
Limited Link between Oxidative Stress and Ochratoxin A—Induced Renal Injury in an Acute Toxicity Rat Model

Abstract
:1. Introduction
2. Results
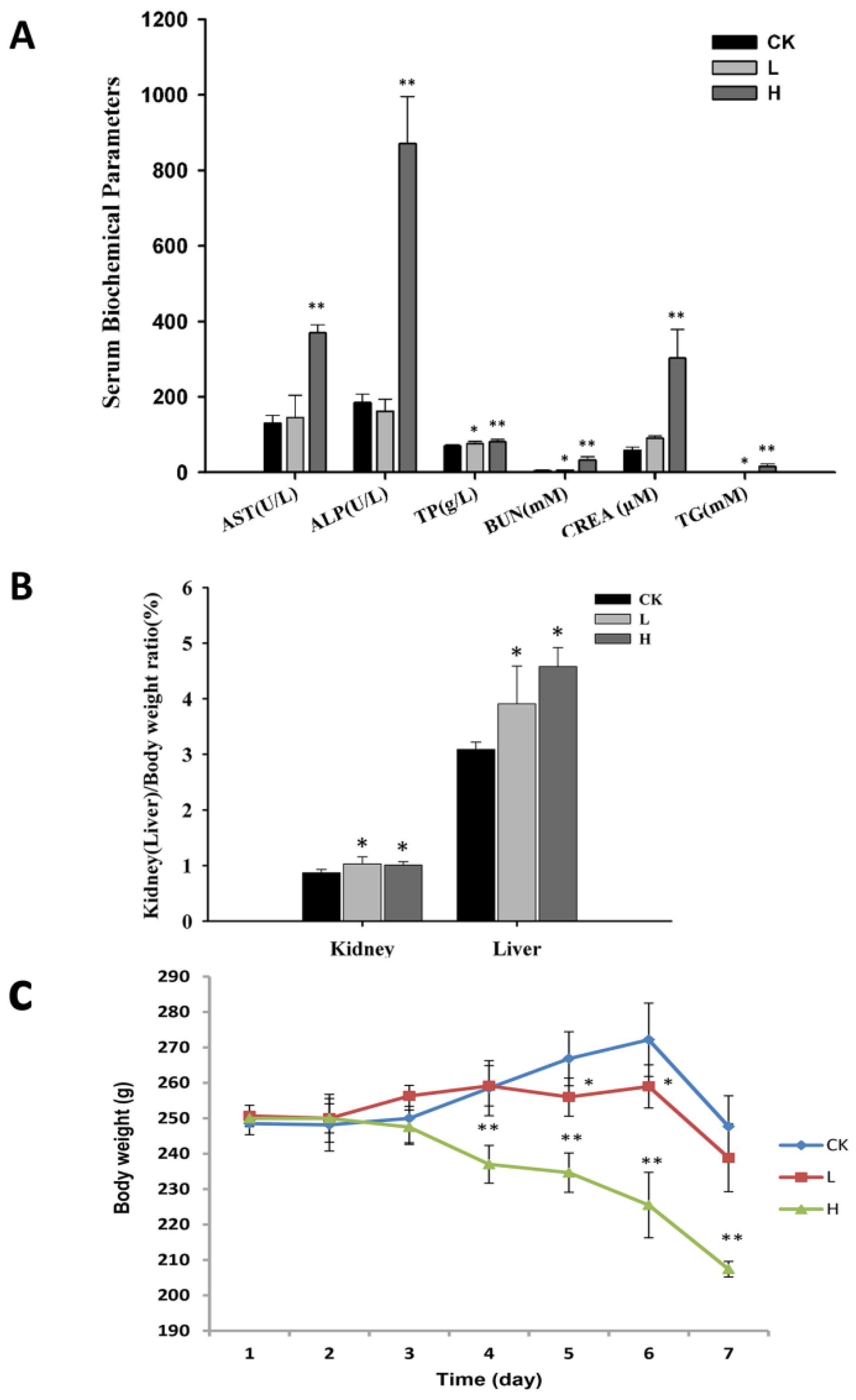
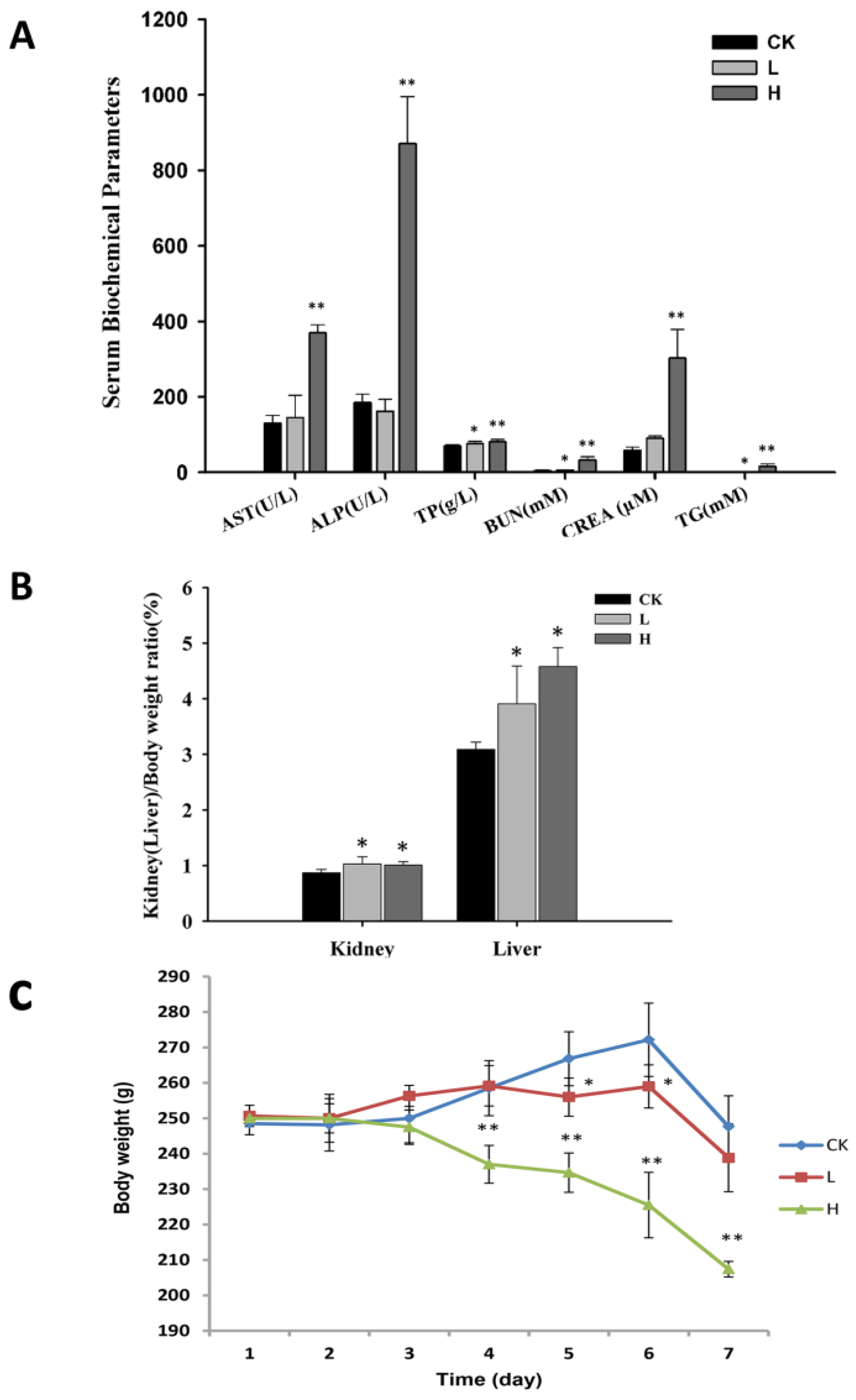
2.1. OTA Affects the Physiological Status of Rats
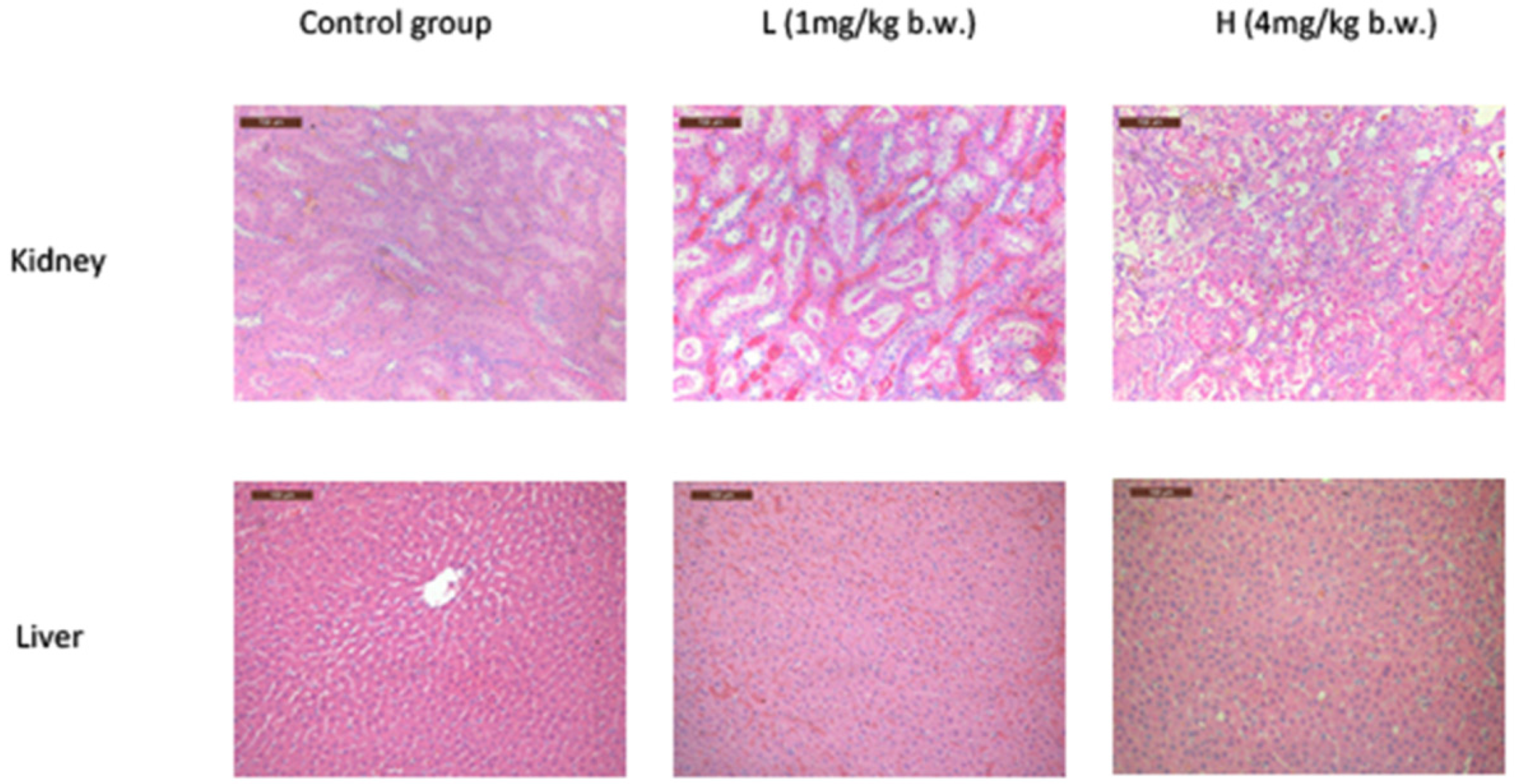
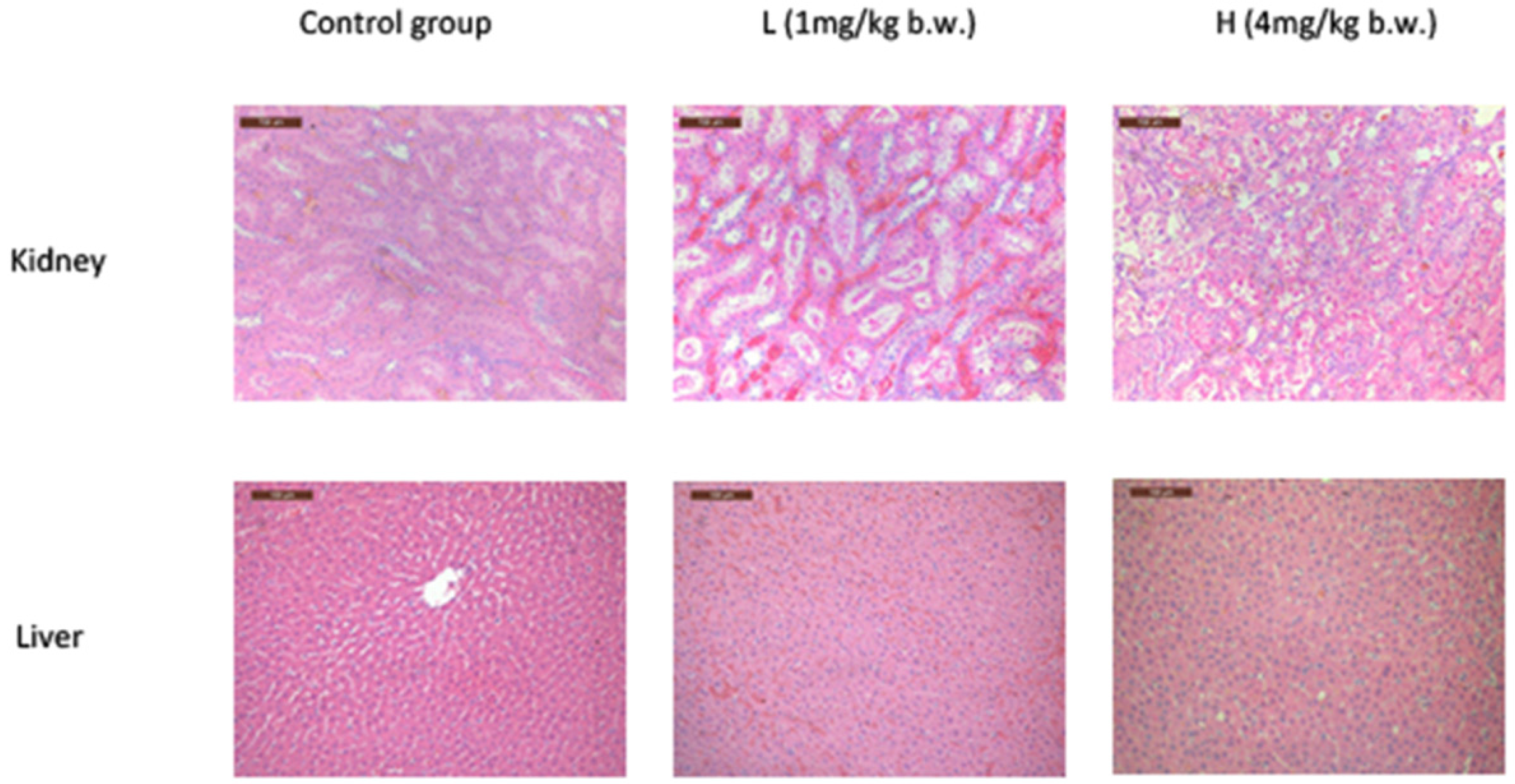
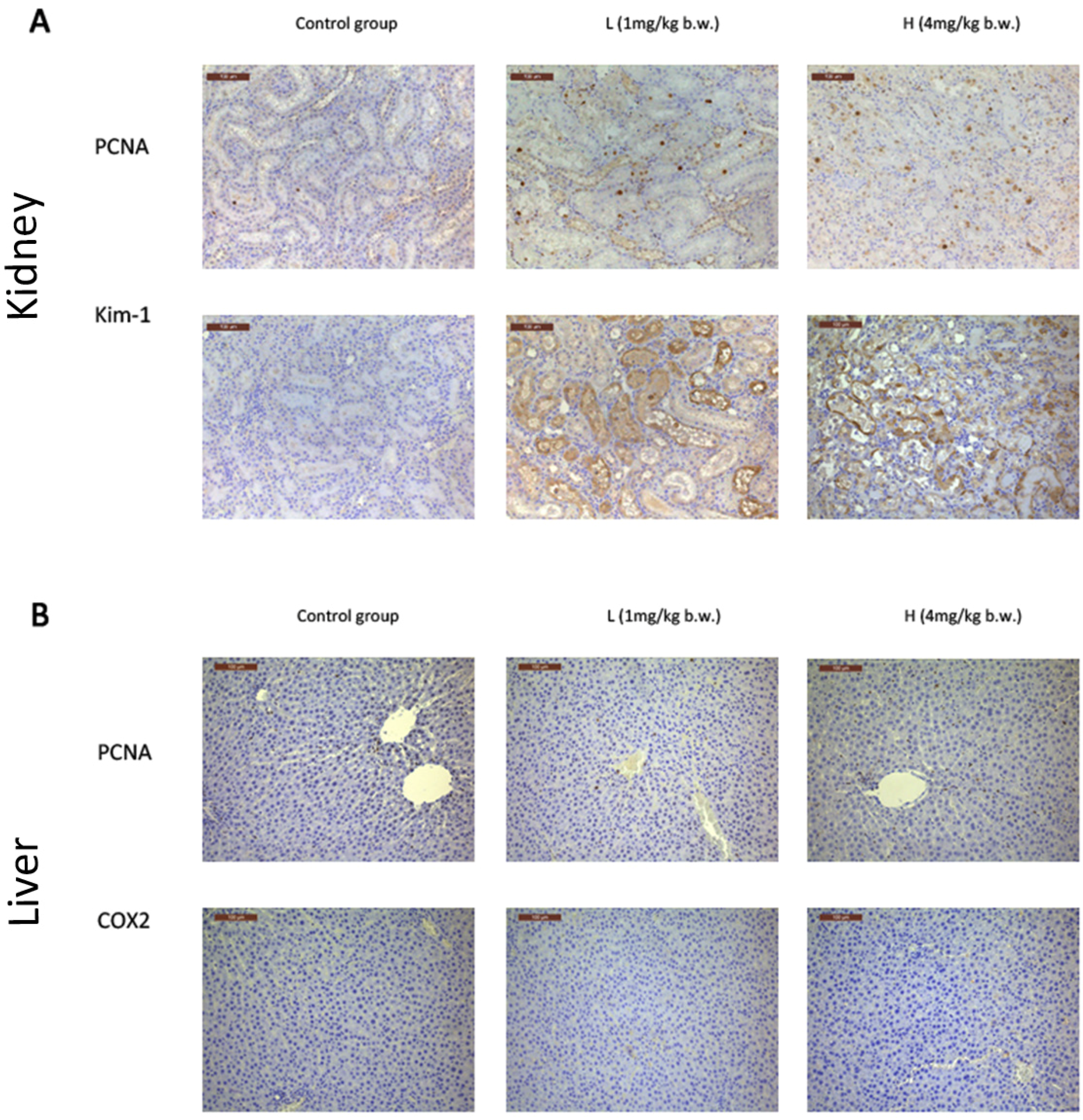
2.2. OTA Caused Kidney Damage in Rats
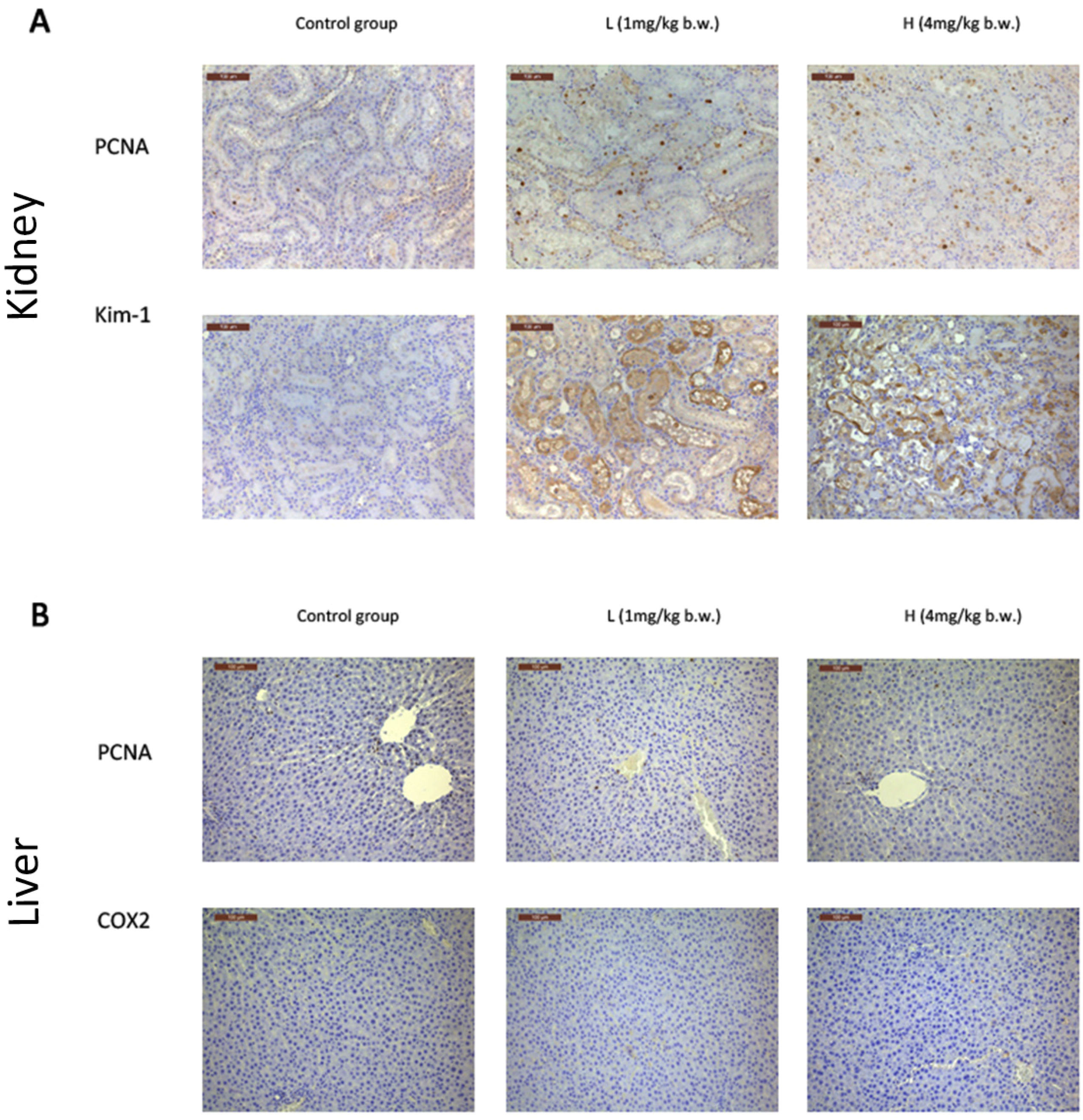
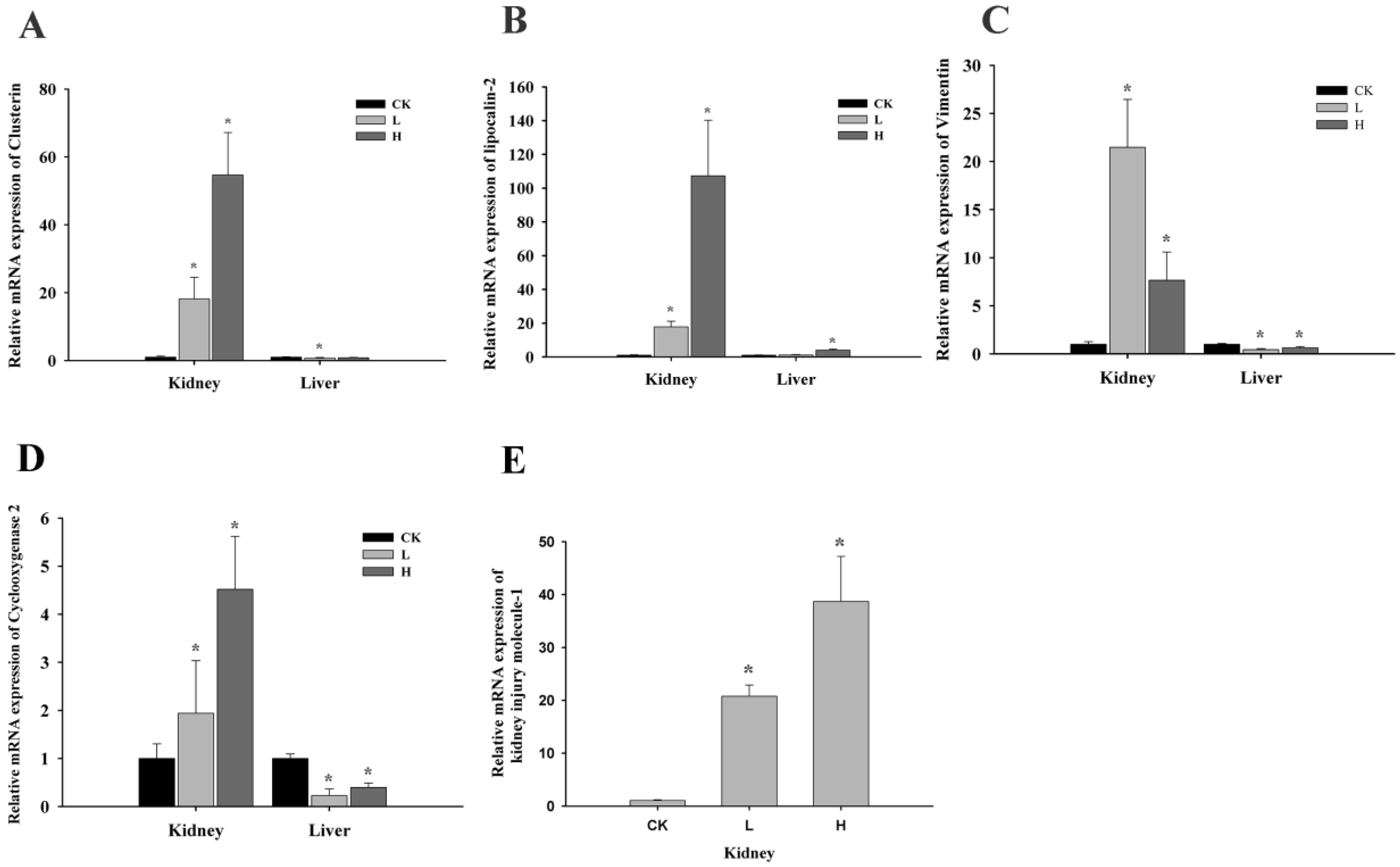
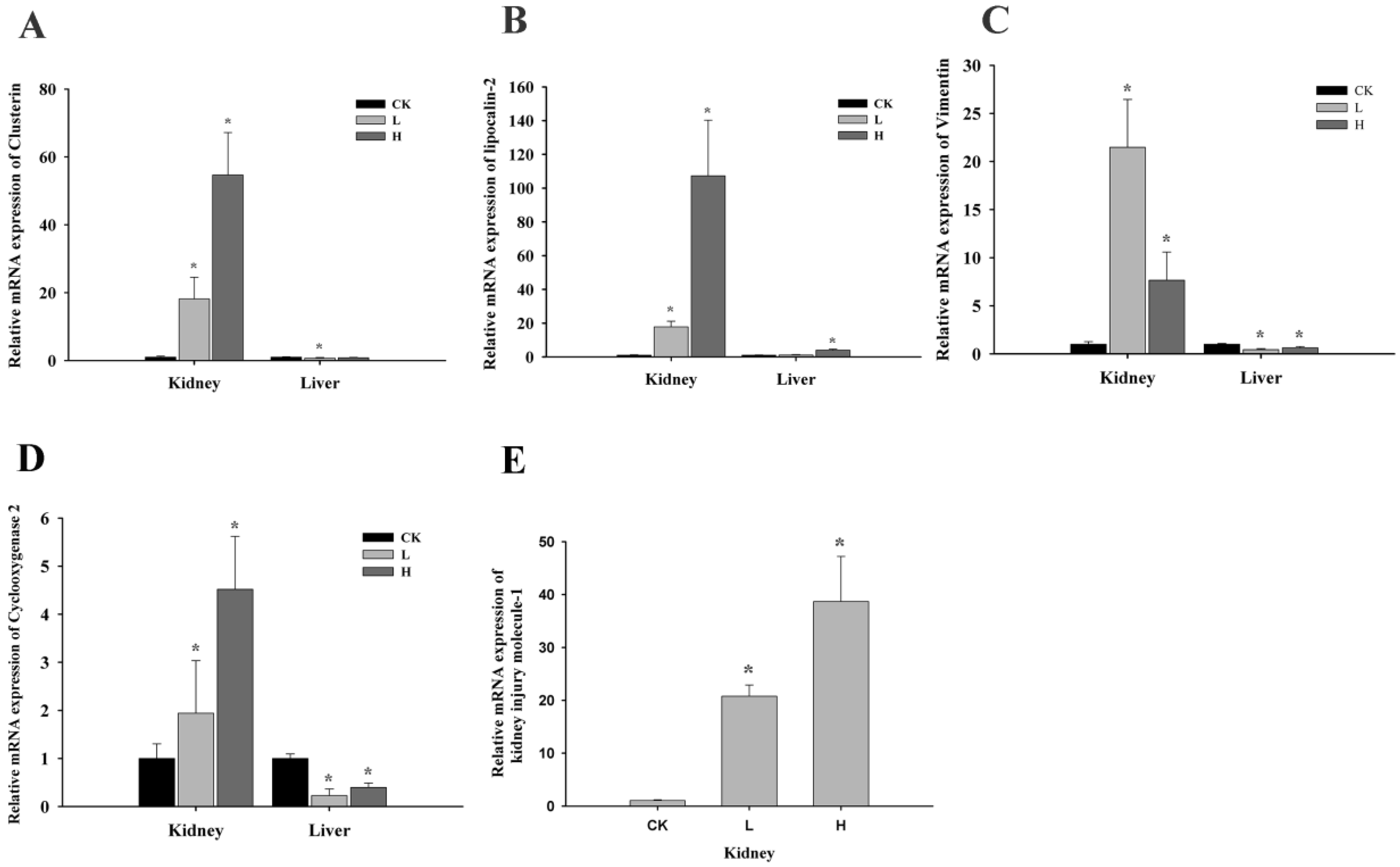
2.3. OTA Induced Putative Biomarkers of Nephrotoxicity
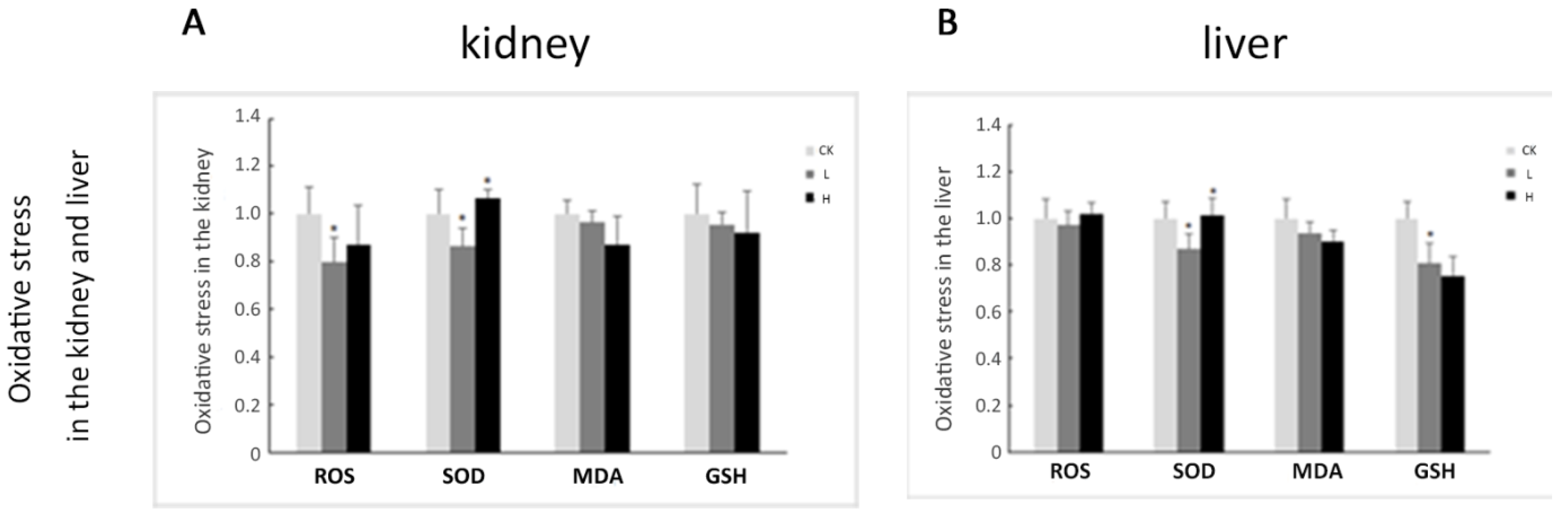
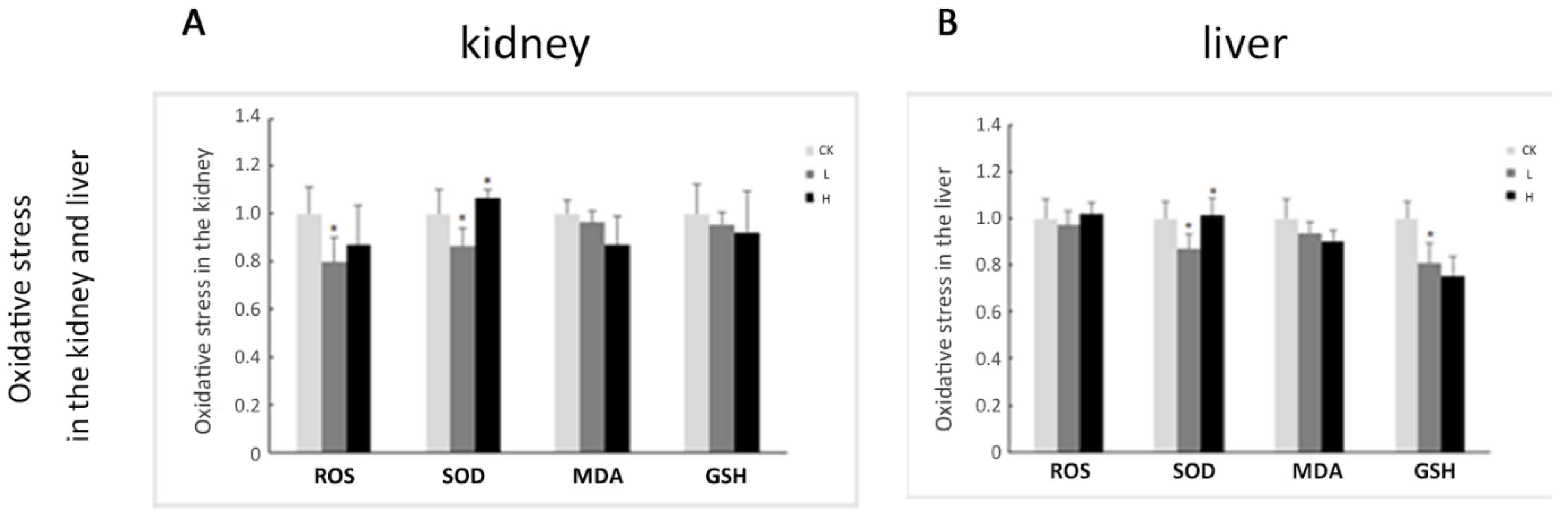
2.4. Oxidative Stress Plays a Limited Role in OTA-Induced Rat Acute Kidney Injury
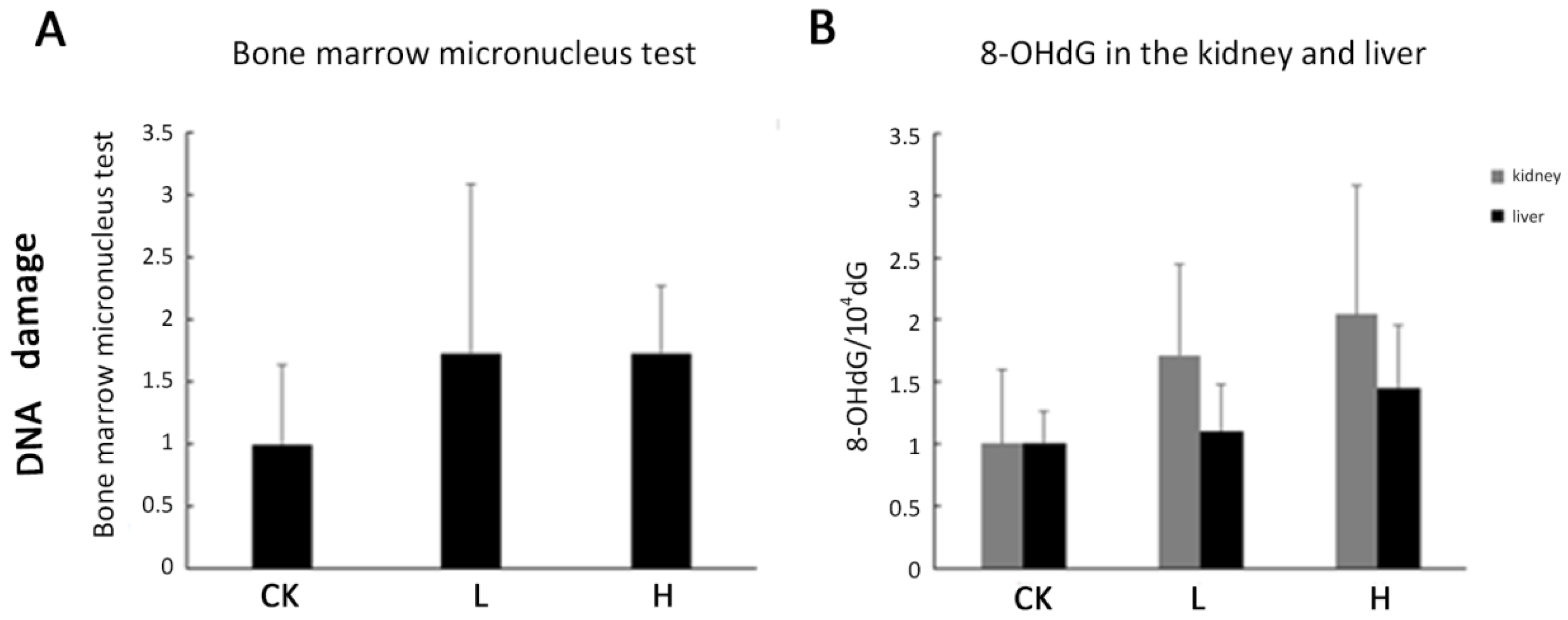
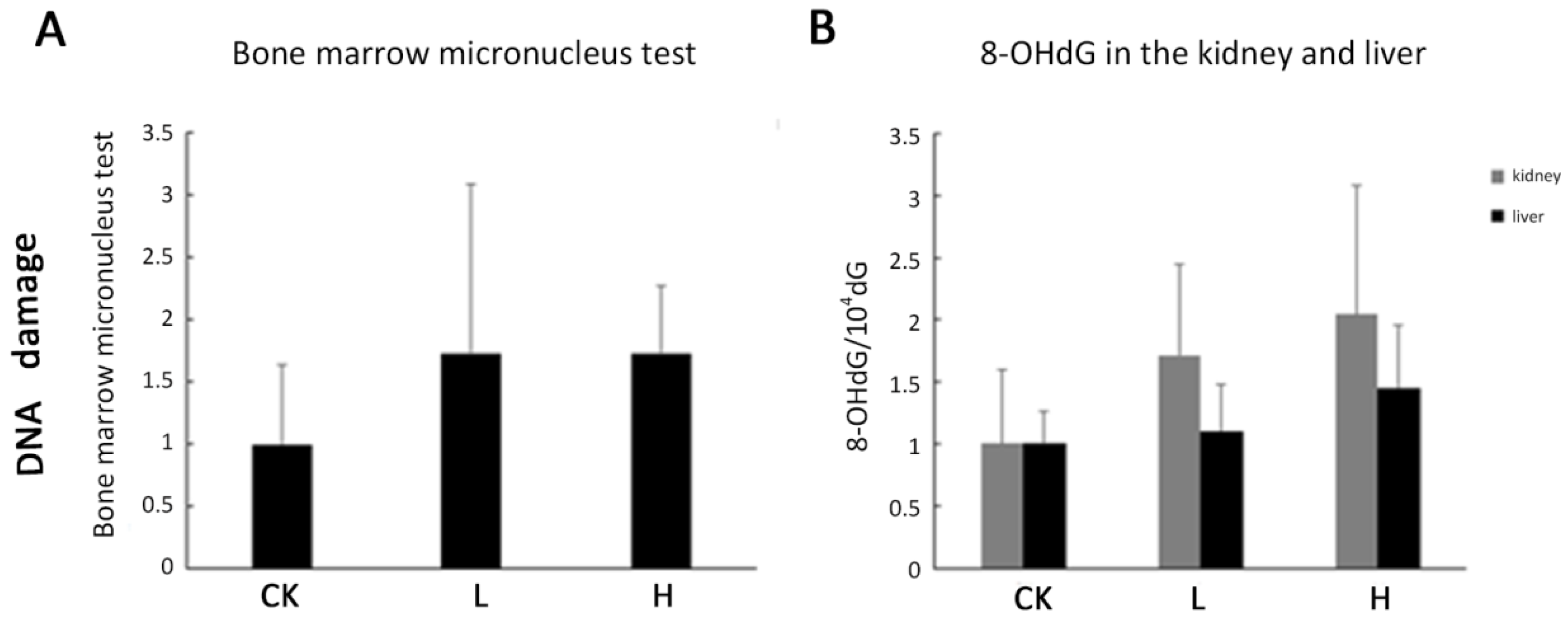
2.5. DNA Damage Was Not Observed in Acute Toxicity
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals
4.3. Immunohistochemistry
4.4. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.5. Determination of Oxidation-Related Parameters
4.6. Analysis of 8-Hydroxydeoxyguanosine (8-OHdG)
4.7. Comet Assay
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| OTA | Ochratoxin A |
| AST | aspartate aminotransferase |
| ALP | alkaline phosphatase |
| TP | total serum protein |
| TG | triacylglycerol |
| BUN | blood urea nitrogen |
| ALT | alanine aminotransferase |
| ALB | albumin |
| CREA | creatinine |
| GLU | glucose |
| PCNA | proliferating cell nuclear antigen |
| Kim-1 | kidney injury molecule-1 |
| Cox-2 | Cyclooxygenase 2 |
| Lcn2 | lipocalin-2 |
| Clu | Clusterin |
| Vim | Vimentin |
| ROS | Reactive Oxygen Species |
| MNT | marrow micronucleus test |
| GSH | reduced glutathione |
| SOD | superoxide dismutase |
| MDA | malonaldehyde |
| PI | propidium iodide |
| LDH | lactate dehydrogenase |
| 8-OHdG | 8-hydroxydeoxyguanosine |
| CHO/CHOz | cholesterol |
References
- Kamp, H.G.; Eisenbrand, G.; Janzowski, C.; Kiossev, J.; Latendresse, J.R.; Schlatter, J.; Turesky, R.J. Ochratoxin a induces oxidative DNA damage in liver and kidney after oral dosing to rats. Mol. Nutr. Food Res. 2005, 49, 1160–1167. [Google Scholar] [CrossRef] [PubMed]
- Van der Merwe, K.J.; Steyn, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin a, a toxic metabolite produced by aspergillus ochraceus wilh. Nature 1965, 205, 1112–1113. [Google Scholar] [CrossRef] [PubMed]
- Wilk-Zasadna, I.; Minta, M. Developmental toxicity of ochratoxin A in rat embryo midbrain micromass cultures. Int. J. Mol. Sci. 2009, 10, 37–49. [Google Scholar] [PubMed]
- Krogh, P.; Preusser, H.-J. Causal associations of mycotoxic nephropathy. Zent. Bakteriol. Parasitenkd. Infekt. Hyg. IA 1980, 45, 291–300. [Google Scholar]
- Kanisawa, M.; Suzuki, S. Induction of renal and hepatic tumors in mice by ochratoxin A, a mycotoxin. Gan 1978, 69, 599–600. [Google Scholar] [PubMed]
- O’Brien, E.; Dietrich, D.R. Ochratoxin a: The continuing enigma. Crit. Rev. Toxicol. 2005, 35, 33–60. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, R.R.; Frohlich, A.A. A review of recent advances in understanding ochratoxicosis. J. Anim. Sci. 1992, 70, 3968–3988. [Google Scholar] [CrossRef] [PubMed]
- Lautert, C.; Ferreiro, L.; Wolkmer, P.; Paim, F.C.; da Silva, C.B.; Jaques, J.A.; Lopes, S.T.; Santurio, J.M. Individual in vitro effects of ochratoxin A, deoxynivalenol and zearalenone on oxidative stress and acetylcholinesterase in lymphocytes of broiler chickens. Springerplus 2014, 3. [Google Scholar] [CrossRef] [PubMed]
- Aleo, M.D.; Wyatt, R.D.; Schnellmann, R.G. Mitochondrial dysfunction is an early event in ochratoxin A but not oosporein toxicity to rat renal proximal tubules. Toxicol. Appl. Pharmacol. 1991, 107, 73–80. [Google Scholar] [CrossRef]
- Li, X.; Gao, J.; Huang, K.; Qi, X.; Dai, Q.; Mei, X.; Xu, W. Dynamic changes of global DNA methylation and hypermethylation of cell adhesion-related genes in rat kidneys in response to ochratoxin A. World Mycotoxin J. 2015, 8, 465–476. [Google Scholar] [CrossRef]
- Qi, X.; Yang, X.; Chen, S.; He, X.; Dweep, H.; Guo, M.; Cheng, W.-H.; Xu, W.; Luo, Y.; Gretz, N. Ochratoxin A induced early hepatotoxicity: New mechanistic insights from microRNA, mRNA and proteomic profiling studies. Sci. Rep. 2014. [Google Scholar] [CrossRef]
- Guo, M.; Huang, K.; Chen, S.; Qi, X.; He, X.; Cheng, W.-H.; Luo, Y.; Xia, K.; Xu, W. Combination of metagenomics and culture-based methods to study the interaction between ochratoxin A and gut microbiota. Toxicol. Sci. 2014, 141, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Gagliano, N.; Torri, C.; Donetti, E.; Grizzi, F.; Costa, F.; Bertelli, A.A.; Migliori, M.; Filippi, C.; Bedoni, M.; Panichi, V.; et al. Ochratoxin a-induced renal cortex fibrosis and epithelial-to-mesenchymal transition: Molecular mechanisms of ochratoxin A-injury and potential effects of red wine. Mol. Med. 2005, 11, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Gagliano, N.; Donne, I.D.; Torri, C.; Migliori, M.; Grizzi, F.; Milzani, A.; Filippi, C.; Annoni, G.; Colombo, P.; Costa, F.; et al. Early cytotoxic effects of ochratoxin A in rat liver: A morphological, biochemical and molecular study. Toxicology 2006, 225, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Faucet, V.; Pfohl-Leszkowicz, A.; Dai, J.; Castegnaro, M.; Manderville, R.A. Evidence for covalent DNA adduction by ochratoxin A following chronic exposure to rat and subacute exposure to pig. Chem. Res. Toxicol. 2004, 17, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- Hibi, D.; Kijima, A.; Kuroda, K.; Suzuki, Y.; Ishii, Y.; Jin, M.; Nakajima, M.; Sugita-Konishi, Y.; Yanai, T.; Nohmi, T. Molecular mechanisms underlying ochratoxin A-induced genotoxicity: Global gene expression analysis suggests induction of DNA double-strand breaks and cell cycle progression. J. Toxicol. Sci. 2013, 38, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Pfohl-Leszkowicz, A.; Manderville, R.A. An update on direct genotoxicity as a molecular mechanism of ochratoxin A carcinogenicity. Chem. Res. Toxicol. 2011, 25, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Arbillaga, L.; Azqueta, A.; Ezpeleta, O.; Lopez de Cerain, A. Oxidative DNA damage induced by ochratoxin A in the hk-2 human kidney cell line: Evidence of the relationship with cytotoxicity. Mutagenesis 2007, 22, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Palma, N.; Cinelli, S.; Sapora, O.; Wilson, S.H.; Dogliotti, E. Ochratoxin a-induced mutagenesis in mammalian cells is consistent with the production of oxidative stress. Chem. Res. Toxicol. 2007, 20, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Hundhausen, C.; boesch-saadatmandi, C.; matzner, N.; lang, F.; blank, R.; wolffram, S.; blaschek, W.; Rimbach, G. Ochratoxin a lowers mrna levels of genes encoding for key proteins of liver cell metabolism. Cancer Genom.-Proteom. 2008, 5, 319–332. [Google Scholar]
- Shen, X.L.; Zhang, Y.; Xu, W.; Liang, R.; Zheng, J.; Luo, Y.; Wang, Y.; Huang, K. An itraq-based mitoproteomics approach for profiling the nephrotoxicity mechanisms of ochratoxin A in hek 293 cells. J. Proteom. 2013, 78, 398–415. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhang, Y.; Xu, W.; Luo, Y.; Hao, J.; Shen, X.L.; Yang, X.; Li, X.; Huang, K. Zinc protects hepg2 cells against the oxidative damage and DNA damage induced by ochratoxin A. Toxicol. Appl. Pharmacol. 2013, 268, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Cavin, C.; Delatour, T.; Marin-Kuan, M.; Holzhäuser, D.; Higgins, L.; Bezencon, C.; Guignard, G.; Junod, S.; Richoz-Payot, J.; Gremaud, E. Reduction in antioxidant defenses may contribute to ochratoxin A toxicity and carcinogenicity. Toxicol. Sci. 2007, 96, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Marin-Kuan, M.; Ehrlich, V.; Delatour, T.; Cavin, C.; Schilter, B. Evidence for a role of oxidative stress in the carcinogenicity of ochratoxin A. J. Toxicol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Utan, A.; Cervellati, R.; Speroni, E.; Guerra, M. Catechins: Natural free-radical scavengers against ochratoxin A-induced cell damage in a pig kidney cell line (llc-pk1). Food Chem. Toxicol. 2007, 45, 1910–1917. [Google Scholar] [CrossRef] [PubMed]
- Fusi, E.; Rebucci, R.; Pecorini, C.; Campagnoli, A.; Pinotti, L.; Saccone, F.; Cheli, F.; Purup, S.; Sejrsen, K.; Baldi, A. Alpha-tocopherol counteracts the cytotoxicity induced by ochratoxin A in primary porcine fibroblasts. Toxins 2010, 2, 1265–1278. [Google Scholar] [CrossRef] [PubMed]
- Program, N.T. Toxicology and carcinogenesis studies of ochratoxin A (CAS no. 303-47-9) in f344/n rats (gavage studies). Natl. Toxicol. Program Tech. Rep. Ser. 1989, 358, 1–142. [Google Scholar]
- Meki, A.-R.M.; Hussein, A.A. Melatonin reduces oxidative stress induced by ochratoxin A in rat liver and kidney. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 130, 305–313. [Google Scholar] [CrossRef]
- Petrik, J.; Žanić-Grubišić, T.; Barišić, K.; Pepeljnjak, S.; Radić, B.; Ferenčić, Ž.; Čepelak, I. Apoptosis and oxidative stress induced by ochratoxin A in rat kidney. Arch. Toxicol. 2003, 77, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Yu, T.; Zhu, L.; Gao, J.; He, X.; Huang, K.; Luo, Y.; Xu, W. Ochratoxin a induces rat renal carcinogenicity with limited induction of oxidative stress responses. Toxicol. Appl. Pharmacol. 2014, 280, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Hong, H.; Senior, J.; Tong, W. Biomarkers for drug-induced liver injury. Expert Rev. Gastroenterol. Hepatol. 2010, 4, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Hall, P.; Levison, D.; Woods, A.; Yu, C.W.; Kellock, D.; Watkins, J.; Barnes, D.; Gillett, C.; Camplejohn, R.; Dover, R. Proliferating cell nuclear antigen (pcna) immunolocalization in paraffin sections: An index of cell proliferation with evidence of deregulated expression in some, neoplasms. J. Pathol. 1990, 162, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Bonventre, J.V. Kidney injury molecule-1 (kim-1): A specific and sensitive biomarker of kidney injury. Scand. J. Clin. Lab. Investig. Suppl. 2008, 241, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Liochev, S.I. Reactive oxygen species and the free radical theory of aging. Free Radic. Biol. Med. 2013, 60, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Creppy, E.-E.; Röschenthaler, R.; Dirheimer, G. Inhibition of protein synthesis in mice by ochratoxin A and its prevention by phenylalanine. Food Chem. Toxicol. 1984, 22, 883–886. [Google Scholar] [CrossRef]
- Creppy, E.; Kane, A.; Dirheimer, G.; Lafarge-Frayssinet, C.; Mousset, S.; Frayssinet, C. Genotoxicity of ochratoxin A in mice: DNA single-strand break evaluation in spleen, liver and kidney. Toxicol. Lett. 1985, 28, 29–35. [Google Scholar] [CrossRef]
- Pfohl-Leszkowicz, A.; Chakor, K.; Creppy, E.; Dirheimer, G. DNA adduct formation in mice treated with ochratoxin A. IARC Sci. Publ. 1990, 115, 245–253. [Google Scholar]
- Dvergsten, J.; Manivel, J.C.; Correa-Rotter, R.; Rosenberg, M.E. Expression of clusterin in human renal diseases. Kidney Int. 1994, 45, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, T.; Hung, C.C.; Yang, S.A.; Stevens, J.L.; Bonventre, J.V. Kidney injury molecule-1: A tissue and urinary biomarker for nephrotoxicant-induced renal injury. Am. J. Physiol. Ren. Physiol. 2004, 286, F552–F563. [Google Scholar] [CrossRef] [PubMed]
- Viau, A.; El Karoui, K.; Laouari, D.; Burtin, M.; Nguyen, C.; Mori, K.; Pillebout, E.; Berger, T.; Mak, T.W.; Knebelmann, B.; et al. Lipocalin 2 is essential for chronic kidney disease progression in mice and humans. J. Clin. Investig. 2010, 120, 4065–4076. [Google Scholar] [CrossRef] [PubMed]
- Wojcik, M.; Ramadori, P.; Blaschke, M.; Sultan, S.; Khan, S.; Malik, I.A.; Naz, N.; Martius, G.; Ramadori, G.; Schultze, F.C. Immunodetection of cyclooxygenase-2 (cox-2) is restricted to tissue macrophages in normal rat liver and to recruited mononuclear phagocytes in liver injury and cholangiocarcinoma. Histochem. Cell Biol. 2012, 137, 217–233. [Google Scholar] [CrossRef] [PubMed]
- Rached, E.; Hoffmann, D.; Blumbach, K.; Weber, K.; Dekant, W.; Mally, A. Evaluation of putative biomarkers of nephrotoxicity after exposure to ochratoxin A in vivo and in vitro. Toxicol. Sci. 2008, 103, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Mally, A.; Volkel, W.; Amberg, A.; Kurz, M.; Wanek, P.; Eder, E.; Hard, G.; Dekant, W. Functional, biochemical, and pathological effects of repeated oral administration of ochratoxin A to rats. Chem. Res. Toxicol. 2005, 18, 1242–1252. [Google Scholar] [CrossRef] [PubMed]
- Rached, E.; Hard, G.C.; Blumbach, K.; Weber, K.; Draheim, R.; Lutz, W.K.; Ozden, S.; Steger, U.; Dekant, W.; Mally, A. Ochratoxin a: 13-week oral toxicity and cell proliferation in male f344/n rats. Toxicol. Sci. 2007, 97, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Il’ichev, Y.V.; Perry, J.L.; Ruker, F.; Dockal, M.; Simon, J.D. Interaction of ochratoxin A with human serum albumin. Binding sites localized by competitive interactions with the native protein and its recombinant fragments. Chem.-Biol. Interact. 2002, 141, 275–293. [Google Scholar] [CrossRef]
- Kumagai, S. Ochratoxin a: Plasma concentration and excretion into bile and urine in albumin-deficient rats. Food Chem. Toxicol. 1985, 23, 941–943. [Google Scholar] [CrossRef]
- Bolt, H.M.; Foth, H.; Hengstler, J.G.; Degen, G.H. Carcinogenicity categorization of chemicals-new aspects to be considered in a european perspective. Toxicol. Lett. 2004, 151, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Gautier, J.-C.; Holzhaeuser, D.; Markovic, J.; Gremaud, E.; Schilter, B.t.; Turesky, R.J. Oxidative damage and stress response from ochratoxin A exposure in rats. Free Radic. Biol. Med. 2001, 30, 1089–1098. [Google Scholar] [CrossRef]
- Kane, A.; Creppy, E.E.; Roth, A.; Röschenthaler, R.; Dirheimer, G. Distribution of the [3h]-label from low doses of radioactive ochratoxin A ingested by rats, and evidence for DNA single-strand breaks caused in liver and kidneys. Arch. Toxicol. 1986, 58, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Zeljezic, D.; Domijan, A.-M.; Peraica, M. DNA damage by ochratoxin A in rat kidney assessed by the alkaline comet assay. Braz. J. Med. Biol. Res. 2006, 39, 1563–1568. [Google Scholar] [CrossRef] [PubMed]
- Mally, A. Ochratoxin A and mitotic disruption: Mode of action analysis of renal tumor formation by ochratoxin A. Toxicol. Sci. 2012, 127, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Shi, L.; Huang, K.; Xu, W. Protective effect of n-acetylcysteine against DNA damage and s-phase arrest induced by ochratoxin A in human embryonic kidney cells (hek-293). Food Chem. Toxicol. 2014, 70, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Hibi, D.; Suzuki, Y.; Ishii, Y.; Jin, M.; Watanabe, M.; Sugita-Konishi, Y.; Yanai, T.; Nohmi, T.; Nishikawa, A.; Umemura, T. Site-specific in vivo mutagenicity in the kidney of gpt delta rats given a carcinogenic dose of ochratoxin A. Toxicol. Sci. 2011, 122, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, K.; Hibi, D.; Ishii, Y.; Takasu, S.; Kijima, A.; Matsushita, K.; Masumura, K.; Watanabe, M.; Sugita-Konishi, Y.; Sakai, H.; et al. Ochratoxin a induces DNA double-strand breaks and large deletion mutations in the carcinogenic target site of gpt delta rats. Mutagenesis 2014, 29, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Redon, C.E.; Nakamura, A.J.; Martin, O.A.; Parekh, P.R.; Weyemi, U.S.; Bonner, W.M. Recent developments in the use of γ-H2AX as a quantitative DNA double-strand break biomarker. Aging (Albany NY) 2011, 3, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Scully, R.; Xie, A. Double strand break repair functions of histone H2AX. Mutat. Res. 2013, 750, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Taniai, E.; Yafune, A.; Nakajima, M.; Hayashi, S.-M.; Nakane, F.; Itahashi, M.; Shibutani, M. Ochratoxin A induces karyomegaly and cell cycle aberrations in renal tubular cells without relation to induction of oxidative stress responses in rats. Toxicol. Lett. 2014, 224, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Litvak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Nakae, D.; Mizumoto, Y.; Kobayashi, E.; Noguchi, O.; Konishi, Y. Improved genomic/nuclear DNA extraction for 8-hydroxydeoxyguanosine analysis of small amounts of rat liver tissue. Cancer Lett. 1995, 97, 233–239. [Google Scholar] [CrossRef]
- Jackson, P.; Pedersen, L.M.; Kyjovska, Z.O.; Jacobsen, N.R.; Saber, A.T.; Hougaard, K.S.; Vogel, U.; Wallin, H. Validation of freezing tissues and cells for analysis of DNA strand break levels by comet assay. Mutagenesis 2013, 28, 699–707. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Brain | Kidney | Liver | Spleen | Heart | Thymus | Adrenal Gland | Testis |
|---|---|---|---|---|---|---|---|---|
| CK | 0.76 ± 0.05 | 0.87 ± 0.06 | 3.09 ± 0.13 | 0.26 ± 0.04 | 0.4 ± 0.03 | 0.22 ± 0.07 | 0.034 ± 0.005 | 1.3 ± 0.08 |
| L | 0.85 ± 0.04 a | 1.03 ± 0.13 a | 3.91 ± 0.68 a | 0.25 ± 0.04 | 0.44 ± 0.11 | 0.22 ± 0.05 | 0.041 ± 0.007 | 1.31 ± 0.12 |
| H | 0.94 ± 0.05 ab | 1.01 ± 0.06 a | 4.58 ± 0.34 a | 0.29 ± 0.08 | 0.42 ± 0.03 | 0.12 ± 0.04 b | 0.063 ± 0.004 ab | 1.58 ± 0.18 ab |
| Content | Control | L | H |
|---|---|---|---|
| ALT (U/L) | 30.67 ± 8.76 | 45.75 ± 32.4 | 63.33 ± 35.81 |
| AST (U/L) | 131 ± 20 | 145 ± 59 | 370 ± 21 ab |
| TP (g/L) | 70.35 ± 3.76 | 76.85 ± 4.94 a | 80.95 ± 6.43 ab |
| ALB (g/L) | 29.78 ± 1.22 | 33.77 ± 1.69 a | 33.53 ± 5.66 a |
| ALP (U/L) | 185 ± 22 | 162 ± 32 | 871 ± 125 ab |
| GLU (mmol/L) | 2.14 ± 0.73 | 2.24 ± 0.25 | 8.39 ± 0.94 ab |
| BUN (mmol/L) | 5 ± 0.9 | 6.3 ± 0.8 a | 33.3 ± 8.2 ab |
| CREA (μmol/L) | 58 ± 9 | 91 ± 6 a | 303 ± 76 ab |
| CHOz(mg/dl) | 1.29 ± 0.18 | 1.03 ± 0.28 | 3.07 ± 2.21 |
| TG (mmol/L) | 0.68 ± 0.24 | 0.76 ± 0.3 | 15.32 ± 7.25 ab |
| LDH (U/L) | 1182 ± 392 | 702 ± 454 | 2785 ± 803 ab |
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| Clusterin | CACTACGGGCCTCTGAGCTT | ACGTCCATGGCCTGTTGAG |
| Vimentin | GATGCTCCAGAGGGAGGAAG | AAGGTCAAGACGTGCCAGAG |
| lipocalin2 | TCTGGGCCTCAAGGATAACAAC | AGACAGGTGGGACCTGAACCA |
| Kim-1 | TGGCACTGTGACATCCTCAGA | GCAACGGACATGCCAACATA |
| COX-2 | TGTATGCTACCATCTGGCTTCGG | GTTTGGAACAGTCGCTCGTCATC |
| β-actin | TCGTGCGTGACATTAAGGAG | AGGAAGGAAGGCTGGAAGAG |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, L.; Yu, T.; Qi, X.; Gao, J.; Huang, K.; He, X.; Luo, H.; Xu, W. Limited Link between Oxidative Stress and Ochratoxin A—Induced Renal Injury in an Acute Toxicity Rat Model. Toxins 2016, 8, 373. https://doi.org/10.3390/toxins8120373
Zhu L, Yu T, Qi X, Gao J, Huang K, He X, Luo H, Xu W. Limited Link between Oxidative Stress and Ochratoxin A—Induced Renal Injury in an Acute Toxicity Rat Model. Toxins. 2016; 8(12):373. https://doi.org/10.3390/toxins8120373
Chicago/Turabian StyleZhu, Liye, Tao Yu, Xiaozhe Qi, Jing Gao, Kunlun Huang, Xiaoyun He, Haoshu Luo, and Wentao Xu. 2016. "Limited Link between Oxidative Stress and Ochratoxin A—Induced Renal Injury in an Acute Toxicity Rat Model" Toxins 8, no. 12: 373. https://doi.org/10.3390/toxins8120373