Beta Palmitate Improves Bone Length and Quality during Catch-Up Growth in Young Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Feeding Regimens
2.2. Procedure
2.3. Serum Analysis
2.4. Measurement of Crude Liver Lipid Content (Folch Method)
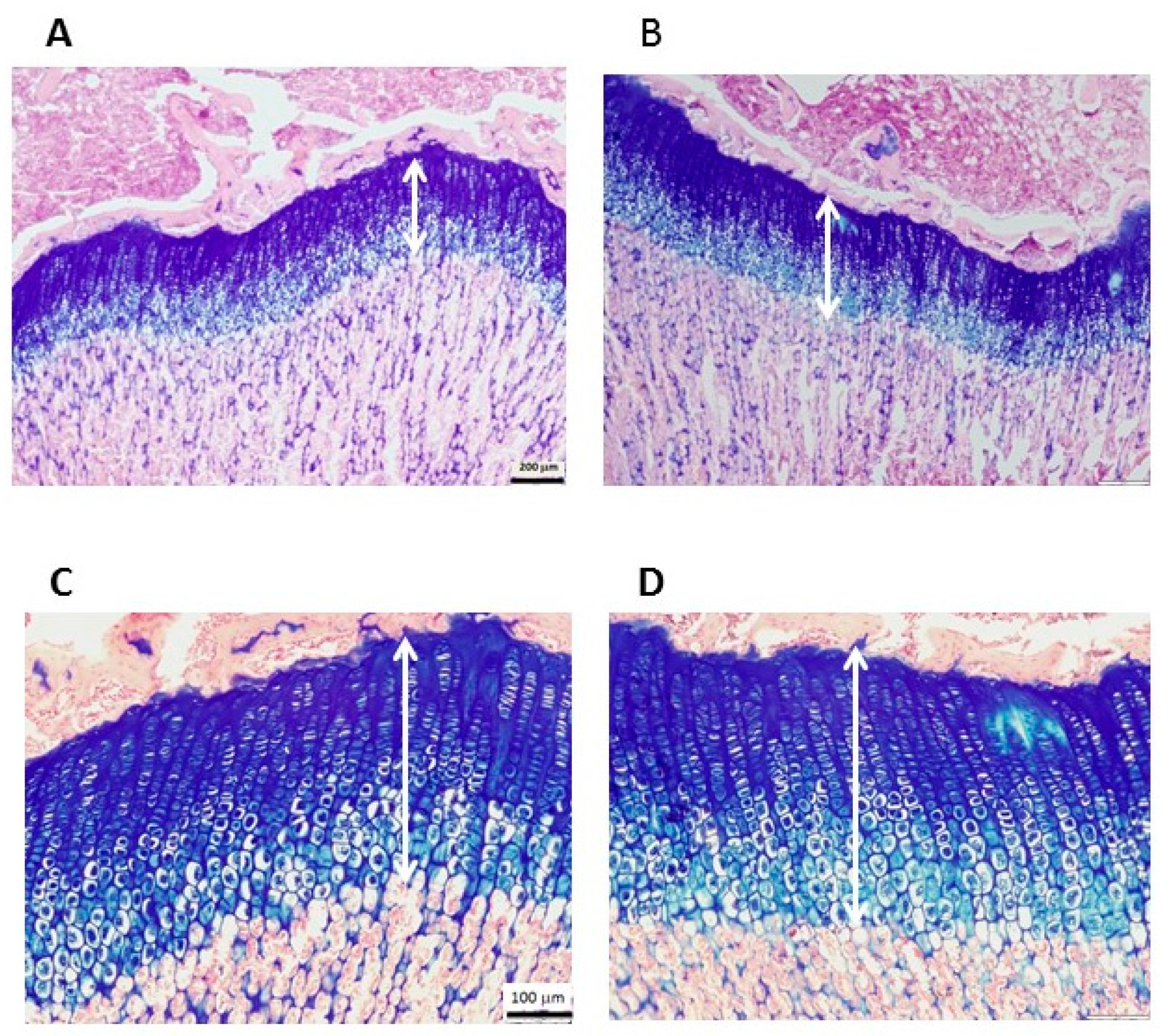
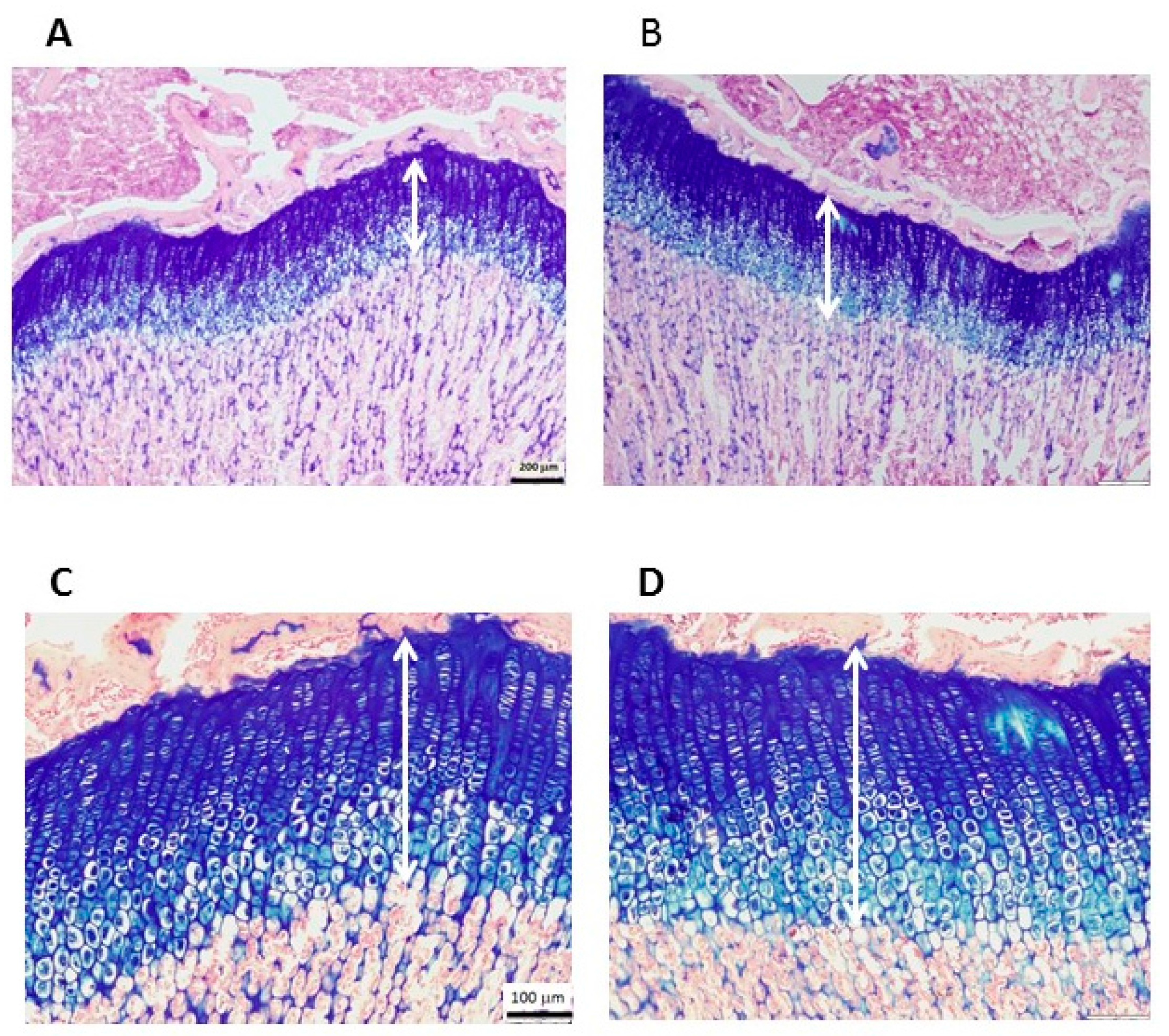
2.5. Histological Staining and Measurement of EGP Height
2.6. RNA Extraction and Affymetrix Analysis
2.7. Reverse Transcription and Real Time PCR for GDF-5
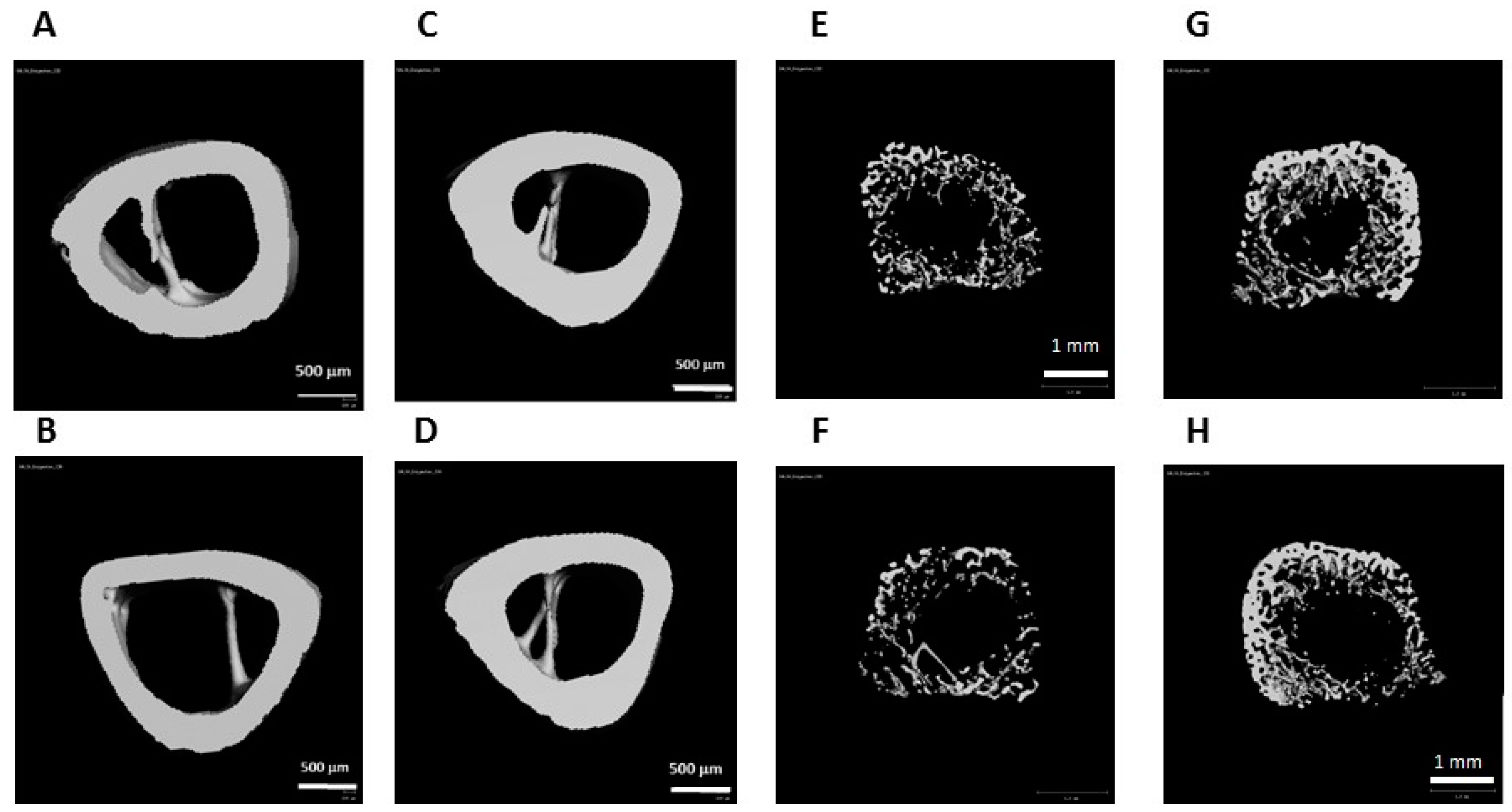
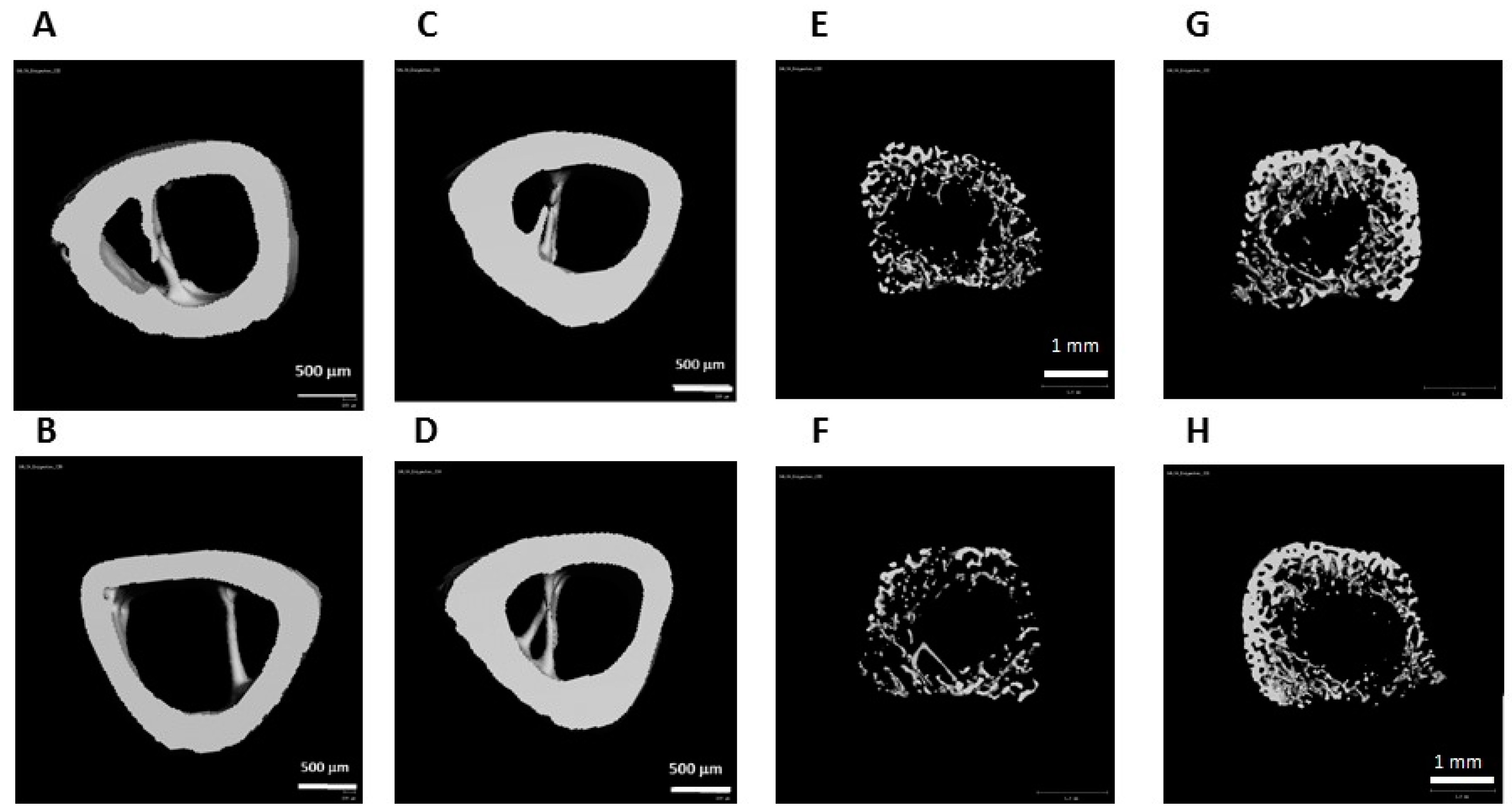
2.8. μCT Analysis
2.9. Biomechanical Analysis
2.10. Statistical Analysis
3. Results
3.1. Effect of the CD and IO Diets on Weight Gain and Serum Values in the Preliminary Experiment
3.2. Effect of the Study Diets on Weight Gain and Serum Values during CU Growth
3.3. Effect of the Study Diets on Linear Growth during CU Growth
3.4. Effect of the Study Diets on Bone Quality during CU Growth
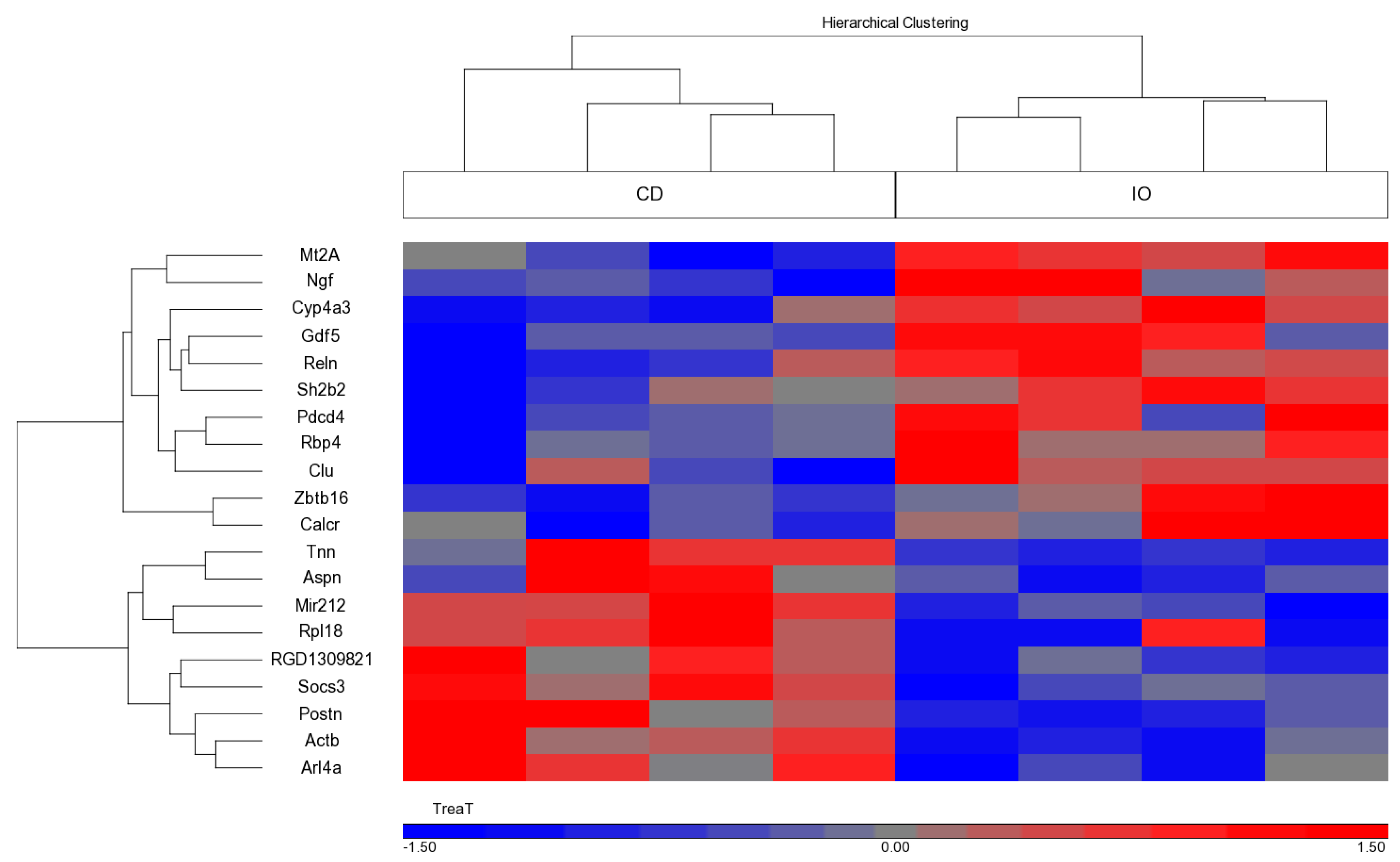
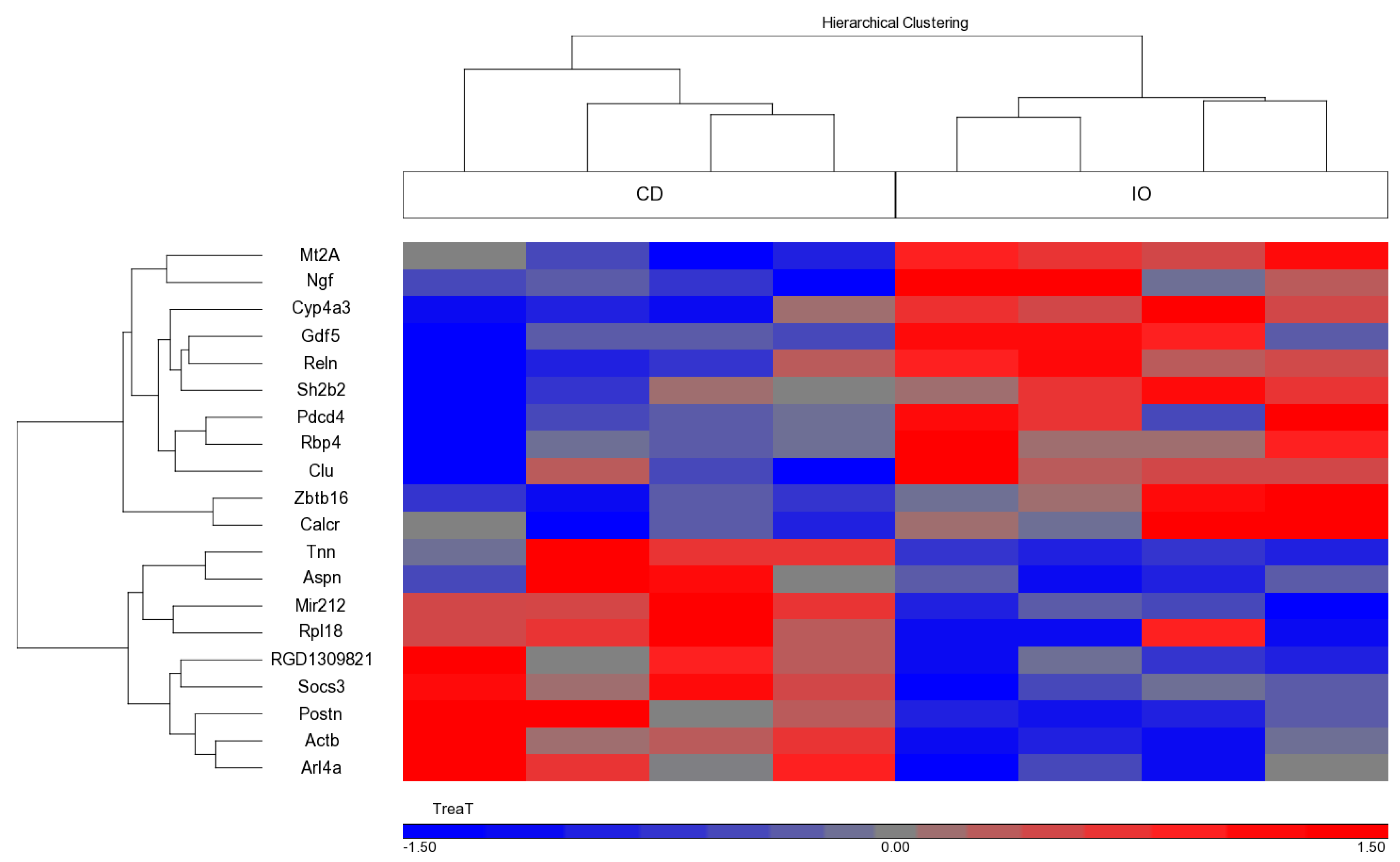
3.5. Effect of the Different Diets on Gene Expression during CU Growth
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Agostoni, C.; Braegger, C.; Desci, T.; Kolacek, S.; Koletzko, B.; Michaelsen, K.F.; Mihatsch, W.; Moreno, L.A.; Puntis, J.; Shamir, R.; et al. Breast feeding. A position paper by the ESPGHAN committee on nutrition. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Yackobovitch-Gavan, M.; Phillip, M.; Gat-Yablonski, G. How milk and its proteins affect growth, bone health, and weight. Horm. Res. Paediatr. 2017, 88. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; Agostoni, C.; Bergmann, R.; Ritzenthaler, K.; Shamir, R. Physiological aspects of human milk lipids and implications for infant feeding: A workshop report. Acta Paediatr. 2011, 100, 1405–1415. [Google Scholar] [CrossRef] [PubMed]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed]
- Havlicekova, Z.; Jesenak, M.; Banovcin, P.; Kuchta, M. Beta-palmitate—A natural component of human milk in supplemental milk formulas. Nutr. J. 2016, 15, 28. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, S.; Hernell, O. Lipid digestion and absorption in early life: An update. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Tomarelli, R.M.; Meyer, B.J.; Weaber, J.R.; Bernhart, F.W. Effect of positional distribution on the absorption of the fatty acids of human milk and infant formulas. J. Nutr. 1968, 95, 583–590. [Google Scholar] [PubMed]
- Innis, S.M.; Dyer, R.; Nelson, C.M. Evidence that palmitic acid is absorbed as sn-2 monoacylglycerol from human milk by breast-fed infants. Lipids 1994, 29, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.M.; Innis, S.M. Plasma lipoprotein fatty acids are altered by the positional distribution of fatty acids in infant formula triacylglycerols and human milk. Am. J. Clin. Nutr. 1999, 70, 62–69. [Google Scholar] [PubMed]
- Quinlan, P.T.; Lockton, S.; Irwin, J.; Lucas, A.L. The relationship between stool hardness and stool composition in breast- and formula-fed infants. J. Pediatr. Gastroenterol. Nutr. 1995, 20, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Carnielli, V.P.; Luijendijk, I.H.; van Goudoever, J.B.; Sulkers, E.J.; Boerlage, A.A.; Degenhart, H.J.; Sauer, P.J. Feeding premature newborn infants palmitic acid in amounts and stereoisomeric position similar to that of human milk: Effects on fat and mineral balance. Am. J. Clin. Nutr. 1995, 61, 1037–1042. [Google Scholar] [PubMed]
- Lucas, A.; Quinlan, P.; Abrams, S.; Ryan, S.; Meah, S.; Lucas, P.J. Randomised controlled trial of a synthetic triglyceride milk formula for preterm infants. Arch. Dis. Child. Fetal Neonatal. Ed. 1997, 77, F178–F184. [Google Scholar] [CrossRef] [PubMed]
- Carnielli, V.P.; Luijendijk, I.H.; Van Goudoever, J.B.; Sulkers, E.J.; Boerlage, A.A.; Degenhart, H.J.; Sauer, P.J. Structural position and amount of palmitic acid in infant formulas: Effects on fat, fatty acid, and mineral balance. J. Pediatr. Gastroenterol. Nutr. 1996, 23, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, K.; Fewtrell, M.S.; Morley, R.; Abbott, R.; Quinlan, P.T.; Wells, J.C.; Bindels, J.G.; Lucas, A. Double-blind, randomized trial of a synthetic triacylglycerol in formula-fed term infants: Effects on stool biochemistry, stool characteristics, and bone mineralization. Am. J. Clin. Nutr. 1999, 70, 920–927. [Google Scholar] [PubMed]
- Bongers, M.E.; de Lorijn, F.; Reitsma, J.B.; Groeneweg, M.; Taminiau, J.A.; Benninga, M.A. The clinical effect of a new infant formula in term infants with constipation: A double-blind, randomized cross-over trial. Nutr. J. 2007, 6. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, J.; Lee, H.C.; Lien, R.; Cheng, S.W.; Li, S.T.; Yao, M.; Northington, R.; Jan, I.; Mutungi, G. Stool fatty acid soaps, stool consistency and gastrointestinal tolerance in term infants fed infant formulas containing high sn-2 palmitate with or without oligofructose: A double-blind, randomized clinical trial. Nutr. J. 2014, 13, 105. [Google Scholar] [CrossRef] [PubMed]
- Litmanovitz, I.; Bar-Yoseph, F.; Lifshitz, Y.; Davidson, K.; Eliakim, A.; Regev, R.H.; Nemet, D. Reduced crying in term infants fed high beta-palmitate formula: A double-blind randomized clinical trial. BMC Pediatr. 2014, 14, 152. [Google Scholar] [CrossRef] [PubMed]
- Savino, F.; Palumeri, E.; Castagno, E.; Cresi, F.; Dalmasso, P.; Cavallo, F.; Oggero, R. Reduction of crying episodes owing to infantile colic: A randomized controlled study on the efficacy of a new infant formula. Eur. J. Clin. Nutr. 2006, 60, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Litmanovitz, I.; Davidson, K.; Eliakim, A.; Regev, R.H.; Dolfin, T.; Arnon, S.; Bar-Yoseph, F.; Goren, A.; Lifshitz, Y.; Nemet, D. High beta-palmitate formula and bone strength in term infants: A randomized, double-blind, controlled trial. Calcif. Tissue Int. 2013, 92, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Yaron, S.; Shachar, D.; Abramas, L.; Riskin, A.; Bader, D.; Litmanovitz, I.; Bar-Yoseph, F.; Cohen, T.; Levi, L.; Lifshitz, Y.; et al. Effect of high beta-palmitate content in infant formula on the intestinal microbiota of term infants. J. Pediatr. Gastroenterol. Nutr. 2013, 56, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Civardi, E.; Garofoli, F.; Longo, S.; Mongini, M.E.; Grenci, B.; Mazzucchelli, I.; Angelini, M.; Castellazzi, A.; Fasano, F.; Grinzato, A.; et al. Safety, growth, and support to healthy gut microbiota by an infant formula enriched with functional compounds. Clin. Nutr. 2015, 36, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Mackie, E.J.; Tatarczuch, L.; Mirams, M. The skeleton: A multi-functional complex organ: The growth plate chondrocyte and endochondral ossification. J. Endocrinol. 2011, 211, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Gebhardt, M.; Golovchenko, S.; Perez-Branguli, F.; Hattori, T.; Hartmann, C.; Zhou, X.; deCrombrugghe, B.; Stock, M.; Schneider, H.; et al. Dual pathways to endochondral osteoblasts: A novel chondrocyte-derived osteoprogenitor cell identified in hypertrophic cartilage. Biol. Open 2015, 4, 608–621. [Google Scholar] [CrossRef] [PubMed]
- Boersma, B.; Houwen, R.H.; Blum, W.F.; van Doorn, J.; Wit, J.M. Catch-up growth and endocrine changes in childhood celiac disease. Endocrine changes during catch-up growth. Horm. Res. 2002, 58, 57–65. [Google Scholar] [PubMed]
- Lebenthal, Y.; Yackobovitch-Gavan, M.; Lazar, L.; Shalitin, S.; Tenenbaum, A.; Shamir, R.; Phillip, M. Effect of a nutritional supplement on growth in short and lean prepubertal children: A prospective, randomized, double-blind, placebo-controlled study. J. Pediatr. 2014, 165, 1190–1193. [Google Scholar] [CrossRef] [PubMed]
- Yackobovitch-Gavan, M.; Lebenthal, Y.; Lazar, L.; Shalitin, S.; Demol, S.; Tenenbaum, A.; Shamir, R.; Phillip, M. Effect of nutritional supplementation on growth in short and lean prepubertal children after 1 year of intervention. J. Pediatr. 2016, 179, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Gat-Yablonski, G.; Yackobovitch-Gavan, M.; Phillip, M. Which dietary components modulate longitudinal growth? Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Rouy, E.; Vico, L.; Laroche, N.; Benoit, V.; Rousseau, B.; Blachier, F.; Tome, D.; Blais, A. Protein quality affects bone status during moderate protein restriction in growing mice. Bone 2014, 59, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Masarwi, M.; Gabet, Y.; Dolkart, O.; Brosh, T.; Shamir, R.; Phillip, M.; Gat-Yablonski, G. Skeletal effect of casein and whey protein intake during catch-up growth in young male Sprague-Dawley rats. Br. J. Nutr. 2016, 116, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Even-Zohar, N.; Jacob, J.; Amariglio, N.; Rechavi, G.; Potievsky, O.; Phillip, M.; Gat-Yablonski, G. Nutrition-induced catch-up growth increases hypoxia inducible factor 1alpha RNA levels in the growth plate. Bone 2008, 42, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P. The laboratory rat: Relating its age with human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar] [PubMed]
- Borjesson, A.E.; Windahl, S.H.; Karimian, E.; Eriksson, E.E.; Lagerquist, M.K.; Engdahl, C.; Antal, M.C.; Krust, A.; Chambon, P.; Savendahl, L.; et al. The role of estrogen receptor-alpha and its activation function-1 for growth plate closure in female mice. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1381–E1389. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, O.; Weise, M.; Landman, E.B.; Meyers, J.L.; Barnes, K.M.; Baron, J. Evidence that estrogen hastens epiphyseal fusion and cessation of longitudinal bone growth by irreversibly depleting the number of resting zone progenitor cells in female rabbits. Endocrinology 2014, 155, 2892–2899. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Gat-Yablonski, G.; Finka, A.; Pinto, G.; Quadroni, M.; Shtaif, B.; Goloubinoff, P. Quantitative proteomics of rat livers shows that unrestricted feeding is stressful for proteostasis with implications on life span. Aging 2016, 8, 1735–1758. [Google Scholar] [CrossRef] [PubMed]
- Bouxsein, M.L.; Boyd, S.K.; Christiansen, B.A.; Guldberg, R.E.; Jepsen, K.J.; Muller, R. Guidelines for assessment of bone microstructure in rodents using micro-computed tomography. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2010, 25, 1468–1486. [Google Scholar] [CrossRef] [PubMed]
- Gabet, Y.; Kohavi, D.; Voide, R.; Mueller, T.L.; Muller, R.; Bab, I. Endosseous implant anchorage is critically dependent on mechanostructural determinants of peri-implant bone trabeculae. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2010, 25, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Broz, J.J.; Simske, S.J.; Greenberg, A.R.; Luttges, M.W. Effects of rehydration state on the flexural properties of whole mouse long bones. J. Biomech. Eng. 1993, 115, 447–449. [Google Scholar] [CrossRef] [PubMed]
- Gabet, Y.; Muller, R.; Levy, J.; Dimarchi, R.; Chorev, M.; Bab, I.; Kohavi, D. Parathyroid hormone 1–34 enhances titanium implant anchorage in low-density trabecular bone: A correlative micro-computed tomographic and biomechanical analysis. Bone 2006, 39, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Shtaif, B.; Dror, N.; Bar-Maisels, M.; Phillip, M.; Gat-Yablonski, G. Growth without growth hormone: Can growth and differentiation factor 5 be the mediator? Growth Factors 2015, 33, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Pando, R.; Masarwi, M.; Shtaif, B.; Idelevich, A.; Monsonego-Ornan, E.; Shahar, R.; Phillip, M.; Gat-Yablonski, G. Bone quality is affected by food restriction and by nutrition-induced catch-up growth. J. Endocrinol. 2014, 223, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Mosier, H.D., Jr.; Jansons, R.A. Growth hormone during catch-up growth and failure of catch-up growth in rats. Endocrinology 1976, 98, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Hermanussen, M.; Rol de Lama, M.A.; Romero, A.P.; Ruiz, C.A.; Burmeister, J.; Tresguerres, J.A. Differential catch-up in body weight and bone growth after short-term starvation in rats. Growth Regul. 1996, 6, 230–237. [Google Scholar] [PubMed]
- Pando, R.; Even-Zohar, N.; Shtaif, B.; Edry, L.; Shomron, N.; Phillip, M.; Gat-Yablonski, G. MicroRNAs in the growth plate are responsive to nutritional cues: Association between miR-140 and SIRT1. J. Nutr. Biochem. 2012, 23, 1474–1481. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.B.; Kubota, S.; Ban, N.; Yoshida, M.; Santeford, A.; Sene, A.; Nakamura, R.; Zapata, N.; Kubota, M.; Tsubota, K.; et al. NAMPT-mediated NAD(+) biosynthesis is essential for vision in mice. Cell. Rep. 2016, 17, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Imai, S.; Yoshino, J. The importance of NAMPT/NAD/SIRT1 in the systemic regulation of metabolism and ageing. Diabetes Obes. Metab. 2013, 15, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Kanfi, Y.; Peshti, V.; Gozlan, Y.M.; Rathaus, M.; Gil, R.; Cohen, H.Y. Regulation of SIRT1 protein levels by nutrient availability. FEBS Lett. 2008, 582, 2417–2423. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, T.; Vaisanen, S.; Rakhshandehroo, M.; Kersten, S.; Carlberg, C. Peroxisome proliferator-activated receptor alpha controls hepatic heme biosynthesis through ALAS1. J. Mol. Biol. 2009, 388, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Bruning, J.C.; Winnay, J.; Cheatham, B.; Kahn, C.R. Differential signaling by insulin receptor substrate 1 (IRS-1) and IRS-2 in IRS-1-deficient cells. Mol. Cell. Biol. 1997, 17, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Tenorio, J.; Mansilla, A.; Valencia, M.; Martinez-Glez, V.; Romanelli, V.; Arias, P.; Castrejon, N.; Poletta, F.; Guillen-Navarro, E.; Gordo, G.; et al. A new overgrowth syndrome is due to mutations in RNF125. Hum. Mutat. 2014, 35, 1436–1441. [Google Scholar] [CrossRef] [PubMed]
- Mesarwi, O.A.; Shin, M.K.; Drager, L.F.; Bevans-Fonti, S.; Jun, J.C.; Putcha, N.; Torbenson, M.S.; Pedrosa, R.P.; Lorenzi-Filho, G.; Steele, K.E.; et al. Lysyl oxidase as a serum biomarker of liver fibrosis in patients with severe obesity and obstructive sleep apnea. Sleep 2015, 38, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Sharma-Bhandari, A.; Park, S.H.; Kim, J.Y.; Oh, J.; Kim, Y. Lysyl oxidase modulates the osteoblast differentiation of primary mouse calvaria cells. Int. J. Mol. Med. 2015, 36, 1664–1670. [Google Scholar] [CrossRef] [PubMed]
- Pasco, M.Y.; Leopold, P. High sugar-induced insulin resistance in Drosophila relies on the lipocalin Neural Lazarillo. PLoS ONE 2012, 7, e36583. [Google Scholar] [CrossRef] [PubMed]
- Scharfmann, R.; Atouf, F.; Tazi, A.; Czernichow, P. Growth hormone and prolactin regulate the expression of nerve growth factor receptors in INS-1 cells. Endocrinology 1994, 134, 2321–2328. [Google Scholar] [CrossRef] [PubMed]
- Duval, E.; Bigot, N.; Hervieu, M.; Kou, I.; Leclercq, S.; Galera, P.; Boumediene, K.; Bauge, C. Asporin expression is highly regulated in human chondrocytes. Mol. Med. 2011, 17, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Belter, A.; Gudanis, D.; Rolle, K.; Piwecka, M.; Gdaniec, Z.; Naskret-Barciszewska, M.Z.; Barciszewski, J. Mature miRNAs form secondary structure, which suggests their function beyond RISC. PLoS ONE 2014, 9, e113848. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, N.; Garnero, P.; Ferrari, S. Periostin action in bone. Mol. Cell. Endocrinol. 2016, 432, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Buxton, P.; Edwards, C.; Archer, C.W.; Francis-West, P. Growth/differentiation factor-5 (GDF-5) and skeletal development. J. Bone Jt. Surg. Am. 2001, 83, S23–S30. [Google Scholar] [CrossRef]
- Erlacher, L.; McCartney, J.; Piek, E.; ten Dijke, P.; Yanagishita, M.; Oppermann, H.; Luyten, F.P. Cartilage-derived morphogenetic proteins and osteogenic protein-1 differentially regulate osteogenesis. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 1998, 13, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Yamano, S.; Haku, K.; Yamanaka, T.; Dai, J.; Takayama, T.; Shohara, R.; Tachi, K.; Ishioka, M.; Hanatani, S.; Karunagaran, S.; et al. The effect of a bioactive collagen membrane releasing PDGF or GDF-5 on bone regeneration. Biomaterials 2014, 35, 2446–2453. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.S.; Ohe, J.Y.; Lee, J.B.; Heo, D.N.; Byun, W.; Bae, H.; Kwon, Y.D.; Kwon, I.K. Photo-cured hyaluronic acid-based hydrogels containing growth and differentiation factor 5 (GDF-5) for bone tissue regeneration. Bone 2014, 59, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Degenkolbe, E.; Schwarz, C.; Ott, C.E.; Konig, J.; Schmidt-Bleek, K.; Ellinghaus, A.; Schmidt, T.; Lienau, J.; Ploger, F.; Mundlos, S.; et al. Improved bone defect healing by a superagonistic GDF5 variant derived from a patient with multiple synostoses syndrome. Bone 2015, 73, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, Y.; Mabuchi, A.; Shi, D.; Kubo, T.; Takatori, Y.; Saito, S.; Fujioka, M.; Sudo, A.; Uchida, A.; Yamamoto, S.; et al. A functional polymorphism in the 5′ UTR of GDF5 is associated with susceptibility to osteoarthritis. Nat. Genet. 2007, 39, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.L.; Vieira-Silva, S.; Liston, A.; Raes, J. How informative is the mouse for human gut microbiota research? Dis. Models Mech. 2015, 8, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Backhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Growth Parameters | Control 1,3 Diet (CD) (n = 8) (Mean ± SD) | Infat Oil (BPA) Diet (IO) (n = 8) (Mean ± SD) | T-Test (p Value) |
|---|---|---|---|
| Weight (g) | 131.3 ± 7.9 | 135.6 ± 8.4 | 0.31 |
| Weight gain (g) | 83.5 ± 5.1 | 87.9 ± 6.4 | 0.17 |
| Liver weight (g) | 6.9 ± 0.3 | 6.8 ± 0.5 | 0.81 |
| Liver fat content (mg/100 mg tissue) | 4.5 ± 0.5 | 4.7 ± 0.7 | 0.64 |
| Full humerus length (mm) | 19.86 ± 0.5 | 20.4 ± 0.3 | 0.042 |
| EGP height (mm) | 0.39 ± 0.02 | 0.41 ± 0.02 | 0.06 |
| Proliferative (mm) | 0.22 ± 0.04 | 0.21 ± 0.03 | 0.44 |
| Hypertrophic (mm) | 0.18 ± 0.02 | 0.2 ± 0.02 | 0.02 |
| Proliferative/hypertrophic | 1.21 ± 0.18 | 1.03 ± 0.14 | 0.05 |
| IGF-I (ng/mL) | 868.1 ± 193.7 | 894.2 ± 108.1 | 0.78 |
| Leptin (pg/mL) | 1461 ± 196 | 1448 ± 434.8 | 0.94 |
| Bone Parameters | Control 1,3 Diet (CD) (n = 8) (Mean ± SD) | Infat Oil (BPA) Diet (IO) (n = 8) (Mean ± SD) | T-Test (p Value) |
|---|---|---|---|
| Full volumetric bone mineral density (vBMD) (mg HA/cm3g) | 296.7 ± 14.85 | 306.58 ± 15.2 | 0.262 |
| (A) Cortical bone parameters | |||
| Tt.Ar (mm2) | 3.15 ± 0.07 | 3.07 ± 0.11 | 0.51 |
| Ct.Ar (mm2) | 1.75 ± 0.1 | 1.83 ± 0.12 | 0.31 |
| Ct.Ar/Tt.Ar (%) | 55 ± 0.02 | 59 ± 0.04 | 0.058 |
| Ct.Th (mm) | 0.31 ± 0.01 | 0.35 ± 0.03 | 0.026 |
| (B) Trabecular bone parameters (tROI) | |||
| BV/TV (%) | 3.9 ± 1.31 | 5.26 ± 2.02 | 0.17 |
| Tb.Th (mm) | 0.05 ± 0.004 | 0.06 ± 0.01 | 0.11 |
| Tb.N (mm-1) | 1.06 ± 0.15 | 1.19 ± 0.26 | 0.28 |
| Tb.Sp (mm) | 0.96 ± 0.14 | 0.89 ± 0.15 | 0.41 |
| (B1) Proximal ROI (proximal metaphysis) | |||
| BV/TV (%) | 5.7 ± 2 | 7.5 ± 3 | 0.22 |
| Tb.Th (mm) | 0.05 ± 0.004 | 0.06 ± 0.01 | 0.11 |
| Tb.N (mm−1) | 1.46 a ± 0.2 | 1.54 ± 0.29 | 0.55 |
| Tb.Sp (mm) | 0.71 ± 0.09 | 0.69 ± 0.11 | 0.73 |
| (B2) Distal ROI (proximal metaphysis) | |||
| BV/TV (%) | 0.72 ± 0.15 | 1.25 ± 0.5 | 0.015 |
| Tb.Th (mm) | 0.05 ± 0.0 | 0.05 ± 0.01 | 0.98 |
| Tb.N (mm−1) | 0.71 ± 0.1 | 0.84 ± 0.21 | 0.154 |
| Tb.Sp (mm) | 1.42 ± 0.21 | 1.25 ± 0.23 | 0.21 |
| (C) Bone biomechanical properties | |||
| Stiffness (N/mm) | 41.68 ± 14.4 | 36.37 ± 7.6 | 0.43 |
| Maximal load (N) | 22.8 ± 4.3 | 21.54 ± 3.5 | 0.53 |
| Energy to maximum (N*mm) | 14.02 ± 3.5 | 16.5 ± 5.0 | 0.3 |
| Gene Symbol | Gene Name | Gene ID | Fold Change (IO/CD) |
|---|---|---|---|
| Mt2A | Metallothionein 2A | NM_001137564 | 1.56 |
| Rbp4 | Retinol binding protein 4 | NM_013162 | 1.5 |
| Calcr | Calcitonin receptor | NM_001034015 | 1.42 |
| Reln | Reelin | NM_080394 | 1.37 |
| Clu | Clusterin | NM_053021 | 1.36 |
| Sh2b2 | SH2B adaptor protein 2 | NM_053669 | 1.35 |
| Ngf | Nerve growth factor (beta polypeptide) | NM_001277055 | 1.35 |
| Pdcd4 | Programmed cell death 4 | NM_022265 | 1.34 |
| Cyp4a3 | Cytochrome P450, family 4, subfamily a, polypeptide 3 | NM_175760 | 1.3 |
| Gdf5 | Growth differentiation factor 5 | ENSRNOT00000073736 | 1.28 |
| Zbtb16 | Zinc finger and DTD domain containing 16 | NM_001013181 | 1.28 |
| Actb | Actin, beta | ENSRNOT00000034844 | −1.26 |
| Socs3 | Suppressor of cytokine signaling 3 | NM_053565 | −1.26 |
| Arl4a | ADP-ribosylation factor like GTPase 4A | NM_019186 | −1.26 |
| Mir212 | microRNA 212 | NR_031925 | −1.27 |
| RGD1309821 | Similar to KIAA1161 protein | NSRNOT00000033235 | −1.34 |
| Rpl18 | Ribosomal protein L18 | FQ229993 | −1.35 |
| Postn | Periostin, osteoblast specific factor | NM_001108550 | −1.43 |
| Tnn | Tenascin N | NM_001107189 | −1.56 |
| Aspn | Asporin | NM_001014008 | −1.72 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bar-Maisels, M.; Gabet, Y.; Shamir, R.; Hiram-Bab, S.; Pasmanik-Chor, M.; Phillip, M.; Bar-Yoseph, F.; Gat-Yablonski, G. Beta Palmitate Improves Bone Length and Quality during Catch-Up Growth in Young Rats. Nutrients 2017, 9, 764. https://doi.org/10.3390/nu9070764
Bar-Maisels M, Gabet Y, Shamir R, Hiram-Bab S, Pasmanik-Chor M, Phillip M, Bar-Yoseph F, Gat-Yablonski G. Beta Palmitate Improves Bone Length and Quality during Catch-Up Growth in Young Rats. Nutrients. 2017; 9(7):764. https://doi.org/10.3390/nu9070764
Chicago/Turabian StyleBar-Maisels, Meytal, Yankel Gabet, Raanan Shamir, Sahar Hiram-Bab, Metsada Pasmanik-Chor, Moshe Phillip, Fabiana Bar-Yoseph, and Galia Gat-Yablonski. 2017. "Beta Palmitate Improves Bone Length and Quality during Catch-Up Growth in Young Rats" Nutrients 9, no. 7: 764. https://doi.org/10.3390/nu9070764