Abstract
Malignant gliomas are heterogeneous neoplasms. Glioma stem-like cells (GSCs) are undifferentiated and self-renewing cells that develop and maintain these tumors. These cells are the main population that resist current therapies. Genomic and epigenomic analyses has identified various molecular subtypes. Bone morphogenetic protein 4 (BMP4) reduces the number of GSCs through differentiation and induction of apoptosis, thus increasing therapeutic sensitivity. However, the short half-life of BMP4 impedes its clinical application. We previously reviewed BMP4 signaling in central nervous system development and glioma tumorigenesis and its potential as a treatment target in human gliomas. Recent advances in understanding both adult and pediatric malignant gliomas highlight critical roles of BMP4 signaling pathways in the regulation of tumor biology, and indicates its potential as a therapeutic molecule. Furthermore, significant progress has been made on synthesizing BMP4 biocompatible delivery materials, which can bind to and markedly extend BMP4 half-life. Here, we review current research associated with BMP4 in brain tumors, with an emphasis on pediatric malignant gliomas. We also summarize BMP4 delivery strategies, highlighting biocompatible BMP4 binding peptide amphiphile nanostructures as promising novel delivery platforms for treatment of these devastating tumors.
1. Introduction
Malignant gliomas are the most aggressive category of primary brain tumor [1]. Despite decades of research, curing these tumors remains a challenge [2]. The incidence of malignant gliomas differs by age. In adults (≥19 years), the average overall annual incidence is 8.82 per 100,000. In children (<19 years), malignant gliomas include anaplastic astrocytoma, glioblastoma and diffuse intrinsic pontine gliomas (DIPGs), with an average annual incidence of 3.48 per 100,000 [1]. Regardless of age, patients with these devastating tumors have a poor clinical prognosis [3,4]. Radical surgical resection followed by adjuvant radiotherapy and/or chemotherapy are standard treatments for these tumors, however, tumor recurrence occurs in nearly all instances, primarily due to intrinsic or acquired resistance to routinely used therapies [5]. Identifying novel therapeutic approaches to improve survival in patients with these malignancies is imperative.
Data from the Central Brain Tumor Registry of the United States (CBTRUS) reveals differences between adult and pediatric patients including tumor incidence and location [1]. Genomic and epigenomic analyses have also shown significant differences between adult and pediatric tumors [6,7]. In adult high-grade gliomas (aHGGs), epidermal growth factor receptor (EGFR) is a commonly altered receptor tyrosine kinase (RTK) and mosaic expression of platelet-derived growth factor receptor-α (PDGFRA), platelet-derived growth factor α (PDGFA), fibroblast growth factor receptor 1(FGFR1), fibroblast growth factor 1 (FGF1), NOTCH2, JAG1 (Jagged Canonical Notch Ligand 1) are common. Additionally, IDH1 mutations have been identified in glioblastomas developed from WHO grade II/III astrocytomas or oligodendrogliomas [8,9,10,11]. In pediatric high-grade gliomas (pHGGs), PDGFRA is a more common RTK alteration and MYC and MYCN are frequently amplified [12]. Furthermore, multiple hotspot histone mutations have been identified in pHGGs, but are rare in aHGG. These histone mutations vary further between different pHGG tumor types. For instance, mutations in H3, family 3A (H3F3A) and histone cluster 1, H3b (HIST1H3B), occur at lysine 27 (K27M) in ~80% of DIPGs [13,14], a subset of pHGGs arising from the brainstem. Mutations on histone H3G34 (G34V/R) are present in ~38% of hemispheric pHGGs [12]. In addition to histone mutations, TP53 and activin receptor type 1 (ACVR1, also known as ALK2) mutations are frequent in DIPG [15,16,17], and chimeric fusions involving the kinase domain of neurotrophic tyrosine kinase receptors are present in ~40% of hemispheric pHGG [13].
Regardless of the aforementioned molecular differences between aHGGs and pHGGs, a small population of glioma stem-like cells (GSCs) are considered a driving force for tumor growth and recurrence, and tumor heterogeneity [18,19,20,21]. GSCs can initiate tumors that reproduce the parental tumors’ cellular heterogeneity. GSCs also resist the cytotoxic effects of radiation and chemotherapy [22,23,24,25,26]. These findings indicate that GSCs may be critical therapeutic targets.
Bone morphogenetic protein 4 (BMP4) can abolish cancer stem cell populations in human cancers [27,28,29,30,31,32], including in malignant gliomas [33,34,35,36,37]. In a current phase I clinical trial (NCT02869243) human recombinant BMP4 is being administered through intratumoral and interstitial convection-enhanced delivery (CED) for adult glioblastoma treatment (https://clinicaltrials.gov/ct2/show/NCT02869243). BMP4 signal pathways appear to play critical roles in the regulation of malignant glioma tumor biology, further suggesting that it is a promising therapeutic molecule. However, to fully elucidate BMP4 therapeutic potential, differential roles of BMP4 in tumor molecular subgroups should be examined. In addition, to take advantage of this potential, novel biocompatible materials for effective BMP4 binding and delivery are being synthesized. Preliminary unpublished results from our laboratory showed that an innovative biocompatible peptide amphiphile nanostructure binds BMP4 and markedly extends its half-life, an important factor for its clinical utility [38]. In this review, we have discussed the recent discoveries elucidating the role of BMP4 signal pathways in malignant gliomas and reviewed innovative biocompatible materials for BMP4 delivery and their prospects for clinical applications.
2. BMP4 Signal Pathways and Glioma Biology
BMP4 is a member of the TGF-β family. BMP4 signal pathways are critical in early embryonic development, central nervous system (CNS) formation and development through regulation of stemness and differentiation of neural stem cells (NSCs) [39,40]. In the subventricular zone of the adult brain, BMP4 promotes NSC differentiation into astrocytes [41]. BMP4 directly binds to BMPR1A, BMPR1B, and BMPRII, resulting in phosphorylation of cytosolic Smad (mothers against decapentaplegic homolog) proteins. Smad proteins translocate to the cell nucleus, where they bring about Smad-mediated gene expression as well as activation of MAPK (mitogen-activated protein kinase) signaling as described [42]. Increasing evidence indicates that BMP4 signaling pathways are relevant to human gliomas. However, the role of BMP4 signaling pathways varies between aHGGs and pHGGs, due to differences in the molecular background. For example, a mutation in ACVR1, a member of the BMPRI family, is more frequent in pHGGs, compared to aHGGs [12,15,16,17,37,43,44]. In light of the molecular background differences, BMP4 action needs to be interpreted with respect to the distinct features of each tumor group.
3. BMP4 Signaling in Adult High-Grade Gliomas
HGGs are the most common solid CNS adult tumors. We analyzed the Data from The Cancer Genome Analysis (TCGA) using the GlioVis data portal for visualization and analysis of brain tumor expression datasets [45] (http://gliovis.bioinfo.cnio.es/). The results showed that low-grade gliomas (LGG) express higher BMP4 levels and exhibit lower mortality rates than HGGs that express lower levels of BMP4 (Figure 1). These results were consistent over multiple data sources [46,47], and suggest that BMP4 can be a robust prognostic marker for adult gliomas. The results further suggest that therapeutic targeting of BMP4 may be an effective strategy for treating aHGG.
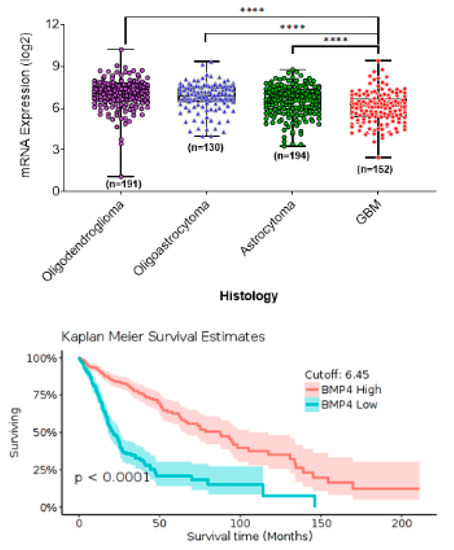
Figure 1.
In silico data analysis from The Cancer Genome Analysis (TCGA) using the Gliovis online portal shows BMP4 mRNA expression associated with tumor category based on histology (top panel) and patient survival in adult gliomas (bottom panel).
GSCs are considered a source for tumors and these cells are resistant to radiation and chemotherapy [48,49]. One strategy to improve treatment outcomes for aHGG is to target GSCs to improve tumor response to conventional therapies. Another strategy is to induce GSC differentiation, resulting in a reduction of the tumorigenic cell population [50]. Treatment with BMPs, including BMP4 provides an approach for inducing GSC differentiation. GSCs express BMP receptors, and have a functional BMP4 signal pathway. The addition of exogenous BMP4 to GSCs enhances SMAD phosphorylation and reduces GSC proliferation [41,51]. Furthermore, in response to BMP4, CD133, a GSC marker, decreases, whereas GFAP, a marker for differentiated astrocytes, increases. Treatment with exogenous BMP4 also decreases GSC tumorigenicity in vivo [51] and reduces tumor cell proliferation [52]. These results, in total, suggest that BMP4 promotes GSC differentiation, and may prove useful in treating HGGs [53]. BMP4 also reduces multidrug resistance in glioma cells and suppresses glioblastoma invasiveness. Multidrug resistance is reduced through the inhibition of B-cell lymphoma 2 (BCL-2) and glial cell derived neurotrophic factor (GDNF), while invasiveness is reduced through increased E-cadherin and claudin expression [33].
With advances in biotechnology, including integrative application of high-throughput sequencing such as single cell RNA-seq (scRNA-seq), 450K DNA methylation profiling, high-throughput m6A-seq, and whole-genomic sequencing (WGS), it is possible to obtain precise molecular signatures, and identify the diverse genetic and epigenetic programs that drive cancers such as gliomas. For example, scRNA-seq reveals proneural, classic and mesenchymal GSC subtypes within individual tumors, thus demonstrating intratumoral cellular heterogeneity [10,54,55]. Preliminary results from our laboratory have indicated that these cell subtypes respond differently to BMP4 (Figure 2A). Proliferation of mesenchymal subtype GSCs does not decrease following treatment with 100ng/ml BMP4 for 4 days. In comparison, proliferation of proneural subtype GCSs does decrease under these conditions. This may reflect different levels of endogenous BMP4 expression (Figure 2B). For instance, the mesenchymal glioblastoma subtype expresses higher levels of BMP4 than proneural and classic GSC subtypes. The pre-glioblastoma subtype within isocitrate dehydrogenase 1 (IDH1) mutant gliomas express low BMP4 (Figure 2C) [56], in comparison to early progenitor-like and neuroblastic subtypes, and is associated with a poor patient prognosis (Figure 2D) [54,57].
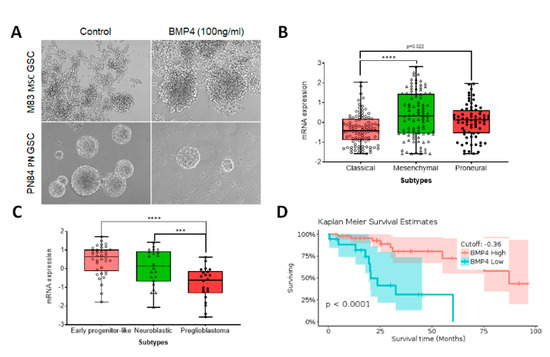
Figure 2.
BMP4 expression in adult glioblastoma subtypes, associated with patient outcome. (A) Representative images show that mesenchymal stem cell (MSC) subtype M83 glioma stem-like cells (GSCs) and proneural (PN) PN84 GSCs respond differentially to BMP4 treatment for 96 h, with only PN84 GSCs showing decreased proliferation. (B–D) In silico TCGA data analysis with Gliovis indicates that: BMP4 levels vary in adult glioblastoma subtypes (B); BMP4 expression varies in isocitrate dehydrogenase 1 (IDH1) mutant early progenitor-like, neuroblastic and preglioblastoma cells (C) and BMP4 level is associated with survival in patients with IDH1 mutant glioblastoma (D).
4. BMP4 Signaling in Pediatric High-Grade Gliomas
Pediatric brain tumors are distinct from their adult counterparts in terms of epidemiology, cellular origins, response to cytotoxic and radiation therapy and clinical outcomes. Recent wide-spread genome-wide profiling that has been applied to pediatric brain tumors has provided full characterization at the molecular genetic level. These large-scale analyses have revealed distinct tumor driving events, gene expression profiles, mutation targets and mutation frequencies [7]. Accordingly, BMP4 involvement in pHGG biology warrants its own examination.
In silico analysis of data from dataset GEO: GSE73038 ([58] shows that BMP4 is differentially expressed among histopathologically-defined pediatric CNS brain tumors (Figure 3, left panel), including in pHGGs (Figure 3, right panel). DIPGs, pHGGs arising in the brainstem, are characterized by an H3K27M mutation in either histone H3.1 or H3.3. H3.3 K27M mutations are also present in other pHGGs from midline regions, including from areas such as the thalamus, cerebellum and spine [59]. While H3.1 K27M mutations are restricted to DIPG [60], they usually occur in conjunction with abnormal signaling pathway activity including pathways associated with BMP4 [14,15,16,29,61]. Recurrent somatic mutations involving ACVR1 have also been discovered in DIPGs [13,15,16]. Interestingly, gain-of-function mutations in ACVR1 appear to be restricted to DIPGs with an H3.1 K27M of the HIST1H3B gene, and are not present in DIPGs with H3.1 K27M mutation of the H3F3A gene. DIPGs harboring ACVR1 mutations exhibit hyperactivation of BMP-ACVR1 signaling, which results in elevation of phosphorylated SMAD1/5/9 and increased expression of BMP downstream response genes [16]. DIPG patients whose tumor harbors an ACVR1 mutation show improved survival [13,17]. Therapeutic targeting of AVCR1 has beneficial anti-tumor effects in preclinical DIPG models [43]. However, the targeting effect is mutation domain dependent [43,44].
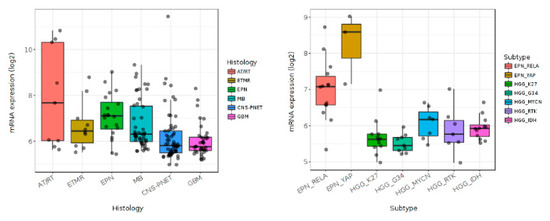
Figure 3.
In silico analysis of pediatric brain tumor datasets (GEO: GSE73038) with Gliovis showing differential expression of BMP4 in pediatric central nervous system tumors (left panel) and pediatric high grade gliomas (right panel).
pHGGs also display other histone mutations, for instance the histone H3G34 (G34V/R) mutation is present in hemispheric pHGGs [12]. Preliminary unpublished results from our laboratory showed increased phosphorylated SMAD1/5/9 and decreased multidrug resistance gene 1 (MDR1) expression, in pediatric glioblastoma KNS42 cells harboring an H3F3A G34V mutation, following BMP4 treatment. Decreasing MDR1 increases tumor cell sensitivity to cytotoxic therapies. Thus, the result with KNS42 cells indicates that, in addition to ACVR1 mutated H3.1 K27M DIPGs, other pHGGs may benefit from BMP4 targeted therapy.
5. BMP4 Delivery Methods for Glioma Treatment
The BMP signaling pathway is a potential therapeutic target for treating gliomas. Therapeutic applications of BMP4 for both adult and pediatric HGGs are based on its ability to induce differentiation and apoptosis of GSCs and thus reduce this cell population. The mechanism for BMP differentiation therapy involves driving GSCs into a post-mitotic state that limits tumor growth. However, there are obstacles that must be overcome relative to BMP4 clinical treatment of malignant gliomas via differentiation therapy [62,63]. For instance, autocrine BMP4 enhances tumor aggressiveness in IDH1 mutant gliomas [64]. It is possible that only certain subsets of GSCs, based on molecular characteristics, are targetable in response to high doses of BMP4. Some cell subsets may show incomplete cell-cycle arrest and/or tumor cell retention of growth-promoting DNA methylation patterns [63]. Further investigation of cell molecular characteristics and differentiation needs to be done to help overcome these obstacles. Another limiting factor for clinical application of BMP4 is its short half-life [65,66,67]. One strategy for overcoming this limitation is the delivery of large doses of BMP4 via polymer beads [51]. Other delivery systems have been designed to overcome the short half-life of BMP4 and improve its biomedical effects, including recent advances in the synthesis of biocompatible BMP4 binding materials. The following discussion reviews delivery systems (Figure 4) and our innovative peptide amphiphile nanostructures as an innovative BMP4 delivery platform (Figure 5).
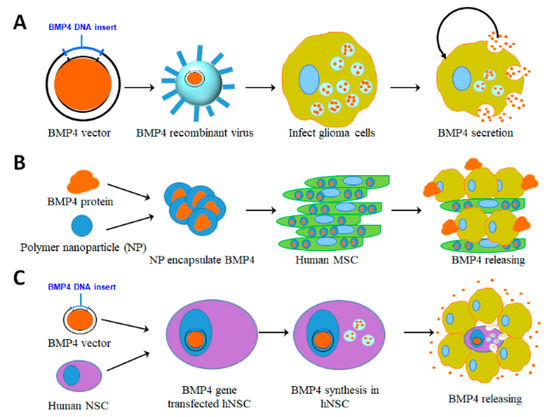
Figure 4.
Representative illustrations indicate BMP4 delivery methods. (A) Viral vector based delivery. BMP4 plasmids are transduced into oncolytic virus to infect glioblastoma cells, which consequently induces apoptosis and differentiation to improve the therapeutic outcome. (B) Human mesenchymal stem cell (hMSC) based delivery. BMP4 is encapsulated into polymer nanoparticles (NP) and transfected into human adipose MSCs, which sustainably release BMP4 to target glioblastoma cells. (C) BMP4 plasmids are transfected into human neural stem cells (NSCs). When these NSCs are co-cultured with glioblastoma cells or injected into glioblastomas, BMP4 is produced and released to target tumor cells.
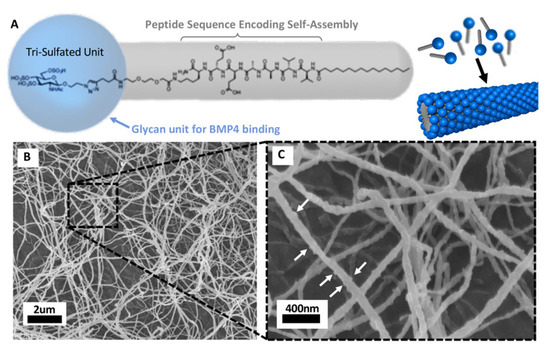
Figure 5.
Peptide amphiphile (PA) nanostructures are novel biocompatible materials for BMP4 delivery. (A) Molecular structure of a glycosylated PA molecule and schematic of the self-assembled glycosylated PA nanofiber. The grey molecular region guides the self-assembly process into nanofibers, the blue molecular region contains the tri-sulfated monosaccharide responsible for protein binding actions. (B) SEM of glycosylated PA nanofiber bundles after exposure to blood proteins for 5 min. (C) Higher magnification of nanofibers with white arrows indicating a rigid surface texture resulting from proteins binding to the fiber surface. (Figures provided by Dr. Samuel Stupp with permission for publication).
5.1. Viral Vector Based Delivery
Viral vectors have been used for high efficiency gene delivery, including for the production of BMP4 in gliomas. An oncolytic vaccinia virus (VACV) expressing BMP4 was delivered both in vitro to primary glioma cultures and in vivo intracranially to xenograft gliomas (Figure 4A). The results of the in vitro study showed cytotoxic activity against GSCs and the in vivo study improved survival rates in treated mice and reduced recurrence of glioma following VACV infection [68]. VACVs, however, are associated with risks including neurodegeneration and demyelination, which limits their clinical application for expressing BMP4 in gliomas [69].
5.2. Human Adipose-Derived Mesenchymal Stem Cell (hAMSC) Based Delivery
Because of their high glioma tropism, human adipose-derived mesenchymal stem cells (hAMSCs) have been touted as a potential therapeutic delivery vehicle for glioma treatment. Though originally derived from bone marrow, large amounts of MSCs can be isolated from adipose tissue, with cells from either source relatively equivalent in treatment efficacy [70]. Furthermore, hAMSCs can be altered with nanoparticles to be more effective than conventional polymers in delivering BMP4 [70]. Nanoparticle-engineered hAMSCs expressing BMP4 cross the blood brain barrier, migrate to and penetrate intracranial tumors, and extend survival. In vivo and in vitro studies showed that hAMSC-BMP4 decreased migration and proliferation of GSCs while promoting differentiation (Figure 4B). Additionally, mice bearing murine GBM experienced improved survival after treatment with hAMSC-BMP4. Significantly, in vivo, hAMSCs maintain their multipotency and hAMSC malignant transformation has not been observed, despite exposure to the GBM microenvironment [71]. However, with the application of tumor growth factors, hAMSCs can transform into fibroblasts and potentially contribute to tumor expansion [71,72]. Further, hAMSCs stop proliferating in vivo after a few days, so that the effect of BMP4 from hAMSC production is limited [73].
5.3. Human Neural Stem Cell (hNSC) Based Delivery
To address deficiencies in the distribution of “free” oncolytic vectors, the use of virally transduced human neural stem cells (hNSCs) has been proposed to treat gliomas. These cells would deliver conditionally replicating adenovirus (CRAd) (Figure 4C). NSCs have shown an intrinsic migratory capacity towards brain tumors, though the mechanisms of this tropism are poorly understood [70,74]. Harnessing the homing ability of hNSCs in conjugation with BMP4 expression inhibits GSC growth both in vivo and in vitro, likely via the Smad signaling pathway. In vivo, hNSC-BMP4 treatment is effective in promoting GSC differentiation and apoptosis in xenograft gliomas, and improves the survival of mice bearing these tumors [74].
5.4. Biocompatible Nanomaterial Based Delivery
Self-assembling materials such as peptide amphiphiles (PAs) have been a focus of medical applications over the past two decades. PAs can be designed to self-assemble in cylindrical nanostructures that resemble the structural characteristics of native extracellular matrix (ECM) fibers. The molecular design of PAs allows for the incorporation of bioactive signals that will be displayed on the surface of the self-assembled nanofibers creating opportunities for exciting novel therapies with broad potential impact in regenerative medicine and cancer. Recently, Srikanth et al. [75] reported that PA nanofibers displaying an IKVAV peptide signal could be used to treat GSCs. They showed that this specific PA potentially increases immobilized β1-integrin at the GSC membrane, activating integrin-linked kinase while inhibiting focal adhesion kinase (FAK), which consequently induces apoptosis in GSCs. PA nanofibers can also be designed to display binding peptide sequences allowing the nanofibers to bind and deliver specific proteins, nucleic acids, drugs and cells [76]. For example, PA nanofibers as a delivery mechanism have been investigated to deliver BMP2. This particular PA nanofiber displays a peptide sequence found through phage-display techniques with an affinity for BMP2. The use of this binding nanofiber led to more efficient delivery and protein activity that resulted in a ten-fold dose reduction of BMP2 required for successful spinal fusion in a rat model [77]. More recently, Lee et al. [38] synthesized a novel sulfated glycopeptide nanostructure that has a binding affinity for multiple proteins including BMP4 (Figure 5). Most importantly, these PA nanostructures are biocompatible, thus they do not cause side effects while providing more efficient delivery to increase therapeutic benefit.
6. Future Prospects
Here, we have summarized recent BMP4 associated progress in aHGG and pHGG. BMP4 treatment could be a valuable adjunct to conventional therapies for these devastating tumors. However, BMP4 mediated differentiation therapies must be used in a patient-specific context since a subset of gliomas do not differentiate in response to BMP4 [21,78,79]. To better predict therapeutic value, the roles of BMP4 in subsets of aHGGs and pHGGs with specific molecular signatures should be further examined. The means delivering BMP4 is also a key factor. We described current BMP4 delivery strategies and proposed that biocompatible nanocarriers could be a novel highly efficient delivery platform. Further studies need to develop PA nanostructures for brain tumor treatment via systemic administration. These nanostructures must be designed to have high BMP4 affinity and to cross the blood-brain barrier. We hope advanced nanotechnology based on self-assembling peptides will enhance BMP4 delivery efficacy and lead to new therapeutic options that, in combination with conventional cytotoxic and/or radiation therapy, will improve outcomes for patients with HGGs.
Author Contributions
Conceptualization, J.A.K. and G.X.; data collection, Y.Y. and G.X.; writing—original draft preparation, S.N., A.M., B.M.-F., G.X.; writing—review and editing, M.M., B.M.-F., J.A.K., C.D.J., S.I.S., S.-Y.C., T.T., G.X.; funding acquisition, G.X., S.I.S. and T.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported, in part, by the National Cancer Institute SPORE grant (P50CA221747-01A1) Career Enhancement Award; and the Rory David Deutsch Foundation, the Surgical Neuro-Oncology Research Fund of Ann & Robert H Lurie Children’s Hospital (A&RLCH) of Chicago, and the Ralph and Marian C. Falk Medical Research Trust.
Acknowledgments
The in silico analyzed results presented here were in part based upon data generated by the TCGA Research Network: https://www.cancer.gov/tcga. We would also like to acknowledge our appreciation of the team who created GlioVis: Data Visualization Tools for Brain Tumor Datasets portal. We apologize to authors whose primary references could not be cited due to the space limitation.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ostrom, Q.T.; Gittleman, H.; Liao, P.; Vecchione-Koval, T.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary brain and other central nervous system tumors diagnosed in the United States in 2010–2014. Neuro-Oncology 2017, 19, v1–v88. [Google Scholar] [CrossRef]
- Aldape, K.; Brindle, K.M.; Chesler, L.; Chopra, R.; Gajjar, A.; Gilbert, M.R.; Gottardo, N.; Gutmann, D.H.; Hargrave, D.; Holland, E.C.; et al. Challenges to curing primary brain tumours. Nature reviews. Clin. Oncol. 2019, 16, 509–520. [Google Scholar] [CrossRef]
- Smith, J.S.; Jenkins, R.B. Genetic alterations in adult diffuse glioma: Occurrence, significance, and prognostic implications. Front. Biosci. 2000, 5, D213–D231. [Google Scholar] [CrossRef]
- Song, K.S.; Phi, J.H.; Cho, B.K.; Wang, K.C.; Lee, J.Y.; Kim, D.G.; Kim, I.H.; Ahn, H.S.; Park, S.H.; Kim, S.K. Long-term outcomes in children with glioblastoma. Journal of neurosurgery. Pediatrics 2010, 6, 145–149. [Google Scholar] [CrossRef]
- Johnson, B.E.; Mazor, T.; Hong, C.; Barnes, M.; Aihara, K.; McLean, C.Y.; Fouse, S.D.; Yamamoto, S.; Ueda, H.; Tatsuno, K.; et al. Mutational analysis reveals the origin and therapy-driven evolution of recurrent glioma. Science 2014, 343, 189–193. [Google Scholar] [CrossRef]
- Nishikawa, R. Pediatric and adult gliomas: How different are they? Neuro-Oncology 2010, 12, 1203–1204. [Google Scholar] [CrossRef]
- Sturm, D.; Bender, S.; Jones, D.T.; Lichter, P.; Grill, J.; Becher, O.; Hawkins, C.; Majewski, J.; Jones, C.; Costello, J.F.; et al. Paediatric and adult glioblastoma: Multiform (epi)genomic culprits emerge. Nature reviews. Cancer 2014, 14, 92–107. [Google Scholar] [CrossRef]
- Yan, H.; Parsons, D.W.; Jin, G.; McLendon, R.; Rasheed, B.A.; Yuan, W.; Kos, I.; Batinic-Haberle, I.; Jones, S.; Riggins, G.J.; et al. IDH1 and IDH2 mutations in gliomas. N. Engl. J. Med. 2009, 360, 765–773. [Google Scholar] [CrossRef]
- Capper, D.; Jones, D.T.W.; Sill, M.; Hovestadt, V.; Schrimpf, D.; Sturm, D.; Koelsche, C.; Sahm, F.; Chavez, L.; Reuss, D.E.; et al. DNA methylation-based classification of central nervous system tumours. Nature 2018, 555, 469–474. [Google Scholar] [CrossRef]
- Patel, A.P.; Tirosh, I.; Trombetta, J.J.; Shalek, A.K.; Gillespie, S.M.; Wakimoto, H.; Cahill, D.P.; Nahed, B.V.; Curry, W.T.; Martuza, R.L.; et al. Single-cell RNA-seq highlights intratumoral heterogeneity in primary glioblastoma. Science 2014, 344, 1396–1401. [Google Scholar] [CrossRef]
- Eskilsson, E.; Rosland, G.V.; Solecki, G.; Wang, Q.; Harter, P.N.; Graziani, G.; Verhaak, R.G.W.; Winkler, F.; Bjerkvig, R.; Miletic, H. EGFR heterogeneity and implications for therapeutic intervention in glioblastoma. Neuro-Oncology 2018, 20, 743–752. [Google Scholar] [CrossRef]
- Mackay, A.; Burford, A.; Carvalho, D.; Izquierdo, E.; Fazal-Salom, J.; Taylor, K.R.; Bjerke, L.; Clarke, M.; Vinci, M.; Nandhabalan, M.; et al. Integrated Molecular Meta-Analysis of 1,000 Pediatric High-Grade and Diffuse Intrinsic Pontine Glioma. Cancer Cell 2017, 32, 520–537.e525. [Google Scholar] [CrossRef]
- Wu, G.; Broniscer, A.; McEachron, T.A.; Lu, C.; Paugh, B.S.; Becksfort, J.; Qu, C.; Ding, L.; Huether, R.; Parker, M.; et al. Somatic histone H3 alterations in pediatric diffuse intrinsic pontine gliomas and non-brainstem glioblastomas. Nat. Genet. 2012, 44, 251–253. [Google Scholar] [CrossRef]
- Schwartzentruber, J.; Korshunov, A.; Liu, X.Y.; Jones, D.T.; Pfaff, E.; Jacob, K.; Sturm, D.; Fontebasso, A.M.; Quang, D.A.; Tonjes, M.; et al. Driver mutations in histone H3.3 and chromatin remodelling genes in paediatric glioblastoma. Nature 2012, 482, 226–231. [Google Scholar] [CrossRef]
- Buczkowicz, P.; Hoeman, C.; Rakopoulos, P.; Pajovic, S.; Letourneau, L.; Dzamba, M.; Morrison, A.; Lewis, P.; Bouffet, E.; Bartels, U.; et al. Genomic analysis of diffuse intrinsic pontine gliomas identifies three molecular subgroups and recurrent activating ACVR1 mutations. Nat. Genet. 2014, 46, 451–456. [Google Scholar] [CrossRef]
- Fontebasso, A.M.; Papillon-Cavanagh, S.; Schwartzentruber, J.; Nikbakht, H.; Gerges, N.; Fiset, P.O.; Bechet, D.; Faury, D.; De Jay, N.; Ramkissoon, L.A.; et al. Recurrent somatic mutations in ACVR1 in pediatric midline high-grade astrocytoma. Nat. Genet. 2014, 46, 462–466. [Google Scholar] [CrossRef]
- Taylor, K.R.; Mackay, A.; Truffaux, N.; Butterfield, Y.; Morozova, O.; Philippe, C.; Castel, D.; Grasso, C.S.; Vinci, M.; Carvalho, D.; et al. Recurrent activating ACVR1 mutations in diffuse intrinsic pontine glioma. Nat. Genet. 2014, 46, 457–461. [Google Scholar] [CrossRef]
- Singh, S.K.; Clarke, I.D.; Hide, T.; Dirks, P.B. Cancer stem cells in nervous system tumors. Oncogene 2004, 23, 7267–7273. [Google Scholar] [CrossRef]
- Hemmati, H.D.; Nakano, I.; Lazareff, J.A.; Masterman-Smith, M.; Geschwind, D.H.; Bronner-Fraser, M.; Kornblum, H.I. Cancerous stem cells can arise from pediatric brain tumors. Proc. Natl. Acad. Sci. USA 2003, 100, 15178–15183. [Google Scholar] [CrossRef]
- Bhaduri, A.; Di Lullo, E.; Jung, D.; Müller, S.; Crouch, E.E.; Espinosa, C.S.; Ozawa, T.; Alvarado, B.; Spatazza, J.; Cadwell, C.R.; et al. Outer Radial Glia-like Cancer Stem Cells Contribute to Heterogeneity of Glioblastoma. Cell Stem Cell 2020, 26, 48–63. [Google Scholar] [CrossRef]
- Das, S.; Srikanth, M.; Kessler, J.A. Cancer stem cells and glioma. Nat. Clin. Pract. Neurol. 2008, 4, 427–435. [Google Scholar] [CrossRef]
- Bao, S.; Wu, Q.; McLendon, R.E.; Hao, Y.; Shi, Q.; Hjelmeland, A.B.; Dewhirst, M.W.; Bigner, D.D.; Rich, J.N. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature 2006, 444, 756–760. [Google Scholar] [CrossRef]
- Weng, Q.; Wang, J.; Wang, J.; He, D.; Cheng, Z.; Zhang, F.; Verma, R.; Xu, L.; Dong, X.; Liao, Y.; et al. Single-Cell Transcriptomics Uncovers Glial Progenitor Diversity and Cell Fate Determinants during Development and Gliomagenesis. Cell Stem Cell 2019, 24, 707–723. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.; Yu, T.S.; McKay, R.M.; Burns, D.K.; Kernie, S.G.; Parada, L.F. A restricted cell population propagates glioblastoma growth after chemotherapy. Nature 2012, 488, 522–526. [Google Scholar] [CrossRef]
- Hussein, D.; Punjaruk, W.; Storer, L.C.; Shaw, L.; Othman, R.; Peet, A.; Miller, S.; Bandopadhyay, G.; Heath, R.; Kumari, R.; et al. Pediatric brain tumor cancer stem cells: Cell cycle dynamics, DNA repair, and etoposide extrusion. Neuro-Oncology 2011, 13, 70–83. [Google Scholar] [CrossRef]
- Parada, L.F.; Dirks, P.B.; Wechsler-Reya, R.J. Brain Tumor Stem Cells Remain in Play. J. Clin. Oncol. 2017, 35, 2428–2431. [Google Scholar] [CrossRef]
- Yokoyama, Y.; Watanabe, T.; Tamura, Y.; Hashizume, Y.; Miyazono, K.; Ehata, S. Autocrine BMP-4 Signaling Is a Therapeutic Target in Colorectal Cancer. Cancer Res. 2017, 77, 4026–4038. [Google Scholar] [CrossRef]
- Chiou, J.; Su, C.-Y.; Jan, Y.-H.; Yang, C.-J.; Huang, M.-S.; Yu, Y.-L.; Hsiao, M. Decrease of FSTL1-BMP4-Smad signaling predicts poor prognosis in lung adenocarcinoma but not in squamous cell carcinoma. Sci. Rep. 2017, 7, 9830. [Google Scholar] [CrossRef]
- Park, W.Y.; Hong, B.J.; Lee, J.; Choi, C.; Kim, M.Y. H3K27 Demethylase JMJD3 Employs the NF-kappaB and BMP Signaling Pathways to Modulate the Tumor Microenvironment and Promote Melanoma Progression and Metastasis. Cancer Res. 2016, 76, 161–170. [Google Scholar] [CrossRef]
- Martinez, V.G.; Rubio, C.; Martinez-Fernandez, M.; Segovia, C.; Lopez-Calderon, F.; Garin, M.; Teijeira, A.; Munera-Maravilla, E.; Varas, A.; Sacedon, R.; et al. BMP4 induces M2 macrophage polarization and favors tumor progression in bladder cancer. Clin. Cancer Res. 2017, 23, 7388–7399. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, H.; Zhao, F.; Lu, P.; Ge, C.; Li, H.; Hou, H.; Yan, M.; Chen, T.; Jiang, G.; et al. BMP4 administration induces differentiation of CD133+ hepatic cancer stem cells, blocking their contributions to hepatocellular carcinoma. Cancer Res. 2012, 72, 4276–4285. [Google Scholar] [CrossRef]
- Yacqub-Usman, K.; Duong, C.V.; Clayton, R.N.; Farrell, W.E. Epigenomic silencing of the BMP-4 gene in pituitary adenomas: A potential target for epidrug-induced re-expression. Endocrinology 2012, 153, 3603–3612. [Google Scholar] [CrossRef]
- Zhao, X.; Sun, Q.; Dou, C.; Chen, Q.; Liu, B. BMP4 inhibits glioblastoma invasion by promoting E-cadherin and claudin expression. Front. Biosci. 2019, 24, 1060–1070. [Google Scholar]
- Jin, X.; Nie, E.; Zhou, X.; Zeng, A.; Yu, T.; Zhi, T.; Jiang, K.; Wang, Y.; Zhang, J.; You, Y. Fstl1 Promotes Glioma Growth Through the BMP4/Smad1/5/8 Signaling Pathway. Cell. Physiol. Biochem. 2017, 44, 1616–1628. [Google Scholar] [CrossRef]
- Liu, S.; Yin, F.; Fan, W.; Wang, S.; Guo, X.R.; Zhang, J.N.; Tian, Z.M.; Fan, M. Over-expression of BMPR-IB reduces the malignancy of glioblastoma cells by upregulation of p21 and p27Kip1. J. Exp. Clin. Cancer Res. 2012, 31, 52. [Google Scholar] [CrossRef]
- Liu, B.; Chen, Q.; Tian, D.; Wu, L.; Dong, H.; Wang, J.; Ji, B.; Zhu, X.; Cai, Q.; Wang, L.; et al. BMP4 reverses multidrug resistance through modulation of BCL-2 and GDNF in glioblastoma. Brain Res. 2013, 1507, 115–124. [Google Scholar] [CrossRef]
- Raja, E.; Komuro, A.; Tanabe, R.; Sakai, S.; Ino, Y.; Saito, N.; Todo, T.; Morikawa, M.; Aburatani, H.; Koinuma, D.; et al. Bone morphogenetic protein signaling mediated by ALK-2 and DLX2 regulates apoptosis in glioma-initiating cells. Oncogene 2017, 36, 4963–4974. [Google Scholar] [CrossRef]
- Lee, S.S.; Fyrner, T.; Chen, F.; Álvarez, Z.; Sleep, E.; Chun, D.S.; Weiner, J.A.; Cook, R.W.; Freshman, R.D.; Schallmo, M.S.; et al. Sulfated glycopeptide nanostructures for multipotent protein activation. Nat. Nanotechnol. 2017, 12, 821–829. [Google Scholar] [CrossRef]
- Bond, A.M.; Peng, C.Y.; Meyers, E.A.; McGuire, T.; Ewaleifoh, O.; Kessler, J.A. BMP signaling regulates the tempo of adult hippocampal progenitor maturation at multiple stages of the lineage. Stem Cells 2014, 32, 2201–2214. [Google Scholar] [CrossRef]
- Bond, A.M.; Bhalala, O.G.; Kessler, J.A. The dynamic role of bone morphogenetic proteins in neural stem cell fate and maturation. Dev. Neurobiol. 2012, 72, 1068–1084. [Google Scholar] [CrossRef]
- Gross, R.E.; Mehler, M.F.; Mabie, P.C.; Zang, Z.; Santschi, L.; Kessler, J.A. Bone morphogenetic proteins promote astroglial lineage commitment by mammalian subventricular zone progenitor cells. Neuron 1996, 17, 595–606. [Google Scholar] [CrossRef]
- Xi, G.; Best, B.; Mania-Farnell, B.; James, C.D.; Tomita, T. Therapeutic Potential for Bone Morphogenetic Protein 4 in Human Malignant Glioma. Neoplasi 2017, 19, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, D.; Taylor, K.R.; Olaciregui, N.G.; Molinari, V.; Clarke, M.; Mackay, A.; Ruddle, R.; Henley, A.; Valenti, M.; Hayes, A.; et al. ALK2 inhibitors display beneficial effects in preclinical models of ACVR1 mutant diffuse intrinsic pontine glioma. Commun. Biol. 2019, 2, 156. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.R.; Vinci, M.; Bullock, A.N.; Jones, C. ACVR1 mutations in DIPG: Lessons learned from FOP. Cancer Res. 2014, 74, 4565–4570. [Google Scholar] [CrossRef]
- Bowman, R.L.; Wang, Q.; Carro, A.; Verhaak, R.G.; Squatrito, M. GlioVis data portal for visualization and analysis of brain tumor expression datasets. Neuro-Oncology 2017, 19, 139–141. [Google Scholar] [CrossRef]
- Bao, Z.; Zhang, C.; Yan, W.; Liu, Y.; Li, M.; Zhang, W.; Jiang, T. BMP4, a strong better prognosis predictor, has a subtype preference and cell development association in gliomas. J. Transl. Med. 2013, 11, 100. [Google Scholar] [CrossRef]
- Wu, Q.; Yao, J. BMP4, a new prognostic factor for glioma. World J. Surg. Oncol. 2013, 11, 264. [Google Scholar] [CrossRef]
- Rich, J.N.; Bao, S. Chemotherapy and cancer stem cells. Cell Stem Cell 2007, 1, 353–355. [Google Scholar] [CrossRef]
- Rich, J.N. Cancer stem cells in radiation resistance. Cancer Res. 2007, 67, 8980–8984. [Google Scholar] [CrossRef]
- Stupp, R.; Hegi, M.E. Targeting brain-tumor stem cells. Nat. Biotechnol. 2007, 25, 193–194. [Google Scholar] [CrossRef]
- Piccirillo, S.G.; Reynolds, B.A.; Zanetti, N.; Lamorte, G.; Binda, E.; Broggi, G.; Brem, H.; Olivi, A.; Dimeco, F.; Vescovi, A.L. Bone morphogenetic proteins inhibit the tumorigenic potential of human brain tumour-initiating cells. Nature 2006, 444, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, R.; Wu, M.; Johnson, K.; Kim, H.; Celebre, A.; Shahzad, U.; Graham, M.S.; Kessler, J.A.; Chuang, J.H.; Karamchandani, J.; et al. BMP signaling mediates glioma stem cell quiescence and confers treatment resistance in glioblastoma. Sci. Rep. 2019, 9, 14569. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.H.; Ewy, J.M.; Chen, J.; Wong, S.Y.; Tharp, K.M.; Stahl, A.; Kumar, S. Transcriptomic analysis reveals that BMP4 sensitizes glioblastoma tumor-initiating cells to mechanical cues. Matrix Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Neftel, C.; Laffy, J.; Filbin, M.G.; Hara, T.; Shore, M.E.; Rahme, G.J.; Richman, A.R.; Silverbush, D.; Shaw, M.L.; Hebert, C.M.; et al. An Integrative Model of Cellular States, Plasticity, and Genetics for Glioblastoma. Cell 2019, 178, 835–849. [Google Scholar] [CrossRef] [PubMed]
- Gorovets, D.; Kannan, K.; Shen, R.; Kastenhuber, E.R.; Islamdoust, N.; Campos, C.; Pentsova, E.; Heguy, A.; Jhanwar, S.C.; Mellinghoff, I.K.; et al. IDH mutation and neuroglial developmental features define clinically distinct subclasses of lower grade diffuse astrocytic glioma. Clin. Cancer Res. 2012, 18, 2490–2501. [Google Scholar] [CrossRef]
- Phillips, H.S.; Kharbanda, S.; Chen, R.; Forrest, W.F.; Soriano, R.H.; Wu, T.D.; Misra, A.; Nigro, J.M.; Colman, H.; Soroceanu, L.; et al. Molecular subclasses of high-grade glioma predict prognosis, delineate a pattern of disease progression, and resemble stages in neurogenesis. Cancer Cell 2006, 9, 157–173. [Google Scholar] [CrossRef]
- Sturm, D.; Orr, B.A.; Toprak, U.H.; Hovestadt, V.; Jones, D.T.W.; Capper, D.; Sill, M.; Buchhalter, I.; Northcott, P.A.; Leis, I.; et al. New Brain Tumor Entities Emerge from Molecular Classification of CNS-PNETs. Cell 2016, 164, 1060–1072. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef]
- Jones, C.; Baker, S.J. Unique genetic and epigenetic mechanisms driving paediatric diffuse high-grade glioma. Nature reviews. Cancer 2014, 14. [Google Scholar] [CrossRef]
- Fukuda, T.; Scott, G.; Komatsu, Y.; Araya, R.; Kawano, M.; Ray, M.K.; Yamada, M.; Mishina, Y. Generation of a mouse with conditionally activated signaling through the BMP receptor, ALK2. Genesis 2006, 44, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Caren, H.; Beck, S.; Pollard, S.M. Differentiation therapy for glioblastoma—Too many obstacles? Mol. Cell. Oncol. 2016, 3, e1124174. [Google Scholar] [CrossRef] [PubMed]
- Caren, H.; Stricker, S.H.; Bulstrode, H.; Gagrica, S.; Johnstone, E.; Bartlett, T.E.; Feber, A.; Wilson, G.; Teschendorff, A.E.; Bertone, P.; et al. Glioblastoma Stem Cells Respond to Differentiation Cues but Fail to Undergo Commitment and Terminal Cell-Cycle Arrest. Stem Cell Rep. 2015, 5, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liu, Y.; Zhang, J.; Yu, D.; Li, A.; Song, H.; Zhang, W.; Davis, D.; Gilbert, M.R.; Liu, F.; et al. Autocrine BMP4 Signaling Enhances Tumor Aggressiveness via Promoting Wnt/beta-Catenin Signaling in IDH1-mutant Gliomas. Transl. Oncol. 2019, 13, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Olmsted, E.A.; Kaplan, F.S.; Shore, E.M. Bone morphogenetic protein-4 regulation in fibrodysplasia ossificans progressiva. Clin. Orthop. Relat. Rew. 2003, 408, 331–343. [Google Scholar] [CrossRef]
- Calpe, S.; Correia, A.C.; Sancho-Serra Mdel, C.; Krishnadath, K.K. Comparison of newly developed anti-bone morphogenetic protein 4 llama-derived antibodies with commercially available BMP4 inhibitors. Mabs 2016, 8, 678–688. [Google Scholar] [CrossRef]
- Glaser, D.L.; Economides, A.N.; Wang, L.; Liu, X.; Kimble, R.D.; Fandl, J.P.; Wilson, J.M.; Stahl, N.; Kaplan, F.S.; Shore, E.M. In vivo somatic cell gene transfer of an engineered Noggin mutein prevents BMP4-induced heterotopic ossification. J. Bone Jt. Surg. Am. 2003, 85, 2332–2342. [Google Scholar] [CrossRef]
- Duggal, R.; Geissinger, U.; Zhang, Q.; Aguilar, J.; Chen, N.G.; Binda, E.; Vescovi, A.L.; Szalay, A.A. Vaccinia virus expressing bone morphogenetic protein-4 in novel glioblastoma orthotopic models facilitates enhanced tumor regression and long-term survival. J. Transl. Med. 2013, 11, 155. [Google Scholar] [CrossRef]
- Choudhury, S.R.; Hudry, E.; Maguire, C.A.; Sena-Esteves, M.; Breakefield, X.O.; Grandi, P. Viral vectors for therapy of neurologic diseases. Neuropharmacology 2017, 120, 63–80. [Google Scholar] [CrossRef]
- Tyler, M.A.; Ulasov, I.V.; Sonabend, A.M.; Nandi, S.; Han, Y.; Marler, S.; Roth, J.; Lesniak, M.S. Neural stem cells target intracranial glioma to deliver an oncolytic adenovirus in vivo. Gene Ther. 2008, 16, 262–278. [Google Scholar] [CrossRef]
- Li, Q.; Wijesekera, O.; Salas, S.J.; Wang, J.Y.; Zhu, M.; Aprhys, C.; Chaichana, K.L.; Chesler, D.A.; Zhang, H.; Smith, C.L.; et al. Mesenchymal stem cells from human fat engineered to secrete BMP4 are nononcogenic, suppress brain cancer, and prolong survival. Clin. Cancer Res. 2014, 20, 2375–2387. [Google Scholar] [CrossRef] [PubMed]
- Spaeth, E.L.; Dembinski, J.L.; Sasser, A.K.; Watson, K.; Klopp, A.; Hall, B.; Andreeff, M.; Marini, F. Mesenchymal stem cell transition to tumor-associated fibroblasts contributes to fibrovascular network expansion and tumor progression. PLoS ONE 2009, 4, e4992. [Google Scholar] [CrossRef] [PubMed]
- Mangraviti, A.; Tzeng, S.Y.; Gullotti, D.; Kozielski, K.L.; Kim, J.E.; Seng, M.; Abbadi, S.; Schiapparelli, P.; Sarabia-Estrada, R.; Vescovi, A.; et al. Non-virally engineered human adipose mesenchymal stem cells produce BMP4, target brain tumors, and extend survival. Biomaterials 2016, 100, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yin, F.; Zhao, M.; Zhou, C.; Ren, J.; Huang, Q.; Zhao, Z.; Mitra, R.; Fan, W.; Fan, M. The homing and inhibiting effects of hNSCs-BMP4 on human glioma stem cells. Oncotarget 2016, 7, 17920–17931. [Google Scholar] [CrossRef]
- Srikanth, M.; Das, S.; Berns, E.J.; Kim, J.; Stupp, S.I.; Kessler, J.A. Nanofiber-mediated inhibition of focal adhesion kinase sensitizes glioma stemlike cells to epidermal growth factor receptor inhibition. Neuro-Oncology 2013, 15, 319–329. [Google Scholar] [CrossRef]
- Webber, M.J.; Kessler, J.A.; Stupp, S.I. Emerging peptide nanomedicine to regenerate tissues and organs. J. Intern. Med. 2010, 267, 71–88. [Google Scholar] [CrossRef]
- Lee, S.S.; Hsu, E.L.; Mendoza, M.; Ghodasra, J.; Nickoli, M.S.; Ashtekar, A.; Polavarapu, M.; Babu, J.; Riaz, R.M.; Nicolas, J.D.; et al. Gel scaffolds of BMP-2-binding peptide amphiphile nanofibers for spinal arthrodesis. Adv. Healthc. Mater. 2015, 4, 131–141. [Google Scholar] [CrossRef]
- Lee, J.; Son, M.J.; Woolard, K.; Donin, N.M.; Li, A.; Cheng, C.H.; Kotliarova, S.; Kotliarov, Y.; Walling, J.; Ahn, S.; et al. Epigenetic-Mediated Dysfunction of the Bone Morphogenetic Protein Pathway Inhibits Differentiation of Glioblastoma-Initiating Cells. Cancer Cell 2008, 13, 69–80. [Google Scholar] [CrossRef]
- Yan, K.; Wu, Q.; Yan, D.H.; Lee, C.H.; Rahim, N.; Tritschler, I.; DeVecchio, J.; Kalady, M.F.; Hjelmeland, A.B.; Rich, J.N. Glioma cancer stem cells secrete Gremlin1 to promote their maintenance within the tumor hierarchy. Genes Dev. 2014, 28, 1085–1100. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).