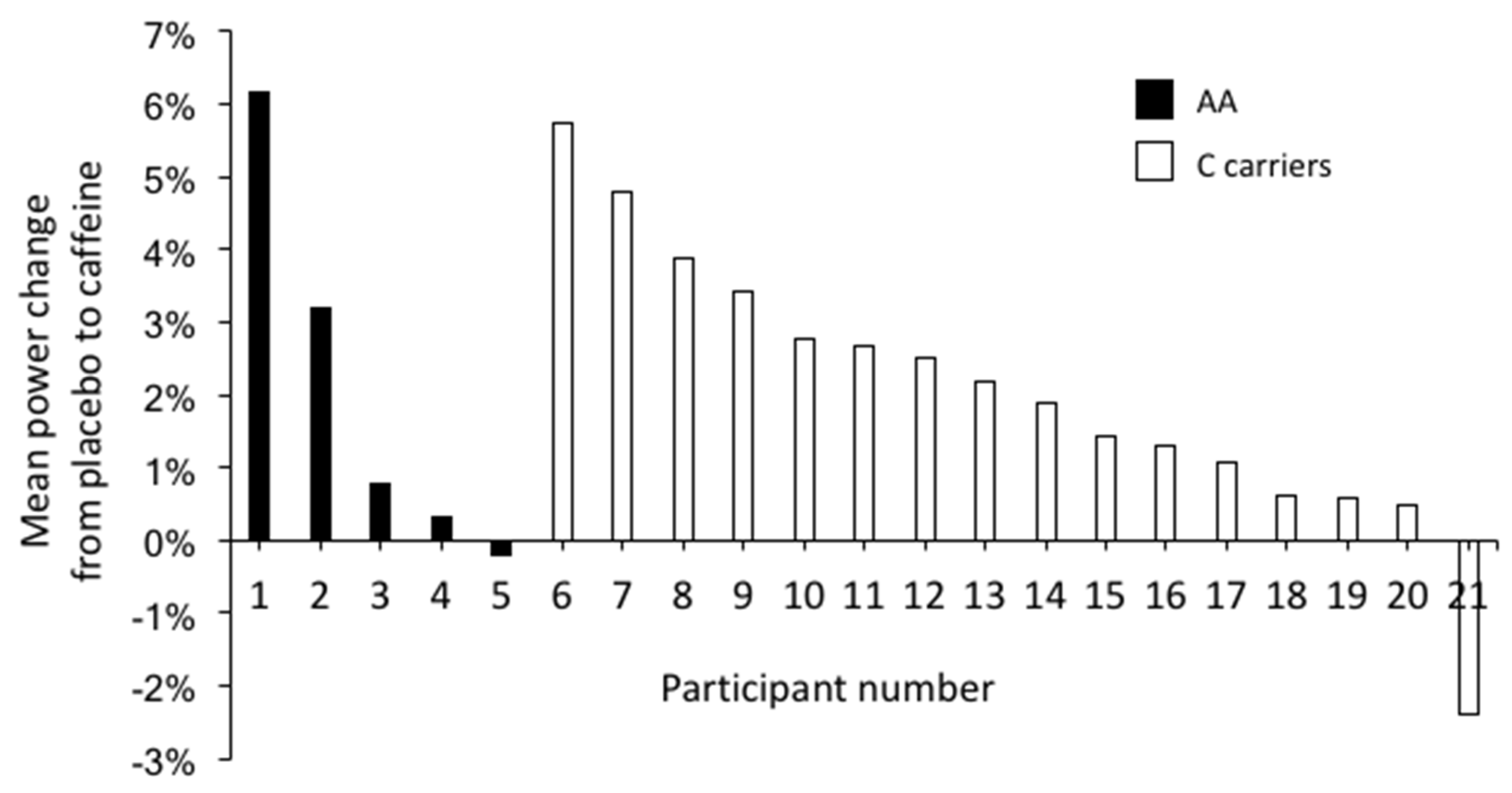
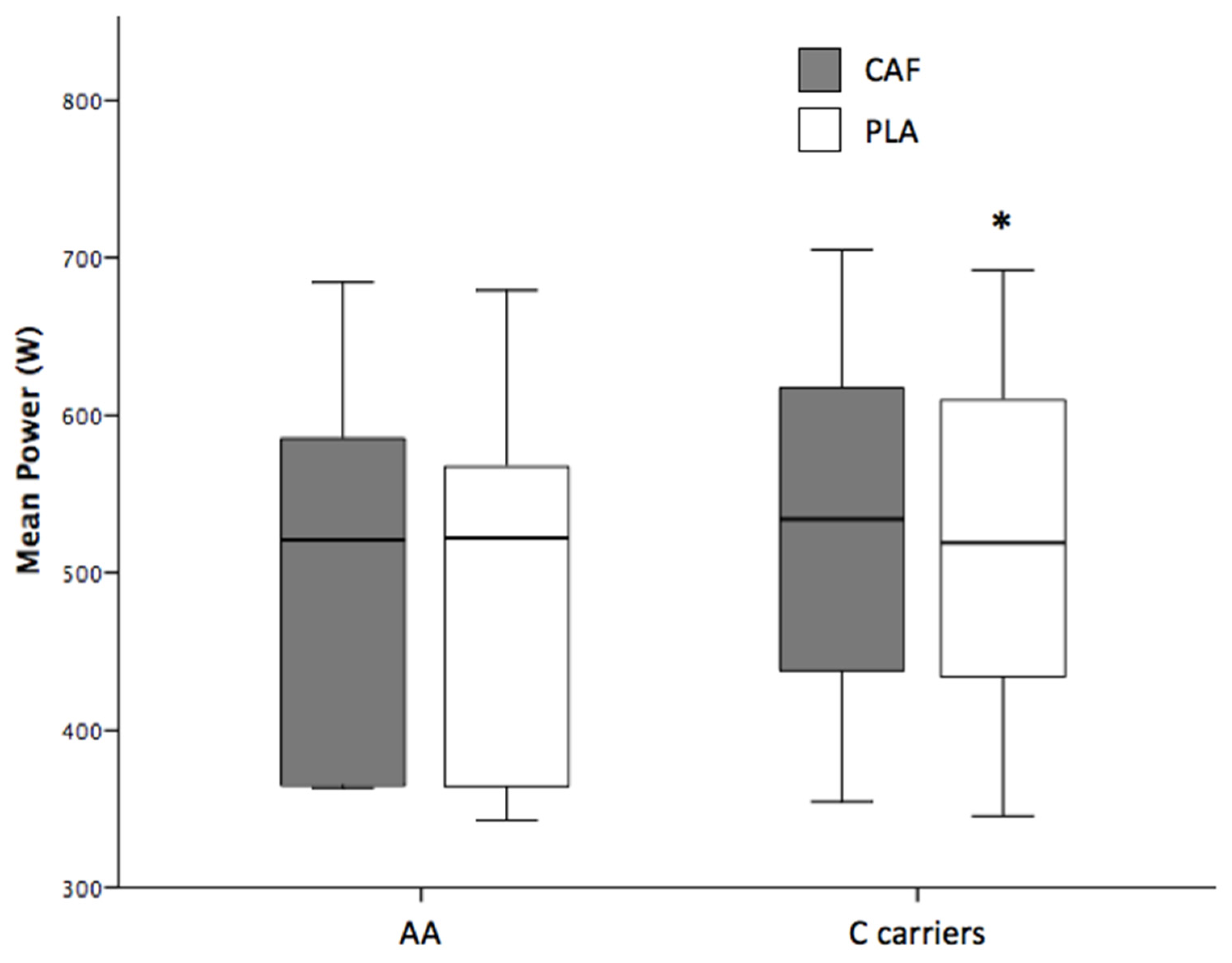
| Mean Power (W) | 511 ± 113 | 521 ± 115 * | 10 | (5.3; 15.0) | 0.000 |
| AA homozygotes | 494 ± 142 | 504 ± 140 | 9 | (–0.9; 18.5) | 0.120 |
| C allele carriers | 516 ± 107 | 527 ± 110 * | 11 | (4.6; 16.4) | 0.002 |
| Peak Power (W) | 663 ± 143 | 680 ± 146 * | 16 | (3.7; 29.0) | 0.014 |
| AA homozygotes | 637 ± 200 | 646 ± 201 | 9 | (–0.9; 18.5) | 0.065 |
| C allele carriers | 671 ± 128 | 690 ± 131 * | 19 | (1.9; 35.5) | 0.031 |
| Fatigue index (%) | 44.2 ± 6.8 | 45.0 ± 7.2 | 0.8 | (–2.0; 3.6) | 0.571 |
| AA homozygotes | 42.3 ± 6.5 | 42.2 ± 6.7 | –0.1 | (–2.1; 1.8) | 0.844 |
| C allele carriers | 44.8 ± 7.0 | 45.9 ± 7.3 | 1.1 | (–2.7; 4.8) | 0.555 |
| Perceived power | 6 ± 2 | 7 ± 2 | 1 | (–0.3; 1.7) | 0.158 |
| AA homozygotes | 6 ± 2 | 7 ± 2 | 1 | (–2.6; 3.8) | 0.634 |
| C allele carriers | 6 ± 2 | 7 ± 2 | 1 | (–0.4; 1.9) | 0.188 |
| Perceived exertion | 6 ± 2 | 6 ± 2 | 0 | (–0.9; 1.1) | 0.836 |
| AA homozygotes | 6 ± 1 | 6 ± 2 | 0 | (–2.2; 3.0) | 0.688 |
| C allele carriers | 6 ± 2 | 6 ± 2 | 0 | (–1.2; 1.2) | 1.000 |
| Variable | Placebo Frequency (%) | Caffeine Frequency (%) |
|---|---|---|
| Nervousness | 1 (4.8) | 5 (23.8) |
| AA homozygotes | 1 (20) | 0 (0) |
| C allele carriers | 0 (0) | 5 (31.3) |
| Insomnia | 3 (14.3) | 7 (33.3) |
| AA homozygotes | 0 (0) | 2 (40.0) |
| C allele carriers | 3 (18.8) | 5 (31.3) |
| Gastrointestinal problems | 0 (0) | 3 (14.3) |
| AA homozygotes | 0 (0) | 1 (20.0) |
| C allele carriers | 0 (0) | 2 (12.5) |
| Activeness | 3 (14.3) | 6 (28.6) |
| AA homozygotes | 2 (40.0) | 1 (20.0) |
| C allele carriers | 1 (6.3) | 5 (31.3) |
| Irritability | 0 (0) | 2 (9.5) |
| AA homozygotes | 0 (0) | 0 (0) |
| C allele carriers | 0 (0) | 2 (12.5) |
| Muscular pain | 4 (19) | 3 (14.3) |
| AA homozygotes | 0 (0) | 0 (0) |
| C allele carriers | 4 (25.0) | 3 (18.8) |
| Headache | 5 (23.8) | 4 (19.0) |
| AA homozygotes | 1 (20) | 1 (20) |
| C allele carriers | 4 (25.0) | 3 (18.8) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salinero, J.J.; Lara, B.; Ruiz-Vicente, D.; Areces, F.; Puente-Torres, C.; Gallo-Salazar, C.; Pascual, T.; Del Coso, J. CYP1A2 Genotype Variations Do Not Modify the Benefits and Drawbacks of Caffeine during Exercise: A Pilot Study. Nutrients 2017, 9, 269. https://doi.org/10.3390/nu9030269
Salinero JJ, Lara B, Ruiz-Vicente D, Areces F, Puente-Torres C, Gallo-Salazar C, Pascual T, Del Coso J. CYP1A2 Genotype Variations Do Not Modify the Benefits and Drawbacks of Caffeine during Exercise: A Pilot Study. Nutrients. 2017; 9(3):269. https://doi.org/10.3390/nu9030269
Chicago/Turabian StyleSalinero, Juan J., Beatriz Lara, Diana Ruiz-Vicente, Francisco Areces, Carlos Puente-Torres, César Gallo-Salazar, Teodoro Pascual, and Juan Del Coso. 2017. "CYP1A2 Genotype Variations Do Not Modify the Benefits and Drawbacks of Caffeine during Exercise: A Pilot Study" Nutrients 9, no. 3: 269. https://doi.org/10.3390/nu9030269