
The Recombinant Maize Ribosome-Inactivating Protein Transiently Reduces Viral Load in SHIV89.6 Infected Chinese Rhesus Macaques

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Antiviral Effects of Maize RIP on Infected Macaque PBMC

{kind=link}
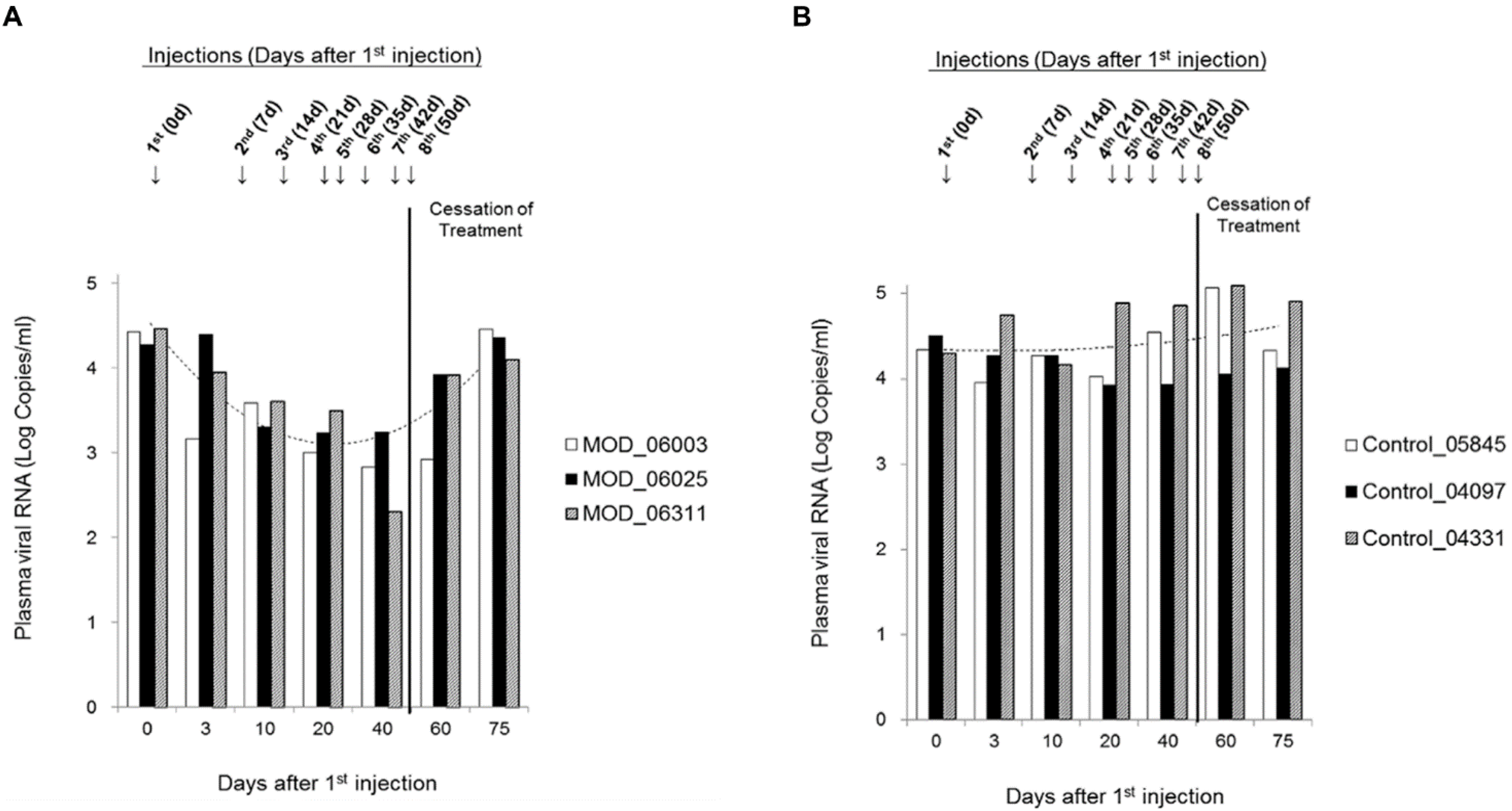{kind=link}
{kind=link}
| RIP variants | Cytotoxicity CC50 (μM) | p27 Antigen reduction EC50 (μM) | |
|---|---|---|---|
| Uninfected | SHIV89.6 | SIVmac239 | |
| His-TAT-Pro | >15 | >24.85 | >24.85 |
| His-TAT-MOD | 8.98 ± 0.36 | 5.53 | 11.23 |
2.2. Antiviral Activities of Maize RIP in Rhesus Macaques


| Markers | Days after first injection of His-TAT-MOD | |||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 3 | 10 | 20 | 40 | 60 | 75 | p value | |
| ALT (U/L) | 41.67 ± 31.07 | 40.33 ± 32.08 | 41.33 ± 36.02 | 28.33 ± 19.40 | 24.00 ± 16.70 | 21.67 ± 16.01 | 22.00 ± 17.09 | 0.022 |
| AST (U/L) | 24.67 ± 15.63 | 34.67 ± 21.39 | 28.00 ± 23.90 | 29.00 ± 20.66 | 23.00 ± 15.10 | 23.67 ± 15.57 | 30.33 ± 17.16 | 0.110 |
| WBC (109/L) | 9.21 ± 2.30 | 9.62 ± 3.72 | 9.77 ± 2.17 | 8.63 ± 2.18 | 10.87 ± 2.10 | 9.87 ± 2.97 | 8.07 ± 1.82 | 0.546 |
| Lym (109/L) | 3.94 ± 2.27 | 3.97 ± 2.07 | 3.29 ± 1.81 | 3.60 ± 1.51 | 4.87 ± 1.75 | 3.87 ± 2.10 | 4.17 ± 1.98 | 0.926 |
| Lym (%) | 43.87 ± 22.78 | 47.83 ± 30.31 | 36.03 ± 22.74 | 44.73 ± 22.02 | 45.93 ± 16.35 | 39.27 ± 19.20 | 50.10 ± 12.97 | 0.961 |
| Mon (109/L) | 0.44 ± 0.32 | 0.34 ± 0.19 | 0.29 ± 0.04 | 0.77 ± 0.23 | 1.00 ± 0.17 | 0.97 ± 0.29 | 0.80 ± 0.10 | 0.099 |
| Mon (%) | 4.60 ± 2.33 | 3.37 ± 0.57 | 3.03 ± 0.29 | 8.87 ± 1.91 | 9.70 ± 2.02 | 9.87 ± 2.25 | 9.70 ± 0.52 | 0.090 |
| Gran (109/L) | 5.83 ± 2.19 | 5.31 ± 5.11 | 6.16 ± 3.53 | 4.27 ± 3.17 | 5.00 ± 2.78 | 4.60 ± 1.61 | 3.10 ± 0.35 | 0.049 |
| Gran (%) | 51.53 ± 20.58 | 48.80 ± 29.74 | 60.93 ± 22.98 | 46.40 ± 23.09 | 44.37 ± 18.24 | 48.73 ± 20.94 | 40.20 ± 12.54 | 0.301 |
| RBC (1012/L) | 5.85 ± 0.86 | 5.80 ± 0.71 | 5.86 ± 0.66 | 6.41 ± 0.60 | 6.39 ± 0.39 | 6.16 ± 0.19 | 5.80 ± 0.11 | 1.121 |
| HGB (g/L) | 127.00 ± 27.73 | 125.00 ± 28.69 | 128.00 ± 32.23 | 128.00 ± 26.63 | 132.67 ± 19.60 | 131.00 ± 16.64 | 118.67 ± 15.70 | 0.917 |
| HCT (%) | 40.83 ± 8.69 | 39.30 ± 8.49 | 39.07 ± 8.68 | 44.37 ± 8.16 | 43.87 ± 6.02 | 42.33 ± 4.82 | 40.00 ± 4.47 | 0.552 |
| MCV (fL) | 69.33 ± 5.19 | 67.20 ± 7.13 | 66.17 ± 8.94 | 69.07 ± 8.23 | 68.70 ± 8.55 | 68.90 ± 8.62 | 69.07 ± 7.52 | 0.069 |
| MCH (pg) | 21.53 ± 1.76 | 21.33 ± 2.54 | 21.67 ± 3.69 | 19.83 ± 2.80 | 20.73 ± 2.72 | 21.30 ± 3.03 | 20,37 ± 2.66 | 0.067 |
| PLT (109/L) | 422.00 ± 299.52 | 415.33 ± 298.26 | 482.00 ± 319.24 | 556.33 ± 340.35 | 536.33 ± 334.32 | 483.67 ± 296.48 | 482.00 ± 336.72 | 0.017 |
| MPV (fL) | 10.70 ± 0.98 | 10.53 ± 1.07 | 10.43 ± 1.32 | 7.87 ± 0.78 | 7.90 ± 1.06 | 7.73 ± 1.34 | 7.20 ± 1.14 | 0.018 |
| Markers | Days after first injection of normal saline | |||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 3 | 10 | 20 | 40 | 60 | 75 | p value | |
| ALT (U/L) | 79.33 ± 42.67 | 90.00 ± 51.10 | 68.00 ± 31.10 | 49.00 ± 19.92 | 23.33 ± 10.41 | 31.00 ± 21.66 | 39.33 ± 17.67 | 0.035 |
| AST (U/L) | 44.00 ± 25.16 | 80.67 ± 37.86 | 28.67 ± 10.02 | 32.00 ± 2.65 | 25.33 ± 6.66 | 26.33 ± 5.86 | 27.67 ± 3.79 | 0.451 |
| WBC (109/L) | 7.18 ± 1.62 | 6.41 ± 2.20 | 6.24 ± 1.55 | 6.40 ± 2.39 | 7.10 ± 1.47 | 9.17 ± 0.64 | 6.87 ± 1.10 | 0.754 |
| Lym (109/L) | 2.39 ± 1.33 | 3.23 ± 1.57 | 2.36 ± 1.16 | 2.57 ± 0.95 | 2.60 ± 0.75 | 2.80 ± 0.87 | 2.83 ± 1.12 | 0.038 |
| Lym (%) | 31.70 ± 14.81 | 43.40 ± 9.55 | 36.47 ± 10.90 | 40.00 ± 4.39 | 36.30 ± 6.22 | 32.13 ± 12.63 | 44.47 ± 5.33 | 0.014 |
| Mon (109/L) | 0.45 ± 0.22 | 0.42 ± 0.29 | 0.47 ± 0.15 | 0.53 ± 0.21 | 0.73 ± 0.21 | 0.73 ± 0.12 | 0.60 ± 0.10 | 0.059 |
| Mon (%) | 6.13 ± 2.14 | 6.00 ± 0.96 | 7.60 ± 2.04 | 8.70 ± 2.70 | 10.63 ± 1.05 | 8.50 ± 1.55 | 9.90 ± 1.15 | 0.015 |
| Gran (109/L) | 4.35 ± 0.98 | 2.70 ± 0.54 | 3.41 ± 0.58 | 3.30 ± 1.39 | 3.77 ± 0.86 | 5.53 ± 1.53 | 2.80 ± 0.40 | 0.129 |
| Gran (%) | 62.17 ± 17.15 | 44.63 ± 11.46 | 55.93 ± 12.18 | 51.30 ± 12.18 | 53.07 ± 6.98 | 59.37 ± 14.01 | 45.63 ± 4.37 | 0.007 |
| RBC (1012/L) | 7.05 ± 1.58 | 6.85 ± 0.848 | 6.48 ± 1.33 | 6.37 ± 1.00 | 6.59 ± 1.28 | 6.90 ± 0.27 | 6.80 ± 0.56 | 0.007 |
| HGB (g/L) | 155.67 ± 21.13 | 149.67 ± 14.67 | 144.00 ± 18.19 | 136.67 ± 15.18 | 146.67 ± 20.50 | 159.00 ± 2.65 | 142.67 ± 19.73 | 0.030 |
| HCT (%) | 50.50 ± 7.73 | 46.70 ± 5.99 | 45.47 ± 6.69 | 46.20 ± 5.12 | 47.77 ± 7.56 | 50.33 ± 0.29 | 46.93 ± 6.90 | 0.005 |
| MCV (fL) | 72.30 ± 4.90 | 70.30 ± 3.32 | 70.67 ± 3.78 | 72.90 ± 3.08 | 72.97 ± 3.16 | 73.97 ± 2.58 | 72.63 ± 2.39 | 0.699 |
| MCH (pg) | 22.33 ± 1.87 | 22.63 ± 1.84 | 22.43 ± 1.65 | 21.50 ± 1.20 | 22.37 ± 1.19 | 23.03 ± 1.00 | 22.03 ± 0.81 | 0.994 |
| PLT (109/L) | 374.33 ± 72.67 | 350.67 ± 39.02 | 346.00 ± 37.16 | 397.00 ± 74.48 | 413.67 ± 81.59 | 420.33 ± 9.71 | 327.33 ± 30.17 | 0.929 |
| MPV (fL) | 10.40 ± 0.53 | 10.40 ± 0.10 | 10.40 ± 0.17 | 8.13 ± 0.32 | 8.50 ± 0.17 | 7.90 ± 0.17 | 8.30 ± 0.26 | 0.026 |
| Days after first injection | Weight (kg) |
|---|---|
| 0 | 6.43 ± 1.30 |
| 17 | 6.18 ± 1.50 |
| 58 | 6.21 ± 1.15 |
| 70 | 6.61 ± 1.83 |
| Markers | His-TAT-MOD | Control | Reference values |
|---|---|---|---|
| Red blood cell (1012/L) | 4.86–7.10 | 5.41–8.81 | 7.0 ± 0.6 |
| White blood cell (109/L) | 6.69–13.83 | 4.10–9.90 | 14.58 ± 2.35 |
| Lymphocyte (109/L) | 1.80–6.80 | 0.90–4.61 | 9.27 ± 2.1 |
| CD4+ T cell (109/L) | 0.46–1.55 | 0.25–1.22 | N.D. |
| CD8+ T cell (109/L) | 0.25–2.37 | 0.25–2.37 | N.D. |
3. Discussion
4. Materials and Methods
4.1. Cloning, Expression and Purification of Maize RIP Variants

4.2. Animals for Experiment
4.3. Cells and Viruses
4.4. Cytotoxicity Assay
4.5. Viral Antigen Reduction Assay
4.6. Protection of Infected Macaque PBMC from Lysis ex vivo
4.7. Administration of Maize RIP and Sample Collection
4.8. Plasma Viral Load Determination
4.9. Flow Cytometry
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stirpe, F. Ribosome-inactivating proteins: From toxins to useful proteins. Toxicon 2013, 67, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Puri, M.; Kaur, I.; Perugini, M.A.; Gupta, R.C. Ribosome-inactivating proteins: Current status and biomedical applications. Drug Discov. Today 2012, 17, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Kaur, I.; Gupta, R.C.; Puri, M. Ribosome inactivating proteins from plants inhibiting viruses. Virol. Sin. 2011, 26, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B.; Chan, W.Y.; Yeung, H.W. Proteins with abortifacient, ribosome inactivating, immunomodulatory, antitumor and anti-AIDS activities from Cucurbitaceae plants. Gen. Pharmacol. 1992, 23, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Battelli, M.G.; Montacuti, V.; Stirpe, F. High sensitivity of cultured human trophoblasts to ribosome-inactivating proteins. Exp. Cell. Res. 1992, 201, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Rajamohan, F.; Venkatachalam, T.K.; Irvin, J.D.; Uckun, F.M. Pokeweed antiviral protein isoforms PAP-I, PAP-II, and PAP-III depurinate RNA of human immunodeficiency virus (HIV)-1. Biochem. Biophys. Res. Commun. 1999, 260, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Au, T.K.; Collins, R.A.; Lam, T.L.; Ng, T.B.; Fong, W.P.; Wan, D.C. The plant ribosome inactivating proteins luffin and saporin are potent inhibitors of HIV-1 integrase. FEBS Lett. 2000, 471, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.L.; Feng, D.; Wu, J.; Sui, S.F. Trichosanthin inhibits integration of human immunodeficiency virus type 1 through depurinating the long-terminal repeats. Mol. Biol. Rep. 2010, 37, 2093–2098. [Google Scholar] [CrossRef] [PubMed]
- Lee-Huang, S.; Kung, H.F.; Huang, P.L.; Bourinbaiar, A.S.; Morell, J.L.; Brown, J.H.; Huang, P.L.; Tsai, W.P.; Chen, A.Y.; Huang, H.I.; et al. Human immunodeficiency virus type 1(HIV-1) inhibition, DNA-binding, RNA-binding, and ribosome inactivation activities in the N-terminal segments of the plant anti-HIV protein GAP31. Proc. Natl. Acad. Sci. USA 1994, 91, 12208–12212. [Google Scholar] [CrossRef]
- Wang, Y.X.; Neamati, N.; Jacob, J.; Palmer, I.; Stahl, S.J.; Kaufman, J.D.; Huang, P.L.; Winslow, H.E.; Pommier, Y.; Wingfield, P.T.; et al. Solution structure of anti-HIV-1 and anti-tumor protein MAP30: Structural insights into its multiple functions. Cell 1999, 99, 433–442. [Google Scholar] [CrossRef]
- Lee-huang, S.; Huang, P.L.; Chen, H.C.; Huang, P.L.; Bourinbaiar, A.; Huang, H.I.; Kung, H.F. Anti-HIV and anti-tumor activities of recombinant MAP30 from bitter melon. Gene 1995, 161, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Li, H.G.; Huang, P.L.; Zhang, D.; Sun, Y.; Chen, H.C.; Zhang, J.; Kong, X.P.; Lee-Huang, S. A new activity of anti-HIV and anti-tumor protein GAP31: DNA adenosine glycosidase-structural and modeling insight into its functions. Biochem. Biophys. Res. Commun. 2010, 391, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Kahn, J.O.; Gorelick, K.J.; Gatti, G.; Arri, C.J.; Lifson, J.D.; Gambertoglio, J.G.; Bostrom, A.; Williams, R. Safety, activity, and pharmacokinetics of GLQ223 in patients with AIDS and AIDS-related complex. Antimicrob. Agents Chemother. 1994, 38, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Uckun, F.M.; Bellomy, K.; O’Neill, K.; Messinger, Y.; Johnson, T.; Chen, C.L. Toxicity, biological activity, and pharmacokinetics of TXU (anti-CD7)-pokeweed antiviral protein in chimpanzees and adult patients infected with human immunodeficiency virus. J. Pharmacol. Exp. Ther. 1999, 291, 1301–1307. [Google Scholar] [PubMed]
- Sandvig, K.; van Deurs, B. Transport of protein toxins into cells: Pathways used by ricin, cholera toxin and shiga toxin. FEBS Lett. 2002, 529, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Bradberry, S.M.; Dickers, K.J.; Rice, P.; Griffiths, G.D.; Vale, J.A. Ricin poisoning. Toxicol. Rev. 2003, 22, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Mak, A.N.; Wong, Y.T.; An, Y.J.; Cha, S.S.; Sze, K.H.; Au, S.W.; Wong, K.B.; Shaw, P.C. Structure-function study of maize ribosome-inactivating protein: Implications for the internal inactivation region and the sole glutamate in the active site. Nucleic Acids Res. 2007, 35, 6259–6267. [Google Scholar] [CrossRef] [PubMed]
- Law, S.K.; Wang, R.R.; Mak, A.N.; Wong, K.B.; Zheng, Y.T.; Shaw, P.C. A switch-on mechanism to activate maize ribosome-inactivating protein for targeting HIV-infected cells. Nucleic Acids Res. 2010, 38, 6803–6812. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Qin, S.; Ding, Y.; Wei, L.; Zhang, J.; Li, H.; Bu, H.; Lu, Y.; Cheng, J. Reference values of clinical chemistry and hematology parameters in rhesus monkeys (Macaca mulatta). Xenotransplantation 2009, 16, 496–501. [Google Scholar] [CrossRef] [PubMed]
- McGrath, M.S.; Hwang, K.M.; Caldwell, S.E.; Gaston, I.; Luk, K.C.; Wu, P.; Ng, V.L.; Crowe, S.; Daniels, J.; Marsh, J.; et al. GLQ223: An inhibitor of human immunodeficiency virus replication in acutely and chronically infected cells of lymphocyte and mononuclear phagocyte lineage. Proc. Natl. Acad. Sci. USA 1989, 86, 2844–2848. [Google Scholar] [CrossRef]
- Huang, P.L.; Sun, Y.T.; Chen, H.C.; Kung, H.F.; Huang, P.L.; Lee-Huang, S. Proteolytic fragments of anti-HIV and anti-tumor proteins MAP30 and GAP31 are biologically active. Biochem. Biophys. Res. Commun. 1999, 262, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, R.A.; Rogers, J.; Katze, M.G.; Bumgarner, R.; Weinstock, G.M.; Mardis, E.R.; Remington, K.A.; Strausberg, R.L.; Venter, J.C.; Wilson, R.K.; et al. Evolutionary and biomedical insights from the rhesus macaque genome. Science 2007, 316, 222–234. [Google Scholar] [CrossRef]
- Gao, F.; Bailes, E.; Robertson, D.L.; Chen, Y.; Rodenburg, C.M.; Michael, S.F.; Cummins, L.B.; Arthur, L.O.; Peeters, M.; Shaw, G.M.; et al. Origin of HIV-1 in the chimpanzee pan troglodytes troglodytes. Nature 1999, 397, 436–441. [Google Scholar] [CrossRef]
- Keele, B.F.; Van Heuverswyn, F.; Li, Y.; Bailes, E.; Takehisa, J.; Santiago, M.L.; Bibollet-Ruche, F.; Chen, Y.; Wain, L.V.; Liegeois, F.; et al. Chimpanzee reservoirs of pandemic and nonpandemic HIV-1. Science 2006, 313, 523–526. [Google Scholar] [CrossRef]
- Chen, Z.; Telfier, P.; Gettie, A.; Reed, P.; Zhang, L.; Ho, D.D.; Marx, P.A. Genetic characterization of new west african simian immunodeficiency virus SIVsm: Geographic clustering of household-derived SIV strains with human immunodeficiency virus type 2 subtypes and genetically diverse viruses from a single feral sooty mangabey troop. J. Virol. 1996, 70, 3617–3627. [Google Scholar] [PubMed]
- Daniel, M.D.; Letvin, N.L.; King, N.W.; Kannagi, M.; Sehgal, P.K.; Hunt, R.D.; Kanki, P.J.; Essex, M.; Desrosiers, R.C. Isolation of T-cell tropic HTLV-III-like retrovirus from macaques. Science 1985, 228, 1201–1204. [Google Scholar] [CrossRef] [PubMed]
- Letvin, N.L.; Daniel, M.D.; Sehgal, P.K.; Desrosiers, R.C.; Hunt, R.D.; Waldron, L.M.; MacKey, J.J.; Schmidt, D.K.; Chalifoux, L.V.; King, N.W. Induction of AIDS-like disease in macaque monkeys with T-cell tropic retrovirus HTLV-III. Science 1985, 230, 71–73. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.J.; Zhang, G.H.; Wang, R.R.; Zheng, Y.T. The influence of age and sex on the cell counts of peripheral blood leukocyte subpopulations in Chinese rhesus macaques. Cell. Mol. Immunol. 2009, 6, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Reimann, K.A.; Li, J.T.; Voss, G.; Lekutis, C.; Tenner-Racz, K.; Racz, P.; Lin, W.; Montefiori, D.C.; Lee-Parritz, D.E.; Lu, Y.; et al. An env gene derived from a primary human immunodeficiency virus type 1 isolate confers high in vivo replicative capacity to a chimeric simian/human immunodeficiency virus in rhesus monkeys. J. Virol. 1996, 70, 3198–3206. [Google Scholar]
- Mellors, J.W.; Rinaldo, C.R.; Gupta, P.; White, R.M.; Todd, J.A.; Kingsley, L.A. Prognosis in HIV-1 infection predicted by the quantity of virus in plasma. Science 1996, 272, 1167–1170. [Google Scholar] [CrossRef] [PubMed]
- Henrard, D.R.; Phillips, J.F.; Muenz, L.R.; Blattner, W.A.; Wiesner, D.; Eyster, M.E.; Goedert, J.J. Natural history of HIV-1 cell-free viremia. JAMA 1995, 274, 554–558. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, T.R.; Blattner, W.A.; Waters, D.; Eyster, E.; Hilgartner, M.W.; Cohen, A.R.; Luban, N.; Hatzakis, A.; Aledort, L.M.; Rosenberg, P.S.; et al. Serum HIV-1 RNA levels and time to development of AIDS in the multicenter hemophilia cohort study. JAMA 1996, 276, 105–110. [Google Scholar] [CrossRef]
- O’Brien, W.A.; Hartigan, P.M.; Martin, D.; Esinhart, J.; Hill, A.; Benoit, S.; Rubin, M.; Simberkoff, M.S.; Hamilton, J.D. Changes in plasma HIV-1 RNA and CD4+ lymphocyte counts and the risk of progression to AIDS. N. Engl. J. Med. 1996, 334, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Ho, D.D. Viral counts count in HIV infection. Science 1996, 272, 1124–1125. [Google Scholar] [CrossRef] [PubMed]
- Cote, H.C.; Brumme, Z.L.; Harrigan, P.R. Human immunodeficiency virus type 1 protease cleavage site mutations associated with protease inhibitor cross-resistance selected by indinavir, ritonavir, and/or saquinavir. J. Virol. 2001, 75, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Harrigan, P.R.; Salim, M.; Stammers, D.K.; Wynhoven, B.; Brumme, Z.L.; McKenna, P.; Larder, B.; Kemp, S.D. A mutation in the 3' region of the human immunodeficiency virus type 1 reverse transcriptase (Y318F) associated with nonnucleoside reverse transcriptase inhibitor resistance. J. Virol. 2002, 76, 6836–6840. [Google Scholar] [CrossRef] [PubMed]
- Shafer, R.W. Genotypic testing for human immunodeficiency virus type 1 drug resistance. Clin. Microbiol. Rev. 2002, 15, 247–277. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.-R.; Au, K.-Y.; Zheng, H.-Y.; Gao, L.-M.; Zhang, X.; Luo, R.-H.; Law, S.K.-Y.; Mak, A.N.-S.; Wong, K.-B.; Zhang, M.-X.; et al. The Recombinant Maize Ribosome-Inactivating Protein Transiently Reduces Viral Load in SHIV89.6 Infected Chinese Rhesus Macaques. Toxins 2015, 7, 156-169. https://doi.org/10.3390/toxins7010156
Wang R-R, Au K-Y, Zheng H-Y, Gao L-M, Zhang X, Luo R-H, Law SK-Y, Mak AN-S, Wong K-B, Zhang M-X, et al. The Recombinant Maize Ribosome-Inactivating Protein Transiently Reduces Viral Load in SHIV89.6 Infected Chinese Rhesus Macaques. Toxins. 2015; 7(1):156-169. https://doi.org/10.3390/toxins7010156
Chicago/Turabian StyleWang, Rui-Rui, Ka-Yee Au, Hong-Yi Zheng, Liang-Min Gao, Xuan Zhang, Rong-Hua Luo, Sue Ka-Yee Law, Amanda Nga-Sze Mak, Kam-Bo Wong, Ming-Xu Zhang, and et al. 2015. "The Recombinant Maize Ribosome-Inactivating Protein Transiently Reduces Viral Load in SHIV89.6 Infected Chinese Rhesus Macaques" Toxins 7, no. 1: 156-169. https://doi.org/10.3390/toxins7010156