Genotypes of Staphylococcus aureus Clinical Isolates Are Associated with Phenol-Soluble Modulin (PSM) Production


1
Department of Laboratory Medicine, Kangdong Sacred Heart Hospital, Hallym University College of Medicine, Seoul 05355, Korea
2
Department of Biotechnology, College of Engineering, Duksung Women’s University, Seoul 01369, Korea
*
Authors to whom correspondence should be addressed.
Toxins 2022, 14(8), 556; https://doi.org/10.3390/toxins14080556
Submission received: 15 July 2022
/
Revised: 6 August 2022
/
Accepted: 12 August 2022
/
Published: 15 August 2022
(This article belongs to the Collection Staphylococcus aureus Toxins)
Abstract
:Phenol-soluble modulins (PSMs) are important S. aureus virulence factors that cause cytolysis, mast cell degranulation, and stimulate inflammatory responses. In this study, PSM production by S. aureus clinical isolates was measured by liquid chromatography/mass spectrometry (LC-MS) and correlated with staphylococcal protein A (spa) type and staphylococcal cassette chromosome mec (SCCmec) type. Of 106 S. aureus clinical isolates, 50 (47.2%) corresponded to methicillin-susceptible S. aureus (MSSA) and 56 (52.8%) to methicillin-resistant S. aureus (MRSA). LC-MS analysis revealed no significant difference in average PSMα3, PSMα4, PSMβ2, and δ-toxin production between MSSA and MRSA isolates, but PSMα1, PSMα2, and PSMβ1 production were higher in MSSA than MRSA. This study demonstrated that average PSMα1–α4, PSMβ1–β2, and δ-toxin production by SCCmec type II strains was significantly lower than the IV, IVA, and V strains. Most of the SCCmec type II strains (n = 17/25; 68.0%) did not produce δ-toxin, suggesting a dysfunctional Agr system. The spa type t111 (except one strain) and t2460 (except one strain producing PSM α1–α4) did not produce PSMα1–α4 and δ-toxin, while average PSM production was higher among the t126 and t1784 strains. This study showed that the genotype of S. aureus, specifically the spa and SCCmec types, is important in characterizing the production of PSMs.
Keywords:
Staphylococcus aureus; MRSA; MSSA; mass spectrometry; PSMs; delta-toxin; spa type; SCCmec typeKey Contribution: MRSA SCCmec type II strain produced significantly lower levels of PSMs than type IV, IVA, and V strains. The spa type t126 and t1784 strains produced the highest amounts of PSMα1–α4, PSMβ1, and δ-toxin, whereas t2460 strains did not produce PSMα1–α4 and δ-toxin, except for one strain that produced low PSMα1–α4.
1. Introduction
The Gram-positive bacterium Staphylococcus aureus (S. aureus) is one of the most frequent causes of both hospital-associated (HA) and community-associated (CA) infections [1,2]. S. aureus infections range from skin and soft tissue infections to severe invasive diseases such as osteomyelitis, pneumonia, and bacteremia [3]. The success of S. aureus in the human host can be attributed to its ability to produce a wide variety of virulence factors that damage the host and evade immunity [4]. S. aureus can produce different virulence factors, such as enterotoxins, alpha toxins, toxic shock syndrome toxins, and phenol-soluble modulins (PSMs) [5]. PSMs, which include δ-toxin, are among the most abundant peptides in an overnight culture and can account for 60% of the total protein produced. The PSM peptide family plays a key role in S. aureus pathogenesis [6,7]. PSM production is regulated by the accessory gene regulator (Agr) quorum-sensing system [8,9,10], which involves the direct binding of response regulator AgrA to psm operon promoters [11].
PSMs are a group of amphipathic α-helical peptides that include PSMα1–PSMα4 (~20–25 amino acids), PSMβ1 and PSMβ2 (~43–45 amino acids), and δ-toxin (~26 amino acids) [6,12,13,14]. PSMs have distinct virulence functions and lyse a variety of eukaryotic cells, including neutrophils, and stimulate inflammatory responses [14,15]. PSMs also contribute to S. aureus pathogenicity in skin and bloodstream infections [7,16]. Compared to other PSMs, δ-toxin is usually a more strongly produced peptide that is more cytolytic to neutrophils and has a moderate capacity to stimulate formyl-peptide receptor 2 (FPR2) [6,17]. Furthermore, PSMs form bacterial functional amyloids [18] that are believed to contribute to biofilm structuring, detachment, and the systemic dissemination of biofilm-associated infection [19,20].
Members of the PSM peptide family are secreted as their primary N-formylated translation products by dedicated ABC exporter systems [21]. PSMs are encoded at three different locations in the S. aureus genome [6]. PSMα1–α4 are encoded by the psmα operon, PSMβ1–β2 are encoded by the psmβ operon, and δ-toxin is encoded by the hld loci. The hld gene is embedded within RNAIII, the RNA effector molecule of the Agr system [14]. Due to the well-known association between the Agr system and δ-toxin production, detection of δ-toxin can be used as evidence of a functional Agr system [22,23]. An additional PSM peptide, PSM-mec, is encoded by specific SCCmec elements (types II, III) of methicillin-resistant S. aureus (MRSA) and is also regulated by Agr [24,25]. Although psm genes are present in all S. aureus genomes sequenced, their expression may differ significantly [6,26]. The increased virulence of the USA300/USA500 sublineage has been attributed to the differential expression of core genome-encoded PSMs [26]. A recent study characterized variation in δ-toxin production across S. aureus strains and identified genetic loci associated with differences between strains [27]; however, there is little information on PSMα1–α4 and PSMβ1–β2 production by S. aureus strains and its association with genotypes.
An association has been reported between PSM production by MRSA and MSSA isolates in vitro and their clinical source of isolation, i.e., skin and soft tissue infection (SSTI), hospital-acquired pneumonia (HAP), and infective endocarditis (IE) [28,29]. MRSA and MSSA isolate from patients with SSTI produced higher levels of PSMα1–α4, PSMβ1, and δ-toxin than HAP or IE isolates [28,29]. However, the impact of S. aureus genotype differences (e.g., S. aureus protein A (spa) type and SCCmec type) on PSM production is largely unknown. Here, irrespective of isolates from multiple infection sites, we performed high-throughput liquid chromatography/mass spectrometry (LC-MS) analysis of 106 S. aureus blood culture isolates to measure in vitro PSM production. We then correlated the PSM production with S. aureus genotypes, specifically the spa and SCCmec types. Moreover, we focused on the clonal lineages or SCCmec types of MRSA and their association with PSM production.
2. Results
2.1. Molecular Characteristics of S. aureus Isolates
Among the 106 S. aureus isolates, 50 (47.2%) were phenotypically MSSA and 56 (52.8%) were MRSA. The spa typing discriminated S. aureus isolates into 47 types, with t189, t2460, t008, t126, t324, and t1784 as the most prevalent (Table 1). MSSA strains were distributed among 31 spa types, with MRSA in 21 spa types. SCCmec II (n = 25) and IV (n = 18) strains were the most predominant, followed by type IVA (n = 10) and V (n = 3).
2.2. In Vitro δ-Toxin Production by S. aureus Clinical Isolates
δ-toxin production is considered to be a surrogate marker of the functional Agr system in S. aureus [23] and dysfunction of Agr was defined as the absence of δ-toxin production [22]. In this study, δ-toxin production by S. aureus blood culture isolates was measured by LC-MS. We found δ-toxin production (sum of the formylated and deformylated δ-toxin) in 80 (75.5%) S. aureus isolates, of which 43 (86.0%) were MSSA and 37 (66.1%) were MRSA isolates (Tables S1 and S2). Of these, three MSSA and five MRSA SCCmec type IV strains were δ-toxin allelic variants. All δ-toxin-deficient MRSA (n = 19/56, 33.9%) belonged to SCCmec type II (n = 17/25, 68.0%) and type IVA (n = 2/10, 20.0%).
2.3. Association between In Vitro PSM Production, Methicillin-Resistance, SCCmec Type, and spa Type
The extent of PSM production in vitro was measured by LC-MS in each of the S. aureus blood culture isolates of different methicillin-resistance phenotypes, SCCmec type, and spa type. Within our set of isolates, no significant difference was found in average PSMα3 (1.69 μM vs. 1.11 μM), PSMα4 (1.46 μM vs. 1.05 μM), PSMβ2 (1.01 μM vs. 0.82 μM), and δ-toxin (6.81 μM vs. 5.39 μM) production between MSSA (n = 50) and MRSA (n = 56) strains by Kruskal–Wallis test, p > 0.05 (Figure 1A; Table S3). However, PSMα1 (1.22 μM vs. 0.88 μM), PSMα2 (0.92 μM vs. 0.63 μM), and PSMβ1 (1.92 μM vs. 1.43 μM) production were higher in MSSA than MRSA isolates. We did not compare the PSM-mec production of S. aureus isolates because its gene is located on the methicillin-resistance cassette, which MSSA lacks.
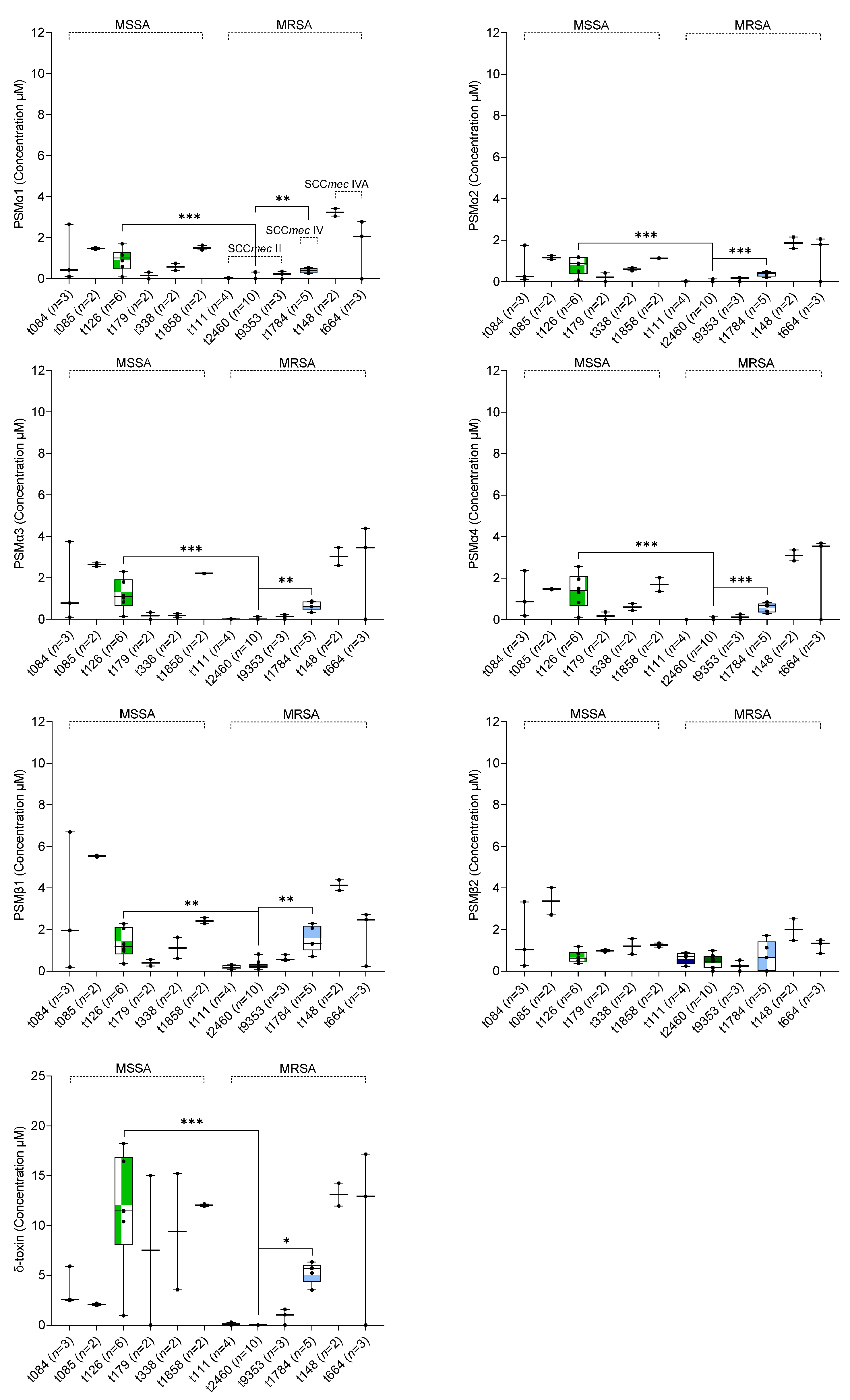
When average PSM production was compared across SCCmec type strains of MRSA using the separate Kruskal–Wallis test, SCCmec type IV (n = 18), IVA (n = 10), and V (n = 3) had significantly higher levels of PSMα1–α4, PSMβ1–β2, and δ-toxin production than SCCmec type II (n = 25) (p < 0.01) (Figure 1B; Table S4), despite each of these SCCmec types comprising a wide variety of spa types. Within SCCmec type II strains, spa type t111 (n = 3) and t2460 (n = 10) did not produce PSMα1-α4 and δ-toxin, except one t2460 strain produced lower amounts of PSMα1 (0.33 μM), PSMα2 (0.13 μM), PSMα3 (0.13 μM), and PSMα4 (0.13 μM). Alternatively, spa type t002 (n = 3) and t9353 (n = 3) of SCCmec type II produced intermediate quantities of all PSMs.
When variation between similar spa types of MSSA and MRSA t002 (1 MSSA and 3 MRSA), t008 (1 MSSA and 6 MRSA), t189 (7 MSSA and 4 MRSA), t304 (1 MSSA and 2 MRSA), and t324 (2 MSSA and 4 MRSA) was analyzed using the separate Kruskal–Wallis test, no significant difference in average PSM production was found (p > 0.05) (Figure 2; Table S5). However, the number of similar spa-type strains in each phenotypic group may be too small to reliably establish an association with PSM production.
When variation between the remaining different spa types of S. aureus irrespective of methicillin-resistance was analyzed using the pairwise Kruskal–Wallis test, t126 (n = 6) and t1784 (n = 5) were found to have significantly higher levels of PSMα1–α4, PSMβ1, and δ-toxin production than t2460 (n = 10, all δ-toxin deficient) (p < 0.05) (Figure 3; Table S6). Furthermore, three spa type t111 strains did not produce PSMα1–α4 and δ-toxin. Interestingly, all spa type t111 and t2460 strains showed a small amount of PSMβ1–β2 production (Figure 3). However, no significant difference in PSMβ2 production across different spa types of MSSA and MRSA isolates was observed (p > 0.05) (Figure 3). An association between the remaining spa types (other than t126, t1784, and t2460) and the production of PSMs could not be established due to the limited number of strains for each spa type.
3. Discussion
This study measured PSM production in S. aureus clinical isolates in vitro and revealed its association with strain genotypes, specifically the spa type and SCCmec type. The prevalence and molecular characteristics of the S. aureus isolates obtained from blood cultures of patients in a hospital showed that spa types t189 and t126 were dominant among MSSA isolates, while t2460, t008, and t1784 were dominant among MRSA isolates. Most of the MRSA isolates belonged to SCCmec type II (n = 25/56, 44.6%) and IV (n = 18/56, 32.1%). The predominance of MRSA isolates with these SCCmec types in this study is consistent with a previous study in Korea showing the abundance of SCCmec type II (n = 282/407, 69.2%) and IV (n = 97/407, 23.8%) strains [30]. Several studies have characterized S. aureus isolates from individual hospitals and found certain genotype strains that appear to be well adapted to the hospital environment as most prevalent [31,32,33,34,35].
The Agr system is of major importance in staphylococcal pathogenesis due to its role in the regulation of PSM production [8,9,10,14], and lack of hemolysin production therefore generally represents a dysfunctional Agr [36]. In this study, 43 (86.0%) MSSA and 37 (66.1%) MRSA isolates possessed a functional Agr system as demonstrated by semiquantitative measurement of δ-toxin production using LC-MS (Tables S1 and S2). Strikingly, most of the δ-toxin deficient MRSA strains were SCCmec type II (n = 17/19, 89.4%), with the spa type t2460 strains (n = 10/17, 58.8%) as predominant. We found that SCCmec type II was a significant negative predictor of δ-toxin production and thereby dysfunctional Agr system. A previous study measured δ-hemolysin activity to determine the Agr functionality and showed that SCCmec type II isolates which are prevalent in hospital-acquired infections in Korea are mostly Agr dysfunctional (n = 274/282, 90.7%) [37]. The agrA or agrC mutations were known as the main cause of Agr dysfunction in S. aureus clinical isolates [36,37,38,39]. Among MRSA isolates, Agr dysfunction is associated with a trend toward persistent bacteremia [37]. Furthermore, we found five SCCmec type IV strains of spa type t1784 as δ-toxin allelic variants. The amino acid sequences of δ-toxin and its allelic variant are the same with the exception at 10 (G10S) [40], due to which their molecular weights are different and can be distinguished by mass spectrometry. δ-Toxin allelic variants with highly similar peptide sequences are often present in many staphylococcal species [41].
Limited information is available about S. aureus PSM production and its association with specific genotypes [28,29]. We found that δ-toxin was the most strongly produced peptide, whereas low to moderate levels of PSMα1–α4 and PSMβ1–β2 were produced by S. aureus clinical isolates under in vitro conditions (Figure 1A). PSMs are known to be produced by all S. aureus strains (except naturally occurring agr mutants) due to the location of encoding genes on the core genome or pathogenicity islands [42], but the expression pattern may differ among them [14]. In our S. aureus blood culture isolates, no significant difference was observed in average PSMα3, PSMα4, PSMβ2, and δ-toxin production between MSSA and MRSA strains, but MSSA strains produced statistically higher levels of PSMα1 (p < 0.04), PSMα2 (p < 0.03), and PSMβ1 than MRSA (p < 0.05). Previous studies showed that the level of PSM production in S. aureus correlates more closely with isolation source, i.e., with the patient disease rather than with methicillin-resistance [28,29]. Another study showed that MSSA strains had significantly higher δ-toxin production than MRSA strains [27]. This difference could be due to either variation in the number of MSSA (n = 86) and MRSA (n = 38) isolates, genotypic background (Agr type and CC), or the continent of origin [27]. Indeed, we found extremely low levels of PSMα1–α4, PSMβ1–β2, and δ-toxin across SCCmec type II strains with nearly all spa type t111 (n = 3) and t2460 (n = 10) did not produce PSMα1–α4 and δ-toxin (except one t2460 strain produced low PSMα1–α4), whereas t002 and t9353 (each n = 3) strains of the same SCCmec type also produced low quantities of all PSMs.
PSM production is mainly controlled by the Agr system and strains with genetic changes leading to impaired Agr activity may be associated with decreased PSM production. A slipped-mispairing mutation in agrA of S. aureus clinical isolates results in delayed activation of Agr and failure to translate α- and δ-hemolysins [36]. The dysfunction of Agr was common among MRSA SCCmec type II bloodstream isolates in Korea [37]. Moreover, the transcription and translation products of PSM-mec, which is encoded by mobile genetic elements in SCCmec type II and III hospital-acquired MRSA strains have been reported to suppress the production of PSMα1–α4 [43,44]. We did not detect significant differences in average PSM production between SCCmec type IV, IVA, and V strains. Finally, we observed differences in average PSM production between spa types, with t126 and t1784 strains producing significantly higher levels of PSMα1–α4, PSMβ1, and δ-toxin than other spa types.
Our study has some limitations. First, S. aureus isolates obtained from blood cultures were only included in this study and not from multiple infection sites including SSTI, pneumonia, and surgical site infections; however, PSMs are known to be produced by all S. aureus strains except agr mutants [42]. Second, PSM production was measured in vitro, but the production of these peptides by the same S. aureus isolates in clinical infection may vary. Third, the significant number of singleton spa type strains in this study prevented meaningful associations between some spa types and PSM production; however, specific regions or even individual hospitals may often show a small number of particular genotype strains. Fourth, while our results do identify an association between average PSM production in vitro and some S. aureus genotypes (SCCmec type II, spa type t2460, t126), we do not establish the molecular basis for these associations; however, agrA or agrC mutations are known as the main cause of Agr dysfunction [36,37,38,39], which thereby suppress the PSM production including δ-toxin. Despite these limitations, our study provided quantitative data on PSM production by certain S. aureus genotypes, specifically spa type t111 and t2460 and SCCmec type II that would suggest low or deficient PSM production by other clinical isolates of the same genotypes, and this may inform anti-virulence strategies to treat infections caused by these genotype strains.
4. Conclusions
Mass spectrometric quantification showed that the production of PSMs can vary greatly depending on certain spa types and SCCmec types of S. aureus isolates. Our results suggest that S. aureus SCCmec type II strains with spa type t111 and t2460 are associated with deficient PSMα1–α4 and δ-toxin production, except for one strain of each spa. Further, S. aureus spa type t126 and t1784 strains are associated with higher levels of PSM production than other spa types. Indeed, this study quantitatively measured PSM production of S. aureus clinical isolates and revealed that it was associated with certain genotype strains.
5. Materials and Methods
5.1. Bacterial Strains and Growth Conditions
A total of 106 S. aureus isolates obtained from blood cultures of patients at Kangdong Sacred Heart Hospital, Seoul, Korea from 2020 to 2021 were included in this study. All the isolates were identified by the matrix-assisted laser desorption ionization/time-of-flight mass spectrometry (MALDI-TOF MS) system (Bruker Microflex LT, Bruker Daltonik GmbH, Bremen, Germany). The presence of a gene encoding methicillin-resistance was confirmed by polymerase chain reaction (PCR) screening for mecA gene [45]. Stock cultures of isolates were stored frozen (−70 °C) in skimmed milk.
5.2. The spa Typing and SCCmec Typing
The spa typing of S. aureus isolates was performed as described previously [46]. A single locus of the repeat region X of the spa gene was sequenced and analyzed using Ridom StaphType software (http://spaserver.ridom.de/, accessed on 13 September 2021). The spa types were assigned using the BioNumerics software v.7.5 (Applied Math, Sint-Martens-Latem, Belgium). All MRSA strains were subjected to SCCmec typing by discriminating the mec gene complex and the cassette chromosome recombinases (ccr) gene complex types [47].
5.3. PSM Quantification by LC–MS
Quantification of PSM production in the culture supernatants was performed using LC-MS as described previously [48]. Briefly, S. aureus strains were grown in 200 μL of TSB in a 96-well microtiter plate (Corning 3596, NY) for 20 h at 37 °C with 200 rpm shaking. Cultures were pelleted at 3100× g for 20 min at 4 °C, and supernatants were used for PSM quantification by LC-MS.
Aliquots (5 μL) of harvested supernatants were eluted from a C8 column (ZORBAX SB-C8, 2.1 × 5 mm, 1.8 µm; Agilent, Santa Clara, CA, USA) on a Waters ZQ 2000 LC-MS system (Waters, Milford, MA) with a gradient of trifluoroacetic acid (TFA; 0.05%) in water and 0.05% TFA in acetonitrile at a 0.3 mL/min flow rate. Electrospray ionization of samples was performed at 3.5 kV and ions were infused into the ion entrance of a mass spectrometer. The m/z values of the analytes were scanned continuously, and mass spectra were recorded. The m/z values of 2+ and 3+ charged ions of α-type PSMs and 3+ and 4+ charged ions of β-type PSMs were used to extract chromatograms for quantification of each PSM. The peptides were quantified by the sum of extracted ion chromatograms of formylated and deformylated forms. The concentration of PSMs was determined by external calibration with synthetic formyl PSMs. Formyl PSM peptides were synthesized by Peptron (Daejeon, Korea) and Cosmogenetech (Daejeon, Korea). The cultivation and mass analysis were performed in duplicate for each isolate.
5.4. Statistical Analysis
Statistical analyses were performed with SPSS software version 24.0 (SPSS Inc., IBM, Chicago, IL, USA). PSM production by each isolate was quantified in duplicate and averaged. The PSM values were compared using the Kruskal–Wallis tests and differences in production across methicillin-resistance, SCCmec type, and spa type were considered statistically significant if * p < 0.05, ** p < 0.01, and *** p < 0.001. The plots were created using GraphPad Prism software version 9.3.0 (GraphPad Software Inc., San Diego, CA, USA).
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/toxins14080556/s1: Table S1. Genotypic characteristics and δ-toxin production of MSSA clinical isolates; Table S2. Genotypic characteristics and δ-toxin production of MRSA clinical isolates; Table S3. Variation in PSM production between MSSA and MRSA strains; Table S4. Variation in PSM production between SCCmec type strains of MRSA; Table S5. Variation in PSM production between similar spa type strains of MSSA and MRSA; Table S6. PSM production by different spa type strains of S. aureus clinical isolates; Figure S1. δ-toxin deficient MSSA (n = 7) and MRSA (n = 19) strains.
Author Contributions
H.-S.J. and J.-S.K. conceived and designed the experiments, analyzed the data, reviewed drafts of the paper, and approved the final draft. H.L. performed the experiments, analyzed the data, prepared figures and/or tables, and authored drafts of the paper. S.H.C. and Y.L. performed the experiments. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Bio and Medical Technology Development Program of the National Research Foundation (NRF) funded by the Korean government, Ministry of Science and ICT (MSIT) 2017M3A9E4077232. This study was also supported by the Hallym University research fund.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article and Supplementary Material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lowy, F.D. Staphylococcus aureus Infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- David, M.Z.; Daum, R.S. Community-associated methicillin-resistant Staphylococcus aureus: Epidemiology and clinical consequences of an emerging epidemic. Clin. Microbiol. Rev. 2010, 23, 616–687. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, D.; Harper, L.; Shopsin, B.; Torres, V.J. Staphylococcus aureus pathogenesis in diverse host environments. Pathog. Dis. 2017, 75, ftx005. [Google Scholar] [CrossRef]
- Tam, K.; Torres, V.J. Staphylococcus aureus Secreted Toxins and Extracellular Enzymes. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Wang, R.; Braughton, K.R.; Kretschmer, D.; Bach, T.-H.L.; Queck, S.Y.; Li, M.; Kennedy, A.D.; Dorward, D.W.; Klebanoff, S.J.; Peschel, A.; et al. Identification of novel cytolytic peptides as key virulence determinants for community-associated MRSA. Nat. Med. 2007, 13, 1510–1514. [Google Scholar] [CrossRef]
- Kobayashi, S.D.; Malachowa, N.; Whitney, A.R.; Braughton, K.R.; Gardner, D.J.; Long, D.; Wardenburg, J.B.; Schneewind, O.; Otto, M.; DeLeo, F.R. Comparative Analysis of USA300 Virulence Determinants in a Rabbit Model of Skin and Soft Tissue Infection. J. Infect. Dis. 2011, 204, 937–941. [Google Scholar] [CrossRef]
- Yarwood, J.M.; Schlievert, P.M. Quorum sensing in Staphylococcus infections. J. Clin. Investig. 2003, 112, 1620–1625. [Google Scholar] [CrossRef]
- Novick, R.P. Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol. Microbiol. 2003, 48, 1429–1449. [Google Scholar] [CrossRef]
- Bronner, S.; Monteil, H.; Prévost, G. Regulation of virulence determinants in Staphylococcus aureus: Complexity and applications. FEMS Microbiol. Rev. 2004, 28, 183–200. [Google Scholar] [CrossRef]
- Queck, S.Y.; Jameson-Lee, M.; Villaruz, A.E.; Bach, T.H.L.; Khan, B.A.; Sturdevant, D.E.; Ricklefs, S.M.; Li, M.; Otto, M. RNAIII-Independent Target Gene Control by the agr Quorum-Sensing System: Insight into the Evolution of Virulence Regulation in Staphylococcus aureus. Mol. Cell 2008, 32, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.S.; Otto, M. How can Staphylococcus aureus phenol-soluble modulins be targeted to inhibit infection? Future Microbiol. 2013, 8, 693–696. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Phenol-soluble modulins. Int. J. Med. Microbiol. 2014, 304, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Peschel, A.; Otto, M. Phenol-soluble modulins and staphylococcal infection. Nat. Rev. Microbiol. 2013, 11, 667–673. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Cheung, G.Y.C.; Rigby, K.M.; Kamenyeva, O.; Kabat, J.; Sturdevant, D.E.; Villaruz, A.E.; Liu, R.; Piewngam, P.; Porter, A.R.; et al. Rapid pathogen-specific recruitment of immune effector cells in the skin by secreted toxins. Nat. Microbiol. 2021, 7, 62–72. [Google Scholar] [CrossRef]
- Fogel, L.A.; Bubeck Wardenburg, J. Staphylococcus aureus PSMs are a double-edged sword. Nat. Microbiol. 2021, 7, 12–13. [Google Scholar] [CrossRef]
- Kretschmer, D.; Gleske, A.-K.; Rautenberg, M.; Wang, R.; Köberle, M.; Bohn, E.; Schöneberg, T.; Rabiet, M.-J.; Boulay, F.; Klebanoff, S.J.; et al. Human Formyl Peptide Receptor 2 Senses Highly Pathogenic Staphylococcus aureus. Cell Host Microbe 2010, 7, 463–473. [Google Scholar] [CrossRef]
- Schwartz, K.; Syed, A.K.; Stephenson, R.E.; Rickard, A.H.; Boles, B.R. Functional Amyloids Composed of Phenol Soluble Modulins Stabilize Staphylococcus aureus Biofilms. PLoS Pathog. 2012, 8, e1002744. [Google Scholar] [CrossRef]
- Periasamy, S.; Joo, H.-S.; Duong, A.C.; Bach, T.-H.L.; Tan, V.Y.; Chatterjee, S.S.; Cheung, G.Y.C.; Otto, M. How Staphylococcus aureus biofilms develop their characteristic structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1281–1286. [Google Scholar] [CrossRef]
- Wang, R.; Khan, B.A.; Cheung, G.Y.C.; Bach, T.-H.L.; Jameson-Lee, M.; Kong, K.-F.; Queck, S.Y.; Otto, M. Staphylococcus epidermidis surfactant peptides promote biofilm maturation and dissemination of biofilm-associated infection in mice. J. Clin. Investig. 2011, 121, 238–248. [Google Scholar] [CrossRef]
- Chatterjee, S.S.; Joo, H.S.; Duong, A.C.; Dieringer, T.D.; Tan, V.Y.; Song, Y.; Fischer, E.R.; Cheung, G.Y.C.; Li, M.; Otto, M. Essential Staphylococcus aureus toxin export system. Nat. Med. 2013, 19, 364–367. [Google Scholar] [CrossRef] [PubMed]
- Gagnaire, J.; Dauwalder, O.; Boisset, S.; Khau, D.; Freydière, A.-M.; Ader, F.; Bes, M.; Lina, G.; Tristan, A.; Reverdy, M.-E.; et al. Detection of Staphylococcus aureus Delta-Toxin Production by Whole-Cell MALDI-TOF Mass Spectrometry. PLoS ONE 2012, 7, e40660. [Google Scholar] [CrossRef] [PubMed]
- Janzon, L.; Löfdahl, S.; Arvidson, S. Identification and nucleotide sequence of the delta-lysin gene, hld, adjacent to the accessory gene regulator (agr) of Staphylococcus aureus. Mol. Gen. Genet. MGG 1989, 219, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Queck, S.Y.; Khan, B.A.; Wang, R.; Bach, T.-H.L.; Kretschmer, D.; Chen, L.; Kreiswirth, B.N.; Peschel, A.; DeLeo, F.R.; Otto, M. Mobile Genetic Element-Encoded Cytolysin Connects Virulence to Methicillin Resistance in MRSA. PLoS Pathog. 2009, 5, e1000533. [Google Scholar] [CrossRef]
- Chatterjee, S.S.; Chen, L.; Joo, H.-S.; Cheung, G.Y.C.; Kreiswirth, B.N.; Otto, M. Distribution and Regulation of the Mobile Genetic Element-Encoded Phenol-Soluble Modulin PSM-mec in Methicillin-Resistant Staphylococcus aureus. PLoS ONE 2011, 6, e28781. [Google Scholar] [CrossRef]
- Li, M.; Diep, B.A.; Villaruz, A.E.; Braughton, K.R.; Jiang, X.; DeLeo, F.R.; Chambers, H.F.; Lu, Y.; Otto, M. Evolution of virulence in epidemic community-associated methicillin-resistant Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2009, 106, 5883–5888. [Google Scholar] [CrossRef]
- Su, M.; Lyles, J.T.; Petit III, R.A.; Peterson, J.; Hargita, M.; Tang, H.; Solis-Lemus, C.; Quave, C.L.; Read, T.D. Genomic analysis of variability in Delta-toxin levels between Staphylococcus aureus strains. PeerJ 2020, 8, e8717. [Google Scholar] [CrossRef]
- Berlon, N.R.; Qi, R.; Sharma-Kuinkel, B.K.; Joo, H.-S.; Park, L.P.; George, D.; Thaden, J.T.; Messina, J.A.; Maskarinec, S.A.; Mueller-Premru, M.; et al. Clinical MRSA isolates from skin and soft tissue infections show increased in vitro production of phenol soluble modulins. J. Infect. 2015, 71, 447–457. [Google Scholar] [CrossRef]
- Qi, R.; Joo, H.-S.; Sharma-Kuinkel, B.; Berlon, N.R.; Park, L.; Fu, C.; Messina, J.A.; Thaden, J.T.; Yan, Q.; Ruffin, F.; et al. Increased in vitro phenol-soluble modulin production is associated with soft tissue infection source in clinical isolates of methicillin-susceptible Staphylococcus aureus. J. Infect. 2016, 72, 302–308. [Google Scholar] [CrossRef]
- Bae, E.; Kim, C.K.; Jang, J.-H.; Sung, H.; Choi, Y.; Kim, M.-N. Impact of Community-Onset Methicillin-Resistant Staphylococcus aureus on Staphylococcus aureus Bacteremia in a Central Korea Veterans Health Service Hospital. Ann. Lab. Med. 2019, 39, 158–166. [Google Scholar] [CrossRef]
- Wang, B.; Xu, Y.; Zhao, H.; Wang, X.; Rao, L.; Guo, Y.; Yi, X.; Hu, L.; Chen, S.; Han, L.; et al. Methicillin-resistant Staphylococcus aureus in China: A multicenter longitudinal study and whole-genome sequencing. Emerg. Microbes Infect. 2022, 11, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Young, B.C.; Wu, C.H.; Charlesworth, J.; Earle, S.; Price, J.R.; Gordon, N.C.; Cole, K.; Dunn, L.; Liu, E.; Oakley, S.; et al. Antimicrobial resistance determinants are associated with Staphylococcus aureus bacteraemia and adaptation to the healthcare environment: A bacterial genome-wide association study. Microb. Genom. 2021, 7, 700. [Google Scholar] [CrossRef] [PubMed]
- Kaku, N.; Sasaki, D.; Ota, K.; Miyazaki, T.; Yanagihara, K. Changing molecular epidemiology and characteristics of MRSA isolated from bloodstream infections: Nationwide surveillance in Japan in 2019. J. Antimicrob. Chemother. 2022, 77, 2130–2141. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-H.; Lee, J.; Jung, J.; Kim, E.S.; Kim, M.J.; Chong, Y.P.; Kim, S.-H.; Lee, S.-O.; Choi, S.-H.; Woo, J.H.; et al. A Longitudinal Study of Adult Patients with Staphylococcus aureus Bacteremia over 11 Years in Korea. J. Korean Med. Sci. 2021, 36, e104. [Google Scholar] [CrossRef] [PubMed]
- Kang, G.S.; Jung, Y.H.; Kim, H.S.; Lee, Y.S.; Park, C.; Lee, K.J.; Cha, J.O. Prevalence of Major Methicillin-Resistant Staphylococcus aureus Clones in Korea Between 2001 and 2008. Ann. Lab. Med. 2016, 36, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Traber, K.; Novick, R. A slipped-mispairing mutation in AgrA of laboratory strains and clinical isolates results in delayed activation of agr and failure to translate δ- and α-haemolysins. Mol. Microbiol. 2006, 59, 1519–1530. [Google Scholar] [CrossRef] [PubMed]
- Chong, Y.P.; Kim, E.S.; Park, S.-J.; Park, K.-H.; Kim, T.; Kim, M.-N.; Kim, S.-H.; Lee, S.-O.; Choi, S.-H.; Woo, J.H.; et al. Accessory Gene Regulator ( agr ) Dysfunction in Staphylococcus aureus Bloodstream Isolates from South Korean Patients. Antimicrob. Agents Chemother. 2013, 57, 1509–1512. [Google Scholar] [CrossRef]
- Traber, K.E.; Lee, E.; Benson, S.; Corrigan, R.; Cantera, M.; Shopsin, B.; Novick, R.P. agr function in clinical Staphylococcus aureus isolates. Microbiology 2008, 154, 2265–2274. [Google Scholar] [CrossRef]
- Painter, K.L.; Krishna, A.; Wigneshweraraj, S.; Edwards, A.M. What role does the quorum-sensing accessory gene regulator system play during Staphylococcus aureus bacteremia? Trends Microbiol. 2014, 22, 676–685. [Google Scholar] [CrossRef]
- Cheung, G.Y.C.; Yeh, A.J.; Kretschmer, D.; Duong, A.C.; Tuffuor, K.; Fu, C.L.; Joo, H.S.; Diep, B.A.; Li, M.; Nakamura, Y.; et al. Functional characteristics of the Staphylococcus aureus δ-toxin allelic variant G10S. Sci. Rep. 2015, 5, 18023. [Google Scholar] [CrossRef]
- Verdon, J.; Girardin, N.; Lacombe, C.; Berjeaud, J.-M.; Héchard, Y. δ-hemolysin, an update on a membrane-interacting peptide. Peptides 2009, 30, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Cheung, G.Y.C.; Joo, H.-S.; Chatterjee, S.S.; Otto, M. Phenol-soluble modulins—Critical determinants of staphylococcal virulence. FEMS Microbiol. Rev. 2014, 38, 698–719. [Google Scholar] [CrossRef]
- Kaito, C.; Saito, Y.; Ikuo, M.; Omae, Y.; Mao, H.; Nagano, G.; Fujiyuki, T.; Numata, S.; Han, X.; Obata, K.; et al. Mobile Genetic Element SCCmec-encoded psm-mec RNA Suppresses Translation of agrA and Attenuates MRSA Virulence. PLoS Pathog. 2013, 9, e1003269. [Google Scholar] [CrossRef]
- Kaito, C.; Saito, Y.; Nagano, G.; Ikuo, M.; Omae, Y.; Hanada, Y.; Han, X.; Kuwahara-Arai, K.; Hishinuma, T.; Baba, T.; et al. Transcription and Translation Products of the Cytolysin Gene psm-mec on the Mobile Genetic Element SCCmec Regulate Staphylococcus aureus Virulence. PLoS Pathog. 2011, 7, e1001267. [Google Scholar] [CrossRef]
- Oliveira, D.C.; De Lencastre, H. Multiplex PCR strategy for rapid identification of structural types and variants of the mec element in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2002, 46, 2155–2161. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, D.; Claus, H.; Witte, W.; Rothgänger, J.; Claus, H.; Turnwald, D.; Vogel, U. Typing of methicillin-resistant Staphylococcus aureus in a university hospital setting by using novel software for spa repeat determination and database management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Ito, T.; Ma, X.X.; Watanabe, S.; Kreiswirth, B.N.; Etienne, J.; Hiramatsu, K. Combination of multiplex PCRs for staphylococcal cassette chromosome mec type assignment: Rapid identification system for mec, ccr, and major differences in junkyard regions. Antimicrob. Agents Chemother. 2007, 51, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.-S.; Otto, M. The Isolation and Analysis of Phenol-Soluble Modulins of Staphylococcus epidermidis. In Staphylococcus Epidermidis. Methods in Molecular Biology (Methods and Protocols); Fey, P.D., Ed.; Humana Press: Totowa, NJ, USA, 2014; pp. 93–100. ISBN 9781627037358. [Google Scholar]
Figure 1.
Association between PSM production and methicillin-resistance or SCCmec type of MRSA. (A) Variation in PSM production between MSSA and MRSA strains and (B) variation in PSM production between SCCmec types (II, IV, IVA, and V) of MRSA. PSM production in the culture supernatants of S. aureus isolates grown in TSB for 20 h at 37 °C with 200 rpm shaking was measured by LC-MS and shown as the sum of formylated and deformylated forms. Each isolate was assayed two times, and the results were averaged for each isolate and PSM. Each symbol represents one strain while the horizontal bars represent the median. Statistical analysis for the difference in average PSM production between MSSA and MRSA as well as between SCCmec types of MRSA was performed using separate Kruskal–Wallis tests. * p < 0.05, ** p < 0.01, *** p < 0.001.
Figure 1.
Association between PSM production and methicillin-resistance or SCCmec type of MRSA. (A) Variation in PSM production between MSSA and MRSA strains and (B) variation in PSM production between SCCmec types (II, IV, IVA, and V) of MRSA. PSM production in the culture supernatants of S. aureus isolates grown in TSB for 20 h at 37 °C with 200 rpm shaking was measured by LC-MS and shown as the sum of formylated and deformylated forms. Each isolate was assayed two times, and the results were averaged for each isolate and PSM. Each symbol represents one strain while the horizontal bars represent the median. Statistical analysis for the difference in average PSM production between MSSA and MRSA as well as between SCCmec types of MRSA was performed using separate Kruskal–Wallis tests. * p < 0.05, ** p < 0.01, *** p < 0.001.

Figure 2.
Variation in PSM production between similar spa types (t002, t008, t189, t304, and t324) of MSSA and MRSA strains. Difference in the average production of each PSM between MSSA and MRSA isolates was calculated using separate Kruskal–Wallis tests. No significant difference in the production of any of the PSMs was found for similar spa types of MSSA and MRSA (p > 0.05).
Figure 2.
Variation in PSM production between similar spa types (t002, t008, t189, t304, and t324) of MSSA and MRSA strains. Difference in the average production of each PSM between MSSA and MRSA isolates was calculated using separate Kruskal–Wallis tests. No significant difference in the production of any of the PSMs was found for similar spa types of MSSA and MRSA (p > 0.05).
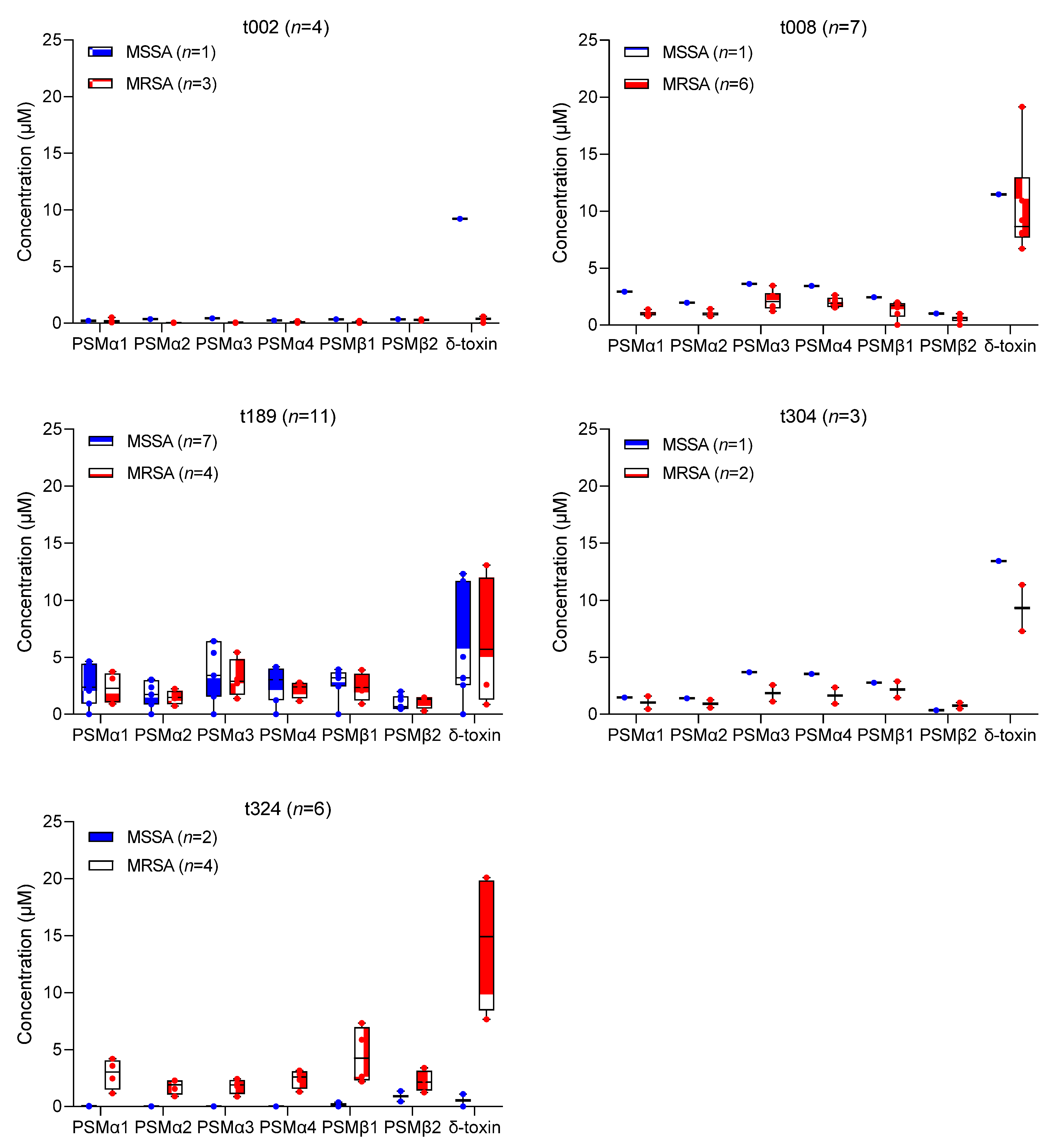
Figure 3.
PSM production by different spa types (>2 strains per spa type) of S. aureus. Singleton spa types were not included in the figure. Difference in the average PSM production between different spa types was calculated using pairwise Kruskal–Wallis tests. * p < 0.05, ** p < 0.01, and *** p < 0.001.
Figure 3.
PSM production by different spa types (>2 strains per spa type) of S. aureus. Singleton spa types were not included in the figure. Difference in the average PSM production between different spa types was calculated using pairwise Kruskal–Wallis tests. * p < 0.05, ** p < 0.01, and *** p < 0.001.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Genotype distribution of S. aureus clinical isolates.
| spa Type | S. aureus Isolates n (%) | Phenotype | SCCmec Type for MRSA | |
|---|---|---|---|---|
| MSSA n (%) | MRSA n (%) | |||
| MSSA and MRSA | ||||
| t189 | 11 (10.4) | 7 (6.6) | 4 (3.8) | IV |
| t008 | 7 (6.6) | 1 (0.9) | 6 (5.7) | IV |
| t324 | 6 (5.7) | 2 (1.9) | 4 (3.8) | IVA |
| t002 | 4 (3.8) | 1 (0.9) | 3 (2.8) | II |
| t304 | 3 (2.8) | 1 (0.9) | 2 (1.9) | IV |
| MSSA | ||||
| t126 | 6 (5.7) | 6 (5.7) | - | O |
| t084 | 3 (2.8) | 3 (2.8) | - | O |
| t085 | 2 (1.9) | 2 (1.9) | - | O |
| t179 | 2 (1.9) | 2 (1.9) | - | O |
| t338 | 2 (1.9) | 2 (1.9) | - | O |
| t1858 | 2 (1.9) | 2 (1.9) | - | O |
| t005 | 1 (0.9) | 1 (0.9) | - | O |
| t019 | 1 (0.9) | 1 (0.9) | - | O |
| t021 | 1 (0.9) | 1 (0.9) | - | O |
| t127 | 1 (0.9) | 1 (0.9) | - | O |
| t177 | 1 (0.9) | 1 (0.9) | - | O |
| t346 | 1 (0.9) | 1 (0.9) | - | O |
| t363 | 1 (0.9) | 1 (0.9) | - | O |
| t386 | 1 (0.9) | 1 (0.9) | - | O |
| t416 | 1 (0.9) | 1 (0.9) | - | O |
| t521 | 1 (0.9) | 1 (0.9) | - | O |
| t571 | 1 (0.9) | 1 (0.9) | - | O |
| t1333 | 1 (0.9) | 1 (0.9) | - | O |
| t1361 | 1 (0.9) | 1 (0.9) | - | O |
| t1767 | 1 (0.9) | 1 (0.9) | - | O |
| t1950 | 1 (0.9) | 1 (0.9) | - | O |
| t4727 | 1 (0.9) | 1 (0.9) | - | O |
| t4956 | 1 (0.9) | 1 (0.9) | - | O |
| t10234 | 1 (0.9) | 1 (0.9) | - | O |
| t10686 | 1 (0.9) | 1 (0.9) | - | O |
| t12605 | 1 (0.9) | 1 (0.9) | - | O |
| undefined | 1 (0.9) | 1 (0.9) | - | O |
| MRSA | ||||
| t2460 | 10 (9.4) | - | 10 (9.4) | II |
| t1784 | 5 (4.7) | - | 5 (4.7) | IV |
| t111 | 4 (3.8) | - | 4 (3.8) | II |
| t664 | 3 (2.8) | - | 3 (2.8) | IVA |
| t9353 | 3 (2.8) | - | 3 (2.8) | II |
| t148 | 2 (1.9) | - | 2 (1.9) | IVA |
| t034 | 1 (0.9) | - | 1 (0.9) | V |
| t062 | 1 (0.9) | - | 1 (0.9) | II |
| t242 | 1 (0.9) | - | 1 (0.9) | IV |
| t264 | 1 (0.9) | - | 1 (0.9) | II |
| t893 | 1 (0.9) | - | 1 (0.9) | II |
| t1081 | 1 (0.9) | - | 1 (0.9) | V |
| t1154 | 1 (0.9) | 1 (0.9) | II | |
| t1560 | 1 (0.9) | - | 1 (0.9) | II |
| t3092 | 1 (0.9) | - | 1 (0.9) | V |
| t4359 | 1 (0.9) | - | 1 (0.9) | IVA |
MRSA, methicillin-resistant S. aureus; MSSA, methicillin-sensitive S. aureus; n, number of isolates; SCCmec, staphylococcal cassette chromosome mec; spa, staphylococcal protein A; -, none; O, non-typeable.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lade, H.; Chung, S.H.; Lee, Y.; Joo, H.-S.; Kim, J.-S. Genotypes of Staphylococcus aureus Clinical Isolates Are Associated with Phenol-Soluble Modulin (PSM) Production. Toxins 2022, 14, 556. https://doi.org/10.3390/toxins14080556
AMA Style
Lade H, Chung SH, Lee Y, Joo H-S, Kim J-S. Genotypes of Staphylococcus aureus Clinical Isolates Are Associated with Phenol-Soluble Modulin (PSM) Production. Toxins. 2022; 14(8):556. https://doi.org/10.3390/toxins14080556
Chicago/Turabian StyleLade, Harshad, Sung Hee Chung, Yeonhee Lee, Hwang-Soo Joo, and Jae-Seok Kim. 2022. "Genotypes of Staphylococcus aureus Clinical Isolates Are Associated with Phenol-Soluble Modulin (PSM) Production" Toxins 14, no. 8: 556. https://doi.org/10.3390/toxins14080556
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.