Subtype Specificity of β-Toxin Tf1a from Tityus fasciolatus in Voltage Gated Sodium Channels

 ,
, 
Abstract
:1. Introduction
2. Results
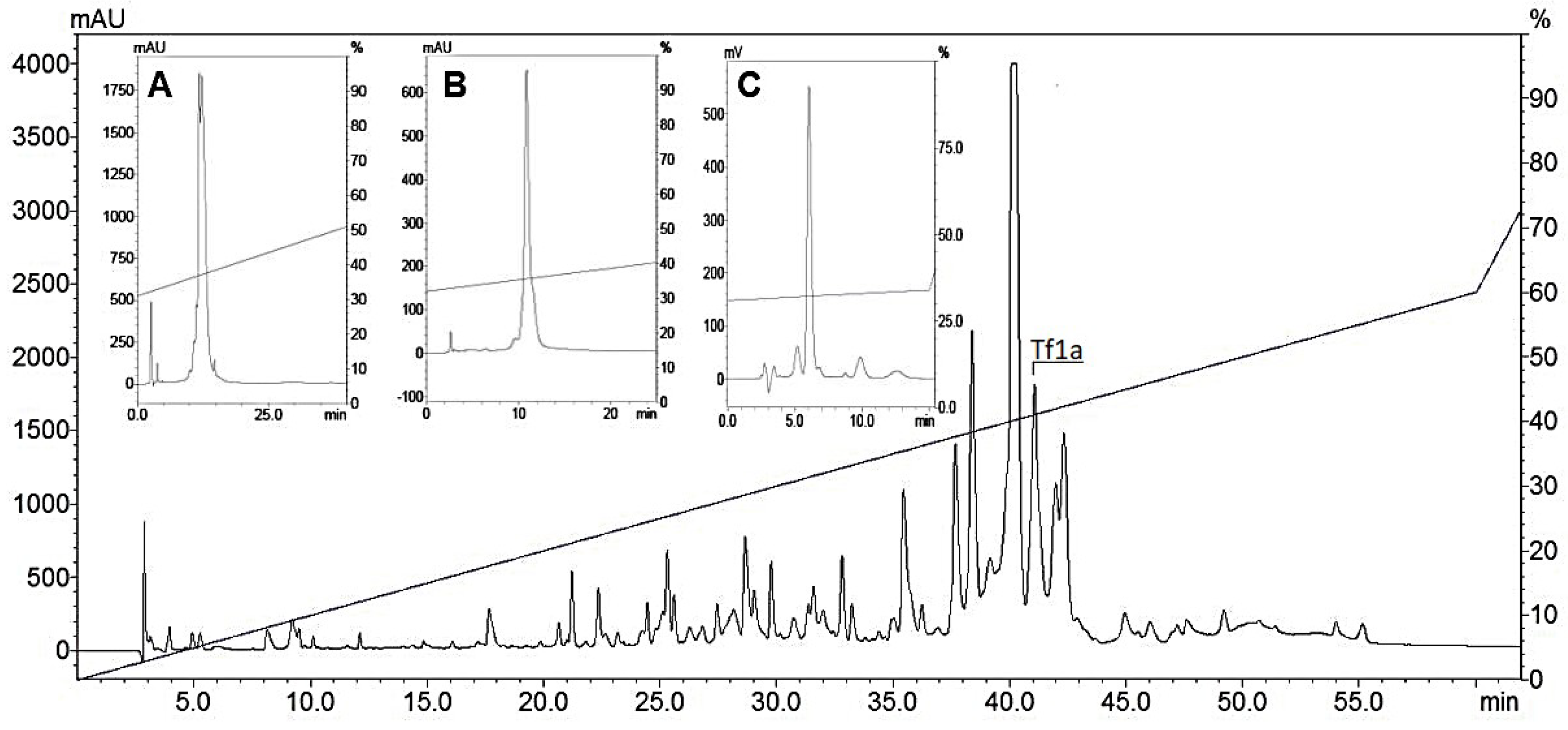
2.1. Toxin Purification
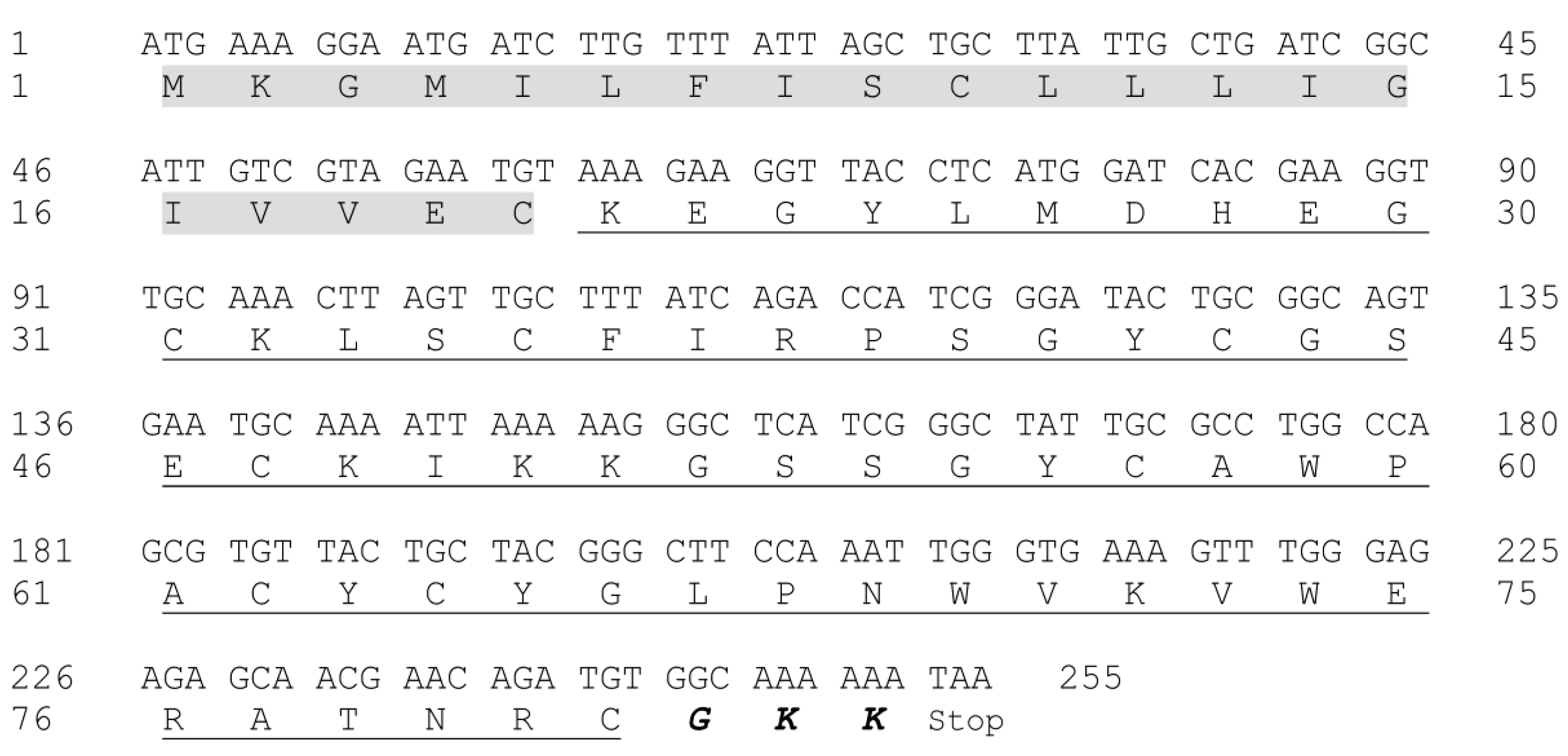
2.2. Molecular Mass Evaluation and Sequence Determination
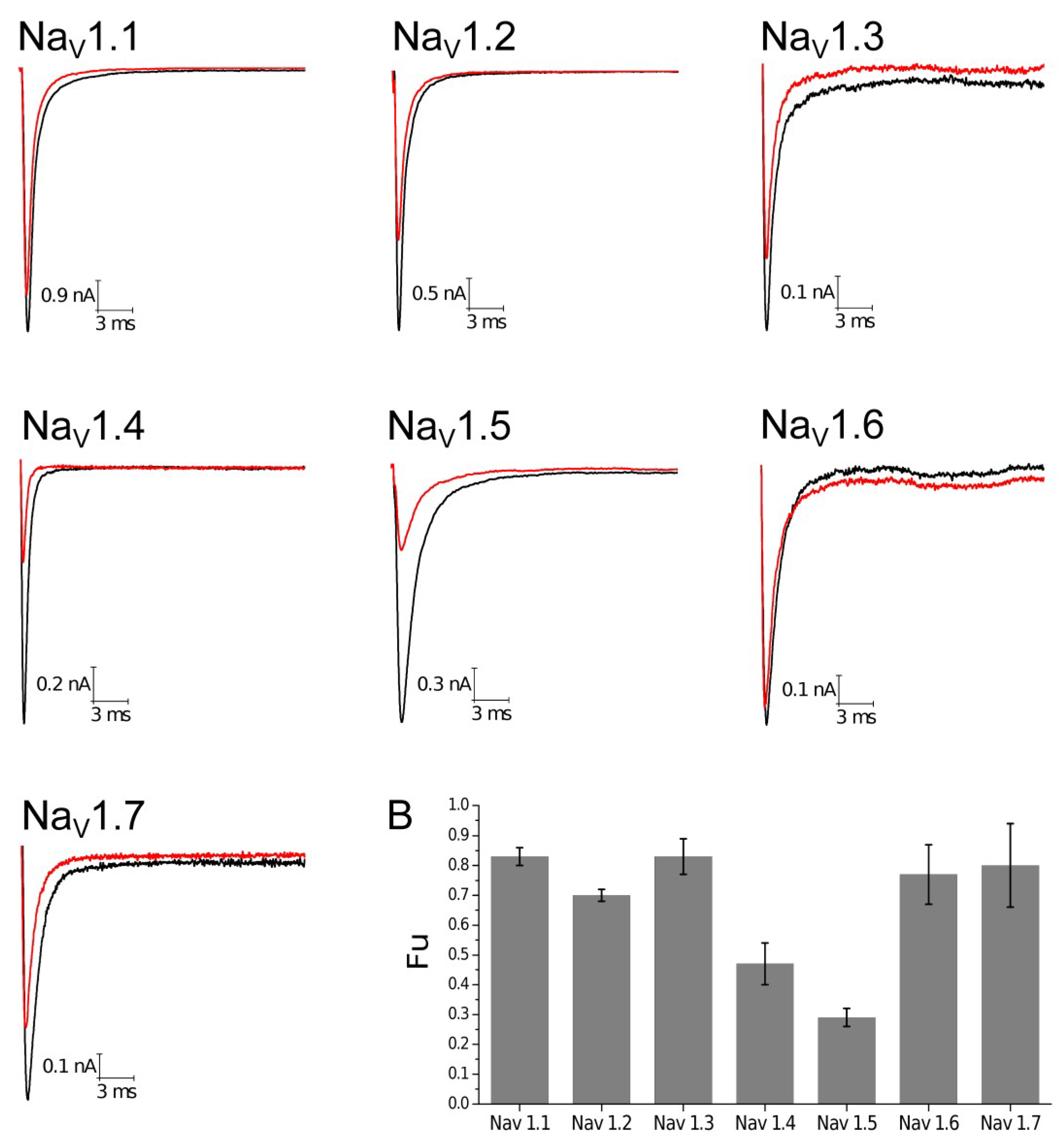
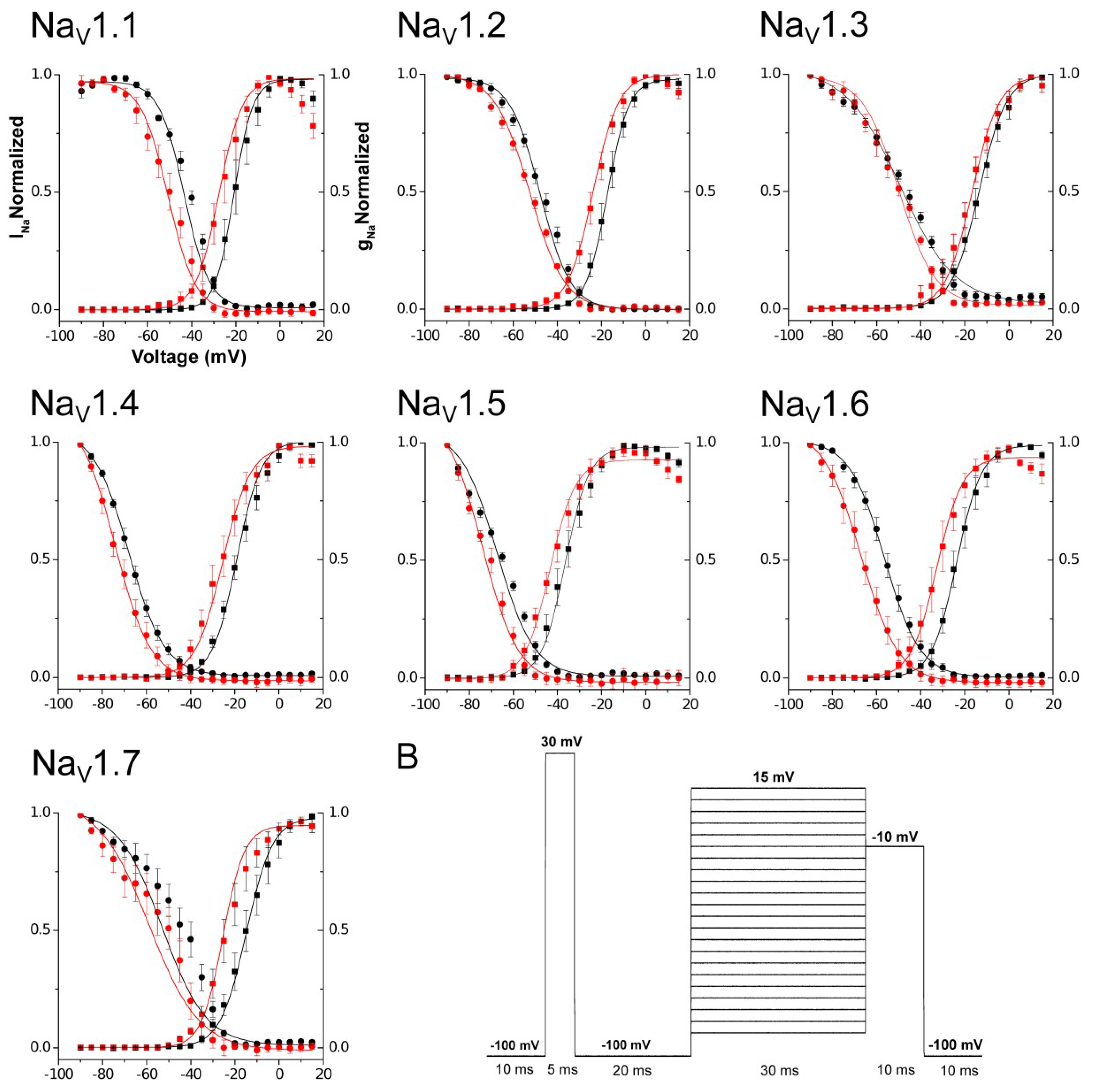
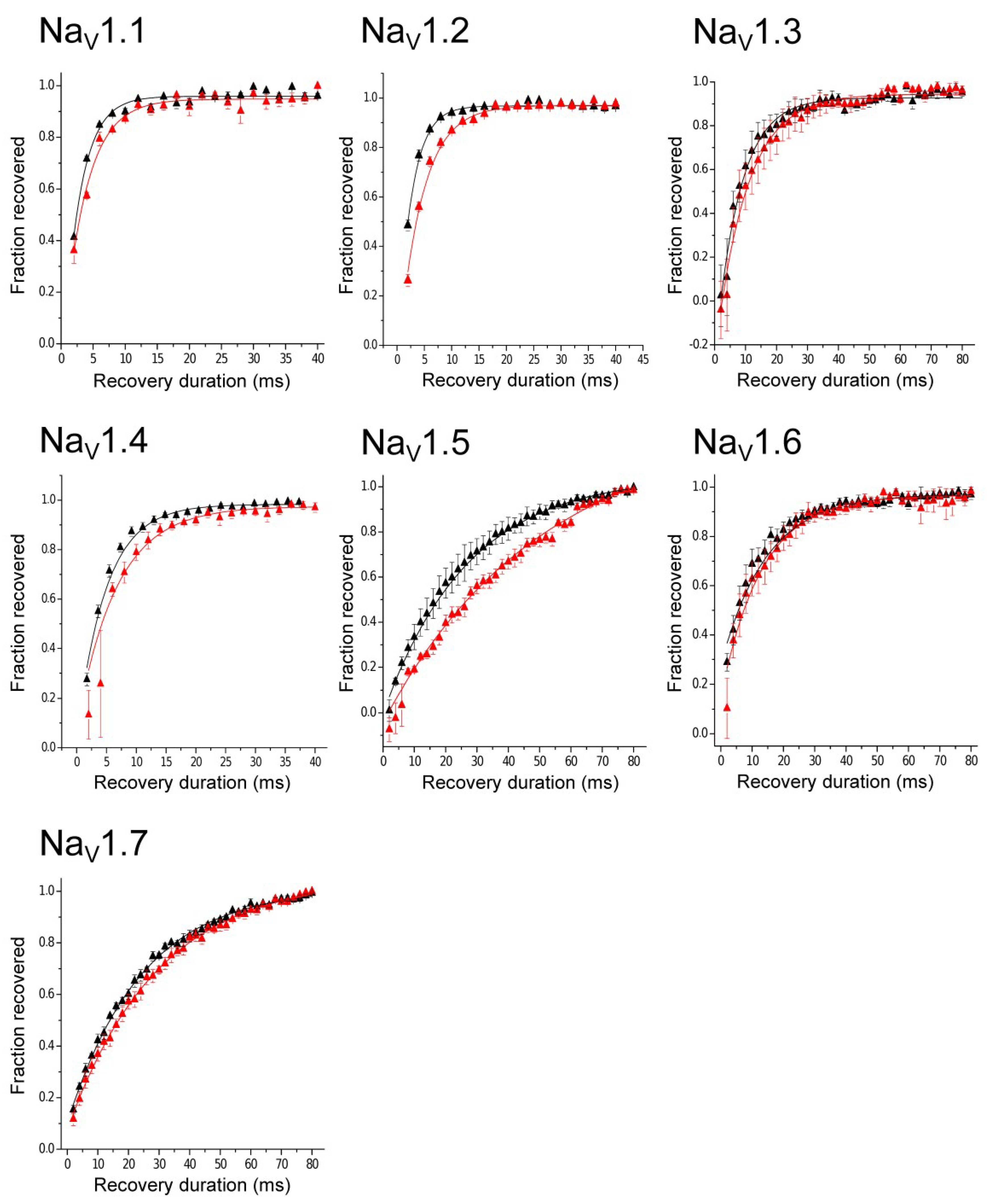
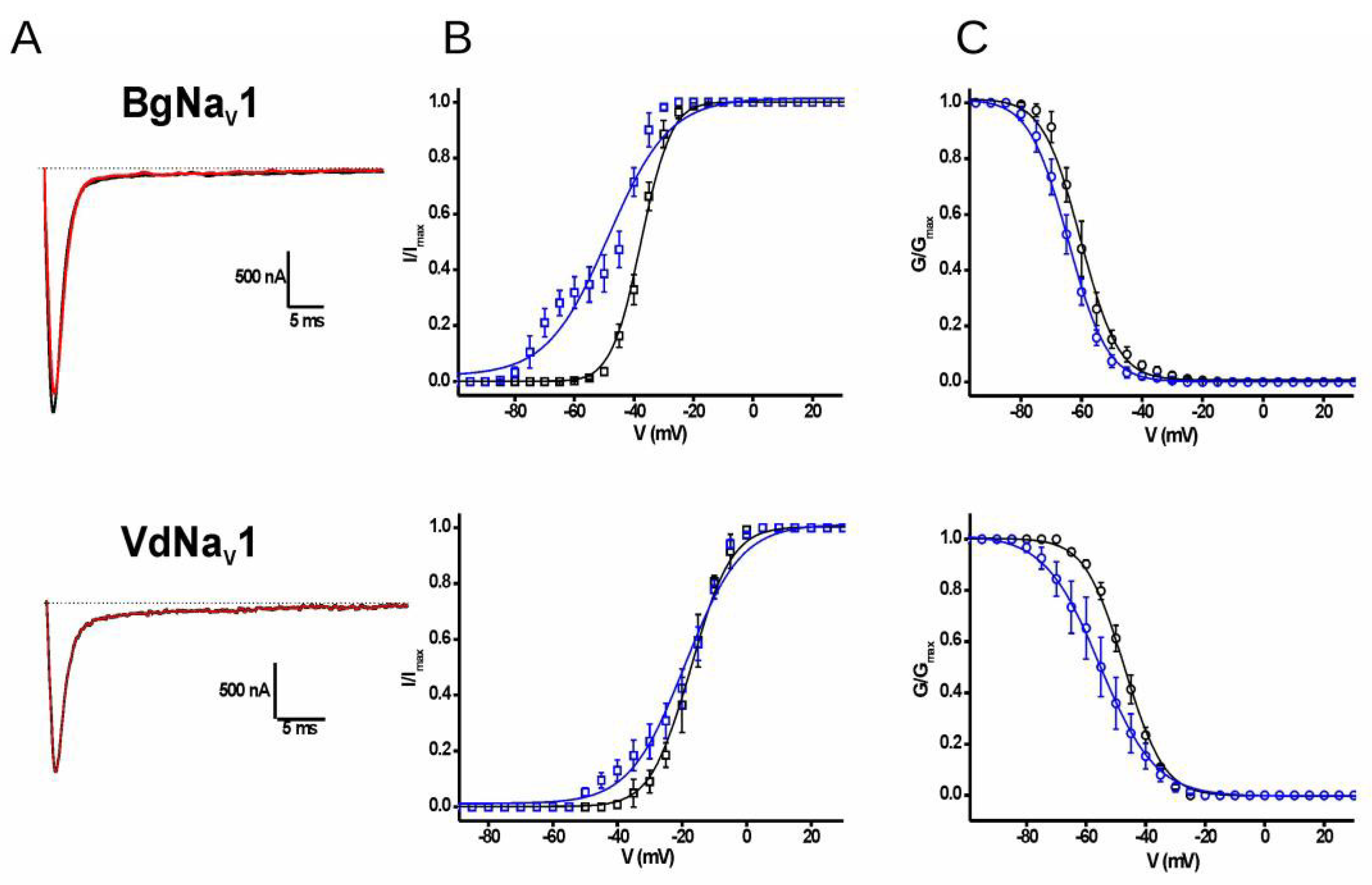
2.3. Electrophysiological Characterization
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals Capture and Venom Extraction
5.2. Toxin Purification
5.3. Molecular Mass and Partial Sequence Determination
5.4. Electrophysiological Assays
5.4.1. Human Channels (hNaV)
Cell Culture
Human NaV Subtypes Current Recording
Data Analysis
5.4.2. Insect and Arachnida Channel Subtypes—Expression of Voltage-Gated Ion Channels in Xenopus Laevis Oocytes
Insect and Arachnida Channels Recordings
Insect and Arachnida Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brazil, T.K.; Porto, T.J. Os Escorpiões; EDUFBA: Salvador, Brasil, 2010; ISBN 9788523207298. Available online: https://repositorio.ufba.br/ri/bitstream/ri/5109/1/Escorpioes-web.pdf (accessed on 17 January 2018).
- Nencioni, A.L.A.; Neto, E.B.; de Freitas, L.A.; Dorce, V.A.C. Effects of Brazilian scorpion venoms on the central nervous system. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.; Castro, M.S.; Barbosa, J.A.R.G.; Fontes, W.; Schwartz, E.N.F.; Sebben, A.; Pires, O.R.; Sousa, M.V.; Schwartz, C.A. Purification and primary structure determination of Tf4, the first bioactive peptide isolated from the venom of the Brazilian scorpion Tityus fasciolatus. Toxicon 2003, 41, 737–745. [Google Scholar] [CrossRef]
- Pinto, M.C.L.; Borboleta, L.R.; Melo, M.B.; Labarrére, C.R.; Melo, M.M. Tityus fasciolatus envenomation induced cardiorespiratory alterations in rats. Toxicon 2010, 55, 1132–1137. [Google Scholar] [CrossRef] [PubMed]
- Mendes, T.M.; Guimarães-Okamoto, P.T.C.; Machado-De-Avila, R.A.; Oliveira, D.; Melo, M.M.; Lobato, Z.I.; Kalapothakis, E.; Chávez-Olórtegui, C. General characterization of Tityus fasciolatus scorpion venom. Molecular identification of toxins and localization of linear B-cell epitopes. Toxicon 2015, 99, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Hernández, V.; Jiménez-Vargas, J.; Gurrola, G.; Valdivia, H.; Possani, L.D. Scorpion venom components that affect ion-channels function. Toxicon 2013, 76, 328–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosmans, F.; Tytgat, J.; Gordon, D.; Ilan, N.; Zilberberg, N.; Gilles, N.; Urbach, D.; Cohen, L.; Karbat, I.; Froy, O.; et al. Voltage-gated sodium channel modulation by scorpion α-toxins. Toxicon 2007, 49, 142–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Possani, L.D.; Merino, E.; Corona, M.; Bolivar, F.; Becerril, B. Peptides and genes coding for scorpion toxins that affect ion-channels. Biochimie 2000, 82, 861–868. [Google Scholar] [CrossRef]
- Guerrero-Vargas, J.A.; Mourão, C.B.F.; Quintero-Hernández, V.; Possani, L.D.; Schwartz, E.F. Identification and Phylogenetic Analysis of Tityus pachyurus and Tityus obscurus Novel Putative Na+-Channel Scorpion Toxins. PLoS ONE 2012, 7, e30478. [Google Scholar] [CrossRef] [PubMed]
- Possani, L.D.; Becerril, B.; Delepierre, M.; Tytgat, J. Scorpion toxins specific for Na+-channels. Eur. J. Biochem. 1999, 264, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.; Savarin, P.; Gurevitz, M.; Zinn-Justin, S. Functional Anatomy of Scorpion Toxins Affecting Sodium Channels. J. Toxicol. Toxin Rev. 1998, 17, 131–159. [Google Scholar] [CrossRef]
- Catterall, W.A. Molecular Properties of Voltage-Sensitive Sodium Channels. Annu. Rev. Biochem. 1986, 55, 953–985. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Forty Years of Sodium Channels: Structure, Function, Pharmacology, and Epilepsy. Neurochem. Res. 2017, 42, 2495–2504. [Google Scholar] [CrossRef] [PubMed]
- Cestèle, S.; Qu, Y.; Rogers, J.C.; Rochat, H.; Scheuer, T.; Catterall, W.A. Voltage Sensor–Trapping: Enhanced Activation of Sodium Channels by β-Scorpion Toxin Bound to the S3–S4 Loop in Domain II Sandrine. Neuron 1998, 21, 919–931. [Google Scholar] [CrossRef]
- Goldschen-Ohm, M.P.; Capes, D.L.; Oelstrom, K.M.; Chanda, B. Multiple pore conformations driven by asynchronous movements of voltage sensors in a eukaryotic sodium channel. Nat. Commun. 2013, 4, 1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldin, A.L. Resurgence of Sodium Channel Research. Annu. Rev. Physiol. 2001, 63, 871–894. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Voltage-gated sodium channels at 60: Structure, function and pathophysiology. J. Physiol. 2012, 590, 2577–2589. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Liu, M.; Yan, S.F.; Yan, N. Structure-based assessment of disease-related mutations in human voltage-gated sodium channels. Protein Cell 2017, 8, 401–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimarães, P.T.C.; Pinto, M.C.L.; Melo, M.M. Perfis clínico e hematológico de camundongos submetidos ao envenenamento escorpiônico experimental por Tityus fasciolatus. Arq. Bras. Med. Veterinária e Zootec. 2011, 63, 1382–1390. [Google Scholar] [CrossRef]
- Camargos, T.S.; Bosmans, F.; Rego, S.C.; Mourão, C.B.F.; Schwartz, E.F. The scorpion toxin Tf2 from Tityus fasciolatus promotes NaV1.3 opening. PLoS ONE 2015, 10, e0128578. [Google Scholar] [CrossRef] [PubMed]
- Coelho, V.A.; Cremonez, C.M.; Anjolette, F.A.P.; Aguiar, J.F.; Varanda, W.A.; Arantes, E.C. Functional and structural study comparing the C-terminal amidated β-neurotoxin Ts1 with its isoform Ts1-G isolated from Tityus serrulatus venom. Toxicon 2014, 83, 15–21. [Google Scholar] [CrossRef] [PubMed]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peigneur, S.; Cologna, C.T.; Cremonez, C.M.; Mille, B.G.; Pucca, M.B.; Cuypers, E.; Arantes, E.C.; Tytgat, J. A gamut of undiscovered electrophysiological effects produced by Tityus serrulatus toxin 1 on NaV-type isoforms. Neuropharmacology 2015, 95, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Duque, H.M.; Mourão, C.B.F.; Tibery, D.V.; Barbosa, E.A.; Campos, L.A.; Schwartz, E.F. To4, the first Tityus obscurus β-toxin fully electrophysiologically characterized on human sodium channel isoforms. Peptides 2017, 95, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Zhijian, C.; Feng, L.; Yingliang, W.; Xin, M.; Wenxin, L. Genetic mechanisms of scorpion venom peptide diversification. Toxicon 2006, 47, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Escalon, M.P.; Possani, L.D. Scorpion beta-toxins and voltage-gated sodium channels: Interactions and effects. Front. Biosci. 2013, 18, 572–587. [Google Scholar] [CrossRef]
- Coronas, F.I.V.; Diego-García, E.; Restano-Cassulini, R.; de Roodt, A.R.; Possani, L.D. Biochemical and physiological characterization of a new Na+-channel specific peptide from the venom of the Argentinean scorpion Tityus trivittatus. Peptides 2015, 68, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.; Alfonzo, M.J.; García, C.C.; Winand, N.J.; Leipold, E.; Heinemann, S.H.; García, C.C.; Winand, N.J.; Leipold, E.; Heinemann, S.H. Isolation, molecular cloning and functional characterization of a novel β-toxin from the Venezuelan scorpion, Tityus zulianus. Toxicon 2004, 43, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Leipold, E.; Hansel, A.; Borges, A.; Heinemann, S.H. Subtype specificity of scorpion beta-toxin Tz1 interaction with voltage-gated sodium channels is determined by the pore loop of domain 3. Mol. Pharmacol. 2006, 70, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Leipold, E.; Borges, A.; Heinemann, S.H. Scorpion β-toxin interference with NaV channel voltage sensor gives rise to excitatory and depressant modes. J. Gen. Physiol. 2012, 139, 305–319. [Google Scholar] [CrossRef] [PubMed]
- Díaz, P.; D’Suze, G.; Salazar, V.; Sevcik, C.; Shannon, J.D.; Sherman, N.E.; Fox, J.W. Antibacterial activity of six novel peptides from Tityus discrepans scorpion venom. A fluorescent probe study of microbial membrane Na+ permeability changes. Toxicon 2009, 54, 802–817. [Google Scholar] [CrossRef] [PubMed]
- Peigneur, S.; Sevcik, C.; Tytgat, J.; Castillo, C.; D’Suze, G. Subtype specificity interaction of bactridines with mammalian, insect and bacterial sodium channels under voltage clamp conditions. FEBS J. 2012, 279, 4025–4038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barona, J.; Batista, C.V.F.; Zamudio, F.Z.; Gomez-Lagunas, F.; Wanke, E.; Otero, R.; Possani, L.D. Proteomic analysis of the venom and characterization of toxins specific for Na+- and K+-channels from the Colombian scorpion Tityus pachyurus. Biochim. Biophys. Acta Proteins Proteom. 2006, 1764, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Z.; Yarov-Yarovoy, V.; Scheuer, T.; Karbat, I.; Cohen, L.; Gordon, D.; Gurevitz, M.; Catterall, W.A. Structure-Function Map of the Receptor Site for β-Scorpion Toxins in Domain II of Voltage-gated Sodium Channels. J. Biol. Chem. 2011, 286, 33641–33651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, L.; Ilan, N.; Gur, M.; Stühmer, W.; Gordon, D.; Gurevitz, M. Design of a Specific Activator for Skeletal Muscle Sodium Channels Uncovers Channel Architecture. J. Biol. Chem. 2007, 282, 29424–29430. [Google Scholar] [CrossRef] [PubMed]
- Balser, J.R. The Cardiac Sodium Channel: Gating Function and Molecular Pharmacology. J. Mol. Cell. Cardiol. 2001, 33, 599–613. [Google Scholar] [CrossRef] [PubMed]
- Campos, F.V.; Chanda, B.; Beirão, P.S.L.; Bezanilla, F. β-Scorpion Toxin Modifies Gating Transitions in All Four Voltage Sensors of the Sodium Channel. J. Gen. Physiol. 2007, 130, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Moran, O.; Nizzari, M.; Conti, F. Endogenous expression of the β1A sodium channel subunit in HEK-293 cells. FEBS Lett. 2000, 473, 132–134. [Google Scholar] [CrossRef] [Green Version]
- Liman, E.R.; Tytgat, J.; Hess, P. Subunit Stoichiometry of a Mammalian K + Channel Determ ined by Construction of Multimeric cDNAs. Neuron 1992, 9, 861–871. [Google Scholar] [CrossRef]
- Pucca, M.; Cerni, F.; Peigneur, S.; Bordon, K.; Tytgat, J.; Arantes, E. Revealing the Function and the Structural Model of Ts4: Insights into the “Non-Toxic” Toxin from Tityus serrulatus Venom. Toxins 2015, 7, 2534–2550. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vg Control (mV) | Vg Toxin (mV) | k Control (mV) | k Toxin (mV) | Fu | |
|---|---|---|---|---|---|
| Mean | Mean | Mean | Mean | Mean | |
| hNaV1.1 | −20.21 ± 2.24 | −27.30 ± 2.51 | 4.29 ± 0.54 | 4.53 ± 0.41 | 0.83 ± 0.03 |
| hNaV1.2 | −17.21 ± 1.71 | −23.30 ± 1.61 | 4.89 ± 0.35 | 5.55 ± 0.08 | 0.70 ± 0.02 |
| hNaV1.3 | −12.91 ± 1.98 | −17.70±2.12 | 6.93 ± 0.46 | 7.38 ± 0.52 | 0.83 ± 0.06 |
| hNaV1.4 | −18.41 ± 1.69 | −26.34 ± 2.93 | 6.71 ± 0.35 | 6.61 ± 0.46 | 0.47 ± 0.07 |
| hNaV1.5 | −36.62 ± 2.10 | −42.91 ± 1.72 | 5.95 ± 0.37 | 6.40 ± 0.41 | 0.29 ± 0.03 |
| hNaV1.6 | −23.40 ± 2.01 | −32.41 ± 2.80 | 5.70 ± 0.58 | 5.72 ± 0.29 | 0.77 ± 0.10 |
| hNaV1.7 | −18 ± 2.69 | −25.93 ± 1.84 | 5.90 ± 0.66 | 5.83 ± 0.53 | 0.80 ± 0.14 |
| Vh Control (mV) | Vh Toxin (mV) | kh Control (mV) | kh Toxin (mV) | |
|---|---|---|---|---|
| Mean | Mean | Mean | Mean | |
| hNaV1.1 | −45.92 ± 1.17 | −54.03 ± 2.24 | 8.48 ± 0.28 | 8.6 ± 0.12 |
| hNaV1.2 | −46.92 ± 1.22 | −52.29 ± 0.94 | 7.98 ± 0.32 | 8.45 ± 0.49 |
| hNaV1.3 | −47.57 ± 1.52 | −50.5 ± 1.71 | 11.34 ± 0.57 | 9.92 ± 1.02 |
| hNaV1.4 | −68.15 ± 1.22 | −74.31 ± 2.25 | 8.19 ± 0.43 | 8.25 ± 0.31 |
| hNaV1.5 | −66.18 ± 0.99 | −72.71 ± 1.43 | 9.38 ± 0.47 | 7.85 ± 0.41 |
| hNaV1.6 | −55.78 ± 1.90 | −67.19 ± 2.52 | 8.03 ± 0.26 | 8.61 ± 0.64 |
| hNaV1.7 | −47.46 ± 3.85 | −55.45 ± 6.04 | 11.20 ± 1.32 | 11.20 ± 1.70 |
| τ Control (ms) | τ Toxin (ms) | ||
|---|---|---|---|
| n | Mean | Mean | |
| hNaV1.1 | 3 | 3.01 ± 0.27 | 4.07 ± 0.41 |
| hNaV1.2 | 4 | 2.42 ± 0.16 | 3.90 ± 0.30 |
| hNaV1.3 | 4 | 10.12 ± 2.62 | 12.55 ± 3.97 |
| hNaV1.4 | 4 | 4.33 ± 0.37 | 5.21 ± 0.48 |
| hNaV1.5 | 3 | 25.0 ± 5.62 | 45.11 ± 3.99 |
| hNaV1.6 | 4 | 11.53 ± 1.30 | 12.76 ± 2.22 |
| hNaV1.7 | 4 | 24.35 ± 0.89 | 29.65 ± 2.20 |
| Vg Control (mV) | Vg Toxin (mV) | Vh Toxin (mV) | Vh Toxin (mV) | |
|---|---|---|---|---|
| Mean | Mean | Mean | Mean | |
| BgNaV1 | −37.6 ± 0.1 | −48.6 ± 1.2 | −60.3 ± 0.2 | −64.4 ± 0.1 |
| VdNaV1 | −17.2 ± 0.1 | −19.0 ± 0.5 | −47.2 ± 0.1 | −55.5 ± 0.2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mata, D.O.d.; Tibery, D.V.; Campos, L.A.; Camargos, T.S.; Peigneur, S.; Tytgat, J.; Schwartz, E.F. Subtype Specificity of β-Toxin Tf1a from Tityus fasciolatus in Voltage Gated Sodium Channels. Toxins 2018, 10, 339. https://doi.org/10.3390/toxins10090339
Mata DOd, Tibery DV, Campos LA, Camargos TS, Peigneur S, Tytgat J, Schwartz EF. Subtype Specificity of β-Toxin Tf1a from Tityus fasciolatus in Voltage Gated Sodium Channels. Toxins. 2018; 10(9):339. https://doi.org/10.3390/toxins10090339
Chicago/Turabian StyleMata, Daniel Oliveira da, Diogo Vieira Tibery, Leandro Ambrósio Campos, Thalita Soares Camargos, Steve Peigneur, Jan Tytgat, and Elisabeth Ferroni Schwartz. 2018. "Subtype Specificity of β-Toxin Tf1a from Tityus fasciolatus in Voltage Gated Sodium Channels" Toxins 10, no. 9: 339. https://doi.org/10.3390/toxins10090339
APA StyleMata, D. O. d., Tibery, D. V., Campos, L. A., Camargos, T. S., Peigneur, S., Tytgat, J., & Schwartz, E. F. (2018). Subtype Specificity of β-Toxin Tf1a from Tityus fasciolatus in Voltage Gated Sodium Channels. Toxins, 10(9), 339. https://doi.org/10.3390/toxins10090339