Hydrogen-Deuterium Exchange Mass Spectrometry Reveals a Novel Binding Region of a Neutralizing Fully Human Monoclonal Antibody to Anthrax Protective Antigen

 ,
,
Abstract
:1. Introduction
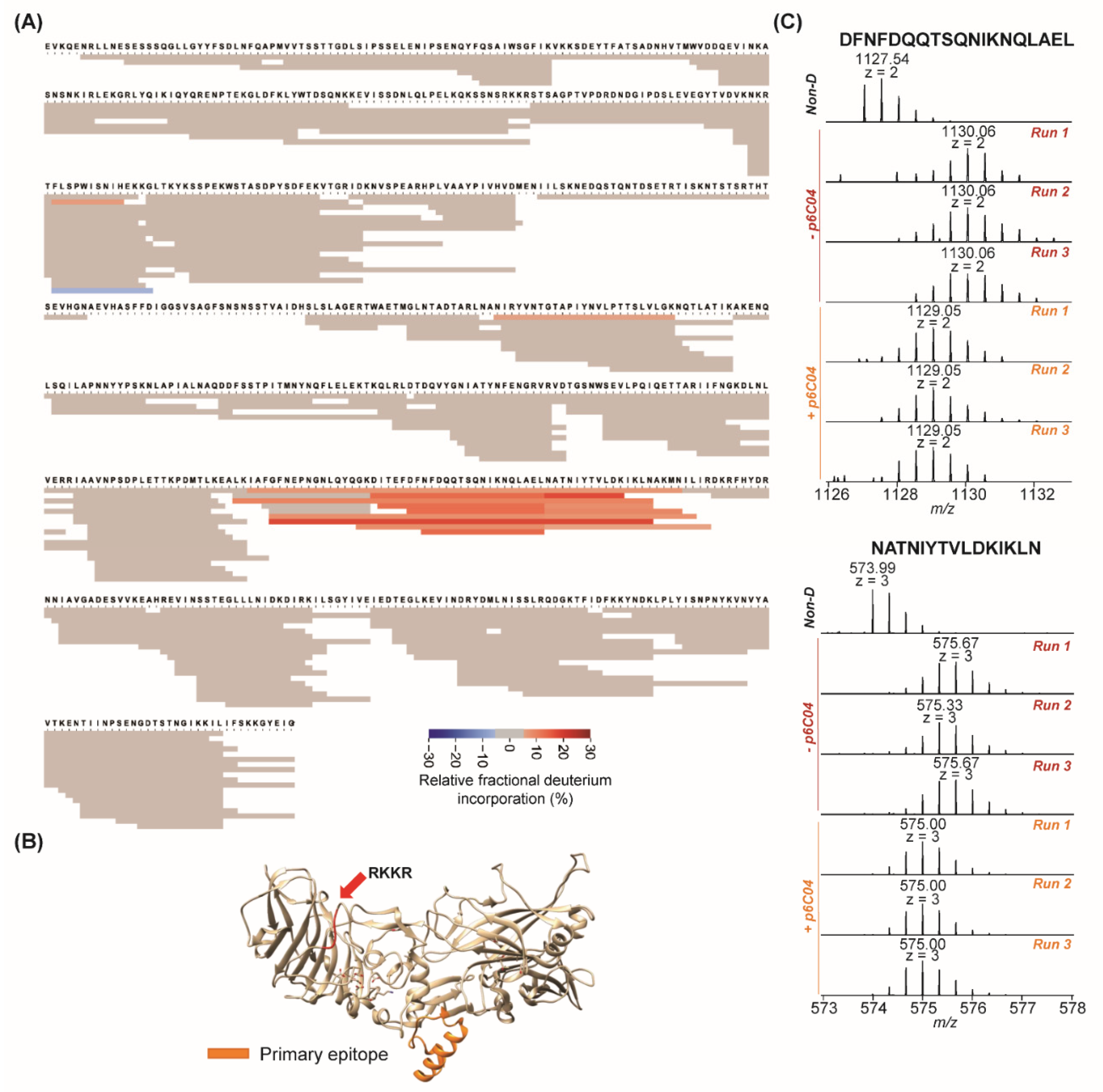
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials and Reagents
5.2. Differential UPLC-HDX-MS
5.3. HDX-MS Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coggeshall, K.M.; Lupu, F.; Ballard, J.; Metcalf, J.P.; James, J.A.; Farris, D.; Kurosawa, S. The sepsis model: An emerging hypothesis for the lethality of inhalation anthrax. J. Cell. Mol. Med. 2013, 17, 914–920. [Google Scholar] [CrossRef]
- Mourez, M. Anthrax toxins. Rev. Physiol. Biochem. Pharmacol. 2004, 152, 135–164. [Google Scholar]
- Scobie, H.M.; Young, J.A. Interactions between anthrax toxin receptors and protective antigen. Curr. Opin. Microbiol. 2005, 8, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Kintzer, A.F.; Sterling, H.J.; Tang, I.I.; Williams, E.R.; Krantz, B.A. Anthrax Toxin Receptor Drives Protective Antigen Oligomerization and Stabilizes the Heptameric and Octameric Oligomer by a Similar Mechanism. PLoS ONE 2010, 5, e13888. [Google Scholar] [CrossRef] [Green Version]
- Klimpel, K.R.; Molloy, S.S.; Thomas, G.; Leppla, S.H. Anthrax toxin protective antigen is activated by a cell surface protease with the sequence specificity and catalytic properties of furin. Proc. Natl. Acad. Sci. USA 1992, 89, 10277–10281. [Google Scholar] [CrossRef] [Green Version]
- Reason, D.; Liberato, J.; Sun, J.; Camacho, J.; Zhou, J. Mechanism of Lethal Toxin Neutralization by a Human Monoclonal Antibody Specific for the PA20 Region of Bacillus anthracis Protective Antigen. Toxins 2011, 3, 979–990. [Google Scholar] [CrossRef] [Green Version]
- Reason, D.; Liberato, J.; Sun, J.; Keitel, W.; Zhou, J. Frequency and Domain Specificity of Toxin-Neutralizing Paratopes in the Human Antibody Response to Anthrax Vaccine Adsorbed. Infect. Immun. 2009, 77, 2030–2035. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Ruther, P.; Jiang, I.; Sawada-Hirai, R.; Sun, S.M.; Nedellec, R.; Morrow, P.R.; Kang, A.S. Human monoclonal antibodies that neutralize anthrax toxin by inhibiting heptamer assembly. Hum. Antibodies 2005, 13, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.; Crowe, S.R.; Garman, L.; Guthridge, C.J.; Muther, J.J.; McKee, E.; Zheng, N.-Y.; Farris, A.D.; Guthridge, J.M.; Wilson, P.C.; et al. Human monoclonal antibodies generated following vaccination with AVA provide neutralization by blocking furin cleavage but not by preventing oligomerization. Vaccine 2012, 30, 4276–4283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brossier, F.; Lévy, M.; Landier, A.; Lafaye, P.; Mock, M. Functional Analysis of Bacillus anthracis Protective Antigen by Using Neutralizing Monoclonal Antibodies. Infect. Immun. 2004, 72, 6313–6317. [Google Scholar] [CrossRef] [Green Version]
- Masson, G.R.; Burke, J.E.; Ahn, N.G.; Anand, G.S.; Borchers, C.; Brier, S.; Bou-Assaf, G.M.; Engen, J.R.; Englander, S.W.; Faber, J.; et al. Recommendations for performing, interpreting and reporting hydrogen deuterium exchange mass spectrometry (HDX-MS) experiments. Nat. Methods 2019, 16, 595–602. [Google Scholar] [CrossRef] [Green Version]
- Konermann, L.; Pan, J.; Liu, Y.-H. Hydrogen exchange mass spectrometry for studying protein structure and dynamics. Chem. Soc. Rev. 2011, 40, 1224–1234. [Google Scholar] [CrossRef]
- Zhu, S.; Liuni, P.; Ettorre, L.; Chen, T.; Szeto, J.; Carpick, B.; James, D.A.; Wilson, D.J. Hydrogen-Deuterium Exchange Epitope Mapping Reveals Distinct Neutralizing Mechanisms for Two Mono-clonal Antibodies against Diphtheria Toxin. Biochemistry 2019, 58, 646–656. [Google Scholar] [CrossRef]
- Grauslund, L.R.; Calvaresi, V.; Pansegrau, W.; Norais, N.; Rand, K.D. Epitope and Paratope Mapping by HDX-MS Combined with SPR Elucidates the Difference in Bacteri-cidal Activity of Two Anti-NadA Monoclonal Antibodies. J. Am. Soc. Mass Spectrom. 2021, 32, 1575–1582. [Google Scholar] [CrossRef]
- Zheng, J.; Wang, C.; Chang, M.R.; Devarkar, S.C.; Schweibenz, B.; Crynen, G.; Garcia-Ordonez, R.D.; Pascal, B.D.; Novick, S.J.; Patel, S.S.; et al. HDX-MS reveals dysregulated checkpoints that compromise discrimination against self RNA during RIG-I mediated autoimmunity. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Malito, E.; Biancucci, M.; Faleri, A.; Ferlenghi, I.; Scarselli, M.; Maruggi, G.; Surdo, P.L.; Veggi, D.; Liguori, A.; Santini, L.; et al. Structure of the meningococcal vaccine antigen NadA and epitope mapping of a bactericidal antibody. Proc. Natl. Acad. Sci. USA 2014, 111, 17128–17133. [Google Scholar] [CrossRef] [Green Version]
- Trabjerg, E.; Nazari, Z.E.; Rand, K.D. Conformational analysis of complex protein states by hydrogen/deuterium exchange mass spectrometry (HDX-MS): Challenges and emerging solutions. TrAC Trends Anal. Chem. 2018, 106, 125–138. [Google Scholar] [CrossRef]
- Fang, M.; Wang, Z.; Cupp-Sutton, K.A.; Welborn, T.; Smith, K.; Wu, S. High-throughput hydrogen deuterium exchange mass spectrometry (HDX-MS) coupled with sub-zero-temperature ultrahigh pressure liquid chromatography (UPLC) separation for complex sample analysis. Anal. Chim. Acta 2021, 1143, 65–72. [Google Scholar] [CrossRef]
- Smith, K.; Garman, L.; Wrammert, J.; Zheng, N.-Y.; Capra, J.D.; Ahmed, R.; Wilson, P.C. Rapid generation of fully human monoclonal antibodies specific to a vaccinating antigen. Nat. Protoc. 2009, 4, 372–384. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Pevzner, P.A. MS-GF+ makes progress towards a universal database search tool for proteomics. Nat. Commun. 2014, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Walters, B.T.; Ricciuti, A.; Mayne, L.; Englander, S.W. Minimizing Back Exchange in the Hydrogen Exchange-Mass Spectrometry Experiment. J. Am. Soc. Mass Spectrom. 2012, 23, 2132–2139. [Google Scholar] [CrossRef] [Green Version]
- Mayne, L. Hydrogen Exchange Mass Spectrometry. Methods Enzymol. 2016, 566, 335–356. [Google Scholar]
- Narang, D.; James, D.A.; Balmer, M.T.; Wilson, D.J. Protein Footprinting, Conformational Dynamics, and Core Interface-Adjacent Neutralization "Hotspots" in the SARS-CoV-2 Spike Protein Receptor Binding Domain/Human ACE2 Interaction. J. Am. Soc. Mass Spectrom. 2021, 32, 1593–1600. [Google Scholar] [CrossRef]
- Hansen, J.; Baum, A.; Pascal, K.E.; Russo, V.; Giordano, S.; Wloga, E.; Fulton, B.O.; Yan, Y.; Koon, K.; Patel, K.; et al. Studies in humanized mice and convalescent humans yield a SARS-CoV-2 antibody cocktail. Science 2020, 369, 1010–1014. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, J.; Bautista, J.; Badithe, A.; Olson, W.; Liu, Y. Epitope Mapping by HDX-MS Elucidates the Surface Coverage of Antigens Associated with High Blocking Efficiency of Antibodies to Birch Pollen Allergen. Anal. Chem. 2018, 90, 11315–11323. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | In Vivo Neutralization (% A/JMouse Survival after Toxin Challenge) * | Domain Reactivity (ELISA) * | Functional * | Epitope(s) |
|---|---|---|---|---|
| p6C04 | 0 | 3 | Oligomerization | DITEFDFNFDQQTSQNIKNQLAELNAT-NIYTVLDKIKLNAKMN |
| p1C03 | 40 | 1A | -- | LSIPSSELENIPSEN; IQYQRENPTEKGLDFKLLSIPSSELENIPSEN |
| p1A06 | 20 | 3 | Oligomerization | DITEFDFNFDQQTSQNIKNQLAELNAT-NIYTVLDKIKLNAKMN |
| p6C01 | 80 | Whole PA | Furin cleavage | AVNPSDPLETTKPDMTL (in domain 3)TDQVYGNIATYN (in domain 2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, M.; Wang, Z.; Norris, K.; James, J.A.; Wu, S.; Smith, K. Hydrogen-Deuterium Exchange Mass Spectrometry Reveals a Novel Binding Region of a Neutralizing Fully Human Monoclonal Antibody to Anthrax Protective Antigen. Toxins 2022, 14, 92. https://doi.org/10.3390/toxins14020092
Fang M, Wang Z, Norris K, James JA, Wu S, Smith K. Hydrogen-Deuterium Exchange Mass Spectrometry Reveals a Novel Binding Region of a Neutralizing Fully Human Monoclonal Antibody to Anthrax Protective Antigen. Toxins. 2022; 14(2):92. https://doi.org/10.3390/toxins14020092
Chicago/Turabian StyleFang, Mulin, Zhe Wang, Kathleen Norris, Judith A. James, Si Wu, and Kenneth Smith. 2022. "Hydrogen-Deuterium Exchange Mass Spectrometry Reveals a Novel Binding Region of a Neutralizing Fully Human Monoclonal Antibody to Anthrax Protective Antigen" Toxins 14, no. 2: 92. https://doi.org/10.3390/toxins14020092