Tb II-I, a Fraction Isolated from Tityus bahiensis Scorpion Venom, Alters Cytokines’: Level and Induces Seizures When Intrahippocampally Injected in Rats


 ,
, 
 and
and
Abstract
:1. Introduction
2. Results
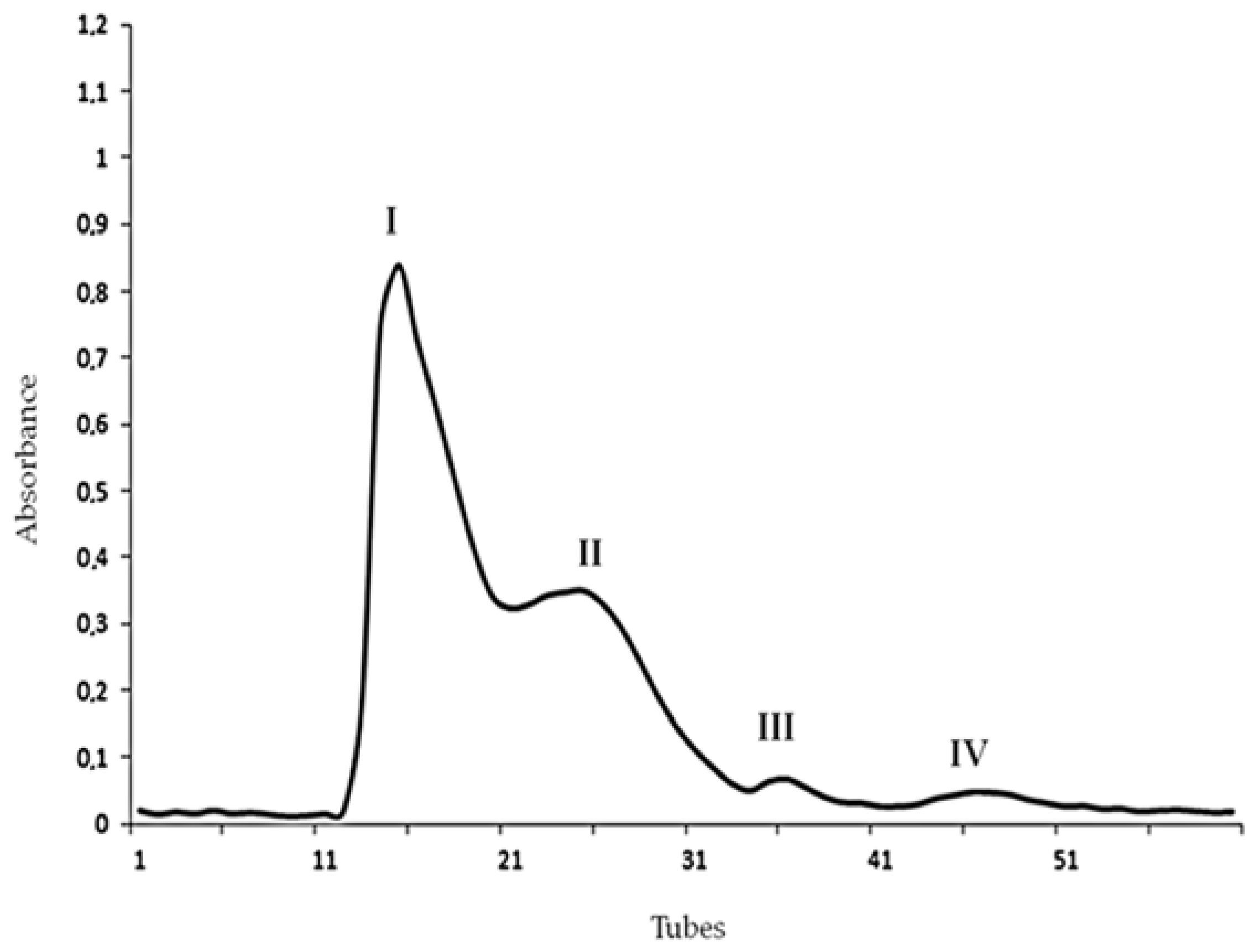
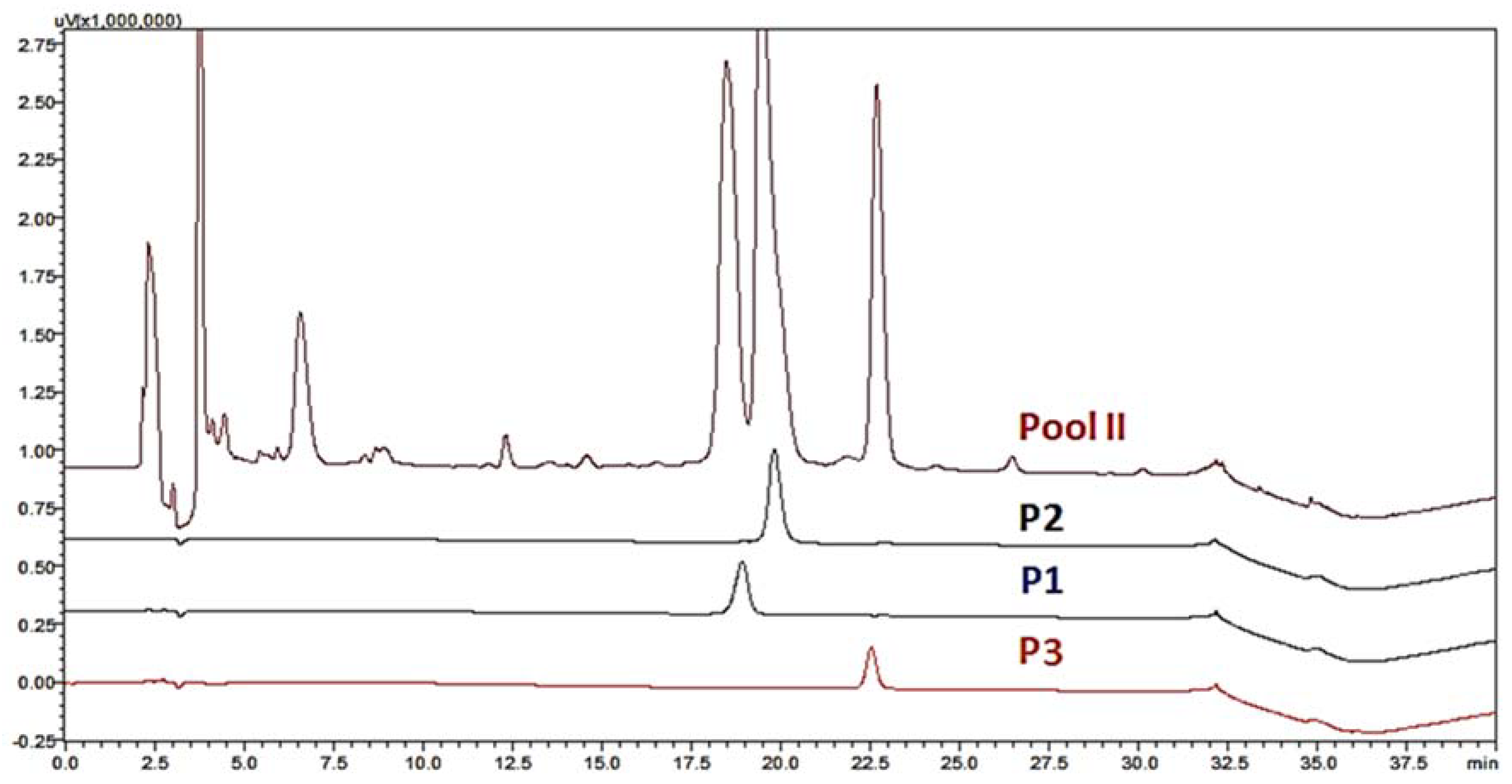
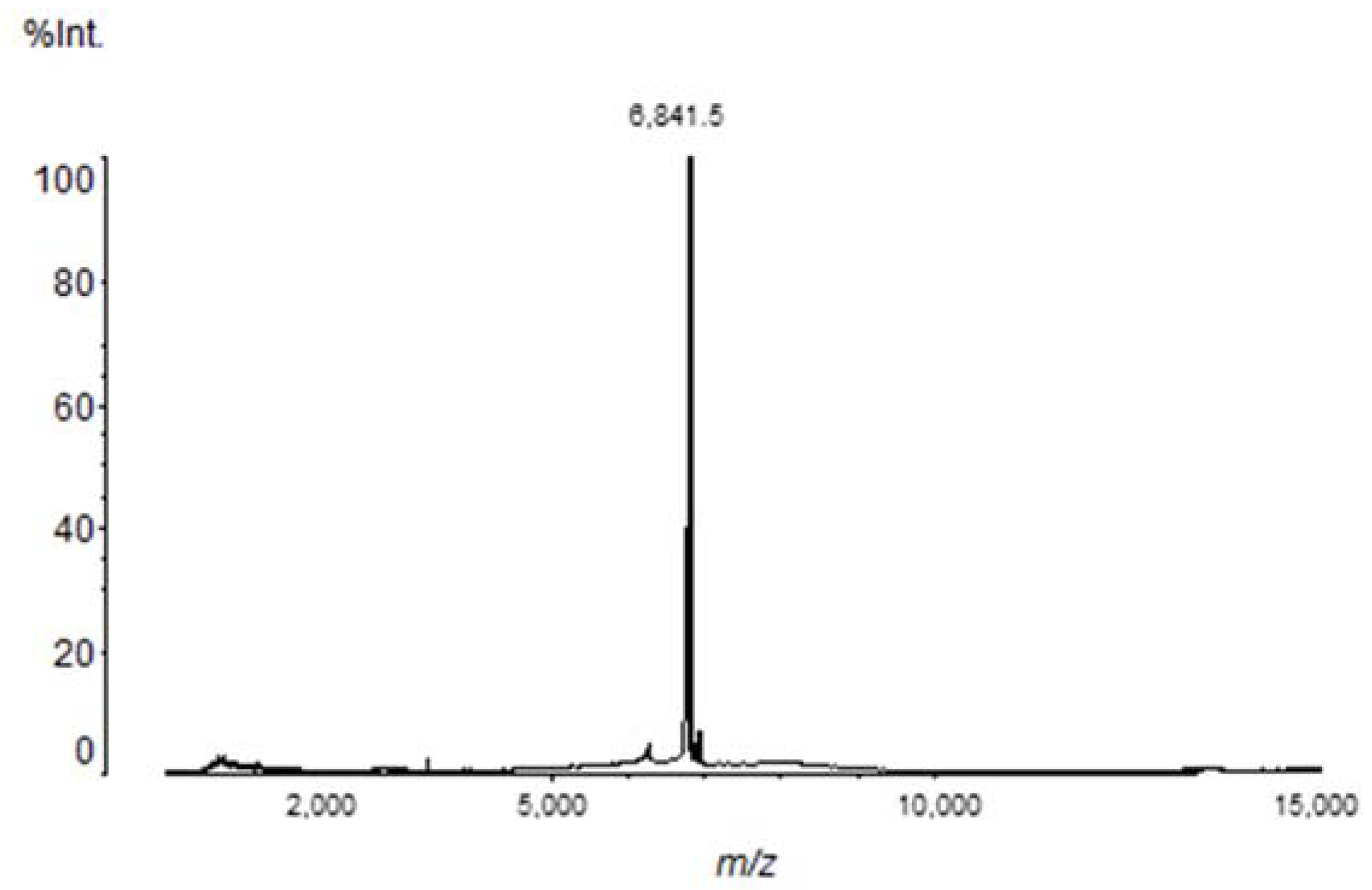
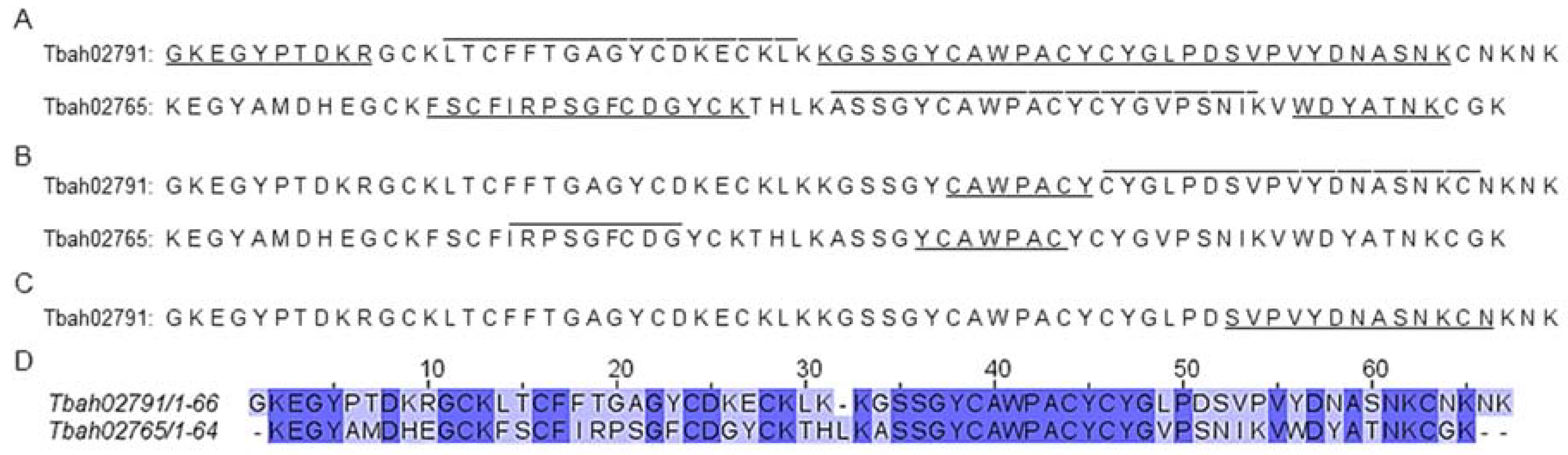
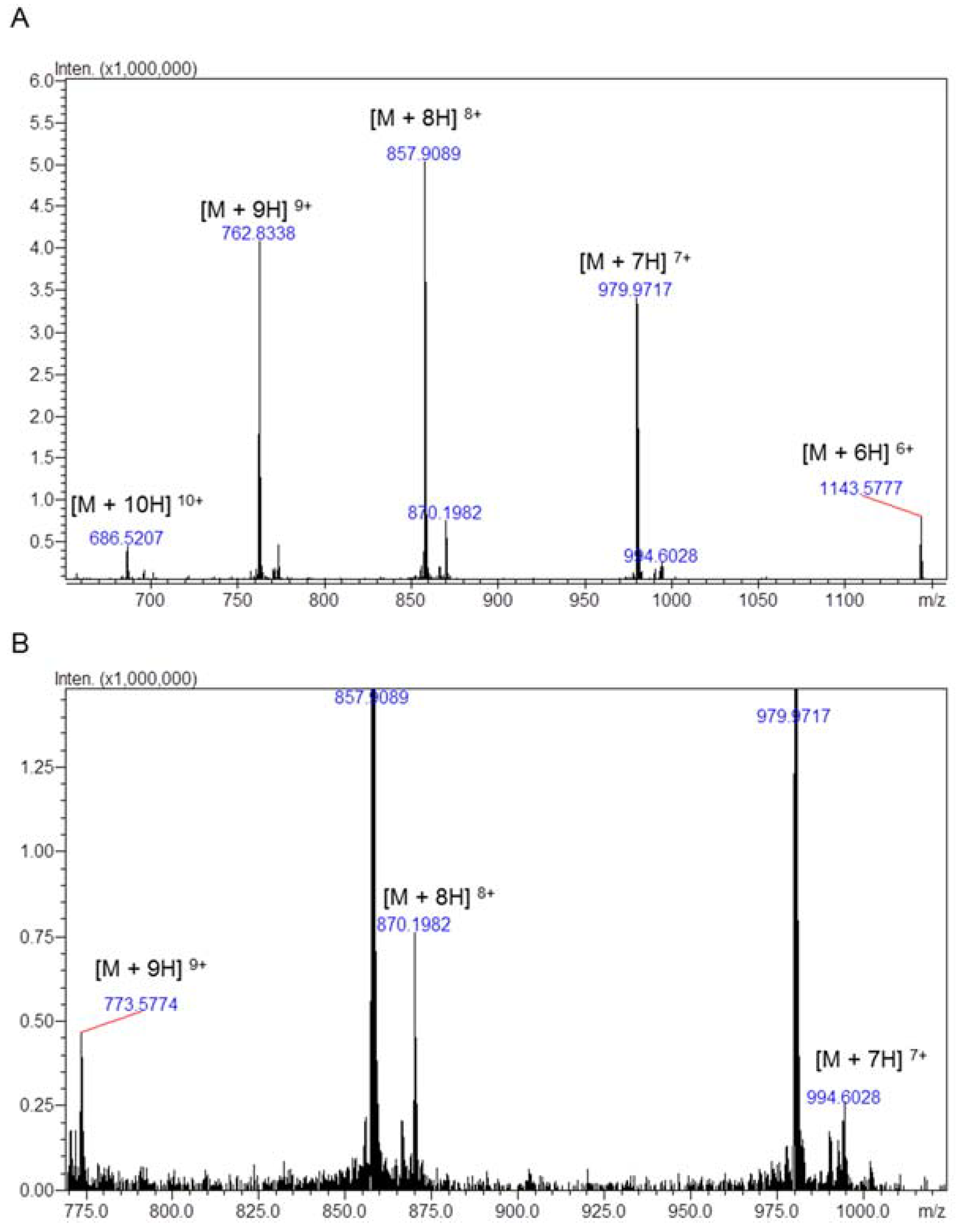
2.1. Chromatography, Biochemical Characterization and Mass Spectrometry Analysis of Tb II-I
2.2. Electroencephalographic Recording and Behavioral Observation
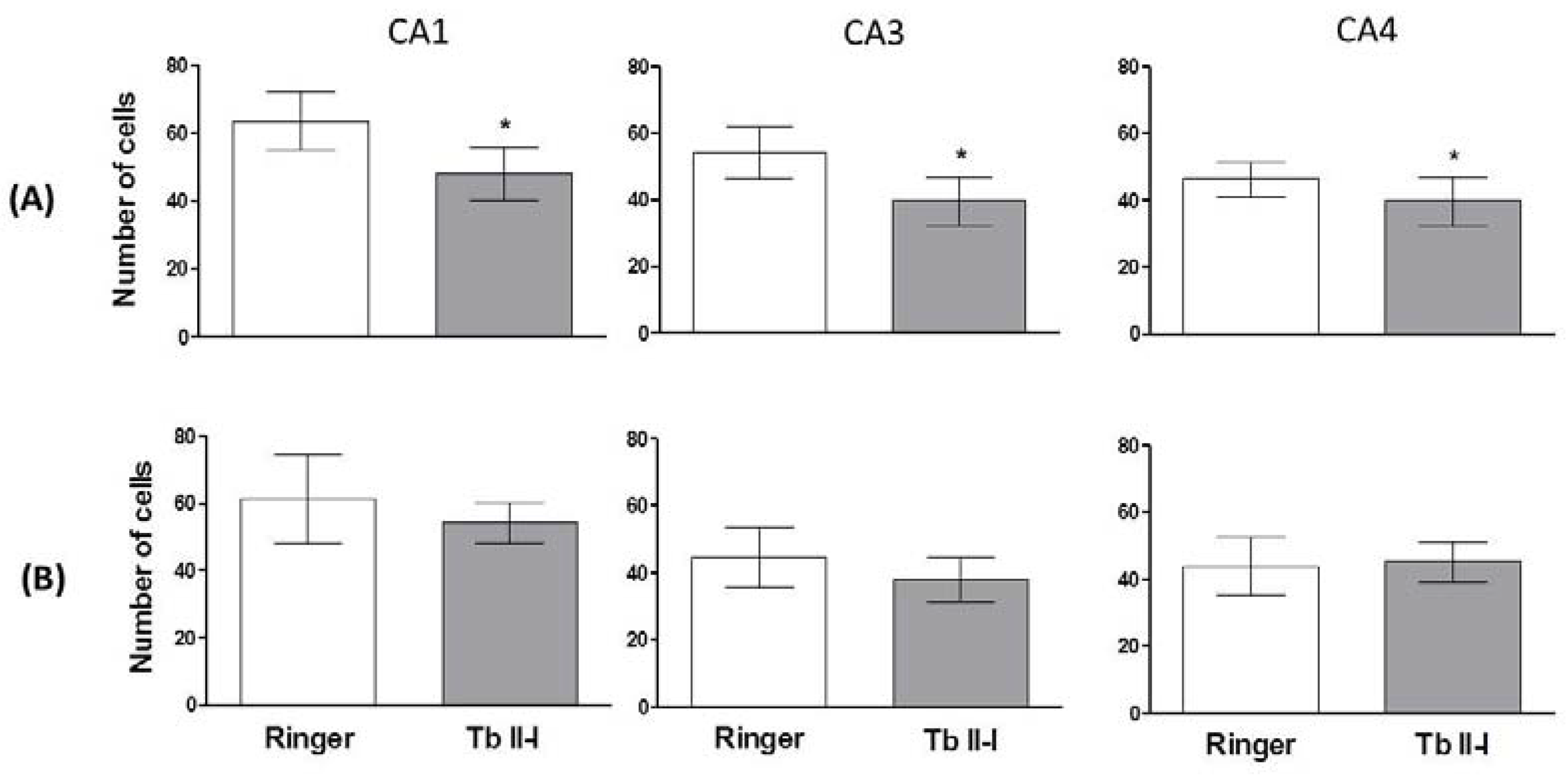
2.3. Histopathological Analysis
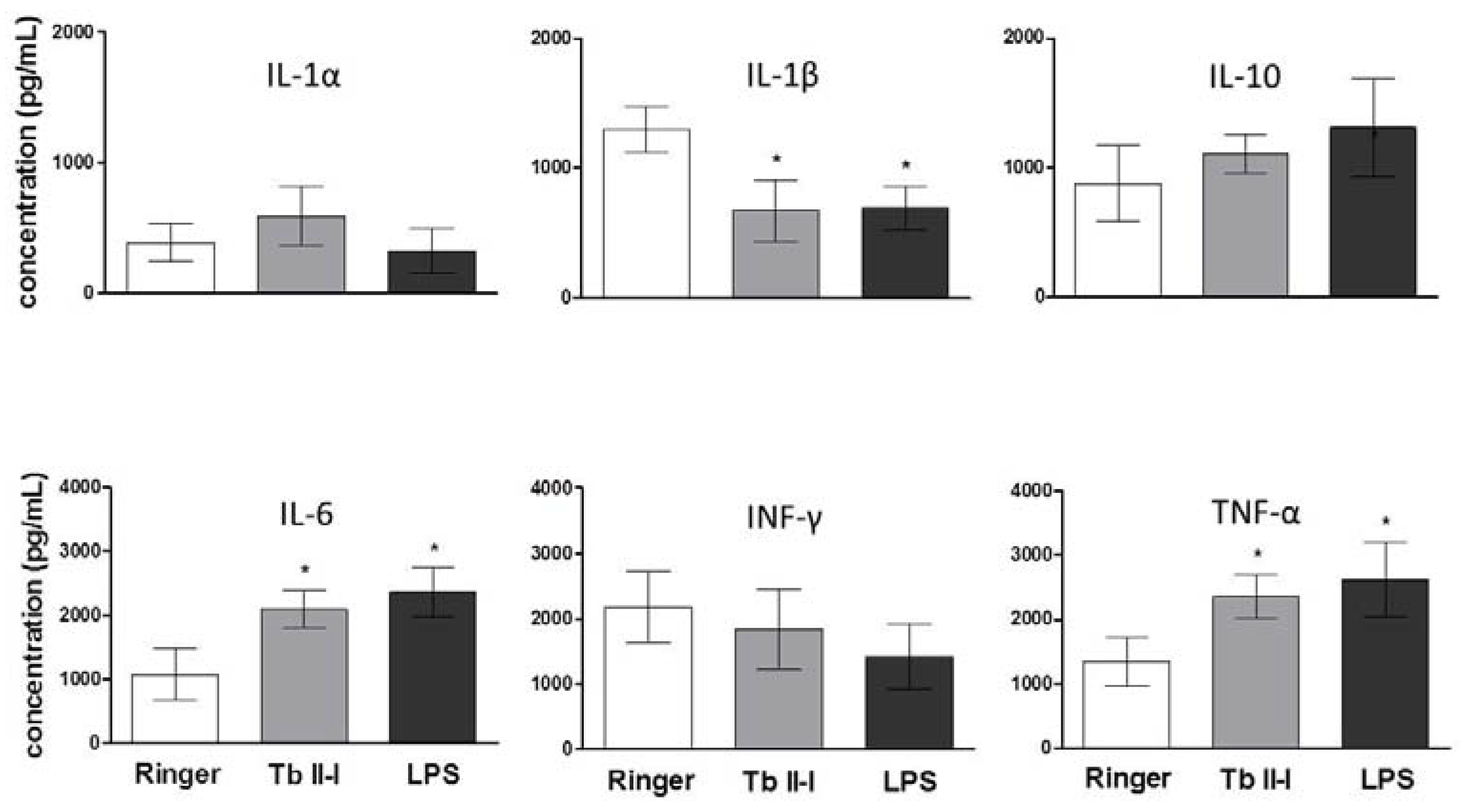
2.4. Cytokine Levels
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Venom Fractionation
5.2. Mass Spectrometry
5.3. Amino Acid Sequencing
5.4. In-Solution Digestion and Proteomic Analysis
5.5. Animals
5.6. Stereotaxic Surgery and Electrographic Recording
5.7. Histological Analysis
5.8. Cytokine Quantification
6. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Cologna, C.T.; Marcussi, S.; Giglio, J.R.; Soares, A.M.; Arantes, E.A. Tityus serrulatus scorpion venom and toxins: An overview. Protein Pept. Lett. 2009, 16, 920–932. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, E.; Gurrola, G.B.; Schwartz, E.F.; Possani, L.D. Scorpion venom components as potential candidates for drug development. Toxicon 2015, 93, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Pucca, M.B.; Cerni, F.A.; Pinheiro Junior, E.L.; Bordon, K.C.F.; Amorim, F.G.; Cordeiro, F.A.; Longhim, H.T.; Cremonez, C.M.; Oliveira, G.H.; Arantes, E.A. Tityus serrulatus venom—A lethal cocktail. Toxicon 2015, 108, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Arantes, E.C. Arachnids of medical importance in Brazil: Main active compounds present in scorpion and spider venoms and tick saliva. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 24. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M. The scorpion envenoming syndrome. Toxicon 1995, 33, 825–858. [Google Scholar] [CrossRef]
- Possani, L.D.; Becerril, B.; Delpierre, M.; Tytgat, J. Scorpion toxins specific for sodium channels. Eur. J. Biochem. 1999, 264, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Petricevich, V.L.; Lebrun, I. Immunomodulatory effects of the Tityus serrulatus venom on murine macrophage functions in vitro. Med. Inflamm. 2005, 24, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Meki, A.R.; Monhey el-Dean, Z.M. Serum interleukin-1 beta, interleukin-6, nitric oxide and alpha-antitrypsin in scorpion envenomed children. Toxicon 1998, 36, 1851–1859. [Google Scholar] [CrossRef]
- Petricevich, V.L.; Peña, C.F. The dynamics of cytokine and nitric oxide secretion in mice injected with Tityus serrulatus scorpion venom. Med. Inflamm. 2002, 11, 173–180. [Google Scholar] [CrossRef] [PubMed]
- D’Suze, G.; Moncada, S.; Gonzalez, C.; Sevcik, C.; Aguilar, V.; Alagon, A. Relationship between plasmatic levels of various cytokines, tumour necrosis factor, enzymes, glucose and venom concentration following Tityus scorpion sting. Toxicon 2003, 41, 367–375. [Google Scholar] [CrossRef]
- Fukuhara, Y.D.M.; Reis, M.L.; Dellalibera-Joviliano, R.; Cunha, F.Q.C.; Donadi, E.A. Increased plasma levels of IL-1a, IL-6, IL-8, IL-10 and TNF-a in patients moderately or severely envenomed by Tityus serrulatus scorpion sting. Toxicon 2003, 41, 49–55. [Google Scholar] [CrossRef]
- Petricevich, V.L. Scorpion venom and the inflammatory response. Med. Inflamm. 2010, 16. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.V.; Dorce, V.A.C.; Freitas, L.A.; Dorce, A.L.C.; Lebrun, I.; Sobral, A.C.M.; Portaro, F.C.V.; Kuniyoshi, A.K.; Carvalho, D.C.; Nencioni, A.L.A. Intrahippocampal injection of TsTX-I increases the levels of INF-γ in the cerebral tissue but not the levels of glutamate. Toxicon 2015, 103, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Van Fraga, I.T.; Limborco-Filho, M.; Lima, O.C.O.; Lacerda-Queiroz, N.; Guidine, P.A.M.; Moraes, M.F.D.; Araújo, R.N.; Moraes-Santos, T.; Massensini, A.R.; Arantes, R.M.E.; et al. Effects of tityustoxin on cerebral inflammatory response in young rats. Neurosci. Lett. 2015, 588, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Taibi-Djennah, Z.; Matin-Eauclaire, M.F.; Laraba-Djebar, F. Systemic responses following brain injuries and inflammatory process activation induced by a neurotoxin of Androctonus scorpion venom. Neuroimmunomodulation 2015, 22, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Vitkovic, L.; Bockaert, J.; Jacque, C. “Inflammatory” cytokines: Neuromodulators in normal brain? J. Neurochem. 2000, 74, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, A.; Friedman, A.; Dingledine, R.J. The role of inflammation in epileptogenesis. Neuropharmacology 2013, 69, 16–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravizza, T.; Balosso, S.; Vezzani, A. Inflammation and prevention of epileptogenesis. Neurosci. Lett. 2011, 497, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Bonilha, L.; Cendes, F.; Ghizoni, E.; Vieira, R.J.; Li, L.M. Epilepsy due to a destructive brain lesion caused by a scorpion sting. Arch. Neurol. 2004, 61, 1294–1296. [Google Scholar] [CrossRef] [PubMed]
- Souza, D.G.; Tanaka, K.; Algemiro, W.; Dezena, R.A.; Borges, M.M.; Pereira, C.U.; Lima, A.A.; Feitosa, S.C. Hemorrhagic stroke following scorpion sting. A case report. Rev. Chil. Neurocir. 2013, 39, 69–70. [Google Scholar]
- Nencioni, A.L.A.; Beraldo Neto, E.; Freitas, L.A.; Dorce, V.A.C. Effects of Brazilian scorpion venoms on the central nervous system. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 3. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P.; Goyffon, M. Epidemiology of scorpionism: A global appraisal. Acta Trop. 2008, 107, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Lothman, E.W.; Bertram, E.H.; Stinger, J.L. Functional anatomy of hippocampal seizures. Prog. Neurobiol. 1991, 37, 1–82. [Google Scholar] [CrossRef]
- Pimenta, A.M.; Martin-Euclaire, M.F.; Rochat, H.; Figueiredo, S.G.; Kalapothakis, E.; Afonso, L.C.; De Lima, M.E. Purification, amino-acid sequence and partial characterization of two toxins with anti-insect activity from the venom of the South-American scorpion Tityus bahiensis (Buthidae). Toxicon 2001, 39, 1009–1019. [Google Scholar] [CrossRef]
- Becerril, B.; Corona, M.; Coronas, F.I.V.; Zamudio, F.; Calderon-Aranda, E.; Fletcher, P.; Martin, B.M.; Possani, L.D. Toxic peptides and genes encoding toxin γ of the Brazilian scorpions Tityus bahiensis and Tityus stigmurus. Biochem. J. 1996, 313, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.F.; Nencioni, A.L.A.; Lebrun, I.; Sandoval, M.R.L.; Dorce, V.A.C. Behavioral, electroencephalographic, and histopathologic effects of a neuropeptide isolated from Tityus serrulatus scorpion venom in rats. Pharmacol. Biochem. Behav. 1998, 60, 7–14. [Google Scholar] [CrossRef]
- Teixeira, V.F.; Conceição, I.M.; Lebrun, I.; Nencioni, A.L.A.; Dorce, V.A.C. Intrahippocampal injection of TsTX-I, a beta-scorpion toxin, causes alterations in electroencephalographic recording and behavior in rats. Life Sci. 2010, 87, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Ossanai, L.T.T.; Lourenço, G.A.; Nencioni, A.L.A.; Lebrun, I.; Yamanouye, N.; Dorce, V.A.C. Effects of a toxin isolated from Tityus bahiensis scorpion venom on the hippocampus of rats. Life Sci. 2012, 91, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Nencioni, A.L.A.; Lebrun, I.; Dorve, V.A.C. A microdialysis study of glutamate concentration in the hippocampus of rats after TsTX toxin injection and blockade of toxin effects by glutamate receptor antagonists. Pharmacol. Biochem. Behav. 2003, 74, 455–463. [Google Scholar] [CrossRef]
- Vezzani, A.; Moneta, D.; Richichi, C.; Aliprandi, M.; Burrows, S.J.; Ravizza, T.; Perego, C.; De Simoni, M.G. Functional role of inflammatory cytokines and anti-inflammatory molecules in seizures and epileptogenesis. Epilepsia 2002, 43, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, A.; Friedman, A. Brain inflammation as a biomarker in epilepsy. Biomark. Med. 2011, 5, 607–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vezzani, A.; Viviani, B. Neuromodulatory properties of inflammatory cytokines and their impact on neuronal excitability. Neuropharmacology 2015, 96, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Fialho, E.M.; Maciel, M.C.; Silva, A.C.; Reis, A.S.; Assunção, A.K.; Fortes, T.S.; Silva, L.A.; Guerra, R.N.; Kwasniewski, F.H.; Nascimento, F.R. Immune cells recruitment and activation by Tityus serrulatus scorpion venom. Toxicon 2011, 58, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Petricevich, V.L.; Cruz, A.H.; Coronas, F.I.V.; Possani, L.D. Toxin gamma from Tityus serrulatus scorpion venom plays an essential role in immunomodulation of macrophages. Toxicon 2007, 50, 666–675. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, U.C.; Candido, D.M.; Dorce, V.A.; Junqueira-de-Azevedo, I.D.L.M. The transcriptome recipe for the venom cocktail of Tityus bahiensis scorpion. Toxicon 2015, 95, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Marin, I.; Kipnis, J. Learning and memory…and the immune system. Learn. Mem. 2013, 20, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.H.; Haroon, E.; Raison, C.L.; Felger, J.C. Cytokine targets in the brain: Impact on neurotransmitters and neurocircuits. Depression Anxiety 2013, 30, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Devinsky, O.; Vezzani, A.; Najjar, S.; De Lanerolle, N.C.; Rogawski, M.A. Glia and epilepsy: Excitability and inflammation. Trends Neurosci. 2013, 36, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Bauer, S.; Nowak, M.; Norwood, B.; Tackenberg, B.; Rosenow, F.; Knake, S.; Oertel, W.H.; Hamer, H.M. Cytokines and epilepsy. Seizure 2011, 20, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Ambrosini, A.; Louin, G.; Croci, N.; Plotkine, M.; Jafarian-Tehrani, M. Characterization of a rat model to study acute neuroinflammation on histopathological, biochemical and functional outcomes. J. Neurosci. Methods 2005, 144, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, X.; Nai, Y.; Ba, M.; Fu, Q.; Wu, F. Hippocampal infusion of lipopolysaccharide induces immune responses and results in seizures in rats. NeuroReport 2017, 28, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Leonoudakis, D.; Braithwaite, S.P.; Beattie, M.S.; Beattie, E.C. TNFα-induced AMPA-receptors trafficking in CNS neurons; relevance to excitotoxicity? Neuron Glia Biol. 2004, 1, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Olmos, G.; Lladó, J. Tumor necrosis factor alpha: A link between neuroinflammation and excitotoxicity. Med. Inflamm. 2014, 2014, 861231. [Google Scholar] [CrossRef] [PubMed]
- Beattie, E.C.; Stellwagen, D.; Morishita, W.; Bresnahan, J.C.; Ha, B.K.; Von Zastrow, M.; Beattie, M.S.; Malenka, R.C. Control of synaptic strength by glial TNF-α. Science 2002, 295, 2282–2285. [Google Scholar] [CrossRef] [PubMed]
- Stellwagen, D.; Beattie, E.C.; Seo, J.Y.; Malenka, R.C. Differential regulation of AMPA receptor and GABA receptor trafficking by Tumor Necrosis Factor-α. J. Neurosci. 2005, 25, 3219–3228. [Google Scholar] [CrossRef] [PubMed]
- Stuck, E.D.; Christensen, R.N.; Huie, J.R.; Tovar, C.A.; Miller, B.A.; Nout, Y.S.; Bresnahan, J.C.; Beattie, M.S.; Ferguson, A.R. Tumor necrosis Factor Alpha mediates GABAA receptor trafficking to the plasma membrane of spinal cord neurons in vivo. Neural Plast. 2012, 2012, 261345. [Google Scholar] [CrossRef] [PubMed]
- Pickering, M.; Cumiskey, D.; O’Connor, J.J. Actions of TNF-α on glutamatergic synaptic transmission in the central nervous system. Exp. Physiol. 2005, 90, 663–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, D.C.; Wallis, G.; Dahle, E.J.; McElroy, P.B.; Thomson, K.E.; Tesi, R.J.; Szymkowski, D.E.; West, P.J.; Smeal, R.M.; Patel, M.; et al. Hippocampal TNFα signaling contributes to seizure generation in an infection-induced mouse model of limbic epilepsy. eNeuro 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Gruol, D.L. IL-6 regulation of synaptic function in the CNS. Neuropharmacology 2015, 96, 42–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothaug, M.; Becker-Pauly, C.; Rose-John, S. The role of interleukin-6 signaling in nervous tissue. Biochim. Biophys. Acta 2016, 1863, 1218–1227. [Google Scholar] [CrossRef] [PubMed]
- Sallmann, S.; Juttler, E.; Prinz, S.; Petersen, N.; Knopf, U.; Weiser, T.; Schwaninger, M. Induction of interleukin-6 by depolarization of neurons. J. Neurosci. 2000, 20, 8637–8642. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.E.; Kim, G.S.; Chan, P.H. Neuroprotection by interleukin-6 mediated by signal transducer and activator of transcriptase 3 and antioxidative signaling in ischemic stroke. Stroke 2011, 42, 3574–3579. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Oscos, F.; Salgado, H.; Hall, S.; Thomas, F.; Farmer, G.E.; Bermeo, G.; Galindo, L.C.; Ramirez, R.D.; D’Mello, S.; Rose-John, S.; et al. The stress-induced cytokine interleukin-6 decreases the inhibition/excitation ratio in the rat temporal cortex via trans-signaling. Biol. Psychiatry 2012, 71, 574–582. [Google Scholar] [CrossRef] [PubMed]
- D’Arcangelo, G.; Tancredi, V.; Onofri, F.; D’Antuono, M.; Giovedi, S.; Benfenati, F. Interleukin-6 inhibits neurotransmitter release and the spread of excitation in the rat cerebral cortex. Eur. J. Neurosci. 2000, 12, 1241–1252. [Google Scholar] [CrossRef] [PubMed]
- Fogal, B.; Hewett, S.J. Interleukin 1-β: A bridge between inflammation and excitotoxicity? J. Neurochem. 2008, 106, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Yirmiya, R.; Goshen, I. Immune modulation of learning, memory, neural plasticity and neurogenesis. Brain Behav. Immun. 2011, 25, 181–213. [Google Scholar] [CrossRef] [PubMed]
- Viviani, B.; Corsini, E.; Binaglia, M.; Galli, C.L.; Marinovich, M. Reactive oxygen species generated by glia are responsible for neuron death induced by human immunodeficiency virus-glycoprotein 120 in vitro. Neuroscience 2001, 107, 51–58. [Google Scholar] [CrossRef]
- Vezzani, A.; Moneta, D.; Conti, M.; Richichi, C.; Ravizza, T.; De Luigi, A.; De Simoni, M.G.; Sperk, G.; Andell-Jonsson, S.; Lundkvist, J.; et al. Powerful anticonvulsant action of IL-1 receptor antagonist on intracerebral injection and astrocytic overexpression in mice. Proc. Natl. Acad. Sci. USA 2000, 97, 11534–11539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagan, P.; Poole, S.; Bristow, A.F.; Tilders, F.; Silverstein, F.S. Intracerebral NMDA injection stimulates production of interleukin-1 beta in perinatal rat brain. J. Neurochem. 1996, 67, 2215–2218. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, A.; Conti, M.; de Luigi, A.; Ravizza, T.; Moneta, D.; Marchesi, F.; de Simoni, M.G. Interleukin-1β immunoreactivity and microglia are enhanced in the rat hippocampus by focal kainate application: Functional evidence for enhancement of electrographic seizures. J. Neurosci. 1999, 19, 5054–5065. [Google Scholar] [CrossRef] [PubMed]
- Viviani, B.; Bartesaghi, S.; Gardoni, F.; Vezzani, A.; Behrens, M.M.; Bartfai, T.; Binaglia, M.; Corsini, E.; Di Luca, M.; Galli, C.L.; et al. Interleukin-1β enhances NMDA receptor-mediated intracellular calcium increase through activation of the Src family of kinases. J. Neurosci. 2003, 23, 8692–8700. [Google Scholar] [CrossRef] [PubMed]
- Ravizza, T.; Boer, K.; Redeker, S.; Spliet, W.G.M.; van Rijen, P.C.; Troost, D.; Vezzani, A.; Aronica, E. The IL-1β system in epilepsy-associated malformations of cortical development. Neurobiol. Dis. 2006, 24, 128–143. [Google Scholar] [CrossRef] [PubMed]
- Kołosowska, K.; Maciejak, P.; Szyndler, J.; Turzyńska, D.; Sobolewska, A.; Płaźnik, A. The role of interleukin-1β in the pentylenetetrazole-induced kindling of seizures, in the rat hippocampus. Eur. J. Pharmacol. 2014, 731, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Minami, M.; Kuraishi, Y.; Satoh, M. Effects of kainic acid on messenger RNA of IL-1β, IL-6, TNF-α and LIF in the rat brain. Biochem. Biophys. Res. Commun. 1991, 176, 593–598. [Google Scholar] [CrossRef]
- Edman, P.; Begg, G. A protein sequenator. Eur. J. Biochem. 1967, 1, 80–91. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, L.A.; Ferreira, R.S., Jr.; Barraviera, B.; de Carvalho, F.C.T.; de Barros, L.C.; Dos Santos, L.D.; Pimenta, D.C. Crotalus durissus terrificus crotapotin naturally displays preferred positions for amino acid substitutions. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 28, 23–46. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Zhang, K.; Hendrie, C.; Liang, C.; Li, M.; Doherty-Kirby, A.; Lajoie, G. PEAKS: Powerful software for peptide de novo sequencing by tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 2337–2342. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 4th ed.; Academic Press: San Diego, CA, USA, 1998; ISBN 0-12-547619-1. [Google Scholar]
- Nencioni, A.L.A.; Lourenço, G.A.; Lebrun, I.; Florio, J.C.; Dorce, V.A.C. Central effects of Tityus serrulatus and Tityus bahiensis scorpion venoms after intraperitoneal injection in rats. Neurosci. Lett. 2009, 463, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Aboumaâd, B.; Iba, N.; Dersi, N. L’envenimation scorpionique au Maroc: Scorpions du genre Androctonus, Buthus et Hottentota. Bull. Soc. Pathol. Exot. 2014, 107, 39–47. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alteration Observed | Fraction I n = 6 | Fraction II n = 6 | Fraction III n = 6 | Fraction IV n = 6 | Ringer n = 6 |
|---|---|---|---|---|---|
| Prostration | 33% | 33% | 17% | 17% | 0% |
| Respiratory difficulty | 17% | 50% | 0% | 0% | 0% |
| Locomotor difficulty | 0% | 50% | 0% | 0% | 0% |
| Salivary and lachrymal Secretions | 17% | 33% | 0% | 0% | 0% |
| Myoclonus | 0% | 83% | 0% | 0% | 0% |
| WDS | 33% | 83% * | 0% | 0% | 0% |
| Spikes | 33% | 83% * | 0% | 0% | 0% |
| Epileptiform discharges | 33% | 83% * | 0% | 0% | 0% |
| Toxin | Theoretical Mass | Experimental Mass |
|---|---|---|
| Tbah02791/Toxin Tb4 | 6851.04 | 6851.71 ± 0.91 |
| Tbah02765/Tb2 II | 6950.03 | 6950.30 ± 0.98 |
| Treatment | N | Salivary and Lachrymal Secretions | WDS | Penile Erection | Myoclonus | Slow Waves | Grouped Spikes | Strong Discharges |
|---|---|---|---|---|---|---|---|---|
| Tb II-I | 6 | 33% | 67% * | 50% | 33% | 33% | 67% * | 67% * |
| Ringer | 6 | 0% | 0% | 0% | 0% | 0% | 0% | 0% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beraldo Neto, E.; Mariano, D.O.C.; Freitas, L.A.; Dorce, A.L.C.; Martins, A.N.; Pimenta, D.C.; Portaro, F.C.V.; Cajado-Carvalho, D.; Dorce, V.A.C.; Nencioni, A.L.A. Tb II-I, a Fraction Isolated from Tityus bahiensis Scorpion Venom, Alters Cytokines’: Level and Induces Seizures When Intrahippocampally Injected in Rats. Toxins 2018, 10, 250. https://doi.org/10.3390/toxins10060250
Beraldo Neto E, Mariano DOC, Freitas LA, Dorce ALC, Martins AN, Pimenta DC, Portaro FCV, Cajado-Carvalho D, Dorce VAC, Nencioni ALA. Tb II-I, a Fraction Isolated from Tityus bahiensis Scorpion Venom, Alters Cytokines’: Level and Induces Seizures When Intrahippocampally Injected in Rats. Toxins. 2018; 10(6):250. https://doi.org/10.3390/toxins10060250
Chicago/Turabian StyleBeraldo Neto, Emidio, Douglas O. C. Mariano, Lucas A. Freitas, Ana L. C. Dorce, Adriana N. Martins, Daniel C. Pimenta, Fernanda C. V. Portaro, Daniela Cajado-Carvalho, Valquiria A. C. Dorce, and Ana L. A. Nencioni. 2018. "Tb II-I, a Fraction Isolated from Tityus bahiensis Scorpion Venom, Alters Cytokines’: Level and Induces Seizures When Intrahippocampally Injected in Rats" Toxins 10, no. 6: 250. https://doi.org/10.3390/toxins10060250