Effect of Increased Temperature on Native and Alien Nuisance Cyanobacteria from Temperate Lakes: An Experimental Approach

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
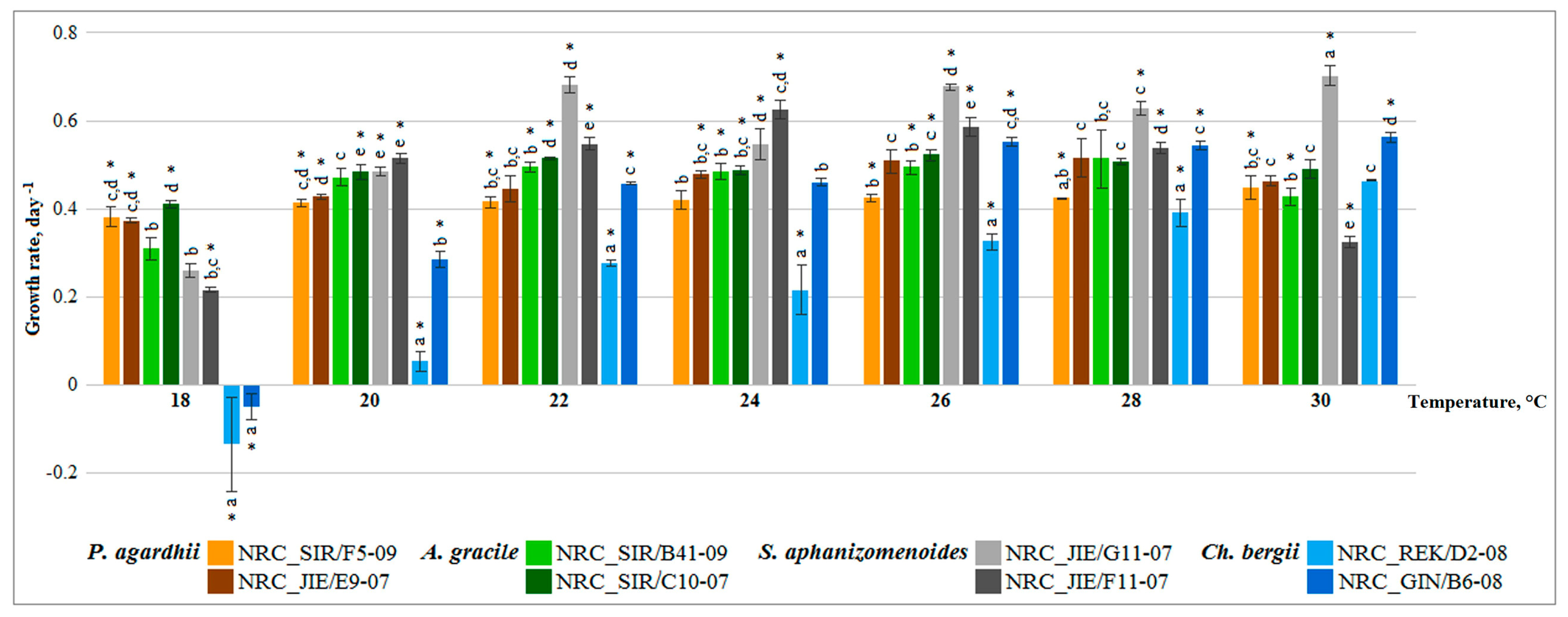
2.1. Growth Rate of Cyanobacteria Monocultures at Different Temperatures
2.2. Cyanotoxins and Oligopeptides Composition
3. Discussion
3.1. Cyanobacteria Response to Temperature Increase
3.2. Secondary Metabolites’ Variation in Response to Temperature
4. Conclusions
5. Materials and Methods
5.1. Cyanobacteria Isolates
5.2. Incubation and Experimental Design
5.3. Growth Evaluation
5.4. Cyanotoxins and Oligopeptides Analysis
5.5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brown, P.T.; Caldeira, K. Greater future global warming inferred from Earth’s recent energy budget. Nature 2017, 552, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgle, P.M. Climate Change 2013: The Physical Science Basis, Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; 1535p, ISBN 978-1-107-66182-0. [Google Scholar]
- Christensen, J.H.; Hewitson, B.; Busuioc, A.; Chen, A.; Gao, X.; Held, R.; Jones, R.; Kolli, R.K.; Kwon, W.K.; Laprise, R.; et al. Regional climate projections. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007; pp. 847–940. ISBN 978-0-521-70596-7. [Google Scholar]
- Kovats, R.S.; Valentini, R.; Bouwer, L.M.; Georgopoulou, E.; Jacob, D.; Martin, E.; Rounsevell, M.; Soussana, J.-F. Europe. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Barros, V.R., Field, C.B., Dokken, D.J., Mastrandrea, M.D., Mach, K.J., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 1267–1326. ISBN 978-92-9169-141-8. [Google Scholar]
- IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Summaries, Frequently Asked Questions, and Cross-Chapter Boxes. A Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; World Meteorological Organization: Geneva, Switzerland, 2014; 190p, ISBN 978-92-9169-141-8. [Google Scholar]
- Meehl, G.A.; Stocker, T.F.; Collins, W.D.; Friedlingstein, P.; Gaye, T.; Gregory, J.M.; Kitoh, A.; Knutti, R.; Murphy, J.M.; Noda, A.; et al. Global climate projections. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007; pp. 747–845. ISBN 978-0-521-70596-7. [Google Scholar]
- Jöhnk, K.D.; Huisman, J.; Sharples, J.; Sommeijer, B.; Visser, P.M.; Stroom, J.M. Summer heatwaves promote blooms of harmful cyanobacteria. Glob. Chang. Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Meerhoff, M.; Davidson, T.A.; Trolle, D.; Sondergaard, M.; Lauridsen, T.L.; Beklioğlu, M.; Brucet, S.; Volta, P.; Gonzalez-Bergonzoni, I.; Nielsen, A. Climate change impacts on lakes: An integrated ecological perspective based on a multi-faceted approach, with special focus on shallow lakes. J. Limnol. 2014, 73, 88–111. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Kronvang, B.; Meerhoff, M.; Sondergaard, M.; Hansen, K.M.; Andersen, H.E.; Lauridsen, T.L.; Liboriussen, L.; Beklioglu, M.; Ozen, A.; et al. Climate change effects on runoff, catchment phosphorus loading and lake ecological state, and potential adaptations. J. Environ. Qual. 2009, 38, 1930–1941. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Lürling, M.; Eshetu, F.; Faassen, E.J.; Kosten, S.; Huszar, V.L.M. Comparison of cyanobacterial and green algal growth rates at different temperatures. Freshw. Biol. 2013, 58, 552–559. [Google Scholar] [CrossRef]
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharm. 2005, 203, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Wiegand, C. Cyanobacterial toxins—Occurrence, biosynthesis and impact on human affairs. Mol. Nutr. Food Res. 2006, 50, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H. Nutrient and other environmental controls of harmful cyanobacterial blooms along the freshwater–marine continuum. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Hudnell, H.K., Ed.; Springer Science & Business Media: Berlin, Germany, 2008; Volume 619, pp. 216–241. [Google Scholar]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Scherer, P.I.; Raeder, U.; Geist, J.; Zwirglmaier, K. Influence of temperature, mixing, and addition of microcystin-LR on microcystin gene expression in Microcystis aeruginosa. MicrobiologyOpen 2016, 6. [Google Scholar] [CrossRef]
- Cirés, S.; Delgado, A.; González-Pleiter, M.; Quesada, A. Temperature Influences the Production and Transport of Saxitoxin and the Expression of sxt Genes in the Cyanobacterium Aphanizomenon gracile. Toxins 2017, 9, 322. [Google Scholar] [CrossRef] [PubMed]
- Ibelings, B.W. Cyanobacterial Waterblooms: The Role of Buoyancy in Water Columns of Varying Stability. Ph.D. Thesis, University of Amsterdam, Amsterdam, The Netherlands, 1992. [Google Scholar]
- Komárek, J. Süßwasserflora von Mitteleuropa (19/3): Cyanoprokaryota. Heterocytous Types; Springer: Berlin/Heidelberg, Germany, 2013; 1130p, ISBN 978-3-8274-0932-4. [Google Scholar]
- Mantzouki, E.; Visser, P.M.; Bormans, M.; Ibelings, B.W. Understanding the key ecological traits of cyanobacteria as a basis for their management and control in changing lakes. Aquat. Ecol. 2016, 50, 333–350. [Google Scholar] [CrossRef]
- Mischke, U.; Nixdorf, B. Equilibrium phase conditions in shallow German lakes: How Cyanoprokaryota species establish a steady state phase in late summer. Hydrobiologia 2003, 502, 123–132. [Google Scholar] [CrossRef]
- Grabowska, M.; Kobos, J.; Toruńska-Sitarz, A.; Mazur-Marzec, H. Non-ribosomal peptides produced by Planktothrix agardhii from Siemianówka Dam Reservoir SDR (northeast Poland). Arch. Microbiol. 2014, 196, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Toporowska, M.; Pawlik-Skowrońska, B.; Kalinowska, R. Mass Development of Diazotrophic Cyanobacteria (Nostocales) and Production of Neurotoxic Anatoxin-a in a Planktothrix (Oscillatoriales) Dominated Temperate Lake. Water Air Soil Pollut. 2016, 227, 321. [Google Scholar] [CrossRef] [PubMed]
- Kokociński, M.; Mankiewicz-Boczek, J.; Jurczak, T.; Spoof, L.; Meriluoto, J.; Rejmonczyk, E.; Hautala, H.; Vehniäinen, M.; Pawełczyk, J.; Soininen, J. Aphanizomenon gracile (Nostocales), a cylindrospermopsin-producing cyanobacterium in Polish lakes. Environ. Sci. Pollut. Res. 2013, 20, 5243–5264. [Google Scholar] [CrossRef] [PubMed]
- Bernard, C.; Ballot, A.; Thomazeau, S.; Maloufi, S.; Furey, A.; Mankiewicz-Boczek, J.; Pawlik-Skowrońska, B.; Capelli, C.; Salmaso, N. Cyanobacteria associated with the production of cyanotoxins. Appendix 2. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; Wiley: Hoboken, NJ, USA, 2017; pp. 501–525. ISBN 978-1-119-06868-6. [Google Scholar]
- Sukenik, A.; Hadas, O.; Kaplan, A.; Quesada, A. Invasion of Nostocales (cyanobacteria) to subtropical and temperate freshwater lakes—Physiological, regional, and global driving forces. Front. Microbiol. 2012, 3, 86. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.K.; Litchman, E. Effects of temperature and nitrogen availability on the growth of invasive and native cyanobacteria. Hydrobiologia 2016, 763, 357–369. [Google Scholar] [CrossRef]
- Padisák, J.; Vasas, G.; Borics, G. Phycogeography of freshwater phytoplankton: Traditional knowledge and new molecular tools. Hydrobiologia 2015, 764, 3–27. [Google Scholar] [CrossRef]
- Sukenik, A.; Quesada, A.; Salmaso, N. Global expansion of toxic and non-toxic cyanobacteria: Effect on ecosystem functioning. Biodivers. Conserv. 2015, 24, 889–908. [Google Scholar] [CrossRef]
- Meriluoto, J.; Blaha, L.; Bojadzija, G.; Bormans, M.; Brient, L.; Codd, G.A.; Drobac, D.; Faassen, E.J.; Fastner, J.; Hiskia, A.; et al. Toxic cyanobacteria and cyanotoxins in European waters—Recent progress achieved through the CYANOCOST Action and challenges for further research. Adv. Oceanogr. Limnol. 2017, 8, 161–178. [Google Scholar] [CrossRef]
- Schembri, M.A.; Neilan, B.A.; Saint, C.P. Identification of genes implicated in toxin production in the cyanobacterium Cylindrospermopsis raciborskii. Environ. Toxicol. 2001, 16, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Sabour, B.; Loudiki, M.; Oudra, B.; Vasconcelos, V.; Oubraim, S.; Fawzi, B. Dynamics and toxicity of Anabaena aphanizomenoides (Cyanobacteria) water-blooms in the shallow brackish Oued Mellah lake (Morocco). Aquat. Ecosyst. Health 2005, 8, 95–104. [Google Scholar] [CrossRef]
- Ledreux, A.; Thomazeau, S.; Catherine, A.; Duval, C.; Yéprémian, C.; Marie, A.; Bernard, C. Evidence for saxitoxins production by the cyanobacterium Aphanizomenon gracile in a French recreational water body. Harmful Algae 2010, 10, 88–97. [Google Scholar] [CrossRef]
- Kokociński, M.; Akçaalan, R.; Salmaso, N.; Stoyneva-Gärtner, M.P.; Sukenik, A. Expansion of Alien and Invasive Cyanobacteria. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; Wiley: Hoboken, NJ, USA, 2017; pp. 28–39. ISBN 978-1-119-06868-6. [Google Scholar]
- Kasperovičienė, J.; Koreivienė, J.; Karosienė, J. Recent appearance of Raphidiopsis mediterranea (Cyanobacteria) in eutrophic Lithuanian lakes. In Biological Invasions in a Changing Word from Science to Management. Neobiota Book of Abstracts, 6th NEOBIOTA Conference, Copenhagen, Denmark, 14–17 September 2010; Kollmann, J., van Mölken, T., Ravn, H.P., Eds.; University of Copenhagen: Copenhagen, Denmark, 2010; p. 100. ISBN 978-87-993976-0-0. [Google Scholar]
- Koreivienė, J.; Kasperovičienė, J. Alien cyanobacteria Anabaena bergii var. limnetica Couté et Preisig from Lithuania: Some aspects of taxonomy, ecology and distribution. Limnologica 2011, 41, 325–333. [Google Scholar] [CrossRef]
- Karosienė, J.; Savadova, K.; Toruńska-Sitarz, A.; Koreivienė, J.; Kasperovičienė, J.; Vitonytė, I.; Błaszczyk, A.; Mazur-Marzec, H. Production of saxitoxins and anatoxin-a by cyanobacteria from Lithuanian lakes. Harmful Algae 2018. under review. [Google Scholar]
- Kavaliauskienė, J. Algae of Lithuanian Lakes; Institute of Geography: Vilnius, Lithuania, 1996; 148p, ISBN 9986-9097-0-8. [Google Scholar]
- Kokociński, M.; Gągała, I.; Jasser, I.; Karosienė, J.; Kasperovičienė, J.; Kobos, J.; Koreivienė, J.; Soininen, J.; Szczurowska, A.; Woszczyk, M.; et al. Distribution of invasive Cylindrospermopsis raciborskii in the East-Central Europe is driven by climatic and local environmental variables. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed]
- Budzyńska, A.; Gołdyn, R. Domination of invasive Nostocales (Cyanoprokaryota) at 52°N latitude. Phycol. Res. 2017, 65, 322–332. [Google Scholar] [CrossRef]
- Toporowska, M.; Pawlik-Skowrońska, B.; Krupa, D.; Kornijów, R. Winter versus summer blooming of phytoplankton in a shallow lake: Effect of hypertrophic conditions. Pol. J. Ecol. 2010, 58, 3–12. [Google Scholar]
- Walls, J.T.; Wyatt, K.H.; Doll, J.C.; Rubenstein, E.M.; Rober, A.R. Hot and toxic: Temperature regulates microcystin release from cyanobacteria. Sci. Total Environ. 2018, 610–611, 786–795. [Google Scholar] [CrossRef] [PubMed]
- Da Gomes, A.M.; Azevedo, S.M.F.O.; Lürling, M. Temperature effect on exploitation and interference competition among Microcystis aeruginosa, Planktothrix agardhii and Cyclotella meneghiniana. Sci. World J. 2015, 2015, 834197. [Google Scholar] [CrossRef] [PubMed]
- Mehnert, G.; Leunert, F.; Cirés, S.; Jöhnk, K.D.; Rücker, J.; Nixdorf, B.; Wiedner, C. Competitiveness of invasive and native cyanobacteria from temperate freshwaters under various light and temperature conditions. J. Plankton Res. 2010, 32, 1009–1021. [Google Scholar] [CrossRef] [Green Version]
- Budzyńska, A.; Rosińska, J.; Pełechata, A.; Toporowska, M.; Napiórkowska-Krzebietke, A.; Kozak, A.; Messyasz, B.; Pęcuła, W.; Kokociński, M.; Szeląg-Wasielewska, E.; et al. Environmental factors driving the occurrence of the invasive cyanobacterium Sphaerospermopsis aphanizomenoides (Nostocales) in temperate lakes. Sci. Total Environ. 2019, 650, 1338–1347. [Google Scholar] [CrossRef] [PubMed]
- Stüken, A.; Rücker, J.; Endrulat, T.; Preussel, K.; Hemm, M.; Nixdorf, B.; Karsten, U.; Wiedner, C. Distribution of three alien cyanobacterial species (Nostocales) in northeast Germany: Cylindrospermopsis raciborskii, Anabaena bergii and Aphanizomenon aphanizomenoides. Phycologia 2006, 45, 696–703. [Google Scholar] [CrossRef]
- Caraus, I. Algae of Romania. A Distributional Checklist of Actual Algae; Studii şi Cercetări, Biologie; University of Bacau: Bacau, Romania, 2002; Volume 7, pp. 1–809. [Google Scholar]
- Whitton, B.A.; John, D.M.; Kelly, M.G.; Haworth, E.Y. A Coded List of Freshwater Algae of the British Isles, 2nd ed.; Natural History Museum: London, UK, 2003; Available online: http://www.nhm.ac.uk/our-science/data/uk-species/checklists/NHMSYS0000591449/index.html (accessed on 30 August 2018).
- De Figueiredo, D.R.; Alves, A.; Pereira, M.J.; Correia, A. Molecular characterization of bloom-forming Aphanizomenon strains isolated from Vela Lake (Western Central Portugal). J. Plankton. Res. 2010, 32, 239–252. [Google Scholar] [CrossRef]
- Kaštovský, J.; Hauer, T.; Mareš, J.; Krautová, M.; Bešta, T.; Komárek, J.; Desortová, B.; Heteša, J.; Hindáková, A.; Houk, V.; et al. A review of the alien and expansive species of freshwater cyanobacteria and algae in the Czech Republic. Biol. Invasions 2010, 12, 3599–3625. [Google Scholar] [CrossRef]
- Pulina, S.; Padedda, B.M.; Sechi, N.; Lugliè, A. The dominance of cyanobacteria in Mediterranean hypereutrophic lagoons: A case study of Cabras Lagoon (Sardinia, Italy). Sci. Mar. 2011, 75, 111–120. [Google Scholar] [CrossRef]
- Wörmer, L.; Cirés, S.; Agha, R.; Verdugo, M.; de Hoyos, C.; Quesada, A. First detection of cyanobacterial PSP (paralytic shellfish poisoning) toxins in Spanish freshwaters. Toxicon 2011, 57, 918–921. [Google Scholar] [CrossRef] [PubMed]
- Karadžić, V.; Subakov Simić, G.; Natić, D.; Ržaničanin, A.; Ćirić, M.; Gačić, Z. Changes in the phytoplankton community and dominance of Cylindrospermopsis raciborskii (Wolosz.) Subba Raju in a temperate lowland river (Ponjavica, Serbia). Hydrobiologia 2013, 711, 43–60. [Google Scholar] [CrossRef]
- Lürling, M.; van Oosterhout, F.; Faassen, E. Eutrophication and Warming Boost Cyanobacterial Biomass and Microcystins. Toxins 2017, 9, 64. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.; Dao, T.S.; Vo, T.G.; Lürling, M. Warming affects growth rates and microcystin production in tropical bloom-forming Microcystis strains. Toxins 2018, 10, 123. [Google Scholar] [CrossRef] [PubMed]
- Gianuzzi, L.; Krock, B.; Crettaz Minaglia, M.C.; Rosso, L.; Houghton, C.; Sedan, D.; Malanga, G.; Espinosa, M.; Andrinolo, D.; Hernando, M. Growth, toxin production, active oxygen species and catalase activity of Microcystis aeruginosa (Cyanophyceae) exposed to temperature stress. Comp. Biochem. Physiol. Part C 2016, 189, 22–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziallas, C.; Grossart, H.P. Increasing Oxygen Radicals and Water Temperature Select for Toxic Microcystis sp. PLoS ONE 2011, 6, e25569. [Google Scholar] [CrossRef] [PubMed]
- Dias, E.; Pereira, P.; Franca, S. Production of paralytic shellfish toxins by Aphanizomenon sp. LMECYA 31 (cyanobacteria). J. Phycol. 2002, 38, 705–712. [Google Scholar] [CrossRef]
- Casero, M.C.; Ballot, A.; Agha, R.; Quesada, A.; Cirés, S. Characterization of saxitoxin production and release and phylogeny of sxt genes in paralytic shellfish poisoning toxin-producing Aphanizomenon gracile. Harmful Algae 2014, 37, 28–37. [Google Scholar] [CrossRef]
- Neilan, B.A.; Pearson, L.A.; Muenchhoff, J.; Moffitt, M.C.; Dittmann, E. Environmental conditions that influence toxin biosynthesis in cyanobacteria. Environ. Microbiol. 2013, 15, 1239–1253. [Google Scholar] [CrossRef] [PubMed]
- Rzymski, P.; Poniedziałek, B. The surprising world of cyanobacteria: Cylindrospermopsin has a soil face. J. Phycol. 2015, 51, 1037–1039. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Kaya, K. Oscillapeptin G, a tyrosinase inhibitor from toxic Oscillatoria agardhii. J. Nat. Prod. 1996, 59, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Ersmark, K.; del Valle, J.R.; Hanessian, S. Chemistry and biology of the aeruginosin family of serine protease inhibitors. Angew. Chem. Int. Ed. 2008, 47, 1202–1223. [Google Scholar] [CrossRef] [PubMed]
- Itou, Y.; Suzuki, S.; Ishida, K.; Murakami, M. Anabaenopeptins G and H, potent carboxypeptidase A inhibitors from the cyanobacterium Oscillatoria agardhii (NIES-595). Bioorg. Med. Chem. Lett. 1999, 9, 1243–1246. [Google Scholar] [CrossRef]
- Spoof, L.; Blaszczyk, A.; Meriluoto, J.; Ceglowska, M.; Mazur-Marzec, H. Structures and activity of new anabaenopeptins produced by Baltic Sea cyanobacteria. Mar. Drugs 2016, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Lebret, K.; Kritzberg, S.; Figueroa, R.; Rengefors, K. Genetic diversity within and genetic differentiation between blooms of a microalgal species. Environ. Microbiol. 2012, 14, 2395–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 2. Teil: Oscillatoriales. In Süßwasserflora von Mitteleuropa; Bridel, B., Gastner, G., Krienitz, L., Schargerl, M., Eds.; Elsevier: Ámsterdam, The Netherlands, 2005; Volume 19, 759p, ISBN 3-8274-0919-5. [Google Scholar]
- Torres, C.; Lürling, M.; Marinho, M.M. Assessment of the Effects of Light vailability on Growth and Competition Between Strains of Planktothrix agardhii and Microcystis aeruginosa. Microb. Ecol. 2016, 71, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Andersen, R.A. (Ed.) Algal Culturing Techniques; Academic Press: Cambridge, MA, USA, 2005; 596p. [Google Scholar]
- Guillard, R.R.L. Division rates. In Handbook of Phycological Methods: Culture Methods and Growth Measurements; Stein, J.R., Ed.; Cambridge University Press: London, UK, 1973; pp. 289–311. [Google Scholar]
- Grabowska, M.; Mazur-Marzec, H. The effect of cyanobacterial blooms in the Siemianówka Dam Reservoir on the phytoplankton structure in the Narew River. Oceanol. Hydrobiol. Stud. 2011, 40, 19–26. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response Variable | Factor | Degrees of Freedom | Mean Square | F Value | p Value |
|---|---|---|---|---|---|
| Growth rate | Temperature | 6 | 0.258 | 15.83 | <0.001 |
| Taxa | 3 | 0.312 | 15.81 | <0.001 | |
| Temperature × Taxa | 18 | 0.047 | 7.79 | <0.001 | |
| Species origin 1 | 1 | 0.003 | 0.14 | 0.711 | |
| Temperature × Species origin 1 | 6 | 0.082 | 5.89 | <0.001 | |
| Toxic strain 2 | 1 | 0.100 | 40.98 | <0.001 | |
| Temperature × Toxic strain 2 | 6 | 0.001 | 0.76 | 0.606 |
| Species | Strain | Lake and Year of Isolation of the Strain | Cyanotoxins | NRPs | |
|---|---|---|---|---|---|
| Native | P. agardhii | NRC_SIR/F5-09 | Širvys, 2014 | MCs | + |
| NRC_JIE/E9-07 | Jieznas, 2015 | - | + | ||
| A. gracile | NRC_SIR/B41-09 | Širvys, 2015 | STX | - | |
| NRC_SIR/C10-07 | Širvys, 2015 | - | - | ||
| Alien in Europe | S. aphanizomenoides | NRC_JIE/G11-07 | Jieznas, 2015 | - | - |
| NRC_JIE/F11-07 | Jieznas, 2015 | - | - | ||
| C. bergii | NRC_REK/D2-08 | Rėkyva, 2015 | - | - | |
| NRC_GIN/B6-08 | Gineitiškės, 2017 | - | - | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savadova, K.; Mazur-Marzec, H.; Karosienė, J.; Kasperovičienė, J.; Vitonytė, I.; Toruńska-Sitarz, A.; Koreivienė, J. Effect of Increased Temperature on Native and Alien Nuisance Cyanobacteria from Temperate Lakes: An Experimental Approach. Toxins 2018, 10, 445. https://doi.org/10.3390/toxins10110445
Savadova K, Mazur-Marzec H, Karosienė J, Kasperovičienė J, Vitonytė I, Toruńska-Sitarz A, Koreivienė J. Effect of Increased Temperature on Native and Alien Nuisance Cyanobacteria from Temperate Lakes: An Experimental Approach. Toxins. 2018; 10(11):445. https://doi.org/10.3390/toxins10110445
Chicago/Turabian StyleSavadova, Ksenija, Hanna Mazur-Marzec, Jūratė Karosienė, Jūratė Kasperovičienė, Irma Vitonytė, Anna Toruńska-Sitarz, and Judita Koreivienė. 2018. "Effect of Increased Temperature on Native and Alien Nuisance Cyanobacteria from Temperate Lakes: An Experimental Approach" Toxins 10, no. 11: 445. https://doi.org/10.3390/toxins10110445