Malting of Fusarium Head Blight-Infected Rye (Secale cereale): Growth of Fusarium graminearum, Trichothecene Production, and the Impact on Malt Quality

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Rye Samples and Fusarium Species
2.2. Development of Trichothecenes during the Malting of Rye
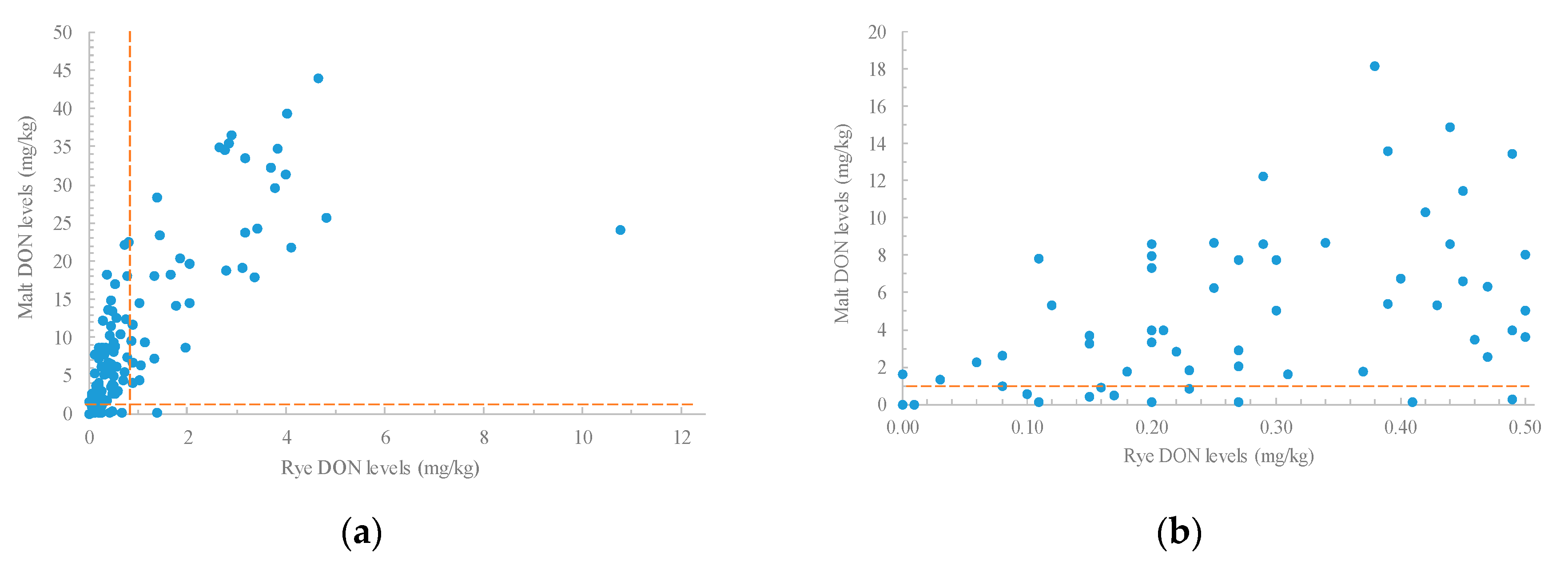
2.3. Relationships between Levels of Trichothecenes in Rye and Malt
2.4. Impact of FHB on Grain and Malt Quality of Rye
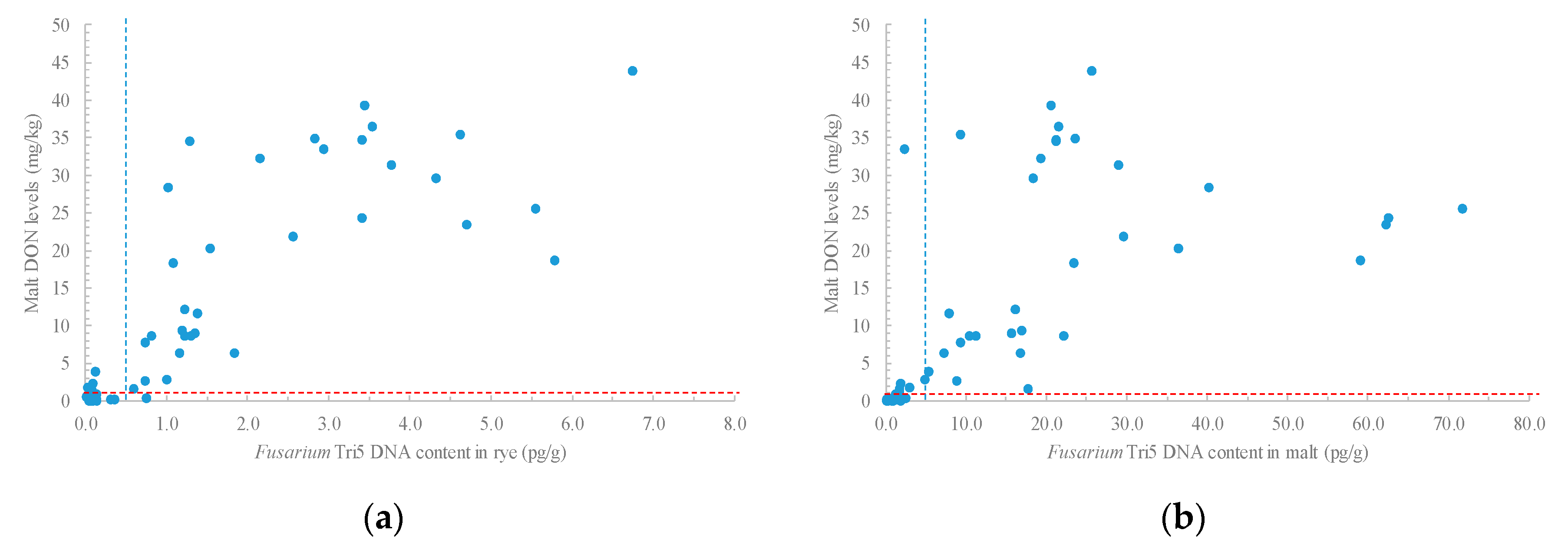
2.5. Relationships between Fusarium Tri5 DNA Levels and Trichotechenes and Malt Quality
3. Conclusions
4. Materials and Methods
4.1. Rye Samples
4.2. Micro-Malting
4.3. Measurement of Malt Quality Parameters
4.4. Determination of Type B Trichothecenes and D3G
4.5. Identification of Fusarium Species
4.6. Quantitative Analysis of Fusarium Tri5 DNA
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Schwarz, P.B.; Casper, H.H.; Beattie, S. Fate and development of naturally-occurring Fusarium mycotoxins during malting and brewing. J. Am. Soc. Brew. Chem. 1995, 53, 121–127. [Google Scholar]
- Lancova, K.; Hajslova, J.; Poustka, J.; Krplova, A.; Zachariasova, M.; Dostalek, P.; Sachambula, L. Transfer of Fusarium mycotoxins and “masked’’ deoxynivalenol (deoxynivalenol-3-glucoside) from field barley through malt to beer. Food Addit. Contam. Part A 2008, 25, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Kostelanska, M.; Hajslova, J.; Zachariasova, M.; Malachova, A.; Kalachova, K.; Poustka, J.; Fiala, J.; Scott, P.M.; Berthiller, F.; Krska, R. Occurrence of deoxynivalenol and its major conjugate, deoxynivalenol-3- glucoside, in beer and some brewing intermediates. J. Agric. Food Chem. 2009, 57, 3187–3194. [Google Scholar] [CrossRef] [PubMed]
- Varga, E.; Malachova, A.; Schwartz, H.; Krska, R.; Berthiller, F. Survey of deoxynivalenol and its conjugates deoxynivalenol-3-glucoside and 3-acetyl-deoxynivalenol in 374 beer samples. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2013, 30, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Kuzdraliński, A.; Solarska, E.; Muszyńska, M. Deoxynivalenol and zearalenone occurence in beers analysed by an enzyme-linked immunosorbent assay method. Food Control 2013, 29, 22–24. [Google Scholar] [CrossRef]
- Bertuzzi, T.; Rastelli, S.; Mulazzi, A.; Donadini, G.; Pietri, A. Mycotoxin occurrence in beer produced in several European countries. Food Control 2011, 22, 2059–2064. [Google Scholar] [CrossRef]
- Bryła, M.; Ksieniewicz-Woźniak, E.; Waśkiewicz, A.; Szymczyk, K.; Jędrzejczak, R. Co-occurrence of nivalenol, deoxynivalenol and deoxynivalenol-3-glucoside in beer samples. Food Control 2018, 92, 319–324. [Google Scholar] [CrossRef]
- Piacentini, K.C.; Rocha, L.O.; Fontes, L.C.; Carnielli, L.; Reis, T.A.; Corrêa, B. Mycotoxin analysis of industrial beers from Brazil: The influence of fumonisin B1 and deoxynivalenol in beer quality. Food Chem. 2017, 218, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Piacentini, K.C.; Savi, G.D.; Olivo, G.; Scussel, V.M. Quality and occurrence of deoxynivalenol and fumonisins in craft beer. Food Control 2015, 50, 925–929. [Google Scholar] [CrossRef]
- Desjardins, A.E. Fusarium mycotoxins Chemistry, Genetics, and Biology; The American Phytopathological Society: St. Paul, MN, USA, 2006; ISBN 0-89054-335-336. [Google Scholar]
- Bond, J.K.; Capehart, T.; Allen, E.; Kim, G. Expanding craft beer production boosts industrial use of barley. Amber Waves 2015, 20–22. [Google Scholar]
- Schwarz, P.B. Fusarium head blight and deoxynivalenol in malting and brewing: Successes and future challenges. Trop. Plant Pathol. 2017, 42, 153–164. [Google Scholar] [CrossRef]
- Beattie, S.; Schwarz, P.B.; Horsley, R.; Barr, J.; Casper, H.H. The effect of grain storage conditions on the viability of Fusarium and deoxynivalenol production in infested malting barley. J. Food Prot. 1998, 61, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Karlovsky, P. Biological detoxification of the mycotoxin deoxynivalenol and its use in genetically engineered crops and feed additives. Appl. Microbiol. Biotechnol. 2011, 91, 491–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slater, A.; (Busch Agricultural Recources, LL., Moorhead, Minnesota); Schwarz, P.B.; (North Dakota State Univerisity, Fargo, ND, USA). Personal communication, 2018.
- Schwarz, P.B. Impact of Fusarium Height Blight on malting and brewing quality of barley. In Fusarium Height Blight of Wheat and Barley; Leonard, K., Bushnell, W., Eds.; APS Press: St. Paul, MN, USA, 2003; pp. 395–419. [Google Scholar]
- Sarlin, T.; Laitila, A.; Pekkarinen, A.; Haikara, A. Effects of three Fusarium species on the quality of barley and malt. J. Am. Soc. Brew. Chem. 2005, 63, 43–49. [Google Scholar] [CrossRef]
- Schwarz, P.B.; Horsley, R.D.; Steffenson, B.J.; Salas, B.; Barr, J. Quality risks associated with the utilization of Fusarium Head Blight infected malting barley. J. Am. Soc. Brew. Chem. 2006, 64, 1–7. [Google Scholar] [CrossRef]
- Nielsen, L.K.; Cook, D.J.; Edwards, S.G.; Ray, R.V. The prevalence and impact of Fusarium Head Blight pathogens and mycotoxins on malting barley quality in UK. Int. J. Food Microbiol. 2014, 179, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Sarlin, T.; Kivioja, T.; Kalkkinen, N.; Linder, M.B.; Nakari-Setälä, T. Identification and characterization of gushing-active hydrophobins from Fusarium graminearum and related species. J. Basic Microbiol. 2012, 52, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Krstanović, V.; Mastanjević, K.; Velić, N.; Pleadin, J.; Perši, N.; Španić, V. The influence of Fusarium culmorum contamination level on deoxynivalenol content in wheat, malt and beer. Rom. Biotechnol. Lett. 2015, 20, 10901–10910. [Google Scholar]
- Spanic, V.; Marcek, T.; Abicic, I.; Sarkanj, B. Effects of Fusarium head blight on wheat grain and malt infected by Fusarium culmorum. Toxins 2017, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Zhou, B.; Gillespie, J.; Gross, T.; Barr, J.; Simsek, S.; Brueggeman, R.; Schwarz, P.B. Production of deoxynivalenol (DON) and DON-3-glucoside during the malting of Fusarium infected hard red spring wheat. Food Control 2017, 85, 6–10. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, Z.; Barr, J.; Gillespie, J.; Simsek, S.; Horsley, R.D.; Schwarz, P.B. Micro-malting for the quality evaluation of rye (Secale cereale) genotypes. Fermentation 2018, 4, 50. [Google Scholar] [CrossRef]
- Schlegel, R.H.J. Rye: Genetics, Breeding, and Cultivation; Schlegel, R.H.J., Ed.; CRC Press Taylor & Fracis Group, LLC: Boca Raton, FL, USA, 2014; ISBN 9781466561434. [Google Scholar]
- Eskola, M.; Parikka, P.; Rizzo, A. Trichothecenes, ochratoxin A and zearalenone contamination and Fusarium infection in Finnish cereal samples in 1998. Food Addit. Contam. 2001, 18, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Döll, S.; Valenta, H.; Kirchheim, U.; Dänicke, S.; Flachowsky, G. Fusarium mycotoxins in conventionally and organically grown grain from Thuringia/Germany. Landbauforsch. Volkenrode 2002, 52, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Arseniuk, E.; Foremska, E.; Góral, T.; Chełkowski, J. Fusarium head blight reactions and accumulation of deoxynivalenol (DON) and some of its derivatives in kernels of wheat, triticale and rye. J. Phytopathol. 1999, 147, 577–590. [Google Scholar] [CrossRef]
- Gräfenhan, T.; Patrick, S.K.; Roscoe, M.; Trelka, R.; Gaba, D.; Chan, J.M.; McKendry, T.; Clear, R.M.; Tittlemier, S.A. Fusarium damage in cereal grains from western Canada. 1. Phylogenetic analysis of moniliformin-producing fusarium species and their natural occurrence in mycotoxin-contaminated wheat, oats, and rye. J. Agric. Food Chem. 2013, 61, 5425–5437. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D. Malting: The latest craft. Brew. Distill. Int. 2013, 47–49. [Google Scholar]
- Salas, B.; Steffenson, B.J.; Casper, H.H.; Tacke, B.; Prom, L.K.; Fetch, T.G.; Schwarz, P.B. Species pathogenic to barley and their associated mycotoxins. Plant Dis. 1999, 83, 667–674. [Google Scholar] [CrossRef]
- Poppenberger, B.; Berthiller, F.; Lucyshyn, D.; Sieberer, T.; Schuhmacher, R.; Krska, R.; Kuchler, K.; Glossl, J.; Luschnig, C.; Adam, G. Detoxification of the Fusarium mycotoxin deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 47905–47914. [Google Scholar] [CrossRef] [PubMed]
- Dall’Erta, A.; Martina, C.; Margherita, D. Masked mycotoxins are efficiently hydrolyzed by human colonic microbiota releasing their aglycones. Chem. Res. Toxicol. 2013, 26, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Nagl, V.; Woechtl, B.; Schwartz-Zimmermann, H.E.; Henni g-Pauka, I.; Moll, W.D.; Adam, G.; Berthiller, F. Metabolism of the masked mycotoxin deoxynivalenol-3-glucoside in pigs. Toxicol. Lett. 2014, 229, 190–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minervini, F.; Fornelli, F.; Flynn, K.M. Toxicity and apoptosis induced by the mycotoxins nivalenol, deoxynivalenol and fumonisin B1 in a human erythroleukemia cell line. Toxicol. In Vitro 2004, 18, 21–28. [Google Scholar] [CrossRef]
- Pinton, P.; Tsybulskyy, D.; Lucioli, J.; Laffitte, J.; Callu, P.; Lyazhri, F.; Grosjean, F.; Bracarense, A.P.; Kolf-clauw, M.; Oswald, I.P. Toxicity of deoxynivalenol and its acetylated derivatives on the intestine: Differential effects on morphology, barrier function, tight junction proteins, and mitogen-activated protein kinases. Toxicol. Sci. 2012, 130, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Habler, K.; Katharina, H.; Cajetan, G.; Jan, S.; Ralph, H.; Michael, H.; Martina, G.; Michael, R. Fate of Fusarium toxins during the malting process. J. Agric. Food Chem. 2016, 64, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Hübner, F.; Schehl, B.D.; Gebruers, K.; Courtin, C.M.; Delcour, J.A.; Arendt, E.K. Influence of germination time and temperature on the properties of rye malt and rye malt based worts. J. Cereal Sci. 2010, 52, 72–79. [Google Scholar] [CrossRef]
- Burr, S.J.; Fry, S.C. Extracellular cross-linking of maize arabinoxylans by oxidation of feruloyl esters to form oligoferuloyl esters and ether-like bonds. Plant J. 2009, 58, 554–567. [Google Scholar] [CrossRef] [PubMed]
- Panina, Y.; Fravel, D.R.; Baker, C.J.; Shcherbakova, L.A. Biocontrol and plant pathogenic Fusarium oxysporum-induced changes in phenolic compounds in tomato leaves and roots. J. Phytopathol. 2007, 155, 475–481. [Google Scholar] [CrossRef]
- Mallett, J. Malt: A Practical Guide from Field to Brewhouse; Hertrich, J., Palmer, J., Eds.; Brewer Publication: Boulder, CO, USA, 2014; ISBN 1938469127. [Google Scholar]
- Prom, L.K.; Horsley, R.D.; Steffenson, B.J.; Schwarz, P.B. Development of Fusarium head blight and accumulation of deoxynivalenol in barley sampled at different growth stages. J. Am. Soc. Brew. Chem. 1999, 57, 60–63. [Google Scholar] [CrossRef]
- Spring Barley Growth and Development Guide. 2018. Available online: https://extension.umn.edu/growing-small-grains/spring-barley-growth-and-development-guide#pollination-792764 (accessed on 14 August 2018).
- Bonnett, O.T. Inflorescences of Maize, Wheat, Rye, Barley, and Oats: Their Initiation and Development; Bulletin (University of Illinois (Urbana-Champaign Campus)) Agricultural Experiment Station: Urbana, IL, USA, 1966. [Google Scholar]
- Góral, T.; Wiśniewska, H.; Ochodzki, P.; Walentyn-Góral, D. Higher Fusarium toxin accumulation in grain of winter triticale lines inoculated with Fusarium culmorum as compared with wheat. Toxins 2016, 8, 301. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, P.B.; Qian, S.Y.; Zhou, B.; Xu, Y.; Barr, J.; Horsley, R.D.; Gillespie, J. Occurrence of deoxynivalenol-3-glucoside on barley from the upper midwestern United States. J. Am. Soc. Brew. Chem. 2014, 72, 208–213. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Location | No. of Samples | DON in Rye (mg/kg) | DON in Malt (mg/kg) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Min | Max | Sample (%) | Mean | Min | Max | Sample (%) | |||||||
| <0.50 | 0.50–1.00 | >1.00 | <0.50 | 0.50–1.00 | 1.01–10.00 | >10.00 | ||||||||
| ND ab | 14 | <0.50 c | <0.20 d | 1.37 | 91 | 9 | 0 | 1.03 | <0.20 | 3.95 | 43 | 21 | 36 | 0 |
| NY a | 22 | 2.04 | <0.50 | 4.66 | 23 | 18 | 59 | 19.64 | 2.04 | 43.92 | 0 | 0 | 50 | 50 |
| MN a | 81 | 0.94 | <0.20 | 10.76 | 54 | 22 | 23 | 9.77 | <0.20 | 28.34 | 11 | 1 | 50 | 38 |
| Total | 117 | 1.06 | <0.20 | 10.76 | 55 | 18 | 27 | 10.60 | <0.20 | 43.92 | 13 | 3 | 49 | 36 |
| Location | No. of Samples | D3G in Rye (mg/kg) | D3G in Malt (mg/kg) | ||||
|---|---|---|---|---|---|---|---|
| Mean | Min | Max | Mean | Min | Max | ||
| ND a | 14 | <0.50 b | <0.10 c | <0.50 | 1.19 | <0.50 | 2.67 |
| NY | 22 | 1.51 | <0.50 | 3.22 | 11.03 | 2.81 | 20.02 |
| MN | 81 | 0.51 | <0.10 | 2.76 | 7.50 | <0.50 | 14.71 |
| total | 117 | 0.65 | <0.10 | 3.22 | 7.41 | <0.50 | 20.02 |
| Location | No. of Samples | 3-ADON in Malt (mg/kg) | 15-ADON in Malt (mg/kg) | NIV in Malt (mg/kg) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Min | Max | Mean | Min | Max | Mean | Min | Max | ||
| ND b | 14 | <0.20 c | <0.20 | <0.50 d | <0.50 | <0.20 | <0.50 | <0.50 | <0.20 | <0.50 |
| NY | 22 | 1.43 | <0.20 | 3.97 | 1.34 | <0.20 | 2.78 | <0.50 | <0.20 | 0.66 |
| MN | 81 | 0.52 | <0.20 | 2.64 | 1.18 | <0.20 | 5.20 | <0.50 | <0.20 | 1.81 |
| total | 117 | 0.63 | <0.20 | 3.97 | 1.08 | <0.20 | 5.20 | <0.50 | <0.20 | 1.81 |
| Trichothecenes | Rye DON | Malt DON | Malt D3G | Malt 3-ADON | Malt 15-ADON | Malt NIV |
|---|---|---|---|---|---|---|
| Rye DON | 1 | |||||
| Rye D3G | 0.81 *** | |||||
| Malt DON | 0.74 *** | 1 | ||||
| Malt D3G | 0.67 *** | 0.96 *** | 1 | |||
| Malt 3-ADON | 0.63 *** | 0.92 *** | 0.85 *** | 1 | ||
| Malt 15-ADON | 0.58 *** | 0.80 *** | 0.82 *** | 0.61 *** | 1 | |
| Malt NIV | 0.52 *** | 0.38 *** | 0.35 *** | 0.45 *** | 1 | |
| Grain quality | ||||||
| Rye thousand kernel weight | 0.37 *** | 0.29 * | 0.28 * | 0.28 * | ||
| Plump kernels | 0.36 *** | 0.31 ** | 0.86 *** | |||
| Thin kernels | −0.78 *** | 0.33 ** | ||||
| Malt quality | ||||||
| Malt α-amylase | −0.32 ** | |||||
| Malt diastatic power (DP) | ||||||
| Malt extract | 0.24 * | |||||
| Wort viscosity | −0.58 *** | −0.64 *** | −0.64 *** | −0.61 *** | −0.61 *** | −0.31 *** |
| Wort soluble protein | 0.31 ** | |||||
| Kolbach index | ||||||
| Wort color | 0.24 * | 0.29 * | 0.25 * | 0.25 * | 0.46 *** | |
| Wort free amino nitrogen (FAN) | 0.27 * | 0.28 * | 0.67 ** | |||
| Wort β-glucan | −0.24 * | −0.34 ** | −0.38 ** | −0.31 ** | −0.31 ** | |
| Wort arabinoxylan | −0.27 * | |||||
| Wort ferulic acid | 0.25 * | |||||
| Wort p-coumaric acid | 0.43 *** | 0.70 *** | 0.78 *** | 0.75 *** | 0.75 *** | |
| Wort vanillic acid | 0.31 ** | 0.27 * | 0.28 * | |||
| Wort total phenolic acids | 0.41 *** | 0.51 *** | 0.49 *** | 0.46 *** | 0.41 *** |
| Parameters | ND rye DON (mg/kg) | MN rye DON (mg/kg) | NY rye DON (mg/kg) | ||||
|---|---|---|---|---|---|---|---|
| <0.20 | 0.20–1.37 | <0.50 | 0.50–2.00 | 2.01–10.76 | <2.00 | 2.01–4.66 | |
| No. of samples | 9 | 5 | 44 | 28 | 9 | 11 | 11 |
| Trichothecenes | |||||||
| Rye D3G (mg/kg) | 0.03 a | 0.13 a | <0.50 a | 0.52 b | 1.69 c | <0.50 a | 2.57 b |
| Malt DON (mg/kg) | 1.10 a | 0.89 a | 5.53 a | 12.80 b | 21.08 c | 4.19 a | 35.09 b |
| Malt D3G (mg/kg) | 1.39 a | 0.82 a | 5.34 a | 9.45 b | 11.98 b | 3.78 a | 18.29 b |
| Malt D3G/DON (mol%) | 74.88 a | 79.47 a | 72.85 a | 53.51 b | 37.15 c | 63.64 a | 34.01 b |
| Malt 3-ADON (mg/kg) | <0.20 a | <0.20 a | <0.50 a | 0.75 b | 0.93 b | <0.50 a | 2.56 b |
| Malt 15-ADON (mg/kg) | <0.20 a | <0.20 a | 0.68 a | 1.61 b | 2.28 c | <0.50 a | 2.04 b |
| Malt NIV (mg/kg) | <0.20 a | <0.20 a | 0.26 a | 0.42 b | 1.04 c | <0.50 a | <0.50 a |
| Grain quality | |||||||
| Rye thousand kernel weight (g) | 30.23 a | 29.84 a | 27.29 a | 30.00 b | 33.32 b | 30.70 a | 30.66 a |
| Rye Plump percent (%) | 49.30 a | 59.45 a | 46.08 a | 62.46 b | 77.23 b | 73.57 a | 66.94 a |
| Rye Thin percent (%) | 2.30 a | 6.94 a | 14.97 ab | 7.80 bc | 7.33 c | 1.71 a | 3.46 b |
| Malt quality | |||||||
| Malt α-amylase (DU) | 69.04 a | 70.58 a | 79.97 a | 71.09 b | 66.58 b | 87.80 a | 83.27 a |
| Malt DP (ASBC) | 143.67 a | 118.10 b | 129.91 a | 147.31 ab | 180.02 bc | 51.45 a | 73.25 b |
| Malt extract (%, db) | 83.17 a | 83.63 a | 83.21 a | 83.77 a | 82.48 a | 88.44 a | 88.28 a |
| Wort viscosity (mPa) | 5.54 a | 6.23 a | 4.57 a | 3.66 b | 2.17 c | 2.83 a | 2.36 a |
| Wort soluble protein (% malt, db) | 10.05 a | 8.14 b | 9.62 a | 8.87 a | 10.61 a | 5.60 a | 5.98 a |
| Kolbach index (%) | 67.95 a | 66.00 a | 66.49 a | 63.92 a | 68.18 a | 67.21 a | 70.05 a |
| Wort color (°SRM) | 6.58 a | 3.83 b | 6.79 a | 7.51 a | 7.70 a | 4.85 a | 6.53 b |
| Wort FAN content (mg/L) | 223.42 a | 220.11 a | 255.10 a | 260.79 ab | 311.72 c | 227.56 a | 233.69 a |
| Wort β-glucan content (mg/L) | 81.39 a | 96.24 a | 33.53 a | 28.91 ab | 18.86 b | 15.82 a | 3.99 b |
| Wort arabinoxylan content (mg/L) | 234.18 a | 215.02 a | 251.21 a | 251.88 ab | 202.13 b | 233.57 a | 234.32 a |
| Wort ferulic acid content (mg/L) | 8.51 a | 6.24 b | 9.37 a | 9.55 a | 8.91 a | 11.34 a | 10.33 a |
| Wort p-coumaric acid content (mg/L) | 0.63 a | 0.52 a | 1.67 a | 2.96 bc | 3.09 c | 2.69 a | 3.66 b |
| Wort vanillic acid content (mg/L) | 1.91 a | 1.92 a | 3.60 a | 2.72 a | 5.84 b | 3.70 a | 6.88 b |
| Wort total phenolic acid content (mg/L) | 11.75 a | 9.26 a | 15.76 a | 16.70 ab | 19.58 b | 18.73 a | 22.28 b |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, Z.; Gillespie, J.; Barr, J.; Wiersma, J.J.; Sorrells, M.E.; Zwinger, S.; Gross, T.; Cumming, J.; Bergstrom, G.C.; Brueggeman, R.; et al. Malting of Fusarium Head Blight-Infected Rye (Secale cereale): Growth of Fusarium graminearum, Trichothecene Production, and the Impact on Malt Quality. Toxins 2018, 10, 369. https://doi.org/10.3390/toxins10090369
Jin Z, Gillespie J, Barr J, Wiersma JJ, Sorrells ME, Zwinger S, Gross T, Cumming J, Bergstrom GC, Brueggeman R, et al. Malting of Fusarium Head Blight-Infected Rye (Secale cereale): Growth of Fusarium graminearum, Trichothecene Production, and the Impact on Malt Quality. Toxins. 2018; 10(9):369. https://doi.org/10.3390/toxins10090369
Chicago/Turabian StyleJin, Zhao, James Gillespie, John Barr, Jochum J. Wiersma, Mark E. Sorrells, Steve Zwinger, Thomas Gross, Jaime Cumming, Gary C. Bergstrom, Robert Brueggeman, and et al. 2018. "Malting of Fusarium Head Blight-Infected Rye (Secale cereale): Growth of Fusarium graminearum, Trichothecene Production, and the Impact on Malt Quality" Toxins 10, no. 9: 369. https://doi.org/10.3390/toxins10090369