1. Introduction
Botulinum neurotoxins (BoNTs) are produced by anaerobic spore-forming Clostridia, including
Clostridium botulinum,
C. butyricum, and
C. baratii. The ingestion of food or feed contaminated with BoNTs can cause flaccid paralysis, called botulism, which can lead to death by respiratory failure [
1,
2]. BoNTs are among the most potent bacterial toxins known [
3]. BoNTs are traditionally grouped into seven serotypes, A−G, based on recognition by antisera. The toxins of different serotypes are zinc metalloproteases that possess differential protease specificity; BoNT/A, C, and E cleave synaptosomal-associated protein (SNAP)-25, and BoNT/B, D, F, and G cleave synaptobrevin-2 (also known as VAMP-2). In addition to SNAP-25, BoNT/C also cleaves syntaxin [
4].
The structure of BoNTs was recently reviewed [
1,
2]. The 150-kDa BoNT holotoxin consists of a 50-kDa light chain (LC) and a 100-kDa heavy chain (HC). The LC and HC are linked by a disulfide bond and are complexed with accessory proteins that allow BoNT to persist in the environment. The HC is composed of a 50-kDa N-terminal domain (H
N) and a 50-kDa C-terminal domain (H
C). The various domains have specific functions. The H
C domain enables binding to gangliosides on neurons. The H
N domain is responsible for the translocation of the LC into the cytoplasm. Finally, the LC proteolyzes the intracellular synaptic vesicle protein SNAP-25 or synaptobrevin-2 resulting in the inhibition of neurotransmitter release into the synaptic cleft causing flaccid paralysis. Many other bacterial toxins, including the diphtheria toxin, anthrax lethal and edema toxins,
C. difficile toxin A and tetanus neurotoxin (TeNT), are similarly composed of three functional domains [
5,
6].
BoNT/A, B, E, F and G are mainly associated with botulism in humans whereas BoNT/C, D, CD and DC are associated with animal botulism. BoNT/DC and CD are naturally existing mosaic toxins related to BoNT/C and D [
7]. The BoNT/DC LC and H
N domains are almost identical to BoNT/D, whereas its H
C domain (H
C/DC) shares about 77% amino acid sequence identity with H
C/C. The protein sequences of BoNT/CD are almost identical to LC/C, H
N/C and H
C/D.
C. botulinum strains can be classified into different groups based on various characteristics. The so-called group III strains produce BoNT/C and D and their mosaic variants [
8,
9]. The work presented here focuses on diagnostics for group III BoNTs.
Due to their high toxicity, there is a need for highly sensitive diagnostics for BoNT detection and identification. For laboratory detection of BoNTs the mouse bioassay is still the gold standard, despite international efforts to develop alternative in vitro tests to reduce animal testing. Although many alternative technologies have been developed, reliable detection of all (sub)serotypes in complex clinical and environmental matrices remains a challenge. One of the most promising methods is the mass spectrometry (MS)-based endopeptidase BoNT activity assay known as Endopep-MS [
10]. Endopep-MS detects the enzymatic action of the LC on a peptide substrate which mimics BoNTs in vivo protein target. By using BoNT-specific peptide substrates and monitoring the cleavage position by examining the mass of the N-terminal (NT) and C-terminal (CT) cleavage products, the various BoNTs can be differentiated. An immunoaffinity step, with capture antibodies on magnetic beads, prior to incubation with the peptide substrate, is a crucial aspect for detecting and differentiating BoNTs in clinical specimens and culture supernatants [
11,
12,
13]. The immunoaffinity not only increases the sensitivity of the test by concentrating BoNT, but it also allows a washing step to remove non-specific proteases that are often present in clinical samples. The sensitivity of Endopep-MS has been shown to equal or exceed that of the mouse bioassay [
13,
14,
15,
16,
17,
18,
19], has shown good performance in an international proficiency test for detection of BoNT/A, B and E [
20], and compares favorably to other in vitro methods for routine diagnostics of clinical specimens and food [
21,
22].
While most advances have been made to improve the Endopep-MS assay for BoNT types related to human botulism [
12,
14,
23], detection and identification of group III BoNTs related to animal botulism has also progressed [
13,
14,
16]. A major challenge is the interference of non-specific proteases that are present in many complex sample matrices, like gastrointestinal contents, liver, feed and environmental samples [
24]. These proteases not only may degrade the capture antibodies on the magnetic beads but can degrade the peptides used as a substrate for the enzymatic cleavage by BoNT. A wash with 2 M NaCl in PBS (high salt wash) after immunocapture of BoNTs from complex matrices has been shown to reduce non-specific protease activity [
13,
16,
25]. Endopep-MS was also improved by the optimization of peptide substrates [
19,
26]. For efficient immunocapture, especially for BoNT/A, B, E and F, monoclonal antibodies (mAbs) have been optimized by increasing affinity using various molecular evolution approaches. Large increases in affinity have been obtained, resulting in up to 1000-fold lower affinity constants (
KD) that are due to both an increase in association (
ka) and a decrease in dissociation (
kd) rate constant [
11,
27,
28]. Also, for group III BoNTs capture mAbs have been developed that perform well in Endopep-MS [
14,
15,
16,
26]. More recently, mAbs against group III BoNTs were affinity matured for human therapeutic application [
29]. However, the utilization of many of the published mAbs is conditional, which makes them less suitable for use in routine diagnostics.
In addition to conventional mAbs, various recombinant antibodies against BoNT have been isolated for use in diagnostics or therapy. This includes single-domain antibodies (VHHs) that are derived from camelid heavy-chain antibodies. These VHHs are mostly directed against BoNT/A, B, E and F, whereas some bind group III BoNTs [
30,
31,
32,
33,
34,
35,
36]. Due to their single-gene origin, VHHs are highly suitable for the genetic fusion of different VHHs into multimers to increase affinity for their targets up to 1000-fold [
37,
38,
39,
40]. Multimerization has also been successfully applied to increase the neutralization of several bacterial toxins in animal disease models, including anthrax [
41], ricin [
42,
43], Shiga toxin [
44], TeNT [
5],
C. difficile toxin A [
45] and BoNT/A [
46].
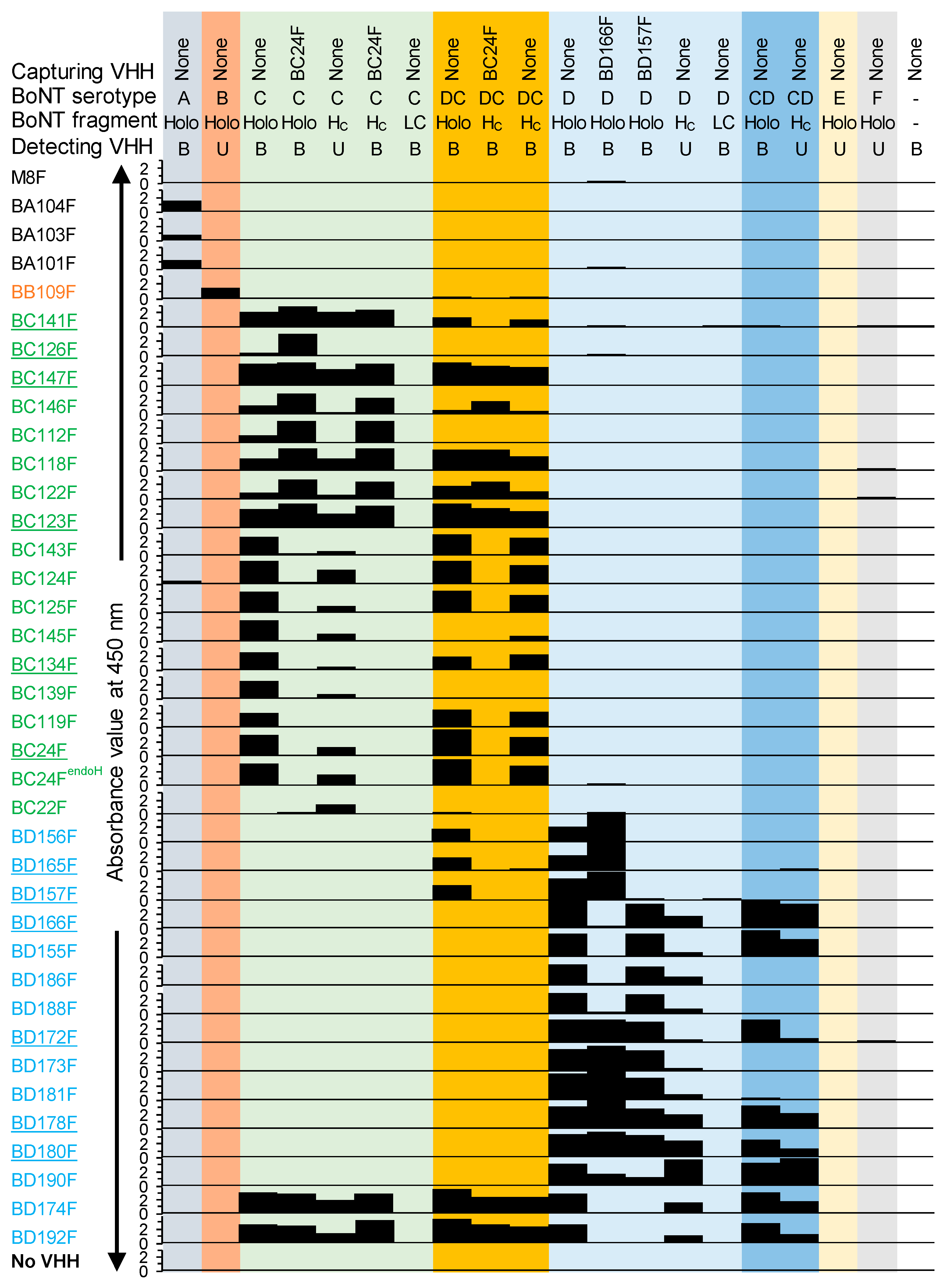
To enable development and, ultimately, implement in-house routine diagnostics for BoNTs, novel antibodies were generated. In this work, 34 VHHs were isolated that bind group III BoNTs (30 VHHs), BoNT/A (3 VHHs) or BoNT/B (1 VHH). These 34 VHHs and 2 published VHHs [
30] were yeast-produced and their antigenic specificity was characterized. VHH binding independent antigenic sites were used for generating 52 VHH multimers by genetic fusion of 2 or 3 VHHs. A selection of 15 multimers with high affinity (
KD < 0.1 nM) for various group III BoNTs was further evaluated for use in routine diagnosis by Endopep-MS.
3. Discussion
We isolated 34 novel VHHs against BoNTs with a focus on group III BoNTs. The VHHs were selected against BoNT/Ci and Di by phage display. Some VHHs were shown to cross-react with BoNT/DCi or CDi mosaic variants. During selections we used, next to the holotoxins, the H
C domains since for endopeptidase-based tests it is important to have a fully active LC. VHHs binding H
C are least likely to interfere with Endopep-MS because it is located distant from the LC domain (
Figure 2E) that carries the enzymatic activity. The use of antibodies against H
C was recommended after the screening of mAbs for use in Endopep-MS showed inhibition of BoNT activity by LC binding mAbs [
53]. Furthermore, BoNTs in complex matrices that are proteolytically cleaved in the H
N domain cannot be detected by Endopep-MS after immunocapture specific for H
C as the LC-carrying activity is not captured. Thus, the use of H
C-specific capture antibodies reduces false positives in Endopep-MS as compared to LC or H
N-specific capture antibodies [
24,
54]. However, BC22F binds a site only available in the H
C protein but not in BoNT/Ci holotoxin. Presumably, this VHH binds to a site close to the H
C/C N-terminus that is not accessible in the BoNT holotoxin due to steric hindrance by the H
N domain, as suggested earlier [
30].
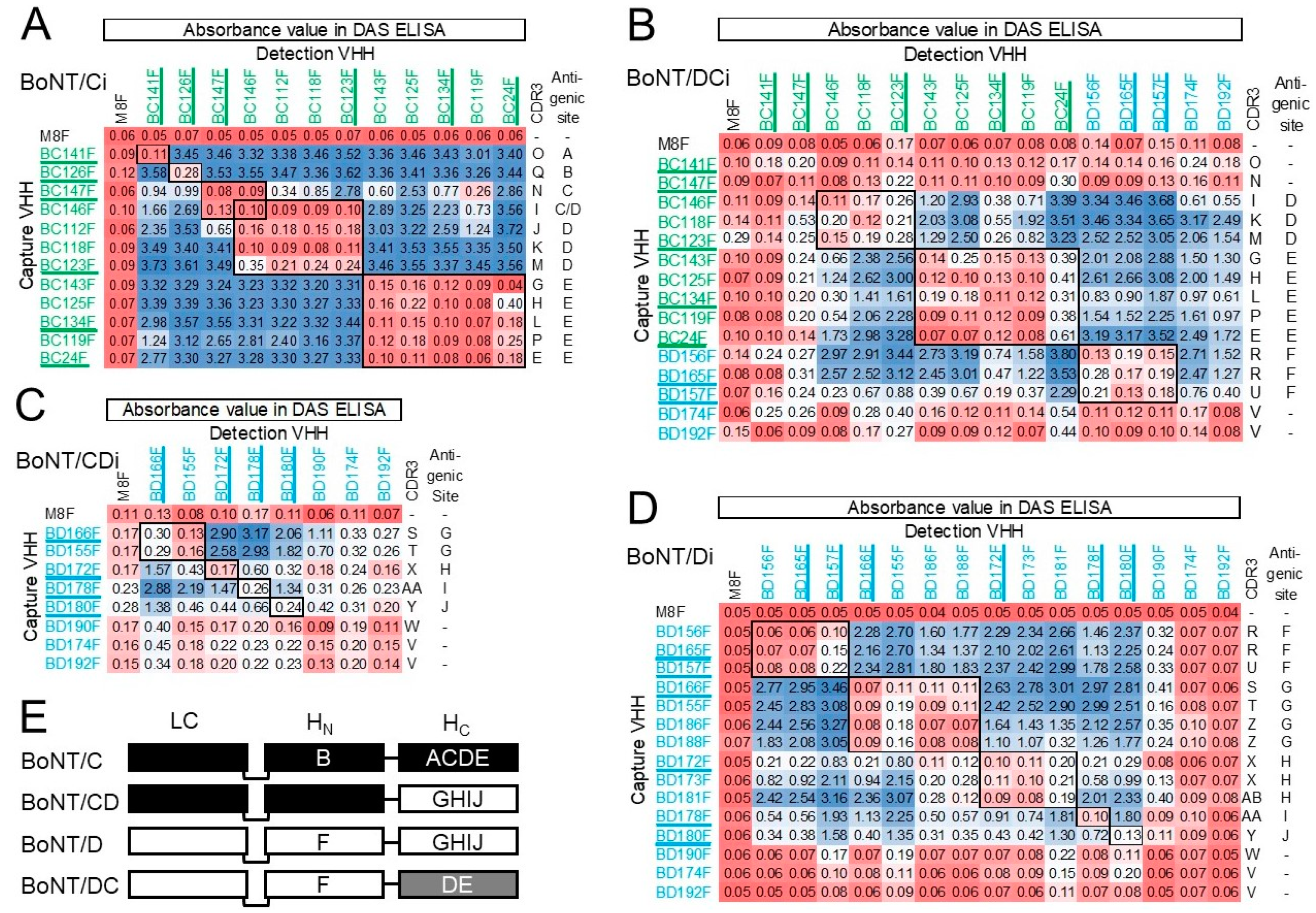
The affinity of VHH multimers generated in this study for BoNT/DCi is generally lower than for BoNT/Ci. This is presumably due to the specificity of most monomers for H
C/C, which shows only about 77% identity with H
C/DC [
7]. Consistent with this conclusion VHH multimers showed much more similar affinities for BoNT/Di and CDi, of which the sequence identity of the H
C domains is much higher.
BD174F and BD192F, both found during phage display selection with H
C/D, were the only two VHHs that bind both BoNT/Ci and Di, and thus also the mosaic variants. None of the other VHHs recognized both BoNT/Ci and Di. MAbs that recognize both BoNT/C and D were isolated earlier and mostly bound H
N [
29,
55], which has a region with high sequence homology of BoNT/C and D [
7]. Furthermore, an mAb that recognized BoNT/A, B, E and F was also shown to bind to H
N [
11]. However, VHHs BD174F and BD192F were found to bind H
C and thus recognize a common epitope in group III H
C domains. Since these VHHs did not perform well in DAS-ELISA their antigenic site(s) could not be mapped by epitope binning. A further two VHHs, BC141F and BC147F, similarly did not bind well to BoNT/DCi in DAS ELISA as well as in biolayer interferometry assays, although they did bind to directly coated BoNT/DCi. This could indicate an epitope that is altered or blocked upon direct coating of BoNT. Such effects of direct coating of antigens by passive adsorption to polystyrene on antigenicity have been observed earlier [
56,
57]. In the case of BC141F and BC147F, the relatively low affinity for BoNT/Ci (
KD > 0.5 nM), which probably is even lower for BoNT/DCi, could also cause reduced binding to soluble BoNT/DCi.
The VHHs that performed well in DAS-ELISA recognized independent antigenic sites A–E for BoNT/Ci and sites F-J for BoNT/Di. At least 20 independent antigenic sites were identified on TeNT [
58], which is structurally highly similar to BoNT, and the 61 kDa ricin toxin [
59]. The lower number of antigenic sites identified by our panel of VHHs could be due to the high proportion of VHHs binding H
C, which encompasses four out of five antigenic sites recognized for both BoNT/C and D.
Oligoclonal mixtures of recombinant antibodies recognizing different antigenic sites were earlier demonstrated to increase the sensitivity of BoNT/A detection [
60]. However, the multimerization of VHHs was proven more effective than making equimolar mixtures of VHHs for increasing affinity, as shown for Shiga toxin [
44] and BoNT [
46]. Therefore, using a set of 12 monomeric VHHs, we chose to produce a broad panel of 52 VHH multimers composed of 2 or 3 constituent VHH domains that each recognize an independent antigenic site. Using biolayer interferometry we identified a number of VHH multimers with increased affinity (avidity) as compared to the monovalent corresponding VHH with highest affinity (
Table 3 and
Table 4). However, some VHH multimers were found to have lower affinities than the optimal corresponding monomers. This was especially apparent for many multimers against BoNT/D containing BD166, which has the highest affinity (
KD = 62 pM) among the BoNT/D binding VHHs. Since the N-terminus of a VHH is close to the antigen-binding site, the genetic fusion of a VHH can affect antigen binding, especially the C-terminal VHH due to steric hindrance by the N-terminal VHH [
37]. Furthermore, mutations Q1E and Q5V of the C-terminal VHH (see
Figure S2) could also affect antigen binding. These two mechanisms can explain compromised antigen binding of the C-terminal VHH domain but not the N-terminal VHH domain in a multimer. However, many multimers containing an N-terminal BD166 domain also show lower affinities than the BD166F monomer (
Table 4). This could be due to prior binding by C-terminal VHH(s) to BoNT affecting binding by the BD166 domain because the two antigenic sites are not properly positioned to allow simultaneous binding of the linked VHH domains.
It is generally assumed that the increased affinity of heterodimeric antibodies is due to the binding of one antigen-binding domain resulting in large increases in “target residence time”, which can lead to rapid rebinding by other domains upon dissociation of a single antigen-binding domain [
61]. However, such a model only applies if both antigen-binding domains recognize epitopes that are suitably oriented to allow simultaneous binding of two genetically linked antibody domains. Long flexible linkers such as the 15 amino acid (GGGGS)
3 linker used here are generally employed for VHH multimerization to increase the conformational flexibility required for such simultaneous binding to different epitopes on a single antigen molecule. We assumed that such simultaneous binding is more often seen for multimers that recognize different toxin domains because conformational flexibility between domains more likely results in properly oriented epitopes. Therefore, we also produced multimers composed of BD157 or BD165 domains, that bind antigenic site F on H
N, in combination with VHHs binding H
C domains for both BoNT/DCi (
Table 3) and BoNT/Di or CDi (
Table 4). However, we did not clearly observe higher binding affinities of VHH multimers containing VHHs that bind to different toxin domains as compared to multimers that bind different antigenic sites on the same (H
C) domain. For multimers with BC126, which is assumed to bind H
N/C, such improved binding is more difficult to demonstrate since BC126F already has a high affinity of 25 pM. Furthermore, the H
N binding of BC126F is only based on the absence of binding to recombinant H
C in direct ELISA and is not confirmed by specificity for BoNT types in ELISAs, as was the case for BD157F and BD165F. The exact epitopes bound by our VHHs were not further determined, beyond the specificity for BoNT domains. More detailed knowledge of epitopes bound by X-ray crystallography was used earlier for the structure-based rational design of VHH multimers with potent BoNT neutralization for therapeutic purposes [
33].
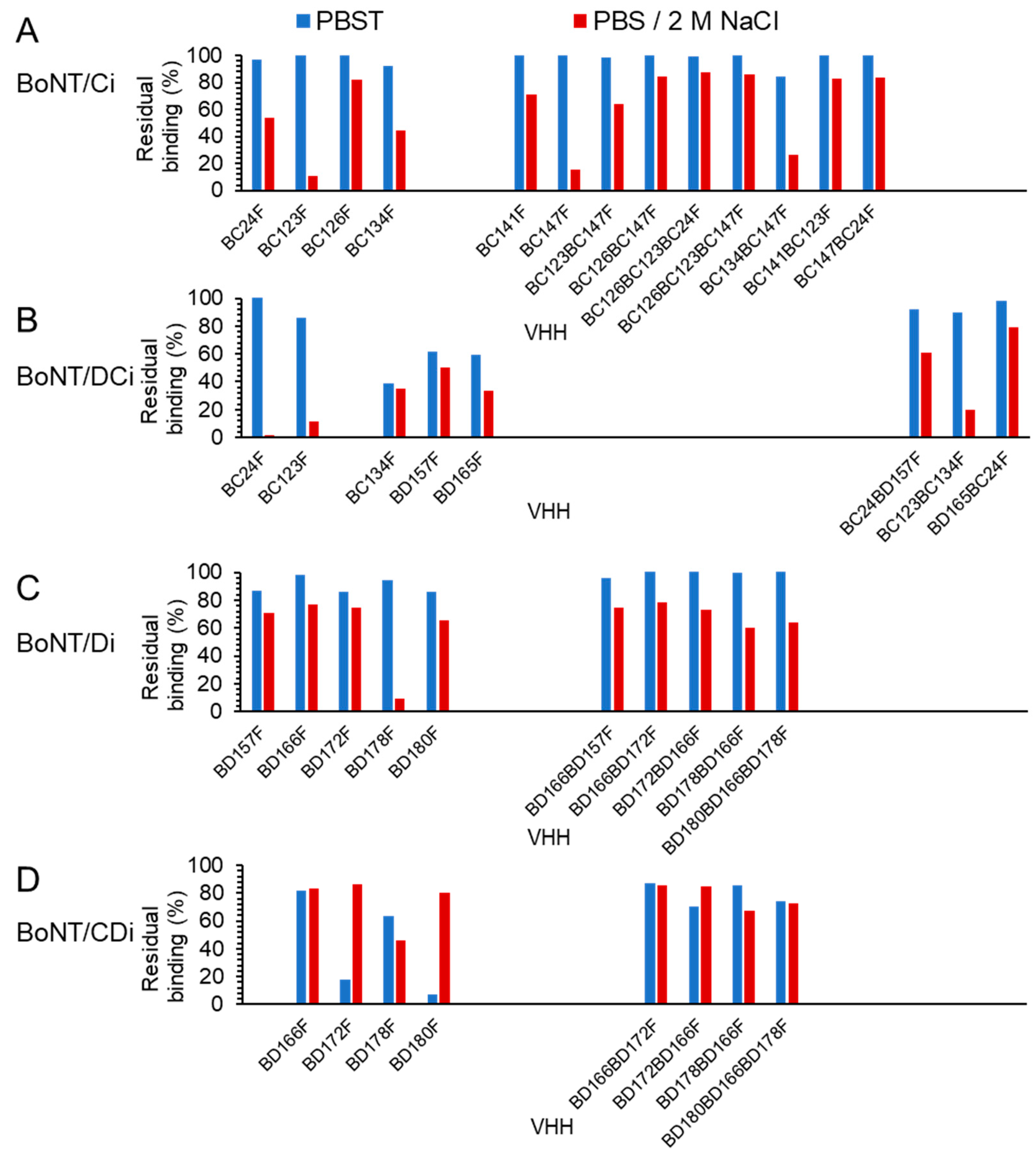
Fifteen multimers were found to have a high affinity (
KD < 0.1 nM;
Table 3 and
Table 4). They were found suitable for binding to BoNTs in the presence of 2 M NaCl, which is often used as a wash step of samples immunocaptured from complex matrices to remove aspecific proteases prior to Endopep-MS [
13,
16,
25]. Using high salt, VHH multimers generally showed decreased dissociation as compared to VHH monomers, consistent with their increased affinity. The 15 high-affinity VHH multimers were found suitable for the detection of the 4 group III BoNTs produced from bacterial culture in Endopep-MS. Unexpectedly, the limit of detection (LOD) in these assays of VHH multimers was not higher than their corresponding monomers. This could be caused by not all constituent VHHs in a multimer contributing to BoNT binding, due to variation in BoNTs from different strains [
62], which often occurs with mosaic variants [
63]. However, other (unidentified) reasons for multimers with increased affinity not resulting in an increased sensitivity of Endopep-MS, cannot be excluded.
Bjornstad et al. [
14] developed an excellent Endopep-MS test design to discriminate the four group III BoNT-types based on different combinations of specificity of the capture antibody and specificity of BoNT proteolytic activity. A similar approach can be achieved if two or more VHH multimers are used that specifically bind either BoNT/C and DC or BoNT/D and CD, in combination with different peptide substrates for LC/C and LC/D. However, the three multimers BC24BD157F, BD165FBC24F and BD166BD157F are not suitable for such discrimination of all four group III BoNTs in Endopep-MS since they contain the VHHs BD165 or BD157 that both recognize site F on H
N of BoNT/D and DC (
Figure 2E). The above considerations will be taken into account in our future studies to determine which high-affinity multimers are most suited for diagnosing BoNT in clinical samples using Endopep-MS.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}



