
Epidermal Growth Factor Receptor Signaling Enhances the Proinflammatory Effects of Staphylococcus aureus Gamma-Toxin on the Mucosa

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
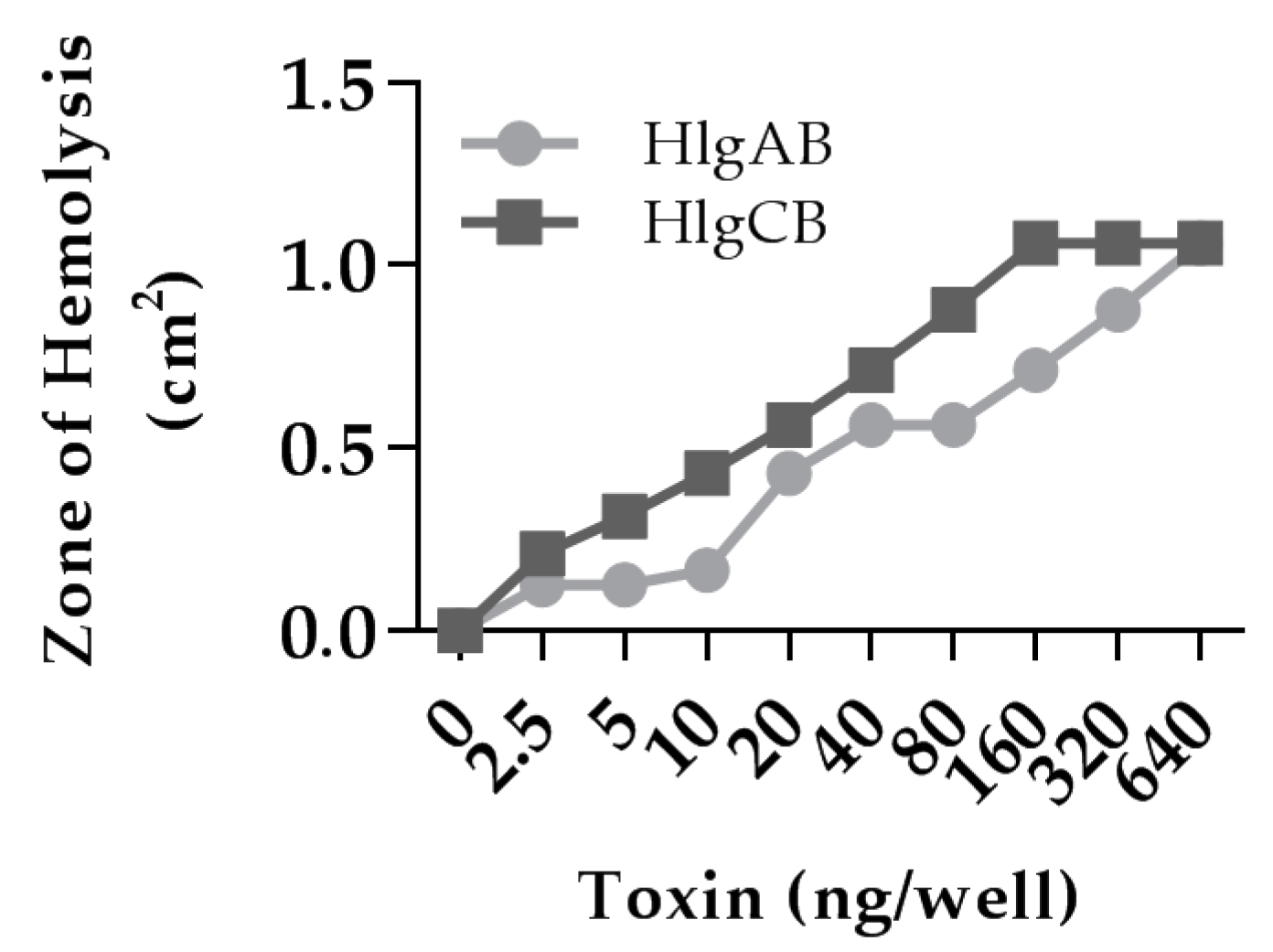
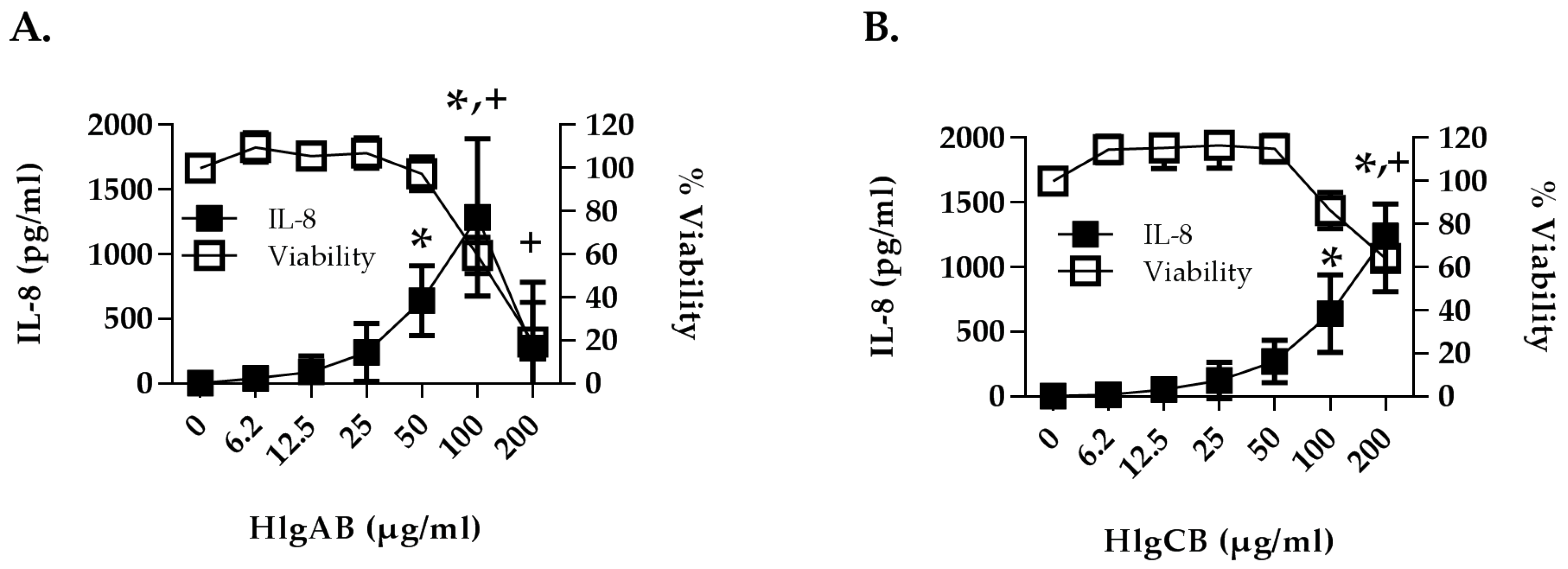
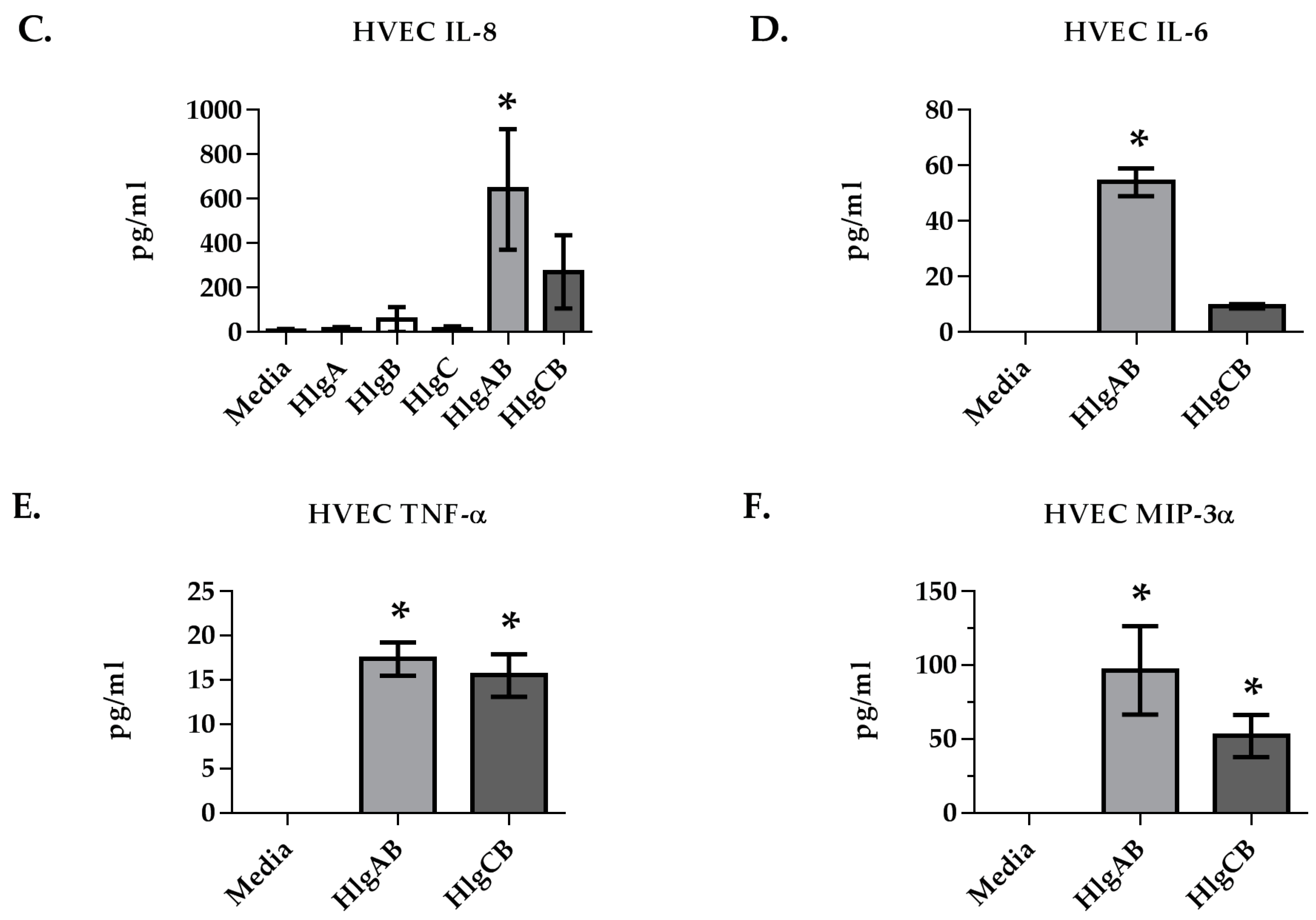
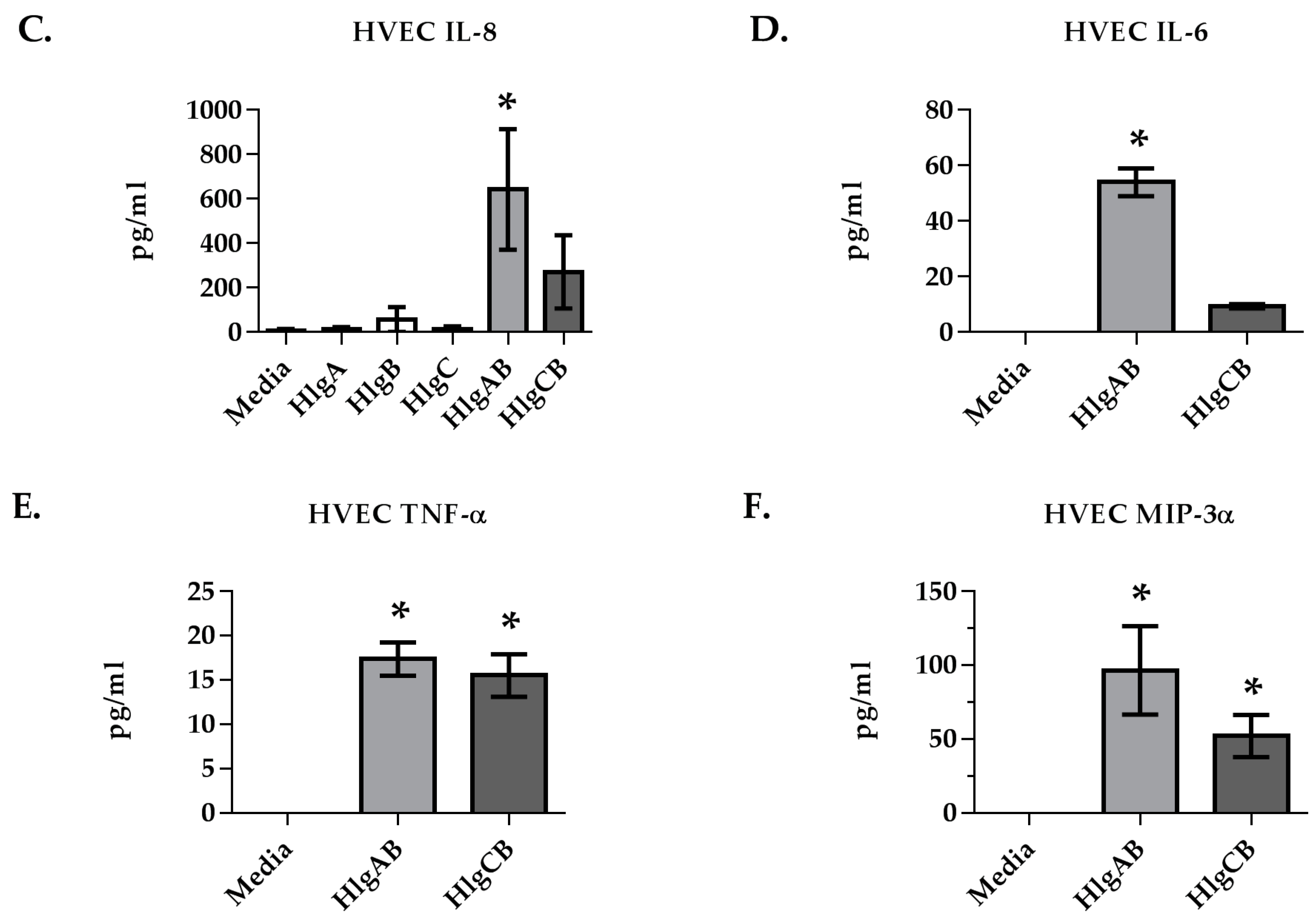
2.1. Gamma-Toxins Are Cytotoxic and Proinflammatory to HVECs
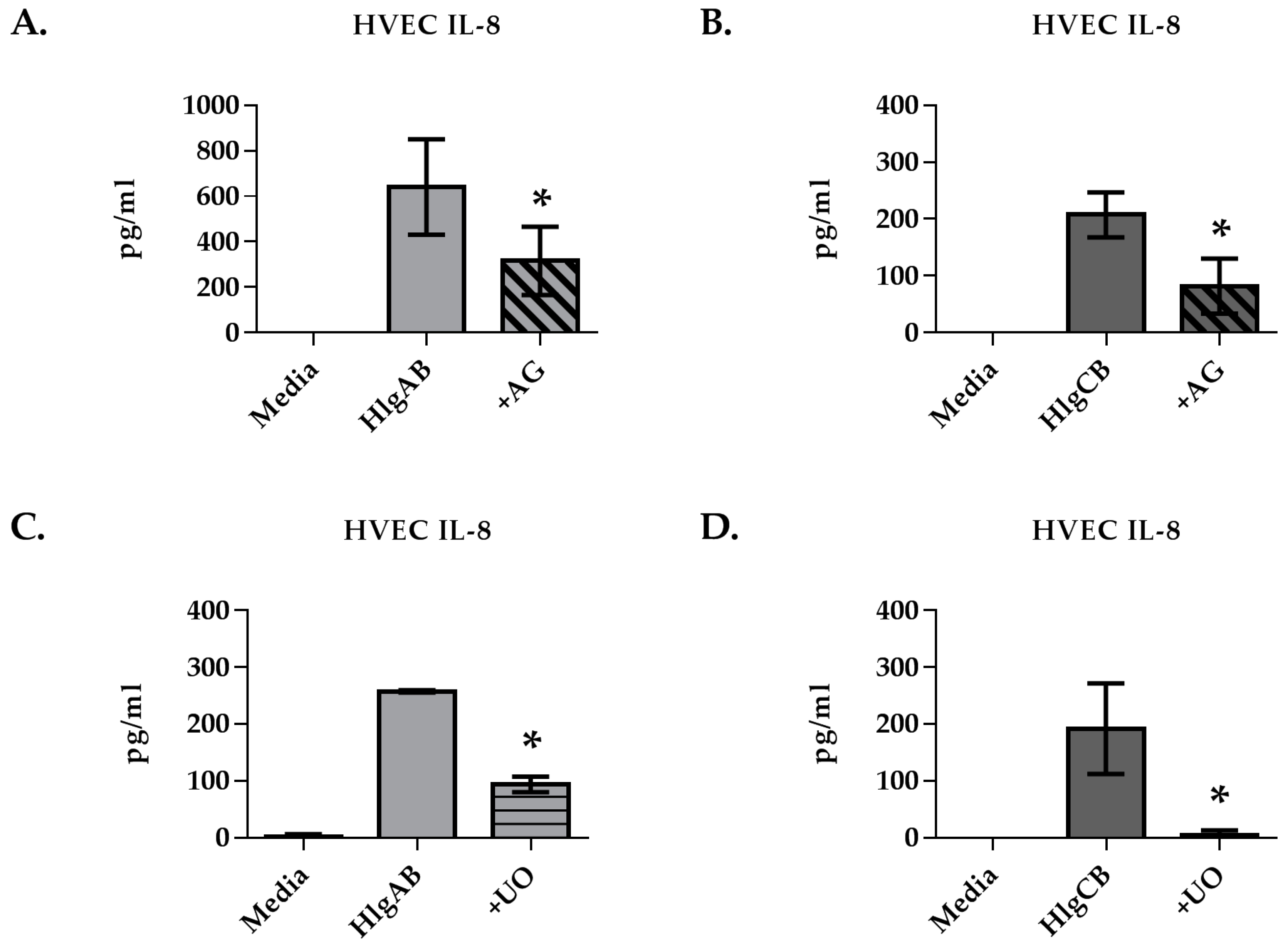
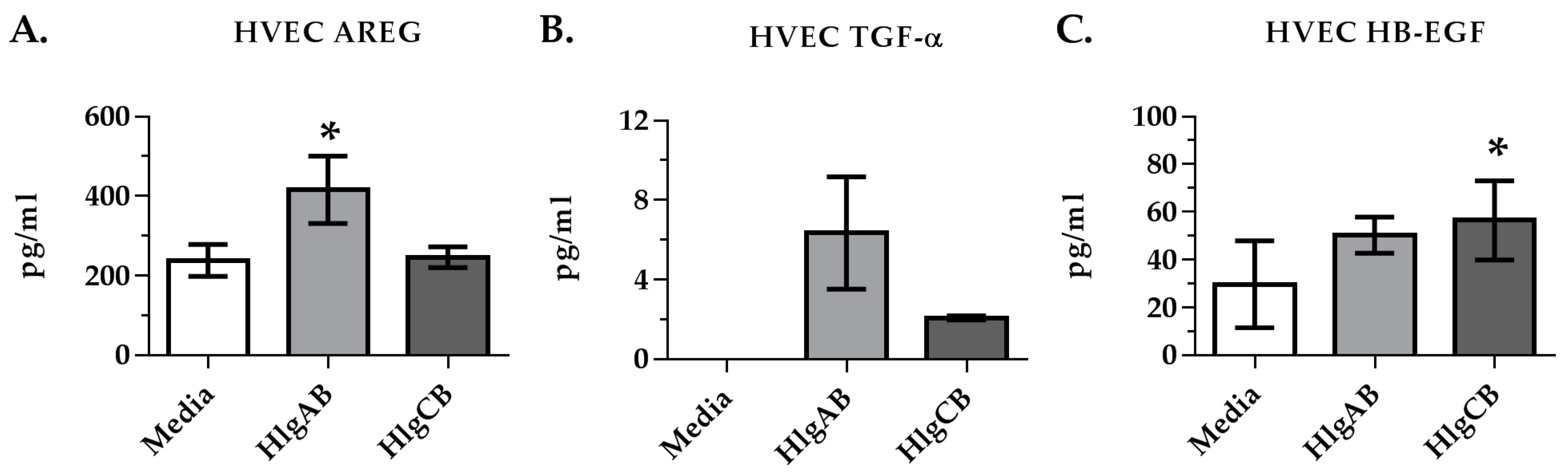
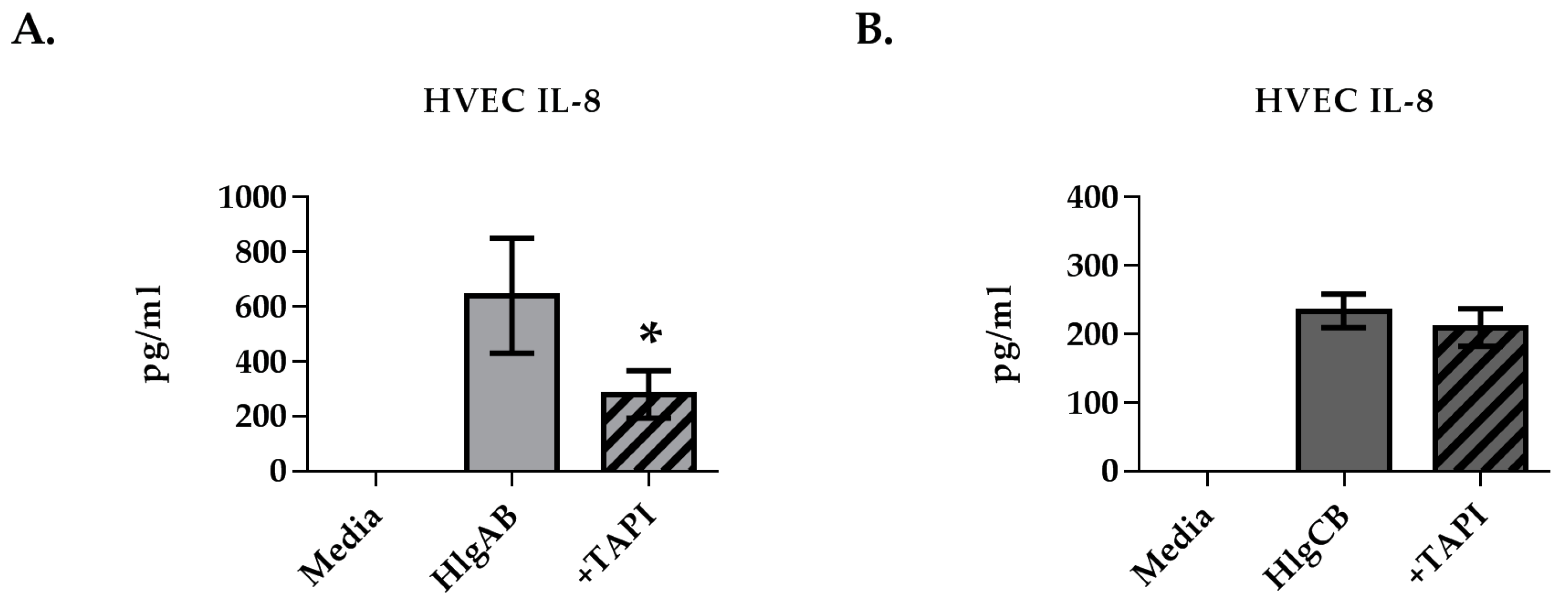
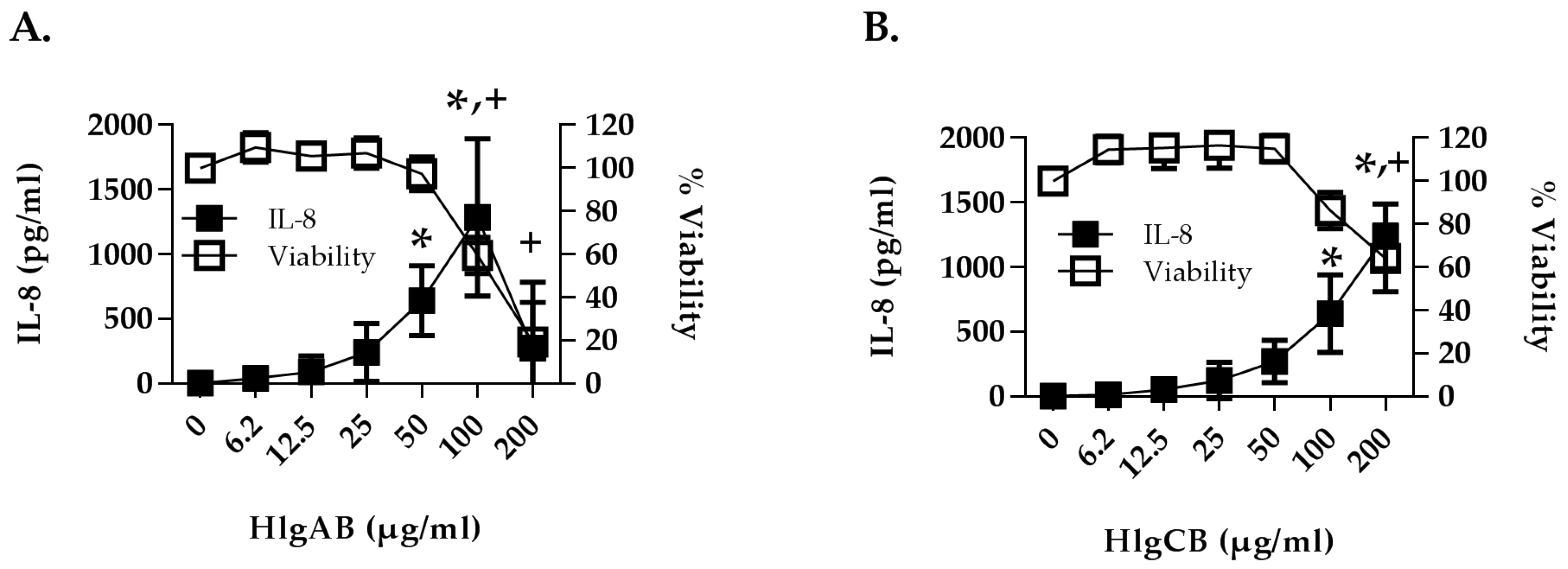
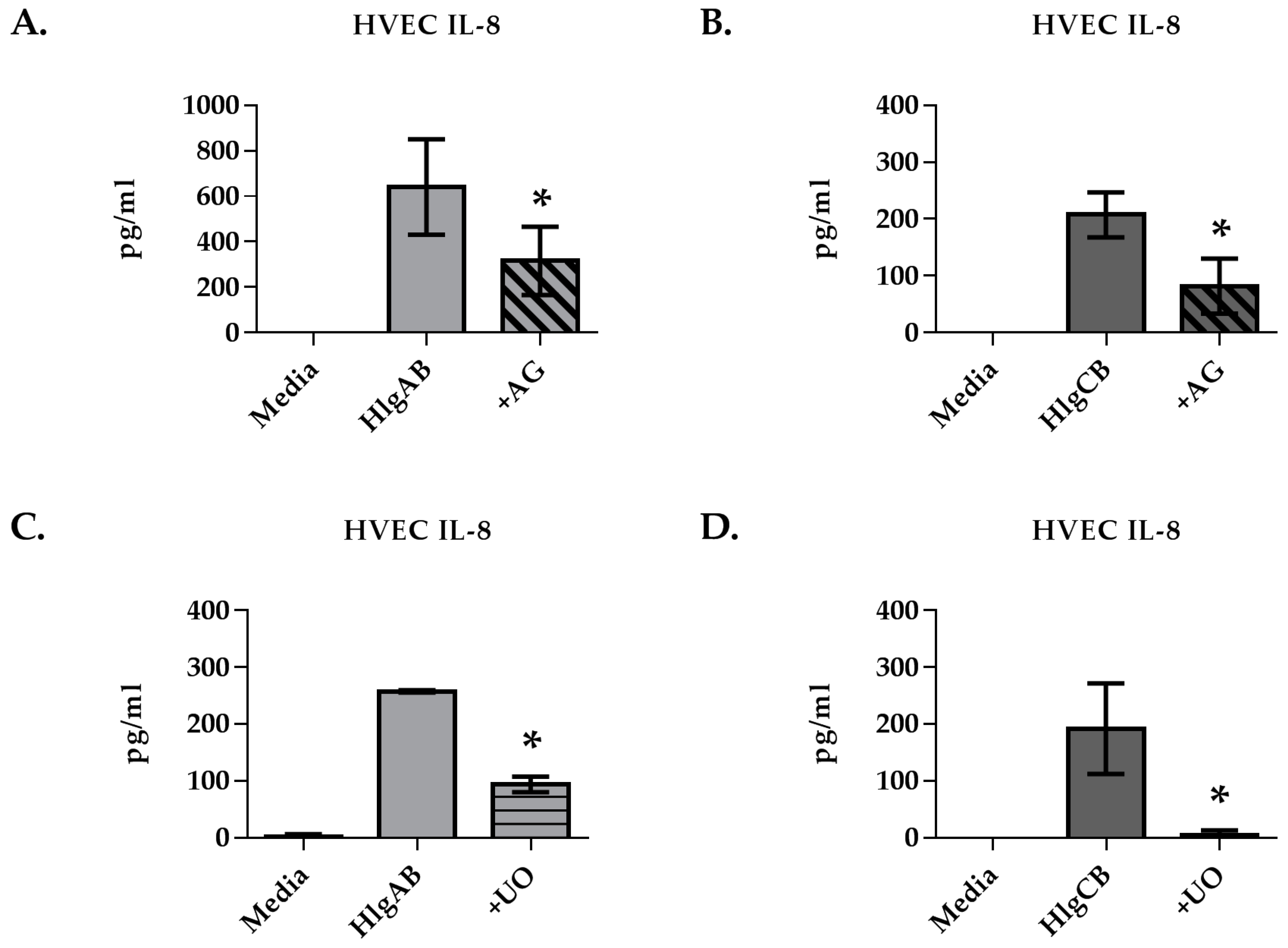
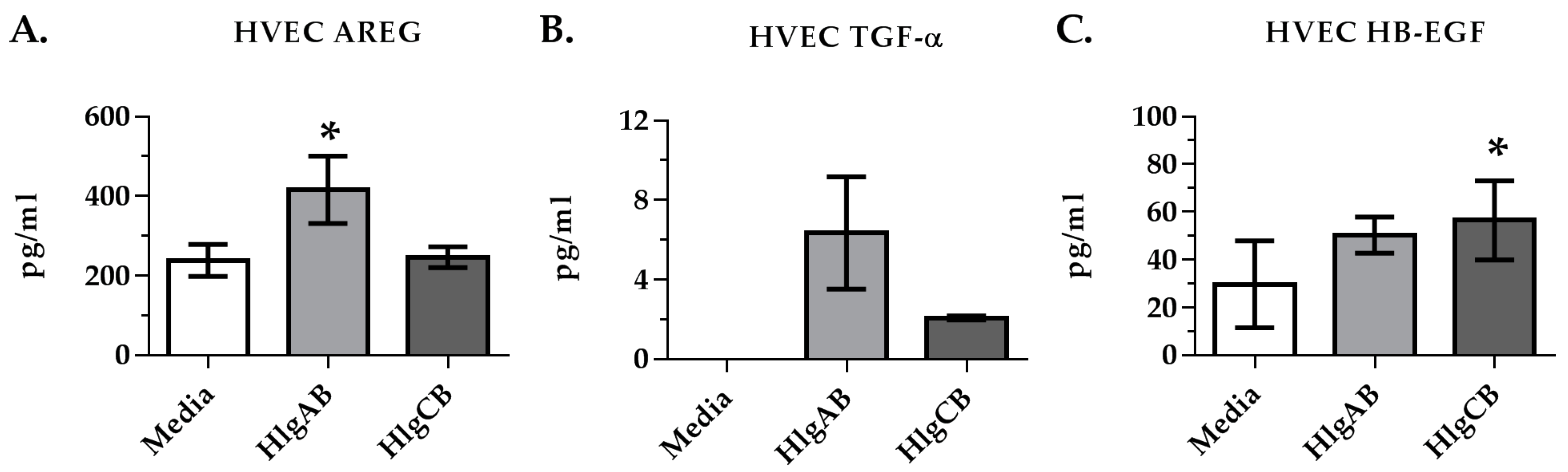
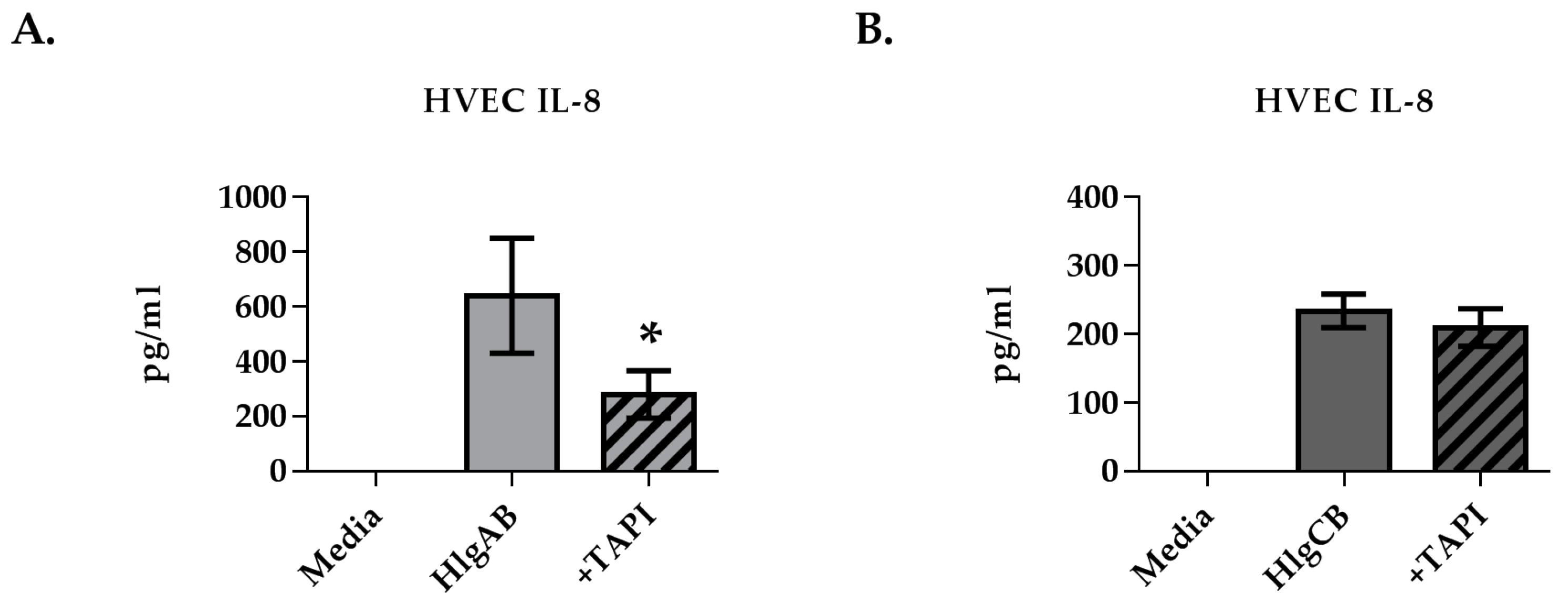
2.2. Gamma-Toxins Stimulation of IL-8 from HVECs Is Enhanced by EGFR Signaling and Involves Shedding of EGFR Ligands
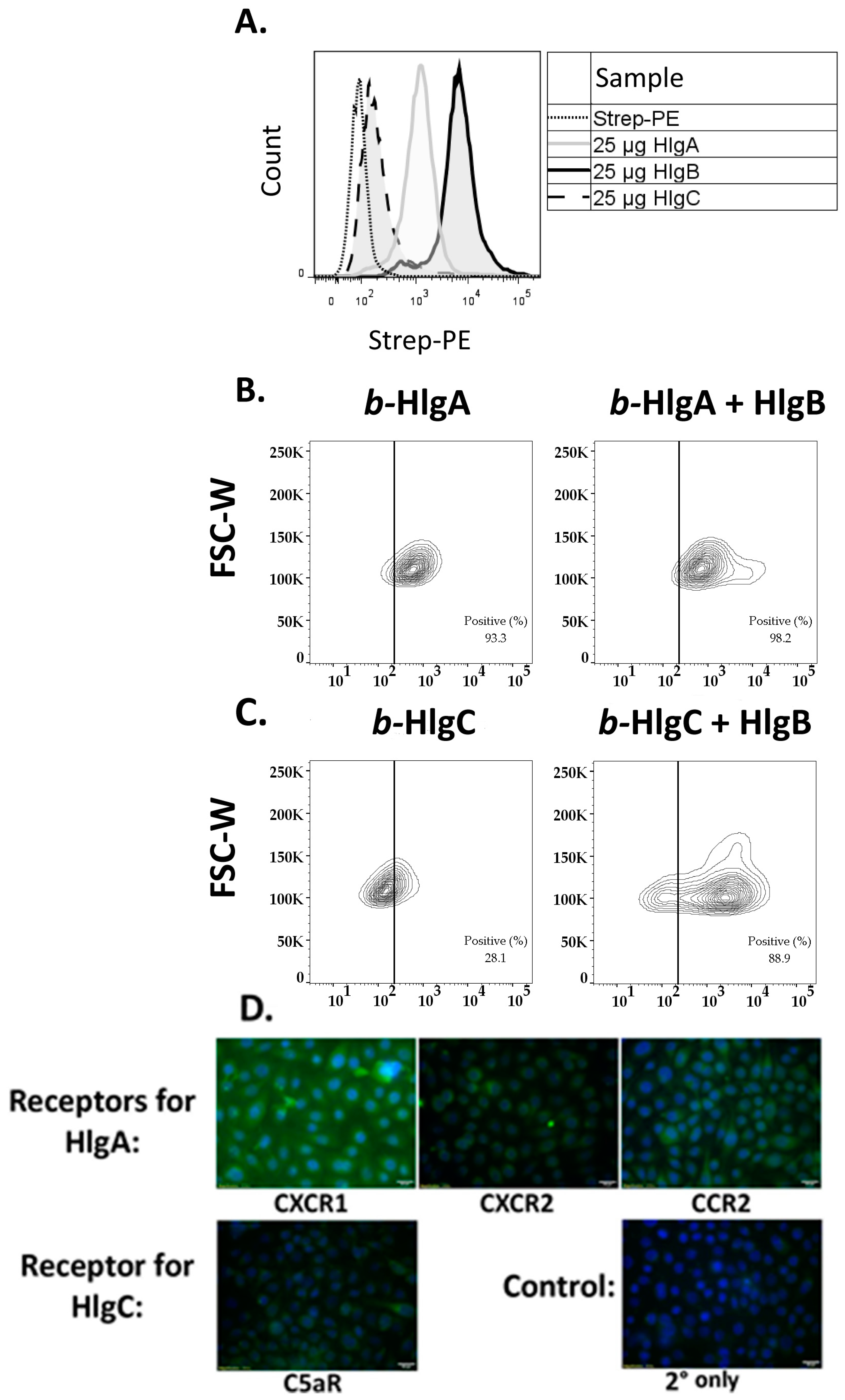
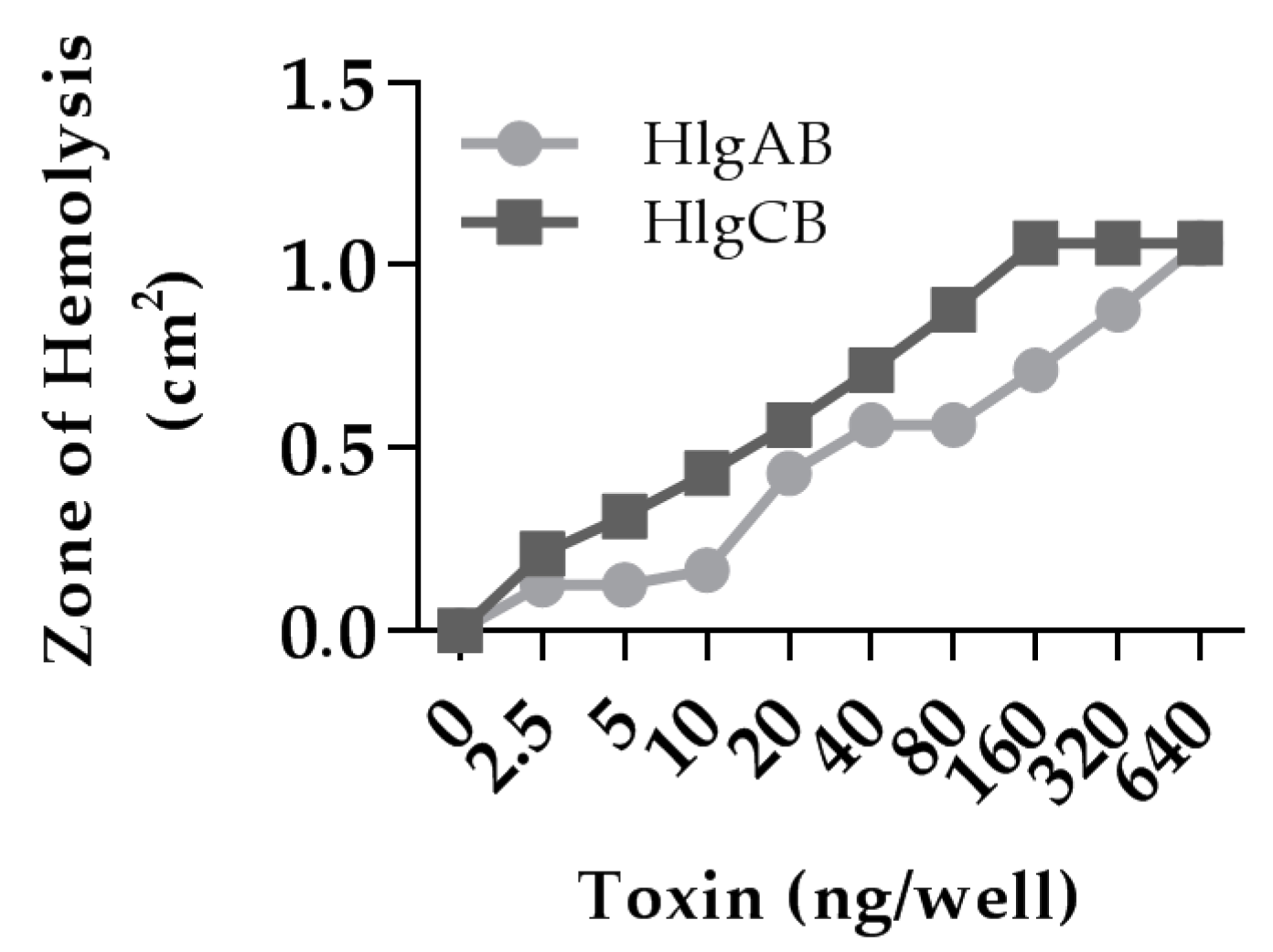
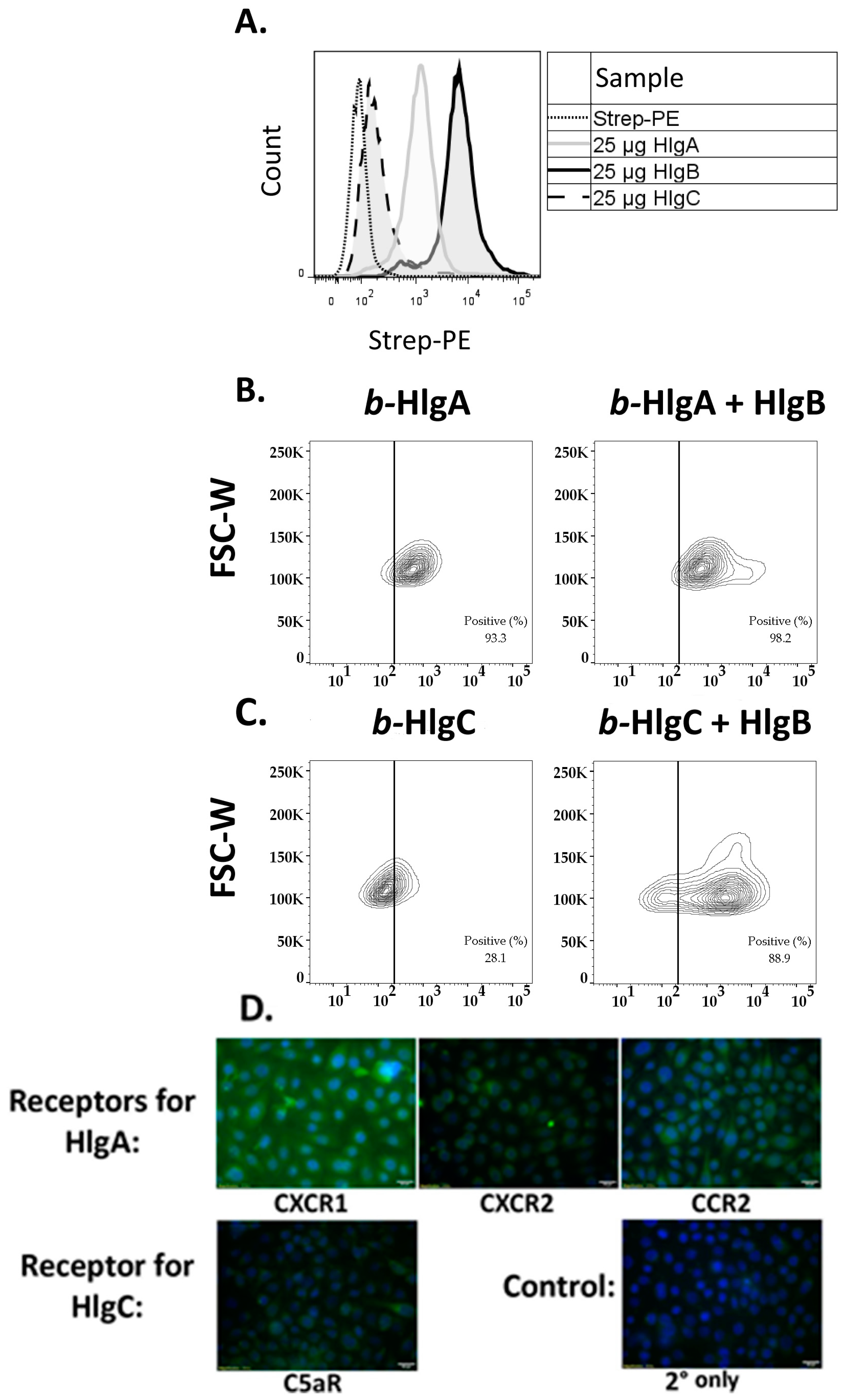
2.3. HlgB Facilitates Binding of HlgA and HlgC to HVECs and Confirmation of the Presence of the Gamma-Toxin Receptors
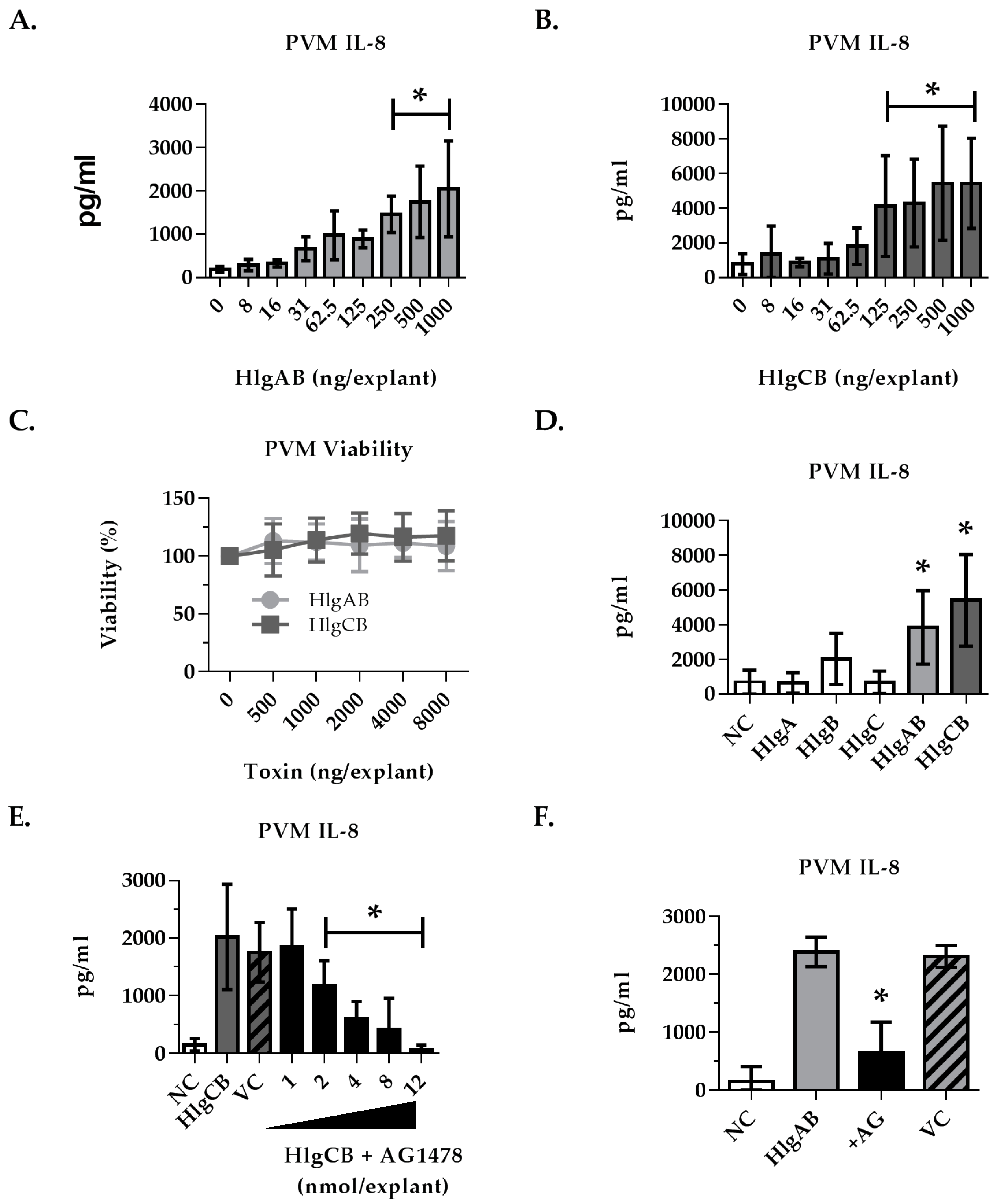
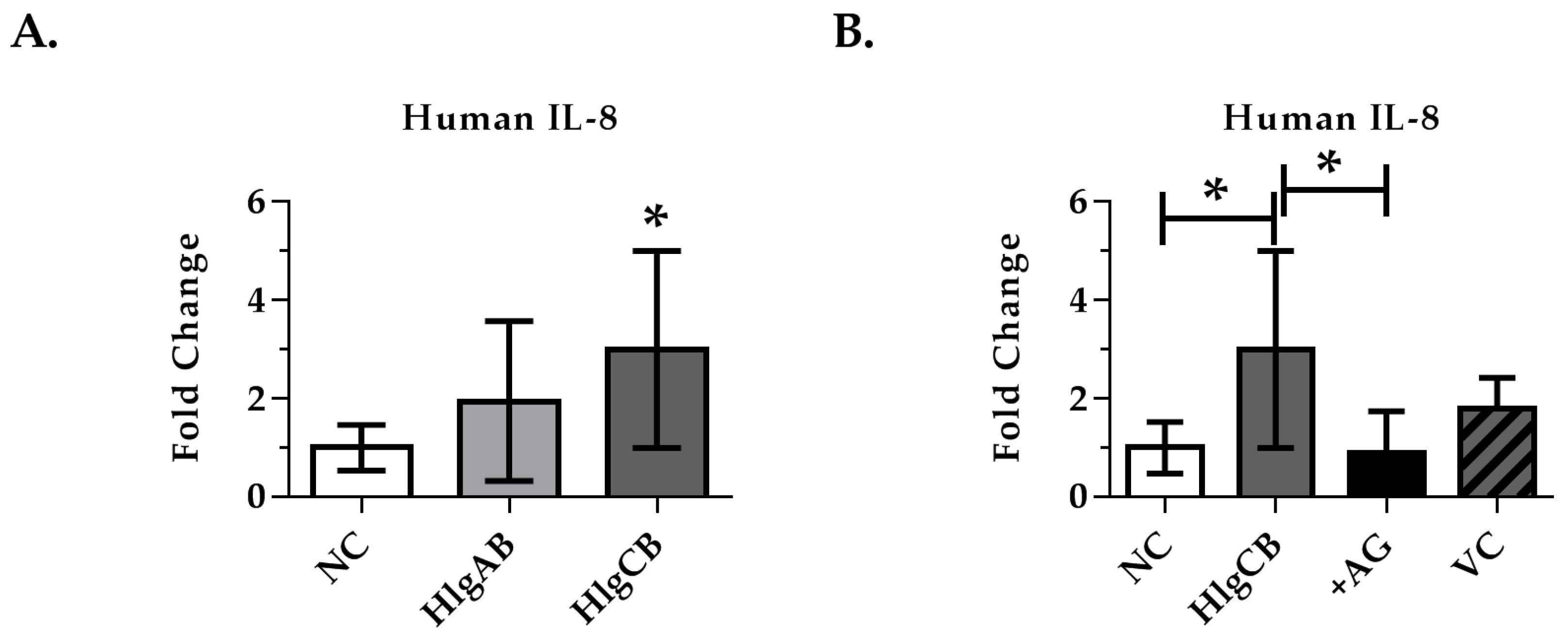
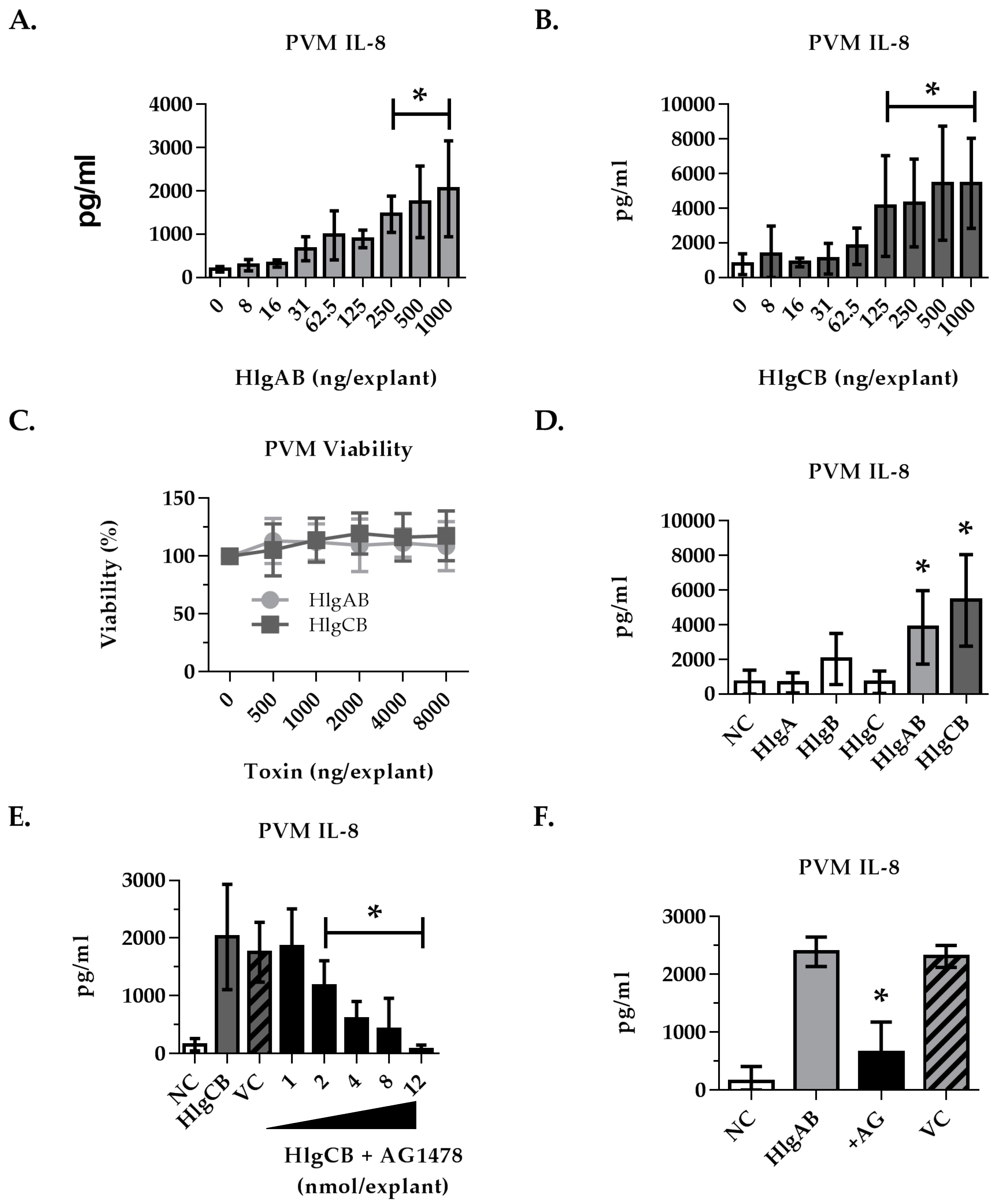
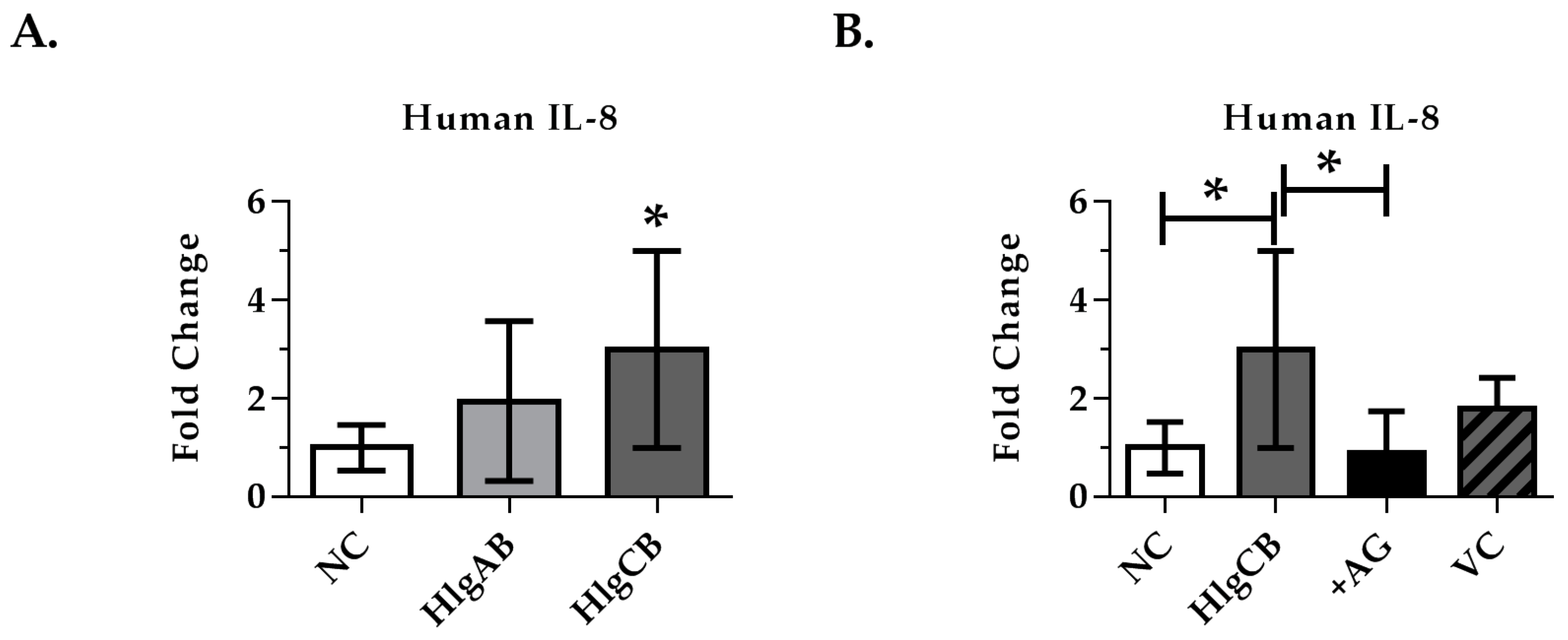
2.4. Gamma-Toxins-Induced IL-8 Production in Ex Vivo Tissue is Enhanced through EGFR Signaling
3. Discussion
4. Materials and Methods
4.1. Gamma-Toxin Purification
4.2. Cell Culture
4.3. Porcine Tissue Procurement and Assays
4.4. Human Tissue
4.5. Statistical Analysis
4.6. Ethics Statement
Supplementary Materials
Author Contributions
Conflicts of Interest
Abbreviations
| Hlg | Gamma-toxin |
| b-Hlg | Biotinylated Gamma-toxin |
| NC | Negative Control |
| VC | Vehicle Control |
| HVEC | Human Vaginal Epithelial Cells |
| PVM | Porcine Vaginal Mucosa |
| TSS | Toxic Shock Syndrome |
| mTSS | Menstrual Toxic Shock Syndrome |
| EGFR | Epidermal Growth Factor Receptor |
| ADAM | A Disintegrin and Metalloprotease |
| PBS | Phosphate Buffered Saline |
| TBS | Tris-Buffered Saline |
| DAPI | 4’,6-Diamidino-2-Phenylindole |
| IL-8 | Interleukin-8 |
| IL-6 | Interleukin-6 |
| TNF-α | Tumor Necrosis Factor Alpha |
| MIP-3 α | Macrophage Inflammatory Protein-3 |
| AREG | Amphiregulin |
| TGF- α | Tumor Growth Factor Alpha |
| HB-EGF | Heparin-Binding Epidermal Growth Factor |
| Strep-PE | Streptavidin r-Phycoerythrin |
| DMSO | Dimethyl Sulfoxide |
| FSC-W | Forward Scatter Width |
| IPTG | Isopropyl β-D-1-thiogalactopyranoside |
| KSFM | Keratinocyte Serum-Free Medium |
| RPMI | Roswell Park Memorial Institute Medium |
References
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- DeVries, A.S.; Lesher, L.; Schlievert, P.M.; Rogers, T.; Villaume, L.G.; Danila, R.; Lynfield, R. Staphylococcal Toxic Shock Syndrome 2000–2006: Epidemiology, Clinical Features, and Molecular Characteristics. PLoS ONE 2011, 6, e22997. [Google Scholar] [CrossRef] [PubMed]
- Spaulding, A.R.; Salgado-Pabon, W.; Kohler, P.L.; Horswill, A.R.; Leung, D.Y.M.; Schlievert, P.M. Staphylococcal and Streptococcal Superantigen Exotoxins. Clin. Microbiol. Rev. 2013, 26, 422–447. [Google Scholar] [CrossRef] [PubMed]
- Vandenesch, F.; Lina, G.; Henry, T. Staphylococcus aureus Hemolysins, bi-component Leukocidins, and Cytolytic Peptides: A Redundant Arsenal of Membrane-Damaging Virulence Factors? Front. Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Alonzo, F.; Torres, V.J. The Bicomponent Pore-Forming Leucocidins of Staphylococcus aureus. Microbiol. Mol. Biol. Rev. 2014, 78, 199–230. [Google Scholar] [CrossRef] [PubMed]
- Malachowa, N.; Whitney, A.R.; Kobayashi, S.D.; Sturdevant, D.E.; Kennedy, A.D.; Braughton, K.R.; Shabb, D.W.; Diep, B.A.; Chambers, H.F.; Otto, M.; et al. Global Changes in Staphylococcus aureus Gene Expression in Human Blood. PLoS ONE 2011, 6, e18617. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.; Hartford, O.; Foster, T.; Tarkowski, A. Alpha-Toxin and Gamma-Toxin Jointly Promote Staphylococcus aureus Virulence in Murine Septic Arthritis. Infect. Immun. 1999, 67, 1045–1049. [Google Scholar] [PubMed]
- Prevost, G.; Cribier, B.; Couppie, P.; Petiau, P.; Supersac, G.; Finck-Barbancon, V.; Monteil, H.; Piemont, Y. Panton-Valentine leucocidin and gamma-hemolysin from Staphylococcus aureus ATCC 49775 are encoded by distinct genetic loci and have different biological activities. Infect. Immun. 1995, 63, 4121–4129. [Google Scholar] [PubMed]
- Perret, M.; Badiou, C.; Lina, G.; Burbaud, S.; Benito, Y.; Bes, M.; Cottin, V.; Couzon, F.; Juruj, C.; Dauwalder, O.; et al. Cross-talk between Staphylococcus aureus leukocidins-intoxicated macrophages and lung epithelial cells triggers chemokine secretion in an inflammasome-dependent manner. Cell. Microbiol. 2012, 14, 1019–1036. [Google Scholar] [CrossRef] [PubMed]
- Schaefers, M.M.; Breshears, L.M.; Anderson, M.J.; Lin, Y.-C.; Grill, A.E.; Panyam, J.; Southern, P.J.; Schlievert, P.M.; Peterson, M.L. Epithelial Proinflammatory Response and Curcumin-Mediated Protection from Staphylococcal Toxic Shock Syndrome Toxin-1. PLoS ONE 2012, 7, e32813. [Google Scholar] [CrossRef] [PubMed]
- Brosnahan, A.J.; Mantz, M.J.; Squier, C.A.; Peterson, M.L.; Schlievert, P.M. Cytolysins Augment Superantigen Penetration of Stratified Mucosa. J. Immunol. 2009, 182, 2364–2373. [Google Scholar] [CrossRef] [PubMed]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Breshears, L.M.; Schlievert, P.M.; Peterson, M.L. A Disintegrin and Metalloproteinase 17 (ADAM17) and Epidermal Growth Factor Receptor (EGFR) Signaling Drive the Epithelial Response to Staphylococcus aureus Toxic Shock Syndrome Toxin-1 (TSST-1). J. Biol. Chem. 2012, 287, 32578–32587. [Google Scholar] [CrossRef] [PubMed]
- Büchau, A.S. EGFR (Trans)activation Mediates IL-8 and Distinct Human Antimicrobial Peptide and Protein Production following Skin Injury. J. Investig. Dermatol. 2010, 130, 929–932. [Google Scholar] [CrossRef] [PubMed]
- Clyne, M.; de Azavedo, J.; Carlson, E.; Arbuthnott, J. Production of gamma-hemolysin and lack of production of alpha-hemolysin by Staphylococcus aureus strains associated with toxic shock syndrome. J. Clin. Microbiol. 1988, 26, 535–539. [Google Scholar] [PubMed]
- Alessandrini, A.; Viero, G.; Dalla Serra, M.; Prévost, G.; Facci, P. γ-Hemolysin Oligomeric Structure and Effect of its Formation on Supported Lipid Bilayers: An AFM Investigation. Biochim. Biophys. Acta BBA-Biomembr. 2013, 1828, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Von Eiff, C.; Friedrich, A.W.; Peters, G.; Becker, K. Prevalence of genes encoding for members of the staphylococcal leukotoxin family among clinical isolates of Staphylococcus aureus. Diagn. Microbiol. Infect. Dis. 2004, 49, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Spaan, A.N.; Vrieling, M.; Wallet, P.; Badiou, C.; Reyes-Robles, T.; Ohneck, E.A.; Benito, Y.; de Haas, C.J.C.; Day, C.J.; Jennings, M.P.; et al. The staphylococcal toxins γ-haemolysin AB and CB differentially target phagocytes by employing specific chemokine receptors. Nat. Commun. 2014, 5, 5438. [Google Scholar] [CrossRef] [PubMed]
- Haugwitz, U.; Bobkiewicz, W.; Han, S.-R.; Beckmann, E.; Veerachato, G.; Shaid, S.; Biehl, S.; Dersch, K.; Bhakdi, S.; Husmann, M. Pore-forming Staphylococcus aureus α-toxin triggers epidermal growth factor receptor-dependent proliferation. Cell. Microbiol. 2006, 8, 1591–1600. [Google Scholar] [CrossRef] [PubMed]
- Geilen, C.; Wieprecht, M.; Orfanos, C. The mitogen-activated protein kinases system (MAP kinase cascade): Its role in skin signal transduction. A review. J. Dermatol. Sci. 1996, 12, 255–262. [Google Scholar] [CrossRef]
- Biswas, D.K.; Iglehart, J.D. Linkage between EGFR family receptors and nuclear factor kappaB (NF-κB) signaling in breast cancer. J. Cell. Physiol. 2006, 209, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Seals, D.F.; Courtneidge, S.A. The ADAMs family of metalloproteases: Multidomain proteins with multiple functions. Genes Dev. 2003, 17, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, T.; Kaneko, J.; Kamio, Y. Essential binding of LukF of staphylococcal gamma-hemolysin followed by the binding of H gamma II for the hemolysis of human erythrocytes. Biosci. Biotechnol. Biochem. 1995, 59, 1181–1183. [Google Scholar] [CrossRef] [PubMed]
- Kuehnert, M.J.; Kruszon-Moran, D.; Hill, H.A.; McQuillan, G.; McAllister, S.K.; Fosheim, G.; McDougal, L.K.; Chaitram, J.; Jensen, B.; Fridkin, S.K. Prevalence of Staphylococcus aureus nasal colonization in the United States, 2001–2002. J. Infect. Dis. 2006, 193, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Peterson, M.L.; Ault, K.; Kremer, M.J.; Klingelhutz, A.J.; Davis, C.C.; Squier, C.A.; Schlievert, P.M. The Innate Immune System Is Activated by Stimulation of Vaginal Epithelial Cells with Staphylococcus aureus and Toxic Shock Syndrome Toxin 1. Infect. Immun. 2005, 73, 2164–2174. [Google Scholar] [CrossRef] [PubMed]
- Rigby, K.M.; DeLeo, F.R. Neutrophils in innate host defense against Staphylococcus aureus infections. Semin. Immunopathol. 2012, 34, 237–259. [Google Scholar] [CrossRef] [PubMed]
- Palazzolo-Ballance, A.M.; Reniere, M.L.; Braughton, K.R.; Sturdevant, D.E.; Otto, M.; Kreiswirth, B.N.; Skaar, E.P.; DeLeo, F.R. Neutrophil microbicides induce a pathogen survival response in community-associated methicillin-resistant Staphylococcus aureus. J. Immunol. 2008, 180, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Harding, M.G.; Zhang, K.; Conly, J.; Kubes, P. Neutrophil Crawling in Capillaries; A Novel Immune Response to Staphylococcus aureus. PLoS Pathog. 2014, 10, e1004379. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Thammavongsa, V.; Schneewind, O.; Missiakas, D. Recurrent infections and immune evasion strategies of Staphylococcus aureus. Curr. Opin. Microbiol. 2012, 15, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Ganem, M.B.; de Marzi, M.C.; Fernández-Lynch, M.J.; Jancic, C.; Vermeulen, M.; Geffner, J.; Mariuzza, R.A.; Fernández, M.M.; Malchiodi, E.L. Uptake and Intracellular Trafficking of Superantigens in Dendritic Cells. PLoS ONE 2013, 8, e66244. [Google Scholar] [CrossRef] [PubMed]
- Alonzo, F.; Torres, V.J. Bacterial Survival Amidst an Immune Onslaught: The Contribution of the Staphylococcus aureus Leukotoxins. PLoS Pathog. 2013, 9, e1003143. [Google Scholar] [CrossRef] [PubMed]
- Yoong, P.; Torres, V.J. The effects of Staphylococcus aureus leukotoxins on the host: Cell lysis and beyond. Curr. Opin. Microbiol. 2013, 16, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Inoshima, I.; Inoshima, N.; Wilke, G.A.; Powers, M.E.; Frank, K.M.; Wang, Y.; Wardenburg, J.B. A Staphylococcus aureus pore-forming toxin subverts the activity of ADAM10 to cause lethal infection in mice. Nat. Med. 2011, 17, 1310–1314. [Google Scholar] [CrossRef] [PubMed]
- Huovila, A.-P.J.; Turner, A.J.; Pelto-Huikko, M.; Kärkkäinen, I.; Ortiz, R.M. Shedding light on ADAM metalloproteinases. Trends Biochem. Sci. 2005, 30, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Oda, K.; Matsuoka, Y.; Funahashi, A.; Kitano, H. A comprehensive pathway map of epidermal growth factor receptor signaling. Mol. Syst. Biol. 2005, 1, E1–E17. [Google Scholar] [CrossRef] [PubMed]
- Ellis, A.G.; Doherty, M.M.; Walker, F.; Weinstock, J.; Nerrie, M.; Vitali, A.; Murphy, R.; Johns, T.G.; Scott, A.M.; Levitzki, A.; et al. Preclinical analysis of the analinoquinazoline AG1478, a specific small molecule inhibitor of EGF receptor tyrosine kinase. Biochem. Pharmacol. 2006, 71, 1422–1434. [Google Scholar] [CrossRef] [PubMed]
- DuMont, A.L.; Yoong, P.; Liu, X.; Day, C.J.; Chumbler, N.M.; James, D.B.A.; Alonzo, F.; Bode, N.J.; Lacy, D.B.; Jennings, M.P.; et al. Identification of a Crucial Residue Required for Staphylococcus aureus LukAB Cytotoxicity and Receptor Recognition. Infect. Immun. 2014, 82, 1268–1276. [Google Scholar] [CrossRef] [PubMed]
- Fichorova, R.N.; Rheinwald, J.G.; Anderson, D.J. Generation of papillomavirus-immortalized cell lines from normal human ectocervical, endocervical, and vaginal epithelium that maintain expression of tissue-specific differentiation proteins. Biol. Reprod. 1997, 57, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J.; Parks, P.J.; Peterson, M.L. A mucosal model to study microbial biofilm development and anti-biofilm therapeutics. J. Microbiol. Methods 2013, 92, 201–208. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gillman, A.N.; Breshears, L.M.; Kistler, C.K.; Finnegan, P.M.; Torres, V.J.; Schlievert, P.M.; Peterson, M.L. Epidermal Growth Factor Receptor Signaling Enhances the Proinflammatory Effects of Staphylococcus aureus Gamma-Toxin on the Mucosa. Toxins 2017, 9, 202. https://doi.org/10.3390/toxins9070202
Gillman AN, Breshears LM, Kistler CK, Finnegan PM, Torres VJ, Schlievert PM, Peterson ML. Epidermal Growth Factor Receptor Signaling Enhances the Proinflammatory Effects of Staphylococcus aureus Gamma-Toxin on the Mucosa. Toxins. 2017; 9(7):202. https://doi.org/10.3390/toxins9070202
Chicago/Turabian StyleGillman, Aaron N., Laura M. Breshears, Charles K. Kistler, Patrick M. Finnegan, Victor J. Torres, Patrick M. Schlievert, and Marnie L. Peterson. 2017. "Epidermal Growth Factor Receptor Signaling Enhances the Proinflammatory Effects of Staphylococcus aureus Gamma-Toxin on the Mucosa" Toxins 9, no. 7: 202. https://doi.org/10.3390/toxins9070202