Daily Intake of Grape Powder Prevents the Progression of Kidney Disease in Obese Type 2 Diabetic ZSF1 Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Cells
2.2. Whole Grape Powder (WGP)
2.3. Food Supplements
2.4. Preparation of WGP Extract
2.5. Primary Outcomes from This Study
2.6. Urine Collection and Determination of Kidney Function
2.7. Blood Collection and Chemical Analysis
2.8. Glomerular Filtration Rate (GFR)
2.9. Renal Histopathology
2.10. PCR Array Analysis of Oxidative Stress
2.11. Flow Cytometric Analysis
2.12. Statistical Analysis
3. Results
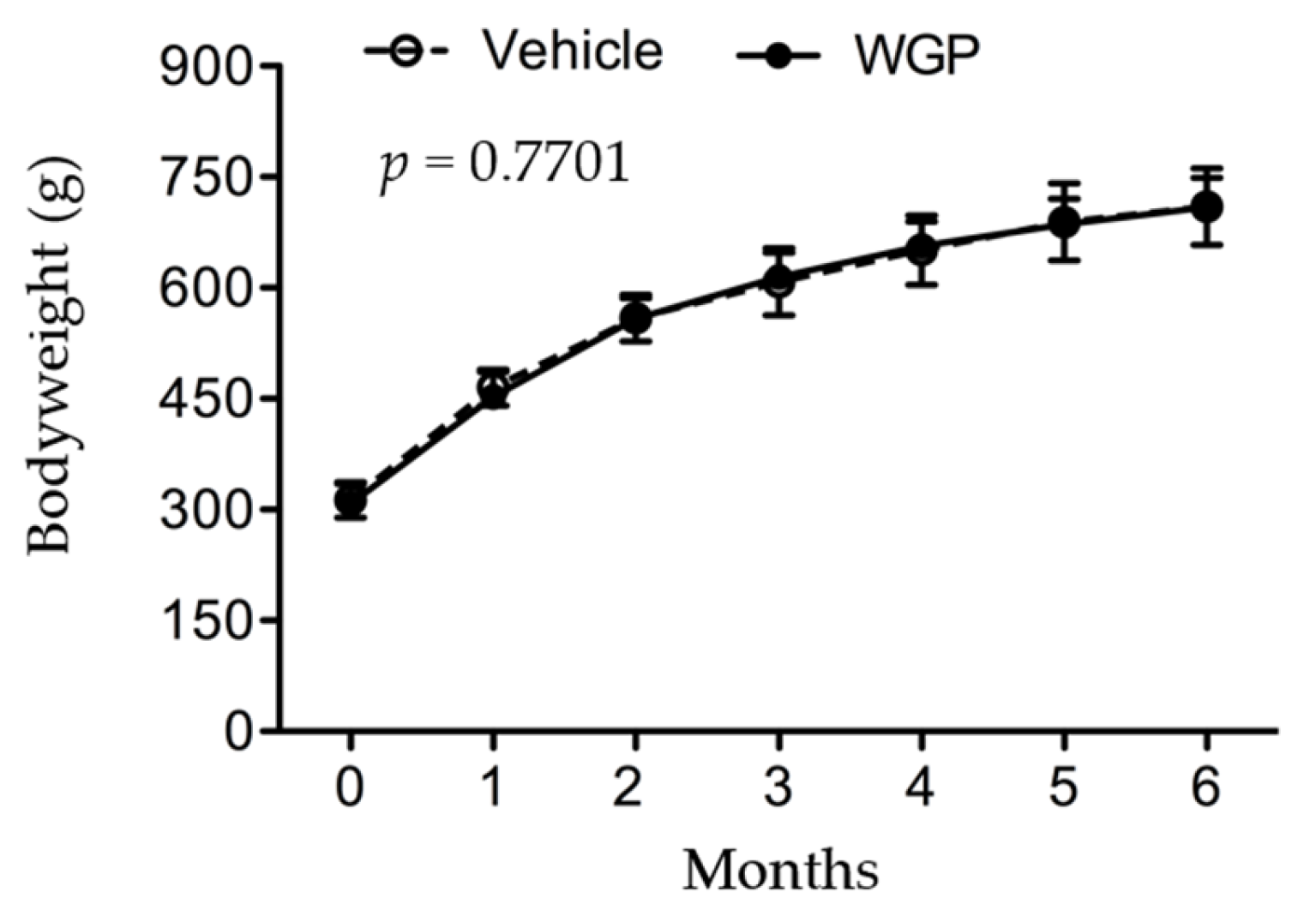
3.1. Daily Intake of WGP Is Associated with Lower Organ Index of the Kidney, Liver and Spleen but Does Not Affect Bodyweight in Obese Diabetic Rats
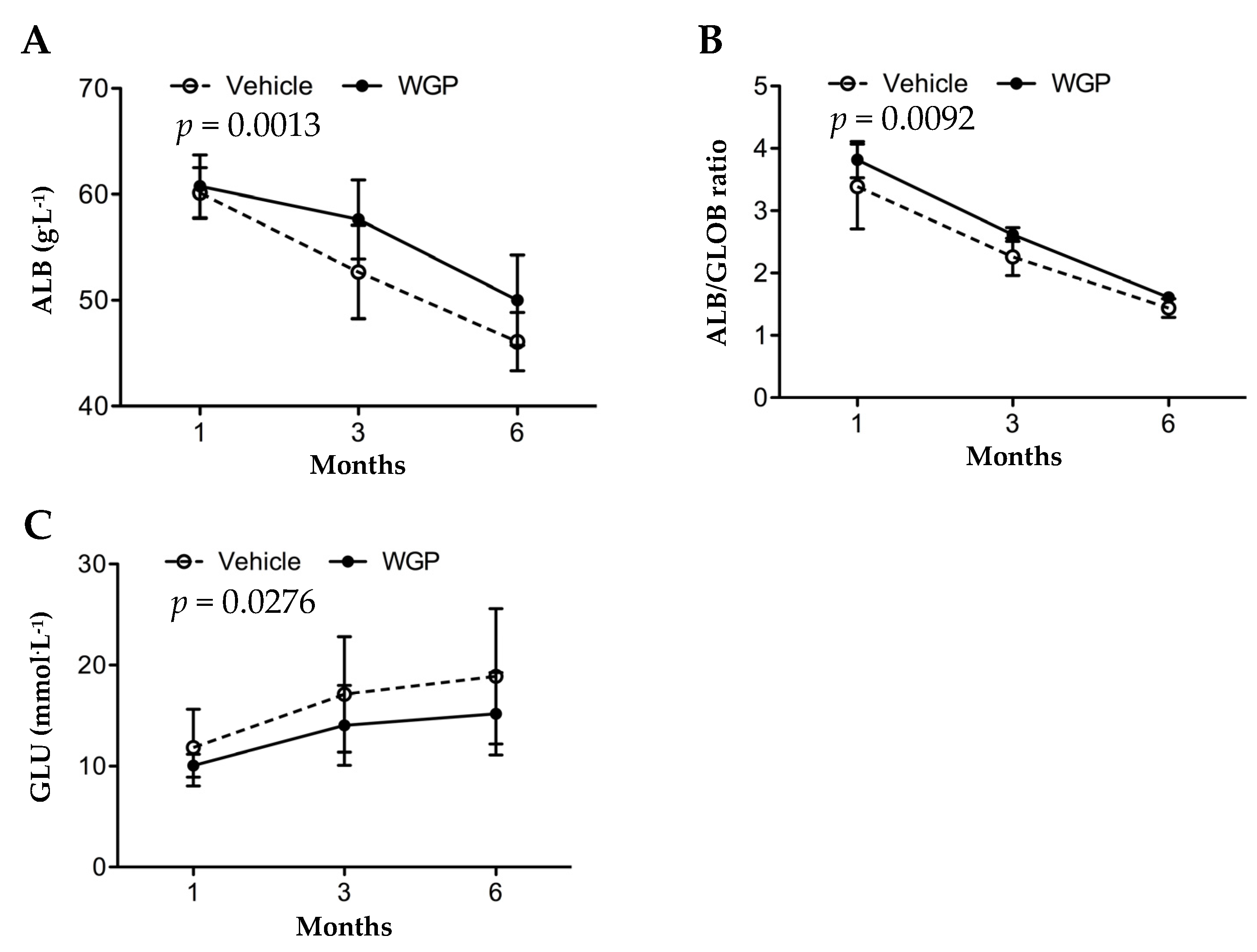
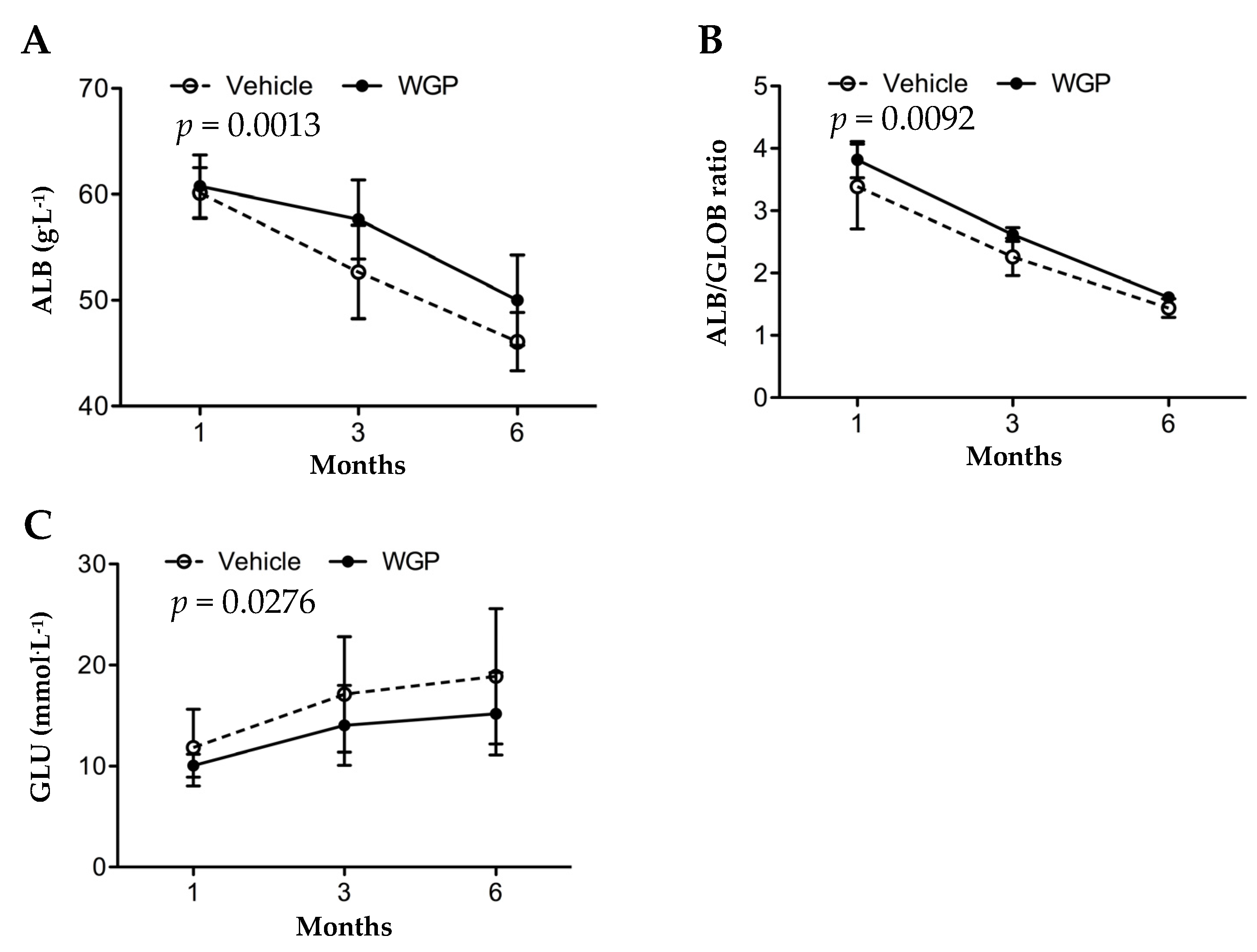
3.2. Daily Intake of WGP Is Associated with Higher Serum Albumin and Lower Blood Glucose in Obese Diabetic Rats
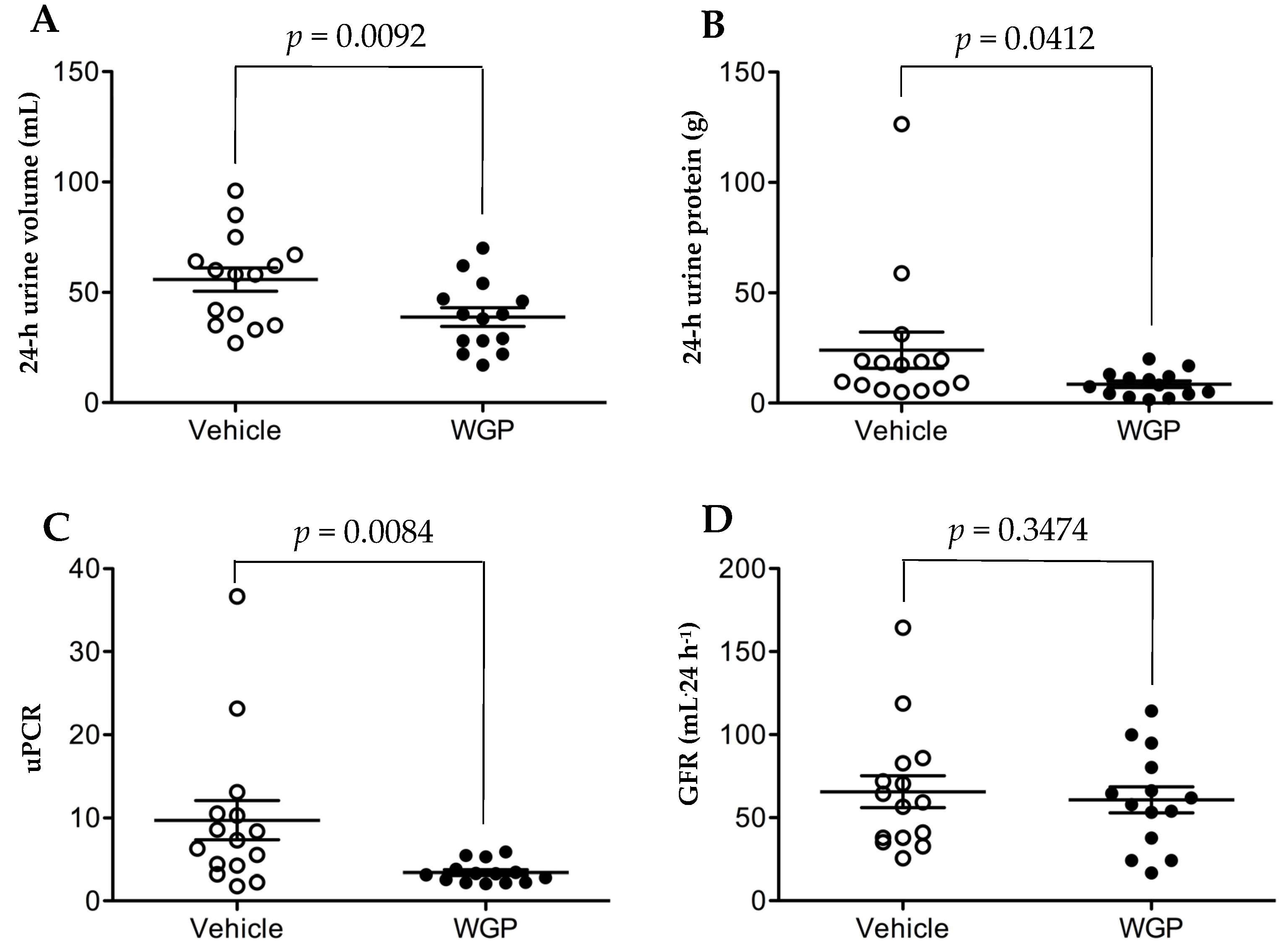
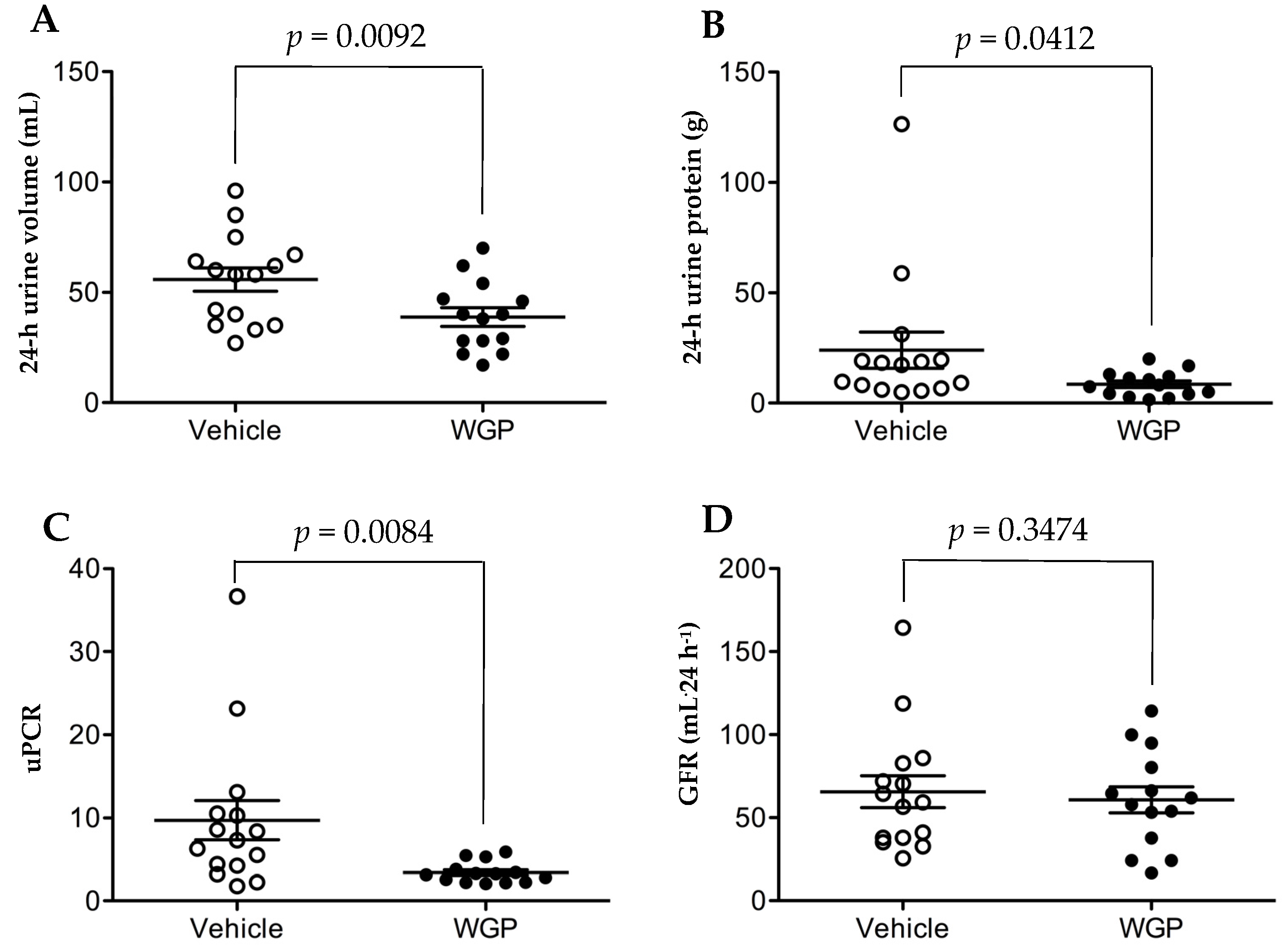
3.3. Daily Intake of WGP Significantly Is Associated with Lower Urine Volume and Urine Protein Excretion but Does Not Impact GFR after Six Months in Obese Diabetic Rats
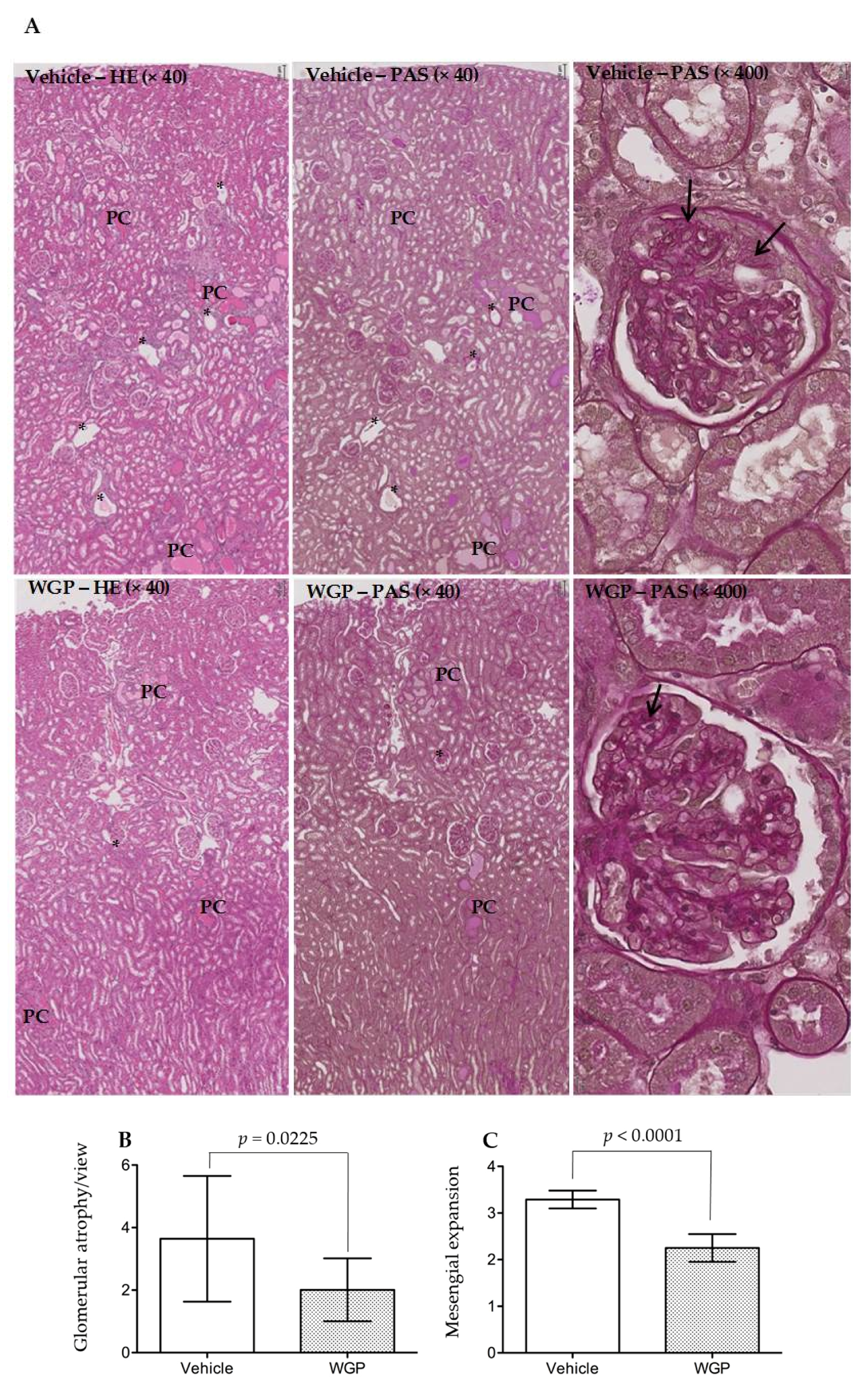
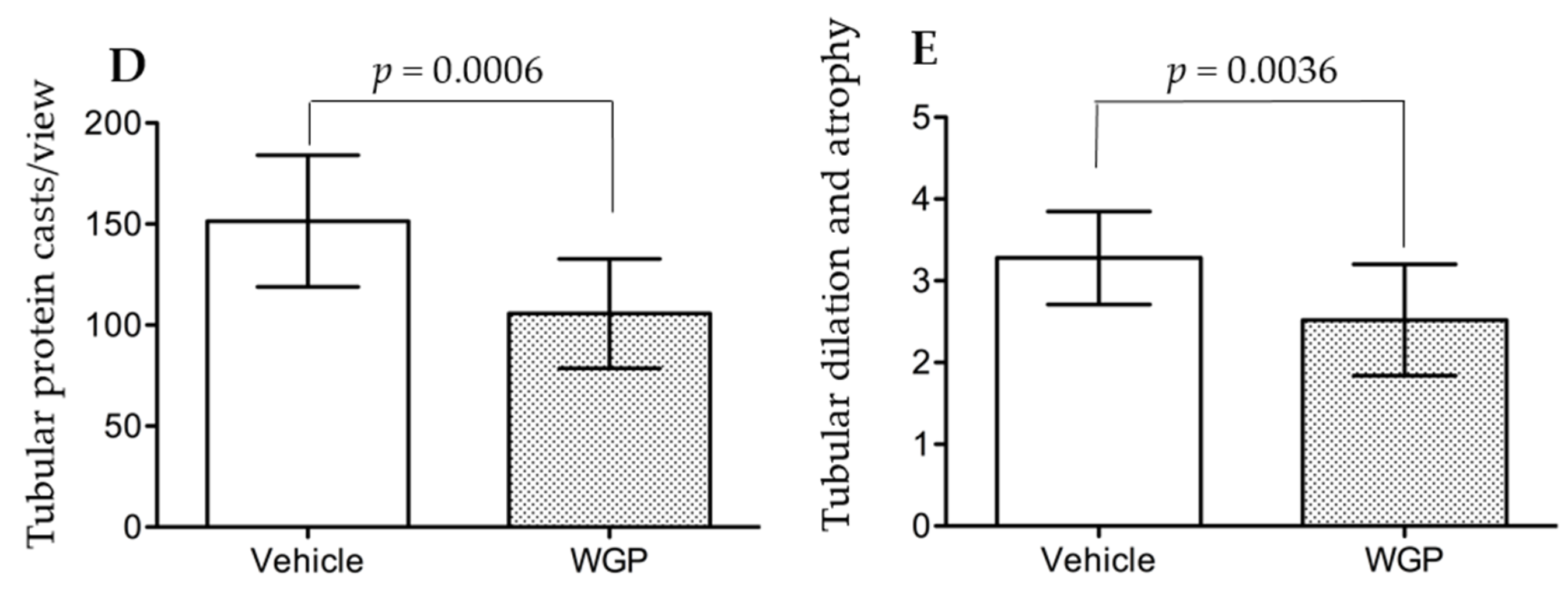
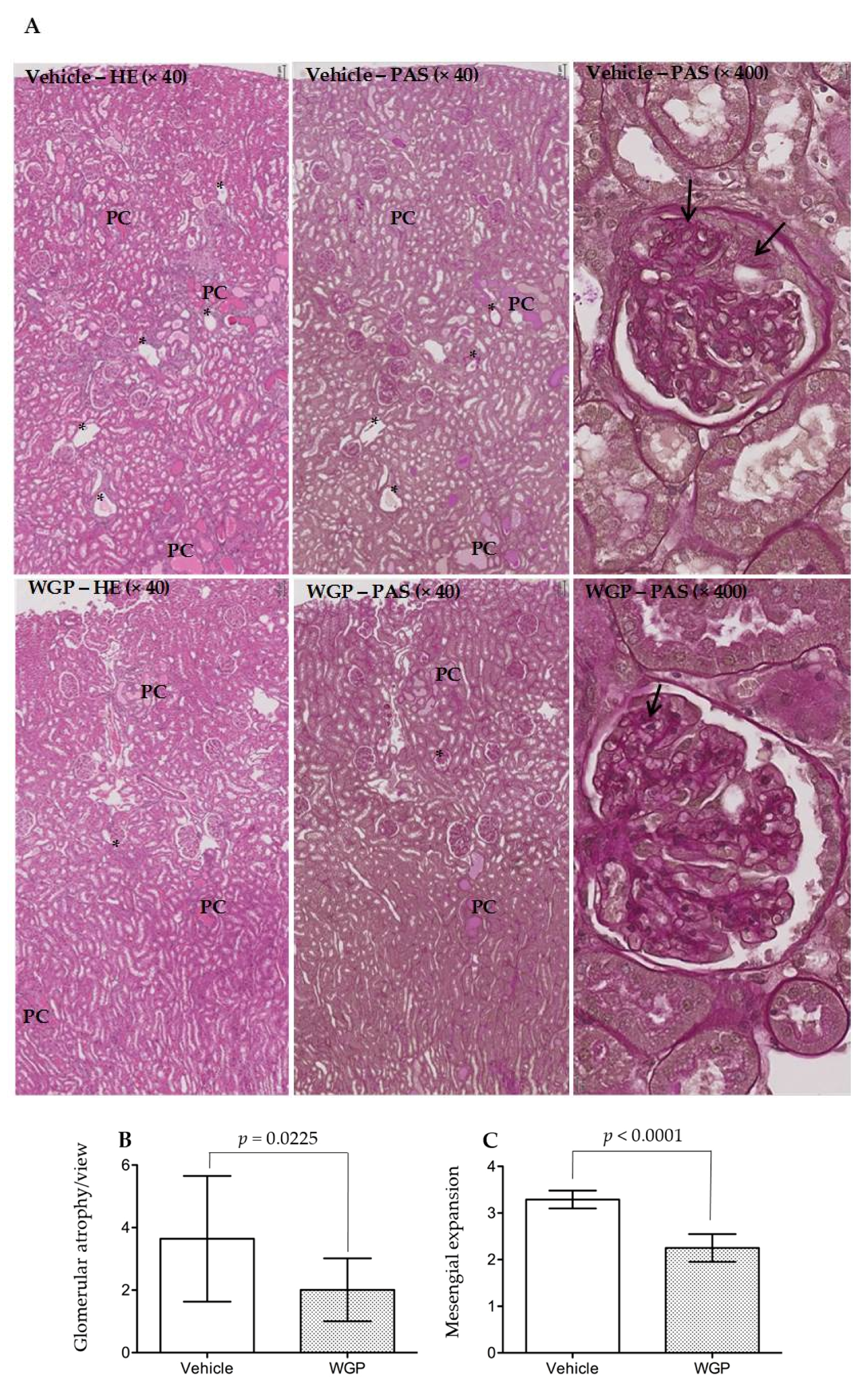
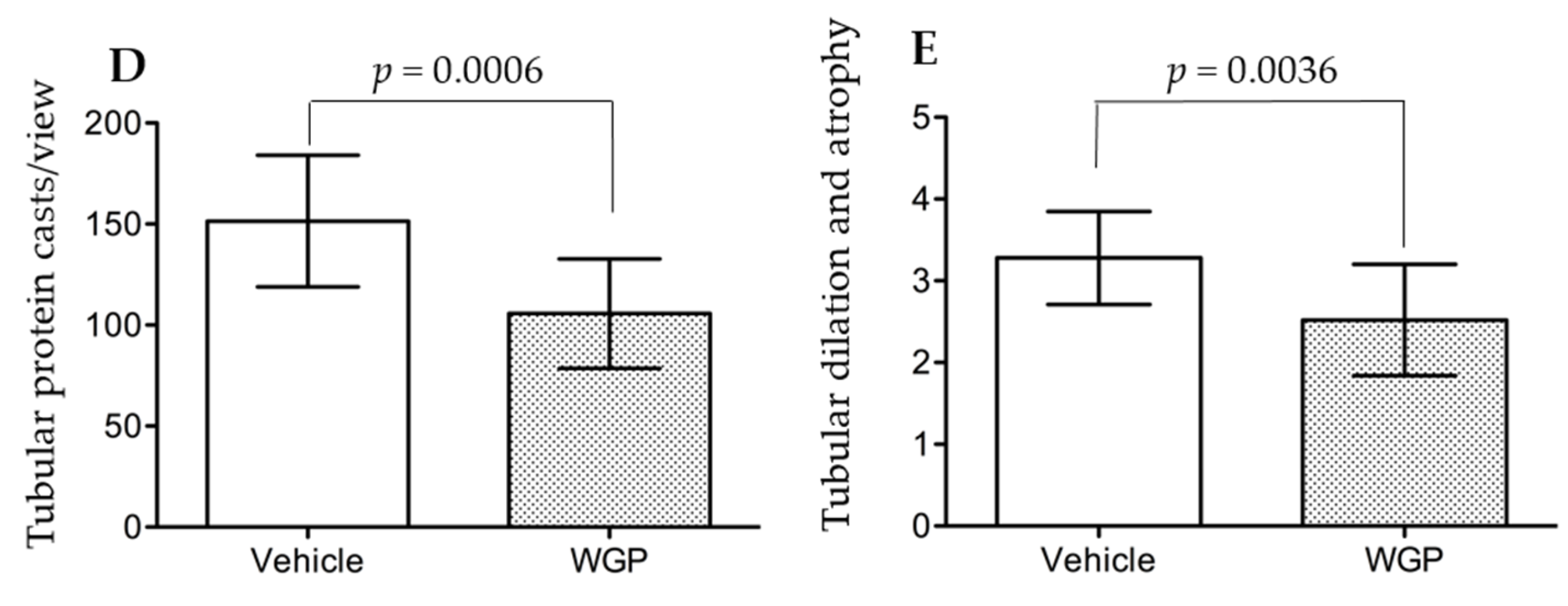
3.4. Daily Intake of WGP Partially Prevents Renal Pathological Changes in Obese Diabetic Rats
3.5. Daily Intake of WGP Is Associated with Increasing Local Antioxidant Defense in the Kidney of Obese Diabetic Rats
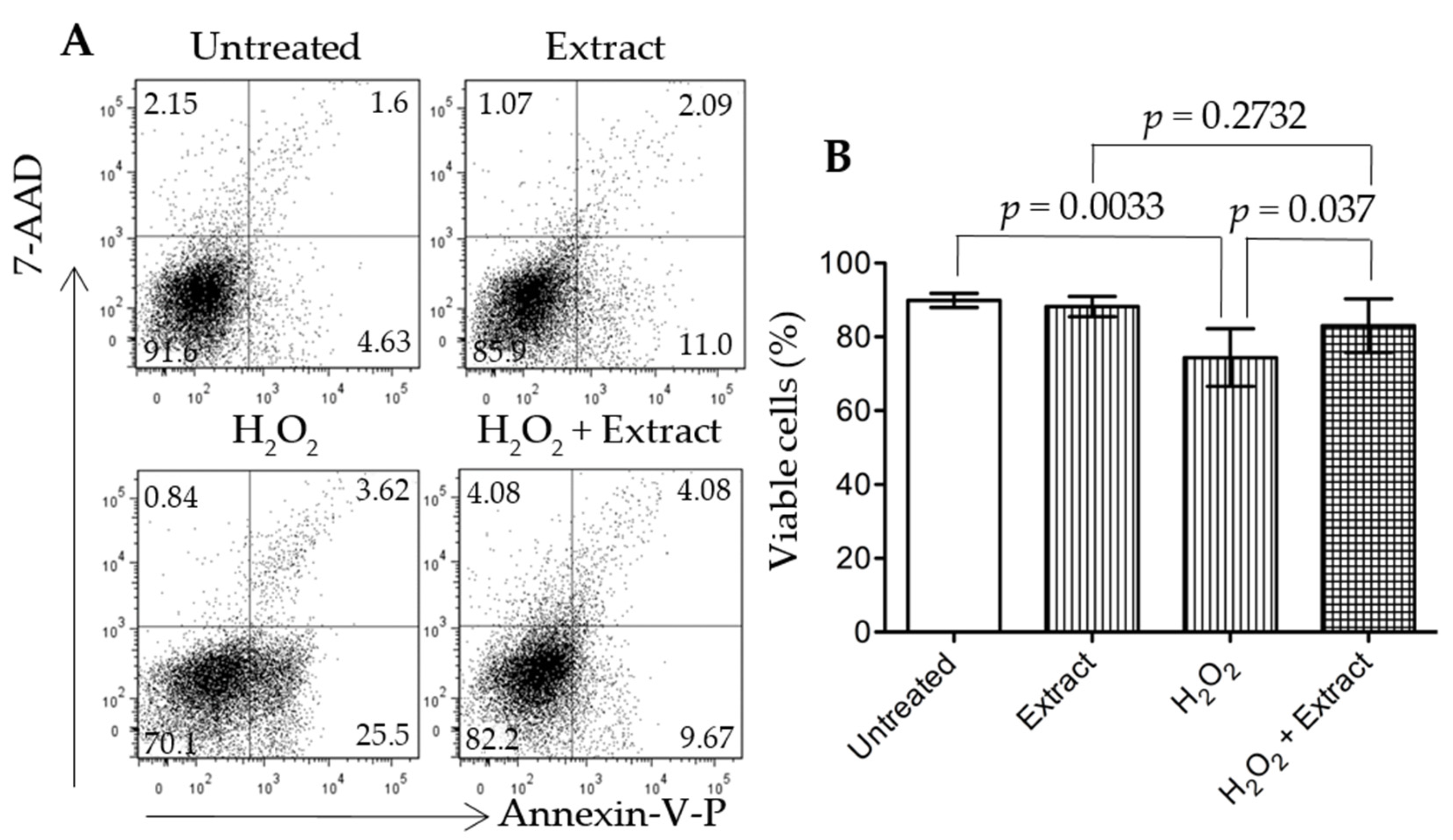
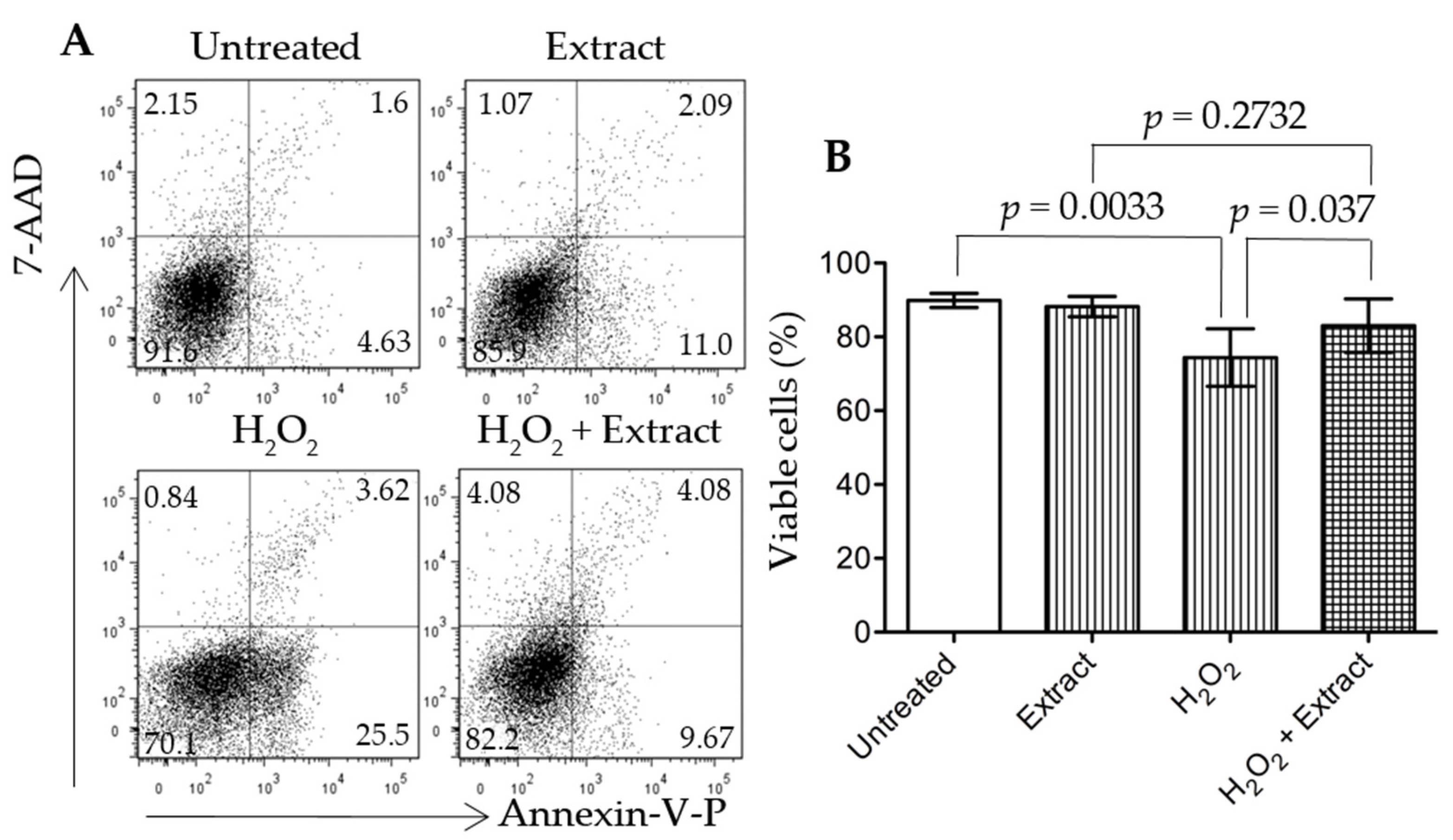
3.6. Addition of WGP Extract Reduced H2O2-Induced Cell Death in Cultured Podocytes
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Coresh, J.; Astor, B.C.; Greene, T.; Eknoyan, G.; Levey, A.S. Prevalence of chronic kidney disease and decreased kidney function in the adult us population: Third national health and nutrition examination survey. Am. J. Kidney Dis. 2003, 41, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.; Hemmelgarn, B.; Culleton, B.; Tobe, S.; McFarlane, P.; Ruzicka, M.; Burns, K.; Manns, B.; White, C.; Madore, F.; et al. Guidelines for the management of chronic kidney disease. CMAJ 2008, 179, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Stigant, C.; Stevens, L.; Levin, A. Nephrology: 4. Strategies for the care of adults with chronic kidney disease. CMAJ 2003, 168, 1553–1560. [Google Scholar] [PubMed]
- Arora, P.; Vasa, P.; Brenner, D.; Iglar, K.; McFarlane, P.; Morrison, H.; Badawi, A. Prevalence estimates of chronic kidney disease in canada: Results of a nationally representative survey. CMAJ 2013, 185, E417–E423. [Google Scholar] [CrossRef] [PubMed]
- Levey, A.S.; Coresh, J. Chronic kidney disease. Lancet 2012, 379, 165–180. [Google Scholar] [CrossRef]
- Kaur, J. A comprehensive review on metabolic syndrome. Cardiol. Res. Pract. 2014, 2014, 943162. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Kari, J.A. Metabolic syndrome and chronic kidney disease. Curr. Opin. Nephrol. Hypertens. 2013, 22, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Prasad, G.V. Metabolic syndrome and chronic kidney disease: Current status and future directions. World J. Nephrol. 2014, 3, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Thomas, G.; Sehgal, A.R.; Kashyap, S.R.; Srinivas, T.R.; Kirwan, J.P.; Navaneethan, S.D. Metabolic syndrome and kidney disease: A systematic review and meta-analysis. Clin. J. Am. Soc. Nephrol. 2011, 6, 2364–2373. [Google Scholar] [CrossRef] [PubMed]
- Nashar, K.; Egan, B.M. Relationship between chronic kidney disease and metabolic syndrome: Current perspectives. Diabetes Metab. Syndr. Obes. 2014, 7, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Asmat, U.; Abad, K.; Ismail, K. Diabetes mellitus and oxidative stress-a concise review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Betteridge, D.J. What is oxidative stress? Metabolism 2000, 49, 3–8. [Google Scholar] [CrossRef]
- Sadowska-Bartosz, I.; Bartosz, G. Effect of antioxidants supplementation on aging and longevity. Biomed. Res. Int. 2014, 2014, 404680. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Faulkner, K.; Plumb, G.W. Glucosinolates and phenolics as antioxidants from plant foods. Eur. J. Cancer Prev. 1998, 7, 17–21. [Google Scholar] [PubMed]
- Vendrame, S.; Del Bo, C.; Ciappellano, S.; Riso, P.; Klimis-Zacas, D. Berry fruit consumption and metabolic syndrome. Antioxidants (Basel) 2016, 5, 34. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, K.; Olejnik, A. Current evidence on the health-beneficial effects of berry fruits in the prevention and treatment of metabolic syndrome. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 446–452. [Google Scholar] [CrossRef] [PubMed]
- El-Mowafy, A.M.; Salem, H.A.; Al-Gayyar, M.M.; El-Mesery, M.E.; El-Azab, M.F. Evaluation of renal protective effects of the green-tea (EGCG) and red grape resveratrol: Role of oxidative stress and inflammatory cytokines. Nat. Prod. Res. 2011, 25, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Gonzalez, P.D.; Lopez-Hernandez, F.J.; Perez-Barriocanal, F.; Morales, A.I.; Lopez-Novoa, J.M. Quercetin reduces cisplatin nephrotoxicity in rats without compromising its anti-tumour activity. Nephrol. Dial. Transplant. 2011, 26, 3484–3495. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, K.; Tamura, Y.; Lanaspa, M.A.; Miyazaki, M.; Suzuki, N.; Sato, W.; Maeshima, Y.; Schreiner, G.F.; Villarreal, F.J.; Johnson, R.J.; et al. Epicatechin limits renal injury by mitochondrial protection in cisplatin nephropathy. Am. J. Physiol. Ren. Physiol. 2012, 303, F1264–F1274. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kang, M.K.; Kim, J.K.; Kim, J.L.; Kang, S.W.; Lim, S.S.; Kang, Y.H. Purple corn anthocyanins retard diabetes-associated glomerulosclerosis in mesangial cells and db/db mice. Eur. J. Nutr. 2012, 51, 961–973. [Google Scholar] [CrossRef] [PubMed]
- Pokkunuri, I.; Ali, Q.; Asghar, M. Grape powder improves age-related decline in mitochondrial and kidney functions in fischer 344 rats. Oxid. Med. Cell. Longev. 2016, 2016, 6135319. [Google Scholar] [CrossRef] [PubMed]
- Laguardia, H.A.; Hamm, L.L.; Chen, J. The metabolic syndrome and risk of chronic kidney disease: Pathophysiology and intervention strategies. J. Nutr. Metab. 2012, 2012, 652608. [Google Scholar] [CrossRef] [PubMed]
- Tofovic, S.P.; Kusaka, H.; Kost, C.K., Jr.; Bastacky, S. Renal function and structure in diabetic, hypertensive, obese ZDFxSHHF-hybrid rats. Ren. Fail. 2000, 22, 387–406. [Google Scholar] [CrossRef] [PubMed]
- Baynes, J.; Murray, D.B. Cardiac and renal function are progressively impaired with aging in zucker diabetic fatty type II diabetic rats. Oxid. Med. Cell. Longev. 2009, 2, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Rafikova, O.; Salah, E.M.; Tofovic, S.P. Renal and metabolic effects of tempol in obese ZSF1 rats—Distinct role for superoxide and hydrogen peroxide in diabetic renal injury. Metab. Clin. Exp. 2008, 57, 1434–1444. [Google Scholar] [CrossRef] [PubMed]
- Vora, J.P.; Zimsen, S.M.; Houghton, D.C.; Anderson, S. Evolution of metabolic and renal changes in the ZDF/DRT-fa rat model of type II diabetes. J. Am. Soc. Nephrol. 1996, 7, 113–117. [Google Scholar] [PubMed]
- Tofovic, S.P.; Jackson, E.K. Rat models of the metabolic syndrome. Methods Mol. Med. 2003, 86, 29–46. [Google Scholar] [PubMed]
- Bilan, V.P.; Salah, E.M.; Bastacky, S.; Jones, H.B.; Mayers, R.M.; Zinker, B.; Poucher, S.M.; Tofovic, S.P. Diabetic nephropathy and long-term treatment effects of rosiglitazone and enalapril in obese ZSF1 rats. J. Endocrinol. 2011, 210, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Tofovic, S.P.; Kusaka, H.; Jackson, E.K.; Bastacky, S.I. Renal and metabolic effects of caffeine in obese (fa/fa(cp)), diabetic, hypertensive ZSF1 rats. Ren. Fail. 2001, 23, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jia, Y.; Jackson, E.K.; Tofovic, S.P. 2-methoxyestradiol and 2-ethoxyestradiol retard the progression of renal disease in aged, obese, diabetic ZSF1 rats. J. Cardiovasc. Pharmacol. 2007, 49, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Griffin, S.V.; Hiromura, K.; Pippin, J.; Petermann, A.T.; Blonski, M.J.; Krofft, R.; Takahashi, S.; Kulkarni, A.B.; Shankland, S.J. Cyclin-dependent kinase 5 is a regulator of podocyte differentiation, proliferation, and morphology. Am. J. Pathol. 2004, 165, 1175–1185. [Google Scholar] [CrossRef]
- Brosius, F.C.; Coward, R.J. Podocytes, signaling pathways, and vascular factors in diabetic kidney disease. Adv. Chronic Kidney Dis. 2014, 21, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.A.; Nguyen, H.A.; Rifkin, D.E. Nutrition interventions in chronic kidney disease. Med. Clin. N. Am. 2016, 100, 1265–1283. [Google Scholar] [CrossRef] [PubMed]
- Chan, M. Protein-controlled versus restricted protein versus low protein diets in managing patients with non-dialysis chronic kidney disease: A single centre experience in Australia. BMC Nephrol. 2016, 17, 129. [Google Scholar] [CrossRef] [PubMed]
- Shah, B.V.; Patel, Z.M. Role of low protein diet in management of different stages of chronic kidney disease —Practical aspects. BMC Nephrol. 2016, 17, 156. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chou, J.; Chang, Y.; Lau, W.L.; Reddy, U.; Rhee, C.M.; Chen, J.; Hao, C.; Kalantar-Zadeh, K. The role of low protein diet in ameliorating proteinuria and deferring dialysis initiation: What is old and what is new. Panminerva Med. 2017, 59, 157–165. [Google Scholar] [PubMed]
- Turki, K.; Charradi, K.; Boukhalfa, H.; Belhaj, M.; Limam, F.; Aouani, E. Grape seed powder improves renal failure of chronic kidney disease patients. EXCLI J. 2016, 15, 424–433. [Google Scholar] [PubMed]
- Bernardi, C.; Moneta, D.; Brughera, M.; DiSalvo, M.; Lamparelli, D.; Mazue, G.; Iatropoulos, M.J. Haematology and clinical chemistry in rats: Comparison of different blood collection sites. Comp. Haematol. Int. 1996, 6, 160–166. [Google Scholar] [CrossRef]
- Vassalotti, J.A.; Stevens, L.A.; Levey, A.S. Testing for chronic kidney disease: A position statement from the national kidney foundation. Am. J. Kidney Dis. 2007, 50, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Arici, M. Management of Chronic Kidney Disease: A Clinician’s Guide; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Spira, A.; Gowrishankar, M.; Halperin, M.L. Factors contributing to the degree of polyuria in a patient with poorly controlled diabetes mellitus. Am. J. Kidney Dis. 1997, 30, 829–835. [Google Scholar] [CrossRef]
- Ahloulay, M.; Schmitt, F.; Dechaux, M.; Bankir, L. Vasopressin and urinary concentrating activity in diabetes mellitus. Diabetes Metab. 1999, 25, 213–222. [Google Scholar] [PubMed]
- Mogensen, C.E. Early glomerular hyperfiltration in insulin-dependent diabetics and late nephropathy. Scand. J. Clin. Lab. Investig. 1986, 46, 201–206. [Google Scholar] [CrossRef]
- Ahmed, S.; Lowder, G. Severity and stages of chronic kidney disease. Age 2012, 140, 13–25. [Google Scholar]
- Griffin, K.A.; Abu-Naser, M.; Abu-Amarah, I.; Picken, M.; Williamson, G.A.; Bidani, A.K. Dynamic blood pressure load and nephropathy in the ZSF1 (fa/fa cp) model of type 2 diabetes. Am. J. Physiol. Renal. Physiol. 2007, 293, F1605–F1613. [Google Scholar] [CrossRef] [PubMed]
- Tofovic, S.P.; Salah, E.M.; Jackson, E.K.; Melhem, M. Early renal injury induced by caffeine consumption in obese, diabetic zsf1 rats. Ren. Fail. 2007, 29, 891–902. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.; Hernandez-Mijares, A.; Garcia-Malpartida, K.; Banuls, C.; Bellod, L.; Victor, V.M. Mitochondria-targeted antioxidant peptides. Curr. Pharm. Des. 2010, 16, 3124–3131. [Google Scholar] [CrossRef] [PubMed]
- Novo, E.; Parola, M. Redox mechanisms in hepatic chronic wound healing and fibrogenesis. Fibrogenesis Tissue Repair 2008, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Babior, B.M. NADPH oxidase: An update. Blood 1999, 93, 1464–1476. [Google Scholar] [PubMed]
- Forman, H.J.; Torres, M. Redox signaling in macrophages. Mol. Aspects Med. 2001, 22, 189–216. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Wang, X.; Chen, H. Oxidative stress to renal tubular epithelial cells—A common pathway in renal pathologies. In Systems Biology of Free Radicals and Anti-Oxidants; Laher, I., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 2606–2624. [Google Scholar]
- Rhee, S.G. Redox signaling: Hydrogen peroxide as intracellular messenger. Exp. Mol. Med. 1999, 31, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Turkmen, K. Inflammation, oxidative stress, apoptosis, and autophagy in diabetes mellitus and diabetic kidney disease: The four horsemen of the apocalypse. Int. Urol. Nephrol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Sunny, N.E.; Bril, F.; Cusi, K. Mitochondrial adaptation in nonalcoholic fatty liver disease: Novel mechanisms and treatment strategies. Trends Endocrinol. Metab. 2017, 28, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Cruzat, V.F.; Keane, K.N.; Carlessi, R.; de Bittencourt, P.I., Jr. Molecular mechanisms of ROS production and oxidative stress in diabetes. Biochem. J. 2016, 473, 4527–4550. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, Y.; Toledo, R.T. Major flavonoids in grape seeds and skins: Antioxidant capacity of catechin, epicatechin, and gallic acid. J. Agric. Food Chem. 2004, 52, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.F.; Hoye, C., Jr.; Fernandez-Plotka, V.C. Influence of heating on the polyphenolic content and antioxidant activity of grape seed flour. J. Food Sci. 2011, 76, C884–C890. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xiao, Y.Y. Grape phytochemicals and associated health benefits. Crit. Rev. Food Sci. Nutr. 2013, 53, 1202–1225. [Google Scholar] [CrossRef] [PubMed]
- Aloui, F.; Charradi, K.; Hichami, A.; Subramaniam, S.; Khan, N.A.; Limam, F.; Aouani, E. Grape seed and skin extract reduces pancreas lipotoxicity, oxidative stress and inflammation in high fat diet fed rats. Biomed. Pharmacother. 2016, 84, 2020–2028. [Google Scholar] [CrossRef] [PubMed]
- Oueslati, N.; Charradi, K.; Bedhiafi, T.; Limam, F.; Aouani, E. Protective effect of grape seed and skin extract against diabetes-induced oxidative stress and renal dysfunction in virgin and pregnant rat. Biomed. Pharmacother. 2016, 83, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Freireich, E.J.; Gehan, E.A.; Rall, D.P.; Schmidt, L.H.; Skipper, H.E. Quantitative comparison of toxicity of anticancer agents in mouse, rat, hamster, dog, monkey, and man. Cancer Chemother. Rep. 1966, 50, 219–244. [Google Scholar] [PubMed]
- Johnson, D.W. Evidence-based guide to slowing the progression of early renal insufficiency. Intern. Med. J. 2004, 34, 50–57. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kidneys | Liver | Spleen | Heart | Lung | |
|---|---|---|---|---|---|
| Vehicle (n = 15) | 0.0070 ± 0.00072 | 0.0627 ± 0.00418 | 0.00135± 0.00010 | 0.00289 ± 0.00115 | 0.00356 ± 0.00096 |
| WGP (n = 14) | 0.0063 ± 0.00069 | 0.0544 ± 0.00546 | 0.00129 ± 0.00001 | 0.00251 ± 0.00022 | 0.00371 ± 0.00103 |
| p value * | 0.0127 | <0.0001 | 0.0341 | 0.2351 | 0.6880 |
| Gene Symbol | Gene Names | Functions | Fold Change * | p Value (n = 4) |
|---|---|---|---|---|
| Dhcr24 | 24-dehydrocholesterol reductase | H2O2 scavenger, preventing H2O2-induced cell death | 4.265 | 0.00222 |
| Cyba | Cytochrome b-245, alpha polypeptide | NADPH oxidase subunit, optimizing immunity | 4.215 | 0.00875 |
| Gstk1 | Glutathione S-transferase kappa 1 | Cellular detoxification (lipid peroxide detoxification) | 1.1475 | 0.01279 |
| Prdx2 | Peroxiredoxin 2 | H2O2 and Alkyl hydroperoxide antioxidant | 1.7625 | 0.02746 |
| Sod2 | Superoxide dismutase, mitochondrial | Limiting ROS detrimental effect, and moderating ROS release | 2.3375 | 0.02983 |
| Park7 | Parkinson disease (autosomal recessive, early onset) 7 | Redox-sensitive chaperone | 1.24 | 0.03723 |
| Gpx4 | Glutathione peroxidase 4 | H2O2, lipid peroxide and hydroperoxide reduction | 1.96 | 0.03756 |
| Gpx1 | Glutathione peroxidase 1 | H2O2 antioxidant | 3.235 | 0.04614 |
| Hmox1 | Heme oxygenase (decycling) 1 | Heme degradation to CO | −101.533 | 0.01283 |
| Ercc6 | Excision repair cross-complementing rodent repair deficiency, complementation group 6 | Damaged DNA repair | −3.9575 | 0.02908 |
| Gstp1 | Glutathione S-transferase pi 1 | Cellular detoxification | −22.4875 | 0.0405 |
| Txnip | Thioredoxin interacting protein | Increasing ROS production | −37.8825 | 0.04436 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almomen, S.M.K.; Guan, Q.; Liang, P.; Yang, K.; Sidiqi, A.M.; Levin, A.; Du, C. Daily Intake of Grape Powder Prevents the Progression of Kidney Disease in Obese Type 2 Diabetic ZSF1 Rats. Nutrients 2017, 9, 345. https://doi.org/10.3390/nu9040345
Almomen SMK, Guan Q, Liang P, Yang K, Sidiqi AM, Levin A, Du C. Daily Intake of Grape Powder Prevents the Progression of Kidney Disease in Obese Type 2 Diabetic ZSF1 Rats. Nutrients. 2017; 9(4):345. https://doi.org/10.3390/nu9040345
Chicago/Turabian StyleAlmomen, Salwa M. K., Qiunong Guan, Peihe Liang, Kaidi Yang, Ahmad M. Sidiqi, Adeera Levin, and Caigan Du. 2017. "Daily Intake of Grape Powder Prevents the Progression of Kidney Disease in Obese Type 2 Diabetic ZSF1 Rats" Nutrients 9, no. 4: 345. https://doi.org/10.3390/nu9040345