A Review of the Enhancement of Bio-Hydrogen Generation by Chemicals Addition


1
Department of Chemical and Environmental Engineering, University of Nottingham Ningbo, Ningbo 315100, China
2
School of Engineering, Edith Cowan University, 270 Joondalup Drive, Joondalup, WA 6027, Australia
3
State key Laboratory of Biochemical Engineering, Institute of Process Engineering, Chinese Academy of Sciences, Beijing 100190, China
4
The Commonwealth Scientific and Industrial Research Organization (CSIRO), Energy Business Unit, Canberra, WA 6151, Australia
*
Authors to whom correspondence should be addressed.
Catalysts 2019, 9(4), 353; https://doi.org/10.3390/catal9040353
Submission received: 10 March 2019
/
Revised: 30 March 2019
/
Accepted: 7 April 2019
/
Published: 11 April 2019
(This article belongs to the Special Issue Recent Advances in Biocatalysis and Metabolic Engineering for Biomanufacturing)
Abstract
:Bio-hydrogen production (BHP) produced from renewable bio-resources is an attractive route for green energy production, due to its compelling advantages of relative high efficiency, cost-effectiveness, and lower ecological impact. This study reviewed different BHP pathways, and the most important enzymes involved in these pathways, to identify technological gaps and effective approaches for process intensification in industrial applications. Among the various approaches reviewed in this study, a particular focus was set on the latest methods of chemicals/metal addition for improving hydrogen generation during dark fermentation (DF) processes; the up-to-date findings of different chemicals/metal addition methods have been quantitatively evaluated and thoroughly compared in this paper. A new efficiency evaluation criterion is also proposed, allowing different BHP processes to be compared with greater simplicity and validity.
1. Introduction
To effectively curb the world emissions from fossil-based energy by 2030 [1,2], attempts of exploring alternative renewable energy have been made worldwide in both scientific and industrial communities in the past decades [3,4]. Due to its great features, such as having the highest energy density among other fuels and complete cleanness after combustion, hydrogen has attracted a lot of attention as an energy carrier [5]. However, the current existing hydrogen generation processes have been dominated by the conventional routes of natural gas steam reforming (SR), natural gas thermal cracking, coal gasification, and partial oxidation of the heavier-than-naphtha hydrocarbons, which use fossil fuel as feedstock, are energy-intensive, and less environmentally friendly [6,7]. Although direct water-splitting via a semi-conductive photocatalyst to produce renewable hydrogen has recently attracted much interest [8], the significant bottom neck of very low efficiency still remains a big technical hurdle to be overcome for its short- and middle-term industrial application. On the other hand, biological processes for hydrogen generation possess many intrinsic appealing advantages, such as simplicity in operation, wide availability of renewable feed stocks (such as agricultural waste and food waste), carbon neutrality, and cost-effectiveness in operation [9,10,11,12,13,14]. Bio-hydrogen production can be achieved by two kinds of biological processes: (1) light-dependent, and (2) light-independent. For photo-dependent processes, it could be further divided into the photolysis and photo-fermentation subcategories. For the photo-independent processes, hydrogen generation is achieved by dark fermentation (DF). For all of these processes, the bio-catalyst hydrogenase ([FeFe], [NiFe], [Fe]) might be the most significant catalyst for the evolution of hydrogen. Among all of these above-mentioned processes, DF is one of the most promising due to its appealing features of simplicity of operation, relatively high hydrogen conversion, flexibility in cultivation, and simultaneous realization of hydrogen production and organic waste consumptions [15]. Therefore, the effective enhancement of BHP during DF has become a research focus among scholars in the last decades. Many approaches have been found to effectively enhance hydrogen generation during the DF, which include pretreatment (e.g., ultrasonic, acid/base, enzyme hydrolysis), optimized operation (e.g., hydraulic retention time), co-fermentation, genetic engineering, and chemical addition [16,17,18]. Some of those approaches will directly or indirectly affect the hydrogenase biocatalyst, while some others might affect the other metabolic pathways or the growth of microbes, which ultimately accelerate or inhibit hydrogen generation [19,20]. From a practical perspective, the chemical addition is more feasible compared with other approaches mentioned above. This is the reason why the numbers of reports in regard to process intensification by chemical addition have been growing very rapidly in recent years [21]. Therefore, this motivated us to review the recent progress of chemical addition, such as metal monomers, metal oxides, nanoparticles (NPs), and synergistic factors that potentially affected the activities of a hydrogenase biocatalyst and consequently led to increased hydrogen generation. The hydrogen production rates will be quantitatively compared among these different works. In this review paper, to avoid repetitive summary and discussion that had been addressed by other scholars, we only focus on chemical addition that could potentially affect the activity of hydrogenase during DF for BHP.
2. Enzyme System in Bio-Hydrogen Generation
Hydrogen generation via biological processes can be achieved by a series of biological electrochemical reactions. These reactions are facilitated by a series of biocatalyst enzymes that are found to play critical roles during the BHP. There are three main bio-hydrogen production and consumption enzymes, which are responsible for the net bio-hydrogen evolution. These three different enzymes are reversible hydrogenase, membrane-bounded uptake hydrogenase, and nitrogenase enzymes. Among them, nitrogenase and hydrogenase are the two pivotal biocatalysts [22].
2.1. Functions of Nitrogenase
Hydrogen generation can be catalyzed by nitrogenase under an anaerobic environment at photofermentation conditions from photosynthetic bacteria. Nitrogenase is well-known for fixing the nitrogen molecule, and is commonly found in archaea and bacteria. While the nitrogen molecule is catalyzed into ammonia by the nitrogenase, hydrogen gas is generated as a by-product, and the entire chemical redox balance is maintained during this biological catalytic nitrogen fixation process, which is summarized in Equation (1) below:
Hydrogen generation catalyzed by nitrogenase is thermodynamically regarded as an energy-intensive and irreversible reaction, which consumes four moles of adenosine triphosphate (ATP) per mole of bio-hydrogen produced. Ammonia (product) removal and an anaerobic condition is critical to hydrogen generation. A schematic diagram of the structure of nitrogenase is shown in Figure 1.
The typical structure of nitrogenase consists of three metal-containing cofactors, which are the iron-sulfur cluster, P cluster, and FeMo cluster. The iron–sulfur cluster serves a critical role in delivering electrons to the FeMo cluster via the P cluster. The FeMo serves as an active site for dinitrogen reduction to ammonia. The nitrogenase enzyme widely exists in the photofermentation in archaea and bacteria. Factors such as chemical additions that either enhance or suppress the activity of nitrogenase will result in a variation of hydrogen evolution. Taking the purple non-sulfur bacteria (PNSB), for example, under nitrogen-deficient conditions, the turnover from the nitrogenase is continuous, reducing the protons into H2. During each circle, at the Fe–S cluster associated with FeMo-co, 2 ATP are hydrolyzed with the transfer of one electron to the MoFe protein and the complex dissociates. The entire turnover is extremely slow at 6.4 s−1, and added to its additional great deal of enzymatic machinery, energy, and time used for the biosynthesis of these complex metal centers, it consequently results in low efficiency [24].
2.2. Functions and Classification of Hydrogenase
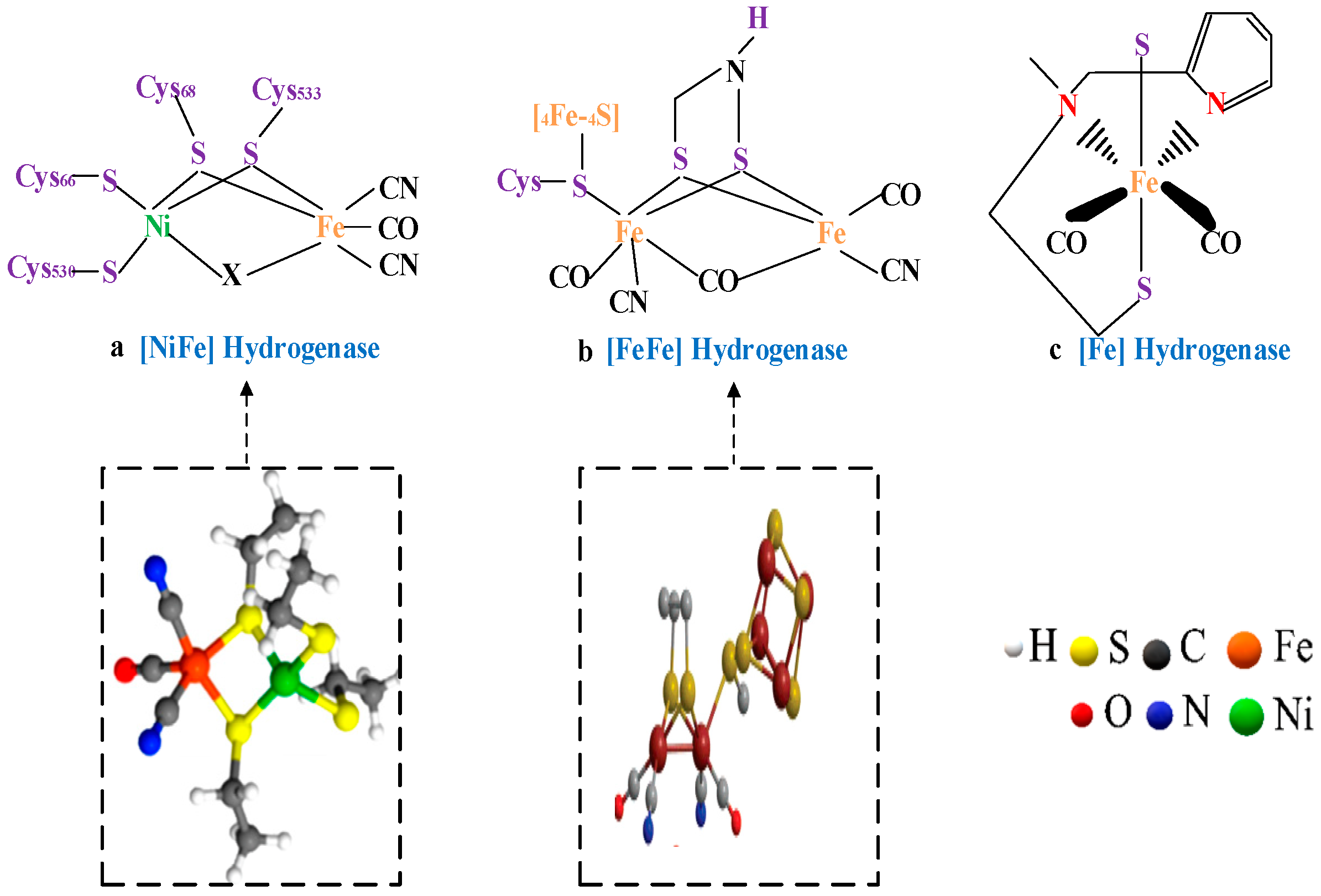
The uncovering of the molecular structure of hydrogenase began from the first report of the atomic structure of the D.gigas enzyme [25]. The metal centers, which are the active sites of the biocatalyst, can be broadly classified into three different types, namely the [NiFe], [FeFe], and [Fe] types [26,27]. The [FeFe] hydrogenase catalyzes the oxidation of H2, as well as the reduction of H+, but the enzyme is mainly found in the H2 generating process, whereas the [NiFe] hydrogenase catalyzes the consumption of hydrogen [28]. The detailed schematic diagram of the molecular structure of hydrogenase is shown in Figure 2. The active site of [NiFe] hydrogenase is a dinuclear thiolate-bridged Ni-Fe complex. The [FeFe]-hydrogenases’ active sites are organized into modular domains with accessory clusters functioning as inter- and intra-molecular electron-transfer centers electronically linked to the catalytic H-cluster. Hydrogenases, especially the [FeFe] hydrogenase, are sensitive to the presence of oxygen (which is only active under strictly anaerobic conditions). However, studies have shown that the [NiFe] hydrogenases present better O2 tolerance than the hydrogenase with [FeFe] metal centers [29]. In the metal center of a [NiFe] type hydrogenase, the active site usually contains two cis nickel (Ni) coordination sites available for substrate binding, a bridging site and a terminal Ni site, with the Ni site terminally bound to the thiolate of Cysteine 530 [30]. [NiFe] widely exists in bacteria during hydrogen fermentation, while the [FeFe]-type hydrogenase can only be found in a few microbial species, such as green algea Chlamydomonas reinhardtii [31,32].
Although the two biocatalysts—namely, the nitrogenase and the hydrogenase—show completely different structures and catalyze hydrogen generation via completely different reaction pathways, these two types of biocatalyst sometimes coexist within the cell of one microbe. Therefore, the addition of metal elements such as nickel or iron will affect the activity of these metal-based biocatalysts, which, in turn, will enhance or inhibit the hydrogen generation.
3. Bio-Hydrogen Production Pathways
Bio-hydrogen production can be achieved by two major processes: (1) light-dependent processes, and (2) light-independent processes. The light-dependent processes can be accomplished by biophotolysis and photofermentation processes, while the light-independent processes can be realized by dark anaerobic fermentation. A conceptual illustration of the bio-hydrogen production pathway is shown in Figure 3.
3.1. Biophotolysis Process
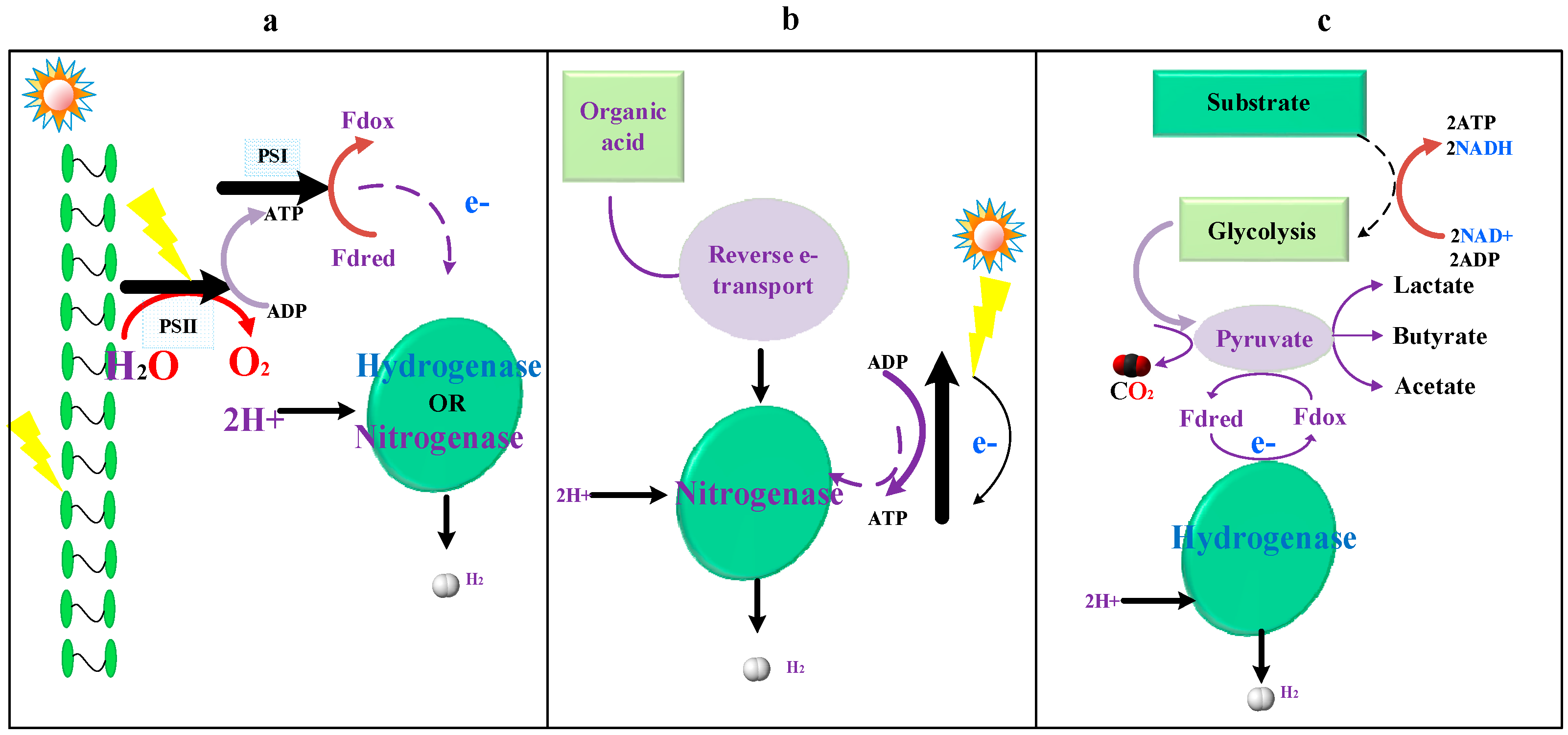
In biophotolysis (BP), the hydrogen ion is catalyzed either by nitrogenase or hydrogenase ([FeFe] [Fe]) to produce hydrogen gas in the presence of light within the cells of microbes (Figure 3a). Species such as algae and cyanobacteria ([NiFe]-type hydrogenase) are able to adopt this pathway to produce this zero-emission hydrogen gas from sunlight radiations [33]. BP can be further classified into two subcategories—direct biophotolysis, and indirect photolysis [34]. Many microbes, such as green algae or cyanobacterium, which are able to harvest solar energy to drive the water-splitting process to produce O2 and reduce the ferredoxin-an electron carrier in the chloroplasts, are able to perform biophotolysis via this direct BP pathway [35]. The water-splitting reaction is catalyzed by the photosynthesis system 2 (PSII) under anaerobic conditions, leading to the formation of hydrogen. The amount of electrons is linearly transferred from water to the ferredoxin, driven by the light energy harvested by PSI and PSII in the absence of oxygen. The reduced ferredoxin then donates the obtained electrons from PSI to the enzyme (hydrogenase or nitrogenase) to form hydrogen gas from protons. This entire pathway is shown in Figure 3a.
The microbes that are able to produce hydrogen via the BP process includes the following: Chlamydomonas reinhardtii, Chlorella fusca, Scenedesmus obliquus, Chlorococcum littorale, and Platymonas subcordiformis [36]. Due to the powerful suppressive effect of the oxygen as a by-product of PSII, the entire hydrogen generation process, including gene expression, mRNA stability, and enzymatic catalysis, will be strongly negatively influenced. Therefore, effective approaches in enhancing direct BP should focus on how to effectively remove or purge the oxygen produced from the system [37,38].
Another route of BP is indirect biophotolysis, of which oxygenic photosynthesis and hypoxic nitrogen fixation reactions are spatially separated from each other. Indirect biophotolysis is widely adopted by cyanobacteria. These are mostly filamentous, and nitrogen fixing in the specialized cell is known as heterocysts. Genera, such as Nostoc, Anabaena, Calothrix, Oscillatoria, are able to produce hydrogen via this indirect photolysis. Some non-nitrogen fixing genera, such as Synechocystis, Synechococcus, and Gloebacter, are also reported to possess this indirect BP pathway [33,36,39].
3.2. Photofermentation Process
In photofermentation (PF), the reduction of ferredoxins and generation of ATP is achieved via the reverse electron flow, driven by harvested solar energy, with the purple non-sulfur photosynthetic bacterium (PNS) under anaerobic conditions [40,41]. Instead of obtaining electrons from water-splitting reactions, as in the direct photolysis that exists in microalgae or cyanobacteria, the organic compounds, such as organic acid, acts as an electron donor under anaerobic conditions in the PNS bacterium. The schematic diagram illustrating this indirect photofermentation is shown in Figure 3b. The hydrogen generated via this pathway has appealing advantages: (1) complete substrate conversion to H2 and CO2; (2) removal of the adverse effect of oxygen that inhibits the activity of [FeFe] hydrogenase, hoxEFUYH [NiFe] hydrogenase, and nitrogenase enzymes [42,43,44,45]; (3) effective utilization of sunlight in both visible (400–700 nm) and near-infrared (700–950 nm) regions of the solar light spectrum; (4) wide availability of organic compounds used as an electron donor; (5) a relatively lower energy barrier to overcome, compared with water-splitting in direct photolysis [46]. Species that are able to produce the hydrogen via this photofermentation process include the following: Rhodospirillum rubrum, Rhodopseudomonas palustris, Rhodobacter sphaeroides, and Rhodobacter capsulatus [47,48].
3.3. Dark Fermentation Process
The essence of dark fermentation (DF) is the catalytic reaction of converting organic substrates into hydrogen under anaerobic conditions. Instead of harvesting the energy from solar light, the energy used to drive the neutralization reaction between the proton (H+) and electrons (e−) to form hydrogen comes from the microbial metabolic oxidation of organic substrates, such as glycolysis of glucose to the pyruvate intermediate. Complicated metabolic products are produced during DF. The product distribution of metabolic products varies significantly with the varieties of microbes, the oxidation of the substrate, and the environmental conditions, such as pH, hydrogen partial pressure, and level of nutrition [49,50]. Taking the glycolysis pathways as an example (Figure 3c), the ATP is generated through the substrate phosphorylation and energy-yielding reactions, including the formation of redox equivalents, such as the reduction of oxidized nicotinamide adenine dinucleotide (NAD+) to nicotinamide adenine dinucleotide (NADH). The produced pyruvate intermediate is then reduced by the produced redox equivalents to form intermediary metabolites, and eventually leads to lactate, CO2, and ethanol formation [51]. Another fermentation pathway includes the transformation of pyruvate to acetyl-coenzyme A (Acetyl-CoA), accompanied by the formation of an additional redox equivalent, CO2 and formate, and eventually leading to the splitting of Acetyl-CoA, and generation of ATPs and acetate [52,53]. The routes for forming molecular hydrogen can be expressed by Equations (2)–(4):
where Fd represents ferredoxins.
These hydrogen-generation reactions are catalyzed by hydrogenase under an anaerobic condition. There are many appealing advantages of generating hydrogen via these DF pathways: (1) relative simplicity in hydrogen generation, with higher conversion, production efficiency, and lower energy input; (2) versatile feedstock, such as organic food waste or inorganic waste, used for the fermentation; (3) the anaerobic conditions will create a favorable state for maintaining better activity of the biocatalyst for both [NiFe] and [FeFe] hydrogenase, and result in a relatively larger yield of hydrogen; (4) the bio-hydrogen fermentation process is flexible to create either a pure or mixed cultivation of the microbes. A brief comparison of different bio-hydrogen pathways, their corresponding technical challenges, and their effective approaches for hydrogen generation enhancement are summarized in Table 1.
Although technical hurdles and challenges still remain for these three hydrogen generation routes, the DF is still among the most promising technical route for BHP, which attracts great research interests and has even been successfully established at a pilot scale [26,78]. Therefore, DF will be the focus of our subsequent discussion for the enhancement of BHP by chemical additions.
4. Metal Additives
Although many attempts at process intensification, such as pretreatment, process optimization, and co-fermentation have been found to be effective in enhancing hydrogen production, the supplementation of additives have attracted much attention in DF due to its simplicity and cost-effectiveness compared with other approaches of process intensification [21]. Among different kinds of supplementation of additives, metal additives are among one of the most widely employed. It has been found that trace metals play a significant role during the anaerobic fermentation process, especially for the activities of the hydrogenase [79]. The addition of metal into fermentation media has been identified to have the following beneficial effects: (1) facilitation of intracellular electron transportation, and (2) provision of essential nutrition for microbial growth. In this paper, attention will be focused on the effects of metal addition on bio-hydrogen generation. For convenience of discussion, the chemical additions are further divided into subcategories, including monomer, metal ion, metal oxide, and others, such as chemical addition, together with the combination of different types of operations, such as immobilizations.
4.1. Metal Monomers
The addition of metal monomers, such as Fe0 and Ni0 during DF, were found to be able to enhance hydrogen generation. The effects of these added metal monomers could be broadly divided into the two categories: (1) directly affects the activity of the biocatalyst; (2) affects the complicated metabolic pathways during DF that leads to enhanced hydrogen generation. Results for the addition of various metal monomers are shown in Table 2. With the addition of different metal monomers, the hydrogen yield was enhanced by different factors, from 10% to 110%, compared with that of the control test without metal addition, depending on the specific conditions such as different inoculum, substrates, or fermentative conditions.
Among the different types of metal monomers, the iron metal monomers are the most promising due to their appealing advantages of relative low cost, and effectiveness in affecting the activity of hydrogenase [21]. In addition, the oxidative-reductive potential (ORP) of fermentation solution could be reduced by the addition of zero-valent iron, which in turn creates a thermodynamically favorable environment for the growth of bacteria. Besides zero-valent metal monomers, the addition of nanosize zero-valent metal monomers, such as iron, nickel, or gold nanoparticles (NPs) began to attract attention due to the unique surface size and quantum size effect. The addition of iron or nickel NPs will facilitate the acceleration of electron transfer between the ferredoxin and hydrogenase to drive hydrogen generation. In addition, the added zero-valent Fe or Ni NPs could be oxidized into metal ions, such as Fe2+ or Ni2+, via the anaerobic corrosion process, which will potentially produce very similar beneficial effects upon BHP as those metal ions of Fe2+ or Ni2+ addition do during the fermentation.
Based upon current reports, one of the highest improvements in hydrogen generation (+110%) could be achieved by adding Ni (37.5 mg/L) and Fe (37.5 mg/L) NPs together during the DF [82]. This surely indicates that the improved electron transfer enhances the overall activity of hydrogenase. However, instead of continuously enhancing the hydrogen generation, an overdose of the metal monomer starts to produce adverse effects upon hydrogen production, as reported in previous research [70]. This indicates that too high a concentration of metal monomers can be harmful for both the activities of hydrogenase, and for other metabolic pathways that indirectly affect hydrogen generation. Therefore, the optimal condition that meets both maximum performance and cost-effectiveness of operation exists. From Table 2, it is not difficult to identify the existing challenges and limitations: (1) there is no consistent quantitative evaluation standard for the assessment of BHP, leading to difficulties in comparing the performance of the different metal monomers; (2) most of the DF focuses on using the sugars, such as glucose or sucrose, and there are very limited efforts at investigating the effect of an addition of metal monomers to the biocatalyst hydrogenase using other types of organic substrates, such as food wastes; (3) the operation of DF was mostly conducted in batch operation, which ends up with continuous inhibitory intermediates accumulation during the DF [90].
4.2. Metal Ions
Metal ion is one of the most common additives that could be used to enhance the catalysts’ performances during the DF. The iron ion is widely employed, not only because of its relative cost-effectiveness compared with other metal ions, but also because of its essential role in the constitutions of hydrogenase and ferredoxin. Like the functionality of iron ions, the role of nickel ion in enhancing the activities of hydrogenase also appears to be obvious. According to the works reported by Grafe and Friedrich, the nickel ion has been found in several hydrogenase and nickel-dependent uptake hydrogenase [91]. According to Zhang et al. [92], the addition of nickel ion directly stimulated the activity of hydrogenase. According to the different hypothesis available, the availability of nickel to a cell may affect the activity of the biocatalyst itself or affect the synthesis of other protein, which, in the end, will contribute to the enhancement of hydrogen evolution [91,92].
A summary of the usage of different metal ions, mainly Fe2+/Fe3+ and Ni2+, as additives during DF using different types of substrates, such as sugars, wastewater, and food waste is shown in Table 3. From this, it can be seen that the addition of metal irons, especially Fe2+ and Ni2+, are effective in enhancing bio-hydrogen fermentation. The role of the metal irons, such as Fe2+/Fe3+ and Ni2+, is found to facilitate both the increase of biomass (cell growth) and hydrogen production during the DF. From the work reported by Hisham et al. [93], it was found that the addition of metal elements, such as Ca2+ or Mg2+ metal ions, led to a significant decrease in hydrogen generation (−30%, −70%), while the biomass experienced a steady increase up to 40%. According to [94], by adding ferrous chloride during DF, the hydrogen generation was enhanced by 650% (increased to 130 ml.g−1). Although the improvement compared to the baseline was significant in that work, the absolute value of hydrogen produced (650 mL of cumulative hydrogen production) was marginal compared with other literature reports (which is often over 1000 mL cumulative hydrogen production within the similar duration of cultivation) [90]. Apart from the addition of singular ion, the hybrid mixtures, such as Fe-Ni or Ni-Mg-Al (hydrotalcite), were also found to be effective in enhancing the hydrogen production. Their addition was found to increase the hydrogen generation to about 70–80%.
From the above discussion, it can be suggested that the roles of the different types of metal ions during DF are completely different. The addition of Fe2+/Fe3+, Ni2+ or the mixture Fe–Ni seems to directly affect the activity of hydrogenase, and therefore the bio-hydrogen generation process could be directly manipulated by adding these types of metal ions. However, in regard to other metal ions, such as Ca2+, Mg2+, Cu2+, Na+, it seems that these metal ions tend to affect the growth of cell mass or indirectly influence other relevant metabolic pathways during the DF, of which no obvious hydrogen production improvement was observed by these types of metal ion additions.
4.3. Metal Oxide
Metal oxides play a very similar role as to metal ions. In recent studies, it has been found that the reduced size of metal oxides (nanoparticle size) will be favorable to the electron transfer between ferredoxin and [NiFe] or [FeFe]-based hydrogenase, which in turn accelerates the catalytic reactions of hydrogen generation [110]. The summary of adding metal oxides and their corresponding BHP performance is shown in Table 4. Various kinds of metal oxides, such as TiO2, CoO, Fe2O3, NiO, and their mixtures (Fe2O3/NiO), and substrates such as glucose, organic wastewater, glucose, and starch, were used in previous studies, conducted mostly in batch operation under mesophilic conditions. The addition of NiO2 NPs, together with a co-addition of other NPs, were also found to enhance hydrogen generation due to facilitations of the electron transfer between ferredoxin and hydrogenase [111]. Therefore, the addition of metal oxides, especially with nanoparticle size, is another effective approach in directly enhancing the activities and performances of hydrogenase, which, in turn, will boost BHP.
4.4. Others
A summary of BHP with synergistic effects are compared in Table 5. Microbial immobilization is one of the most widely employed approaches used to prevent biomass wash-out when hydrogen evolution rate (HRE) is low during continuous operation. The appealing advantages of employing microbial or cell biocatalyst immobilization include: (a) tolerance toward the perturbation of environmental factors, such as temperature, pH, and accumulation of inhibitory intermediates; (b) higher bio-catalytic activity; and (c) higher process stability [121]. Table 6 summarizes the performances of different microbial immobilizer additions during DF [56]. Clearly, with the implementation of immobilization, the BHP is enhanced at different levels. Many researchers have found that immobilization supports, such as activated carbon (AC) or biochar (BC), tend to form a favorable thermodynamic chemical redox potential, which makes the hydrogen generation catalyzed by the hydrogenase run more effectively [70].
Apart from the immobilization approach, the conditions of operation are also found to be effective in enhancing BHP. For example, the hydrogen production yield was higher in continuous BHP, compared with the batch operation [129,131]. This is possibly due to the effective removal of inhibitory metabolic intermediates, which creates a favorable chemical environment for hydrogenase to catalyze the hydrogen formation reaction [132,133]. Nonetheless, although continuous operation could be more appealing compared to batch operation for large-scale production, cells washing-out is one of the critical problems that need to be carefully handled during continuous operation. Furthermore, it is also interesting to find that synergistic effects, such as the addition of immobilized support, together with nanoparticles such as nickel NPs, have a positive effect on BHP. This suggests that these additions directly affect the activities of biocatalyst hydrogenase, the electron transport, and the endurance to environmental perturbation, which in turn boosts hydrogen generation during the DF.
4.5. Results Comparison
In this work, we tried to summarize all the relevant reported works on BHP that we cited in order to find out some quantitative trends on the basis of substrate conversion efficiency (YH2/S) expressed in the mole of hydrogen produced per mole of substrate in mol mol−1, hydrogen evolution rate (HER) expressed in mmol L−1 h−1, and specific hydrogen production rate (qH2) expressed in mmol g−1 h−1. It has been addressed by many scholars that the C-molar-based mass balance is necessary when hydrogen yield and rate are expressed during DF [78]. The failure to present mass balance and kinetic data can lead to poor quality assurance and difficulties in quantitative comparison for dark fermentative BHP. It is necessary to set up presentation standards for hydrogen yield and rate, for the convenience of communication and cross-referencing throughout the scientific community.
Due to the limited number of literature and some inconsistences in the presentation of yields and rates due to the omission of mass balances during DF in some reported works, we only used data from literature reports with complete mass balance and rate expressions, and made a limited number of comparisons in regard to YH2/S versus HER, and YH2/S versus qH2 during DF. The results are presented in Figure 4. In this work, for the convenience of comparisons, we divided the chemical additions into the five different categories, which are metal NPs, metal monomers, metal irons, metal oxide, and others (metals other than iron or nickel), respectively. In addition, among these five different categories for BHP, the different kinds of synergistic effects, such as operations (continuous stirred tank reactor CSTR) or microbial immobilization, were all considered and counted. Within the same category, the YH2/S, HER, and qH2 were averaged, then based upon the calculated mean values (YH2/S, HER and qH2), the deviations were calculated using the following:
where N represents the numbers of sample size, is the sample mean, and xi is one sample value. The detailed calculations of those values of YH2/S, HER, and qH2, together with their corresponding deviations, could be found from previous reports [51,90]. In regard to the variation of the deviations calculated from those reported, there are many different factors that could be attributed to the large deviations observed with the different metal additions for BHP: (a) different microbial strains, (b) different ways of operation, such as batch or CSTR, (c) different substrates, such as glucose and sucrose employed during DF.
From the comparison, the enhancement of the activities of nickel contained hydrogenase can be broadly divided into three different regions. In the first region, the addition of nano-size particles (together with other synergistic factors, such as adding immobilized supports like AC or BC) is found to be relatively more effective in facilitating the bio-catalytic hydrogen reactions, on the basis of both substrate conversion efficiency and hydrogen evolution rate. The added NPs (such as Ni, Fe, or Ni/Fe) not only facilitate the electrons transfer, but also are engaged in other synergistic factors, such as immobilization and improved operations, that ultimately improve the BHP under anaerobic conditions [70]. In the second region, the metal monomer, metal ions, and metal oxides show very similar performances in enhancing the activities of nickel- or iron-containing biocatalysts during BHP. However, their effects tend to be complex: not only do they possibly affect the activities of the hydrogenase, but these additions might also affect other metabolisms or pathways and cell growth, which will eventually contribute to the increase of BHP. The third region is for those metals other than Ni, Fe, which are relatively less effective in enhancing the BHP, and it was found that some of them are even toxic to either the activities of the hydrogenase or the growth of microbes. Therefore, the addition of these materials is not recommended for the enhancement of the activities of hydrogenase during BHP.
5. Economic Perspective of Different Hydrogen Generation Routes
At the present time, hydrogen is predominantly produced by thermal technologies on the commercial scale, via SR, partial oxidation (POX), and autothermal reforming (ATR). The most widely used and economical approach of hydrogen production is via the steam reforming of methane (natural gas) (SMR), which nearly accounts for 90% of the world’s hydrogen generation, at a cost of U.S.$ 7/GJ [7,134]. One of the thorny challenges for these thermochemical processes lies in the simultaneous generation of greenhouse gases that needed to be captured and stored [135,136] or indirectly converting the produced CO2 back into hydrocarbons via catalytic processes, such as Fischer-Tropsch (FT) synthesis [137,138], which will inevitably increase the cost of the entire hydrogen generation process by about 20 to 40% [139]. The alternative route of replacing fossil-based hydrocarbons with carbon-neutral biomass leads to a doubling of the cost of hydrogen production (about US $14–15/GJ, depending on the types of feedstock and conversion routes), which makes the biomass thermochemical process much less competitive and alluring. Another promising technical route of hydrogen production on a large scale is by the electrolysis of water [140]. However, converting higher-grade electrical energy into relatively lower-grade chemical energy, such as hydrogen and oxygen, is found to be contradictory to the practice of energy cascade utilization, let alone it being more mature and cost-effective to store and transport electricity compared to the hydrogen.
Apart from conventional centralized hydrogen production, on-site and decentralized small-scale hydrogen generation, which possesses the advantages of lowering the prices of transport and onsite utilization of non-usable biomass with high water content, has begun to attract more and more interest. The biological hydrogen generation process is found to be perfectly suitable for small-scale and decentralized hydrogen production using those non-usable biomasses with high water content, with the cost of hydrogen production varying from 10 to 20 U.S.$/GJ, which could be further improved via the R&D impetus in the foreseeable future. Holladay et al. recently made a comparison among different technologies for hydrogen generation synoptically [56] and the results are summarized in Table 6.
Clearly, BHP using biomass as feedstock shows very appealing effectiveness, let alone if further considering competitive and beneficial characteristics, such as the reduced environmental impact, and relative simplicity in operation compared with the thermochemical and electrochemical processes. In addition, compared with photo-fermentation, DF presents very high efficiency, has a lower footprint, and is independent of solar energy. Therefore, it is envisioned that the effective approach of enhancing the activities of biocatalysts for BHP via DF is pivotal for highly efficient hydrogen production.
6. Future Perspectives
It is apparent that the enhancement of BHP during DF by process intensification and optimization has begun to approach its technical bottleneck at the current stage. From an energy cascade utilization and material recycling and reused perspective, the future for hydrogen production needs to implement multistage processes to further maximize the harvesting of solar energy [141]. The schematic diagram of a multistage procedure, comprised of four or five different steps or approaches, is proposed in Figure 5. In this system, the feeding flows of this multistage process are solar lights, renewable biomass, and water, and outflows are produced hydrogen gas, oxygen gas, and processed biomasses that could be further converted into value-added organic fertilizer [142]. Within this multistage conversion process, the hydrogen production is initiated by photo-fermentation and photocatalysis (solar water-splitting) with feeding-water and organic substrates. Within the system, the cascade utilization of organic substrates could further maximize the hydrogen production in each individual processing step. Theoretically, it is possible to acquire a maximum hydrogen production rate of 12 moles of hydrogen from 1 mole of substrate (glucose) through this combined approach, using purple non-sulfur photosynthetic bacteria and anaerobic bacteria by integrating DF with the photo-fermentations [143]. In addition, the proposed process also integrates the photocatalysis and micro-electrolysis processes for the sake of maximizing hydrogen productions of the entire process. The challenges of this proposed integrated process lies in: (a) the pH swing between the steps of the photofermentation stage, where ammonia will be generated continuously during the photofermentation catalyzed by nitrogenase, and the nearly neutral pH value of DF; (b) how to best optimize the feeding concentration of organic substrates (C/N/O ratio) [144] and control different metabolic pathways on the level of genetic expressions, as this will significantly affect the level of genetic diversity expressions during the fermentations [145,146,147]; (c) eco-friendly access to the water available.
7. Conclusions
In this paper, biological hydrogen generation produced from renewable bio-resources was found to be a practical route for hydrogen production. Among different BHP routes, the DF has been found to be a practical approach in BHP, especially when it is enhanced by chemical addition. Among the different approaches of chemical addition to improve the activity of hydrogenase, the addition of NPs (Ni, Fe) was found to be relatively more efficient due to its direct effects of facilitating the electron transport between the ferredoxin and the hydrogenase. The order of effectiveness in enhancing the activities of hydrogenase on the basis of substrate conversion efficiency (YH2/S) and hydrogen evolution rate (HER) follows the order of metal NPs > metal monomers/metal ions/metal oxides > other metals (other than Ni, Fe). In order to make the BHP process more feasible and economical enough for industrial applications, future endeavors should focus on the optimized integration of different hydrogen production processes with the energy cascade utilization and material recycling and recovering. By appropriately integrating different approaches, it is potentially possible to approach the theoretical maximum hydrogen yield (12 mol H2 per 1 mol Glucose consumption). These novel approaches of process intensifications, and integration and appropriate combination of several hydrogen generation processes, such as photocatalysis, photofermentation, and DF processes, will eventually facilitate large-scale curbing of the emission footprint and the cost of BHP in the foreseeable future.
Funding
The authors would like to appreciate the funding support from funding support from National Key R&D Program of China (2018YFC1903500), Edith Cowan University for staff support grant and staff excellent awards grant. The Faculty Inspiration Grant of University of Nottingham and Qianjiang Talent Scheme-Grant/Award Number: QJD1803014 are also highly appreciated.
Acknowledgments
The Acid-Based coupled production group at Institute of Process Engineering Chinese Academy of Science is also highly appreciated. The critical and insightful comments raised from three anonymous reviewers in significantly improving the quality of this work were also highly appreciated.
Conflicts of Interest
All authors have no conflict of interest in this work.
References
- Haszeldine, R.S. Carbon Capture and Storage: How Green Can Black Be? Science 2009, 325, 1647–1652. [Google Scholar] [CrossRef] [PubMed]
- Sakimoto, K.K.; Wong, A.B.; Yang, P.D. Self-photosensitization of nonphotosynthetic bacteria for solar-to-chemical production. Science 2016, 351, 74–77. [Google Scholar] [CrossRef]
- Wargacki, A.J.; Leonard, E.; Win, M.N.; Regitsky, D.D.; Santos, C.N.S.; Kim, P.B.; Cooper, S.R.; Raisner, R.M.; Herman, A.; Sivitz, A.B.; et al. An Engineered Microbial Platform for Direct Biofuel Production from Brown Macroalgae. Science 2012, 335, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Al Sadat, W.I.; Archer, L.A. The O-2-assisted Al/CO2 electrochemical cell: A system for CO2 capture/conversion and electric power generation. Sci. Adv. 2016, 2. [Google Scholar] [CrossRef]
- Navlani-Garcia, M.; Mori, K.; Kuwahara, Y.; Yamashita, H. Recent strategies targeting efficient hydrogen production from chemical hydrogen storage materials over carbon-supported catalysts. NPG Asia Mater. 2018, 10, 277–292. [Google Scholar] [CrossRef]
- Benemann, J. Hydrogen biotechnology: Progress and prospects. Nat. Biotechnol. 1996, 14, 1101–1103. [Google Scholar] [CrossRef]
- Shown, I.; Hsu, H.C.; Chang, Y.C.; Lin, C.H.; Roy, P.K.; Ganguly, A.; Wang, C.H.; Chang, J.K.; Wu, C.I.; Chen, L.C.; et al. Highly Efficient Visible Light Photocatalytic Reduction of CO2 to Hydrocarbon Fuels by Cu-Nanoparticle Decorated Graphene Oxide. Nano Lett. 2014, 14, 6097–6103. [Google Scholar]
- Liu, J.; Liu, Y.; Liu, N.Y.; Han, Y.Z.; Zhang, X.; Huang, H.; Lifshitz, Y.; Lee, S.T.; Zhong, J.; Kang, Z.H. Metal-free efficient photocatalyst for stable visible water-splitting via a two-electron pathway. Science 2015, 347, 970–974. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, G.; Jia, Z.H.; Wen, C.; Zhang, L. Acid Hydrolysis of Corn Stover Using Hydrochloric Acid: Kinetic Modeling and Statistical Optimization. Chem. Ind. Chem. Eng. Q. 2014, 20, 531–539. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, J.P.; Yang, G.; Li, Z.H. Production of activated carbon by H3PO4 activation treatment of corncob and its performance in removing nitrobenzene from water. Environ. Prog. 2007, 26, 78–85. [Google Scholar] [CrossRef]
- Sun, Y.; Wei, J.; Yao, M.S.; Yang, G. Preparation of activated carbon from furfural production waste and its application for water pollutants removal and gas separation. Asia-Pac. J. Chem. Eng. 2012, 7, 547–554. [Google Scholar] [CrossRef]
- Bandyopadhyay, A.; Stöckel, J.; Min, H.; Sherman, L.A.; Pakrasi, H.B. High rates of photobiological H2 production by a cyanobacterium under aerobic conditions. Nat. Commun. 2010. [Google Scholar] [CrossRef]
- Han, W.; Yan, Y.T.; Shi, Y.W.; Gu, J.J.; Tang, J.H.; Zhao, H.T. Biohydrogen production from enzymatic hydrolysis of food waste in batch and continuous systems. Sci. Rep. 2016, 6, 38395. [Google Scholar] [CrossRef]
- Hwang, J.H.; Kim, H.C.; Choi, J.A.; Abou-Shanab, R.A.I.; Dempsey, B.A.; Regan, J.M.; Kim, J.R.; Song, H.; Nam, I.H.; Kim, S.N.; et al. Photoautotrophic hydrogen production by eukaryotic microalgae under aerobic conditions. Nat. Commun. 2014, 5, 3234. [Google Scholar] [CrossRef]
- Mishra, P.; Krishnan, S.; Rana, S.; Singh, L.; Sakinah, M.; Wahid, Z. Outlook of fermentative hydrogen production techniques: An overview of dark, photo and integrated dark-photo fermentative approach to biomass. Energy Strategy Rev. 2019, 24, 27–37. [Google Scholar] [CrossRef]
- Trchounian, K.; Sawers, R.G.; Trchounian, A. Improving biohydrogen productivity by microbial dark- and photo-fermentations: Novel data and future approaches. Renew. Sust. Energy Rev. 2017, 80, 1201–1216. [Google Scholar] [CrossRef]
- Gadhe, A.; Sonawane, S.S.; Varma, M.N. Enhanced biohydrogen production from dark fermentation of complex dairy wastewater by sonolysis. Int. J. Hydrog. Energy 2015, 40, 9942–9951. [Google Scholar] [CrossRef]
- Gadhe, A.; Sonawane, S.S.; Varma, M.N. Evaluation of ultrasonication as a treatment strategy for enhancement of biohydrogen production from complex distillery wastewater and process optimization. Int. J. Hydrog. Energy 2014, 39, 10041–10050. [Google Scholar] [CrossRef]
- Hassan, A.H.S.; Mietzel, T.; Brunstermann, R.; Schmuck, S.; Schoth, J.; Kuppers, M.; Widmann, R. Fermentative hydrogen production from low-value substrates. World J. Microb. Biot. 2018, 34, 176. [Google Scholar] [CrossRef]
- Yao, Z.T.; Su, W.P.; Wu, D.D.; Tang, J.H.; Wu, W.H.; Liu, J.; Han, W. A state-of-the-art review of biohydrogen producing from sewage sludge. Int. J. Energy Res. 2018, 42, 4301–4312. [Google Scholar] [CrossRef]
- Yang, J.W.G. Various additives for improving dark fermentative hydrogen production: A review. Renew. Sustain. Energy Rev. 2018, 95, 130–146. [Google Scholar]
- Das, D.; Veziroglu, T.N. Hydrogen production by biological processes: A survey of literature. Int. J. Hydrog. Energy 2001, 26, 13–28. [Google Scholar] [CrossRef]
- Seefeldt, L.C.; Peters, J.W.; Beratan, D.N.; Bothner, B.; Minteer, S.D.; Raugei, S.; Hoffman, B.M. Control of electron transfer in nitrogenase. Curr. Opin. Chem. Biol. 2018, 47, 54–59. [Google Scholar] [CrossRef]
- Hallenbeck, P.C.; Benemann, J.R. Biological hydrogen production; fundamentals and limiting processes. Int. J. Hydrog. Energy 2002, 27, 1185–1193. [Google Scholar] [CrossRef]
- Trofanchuk, O.; Stein, M.; Gessner, C.; Lendzian, F.; Higuchi, Y.; Lubitz, W. Single crystal EPR studies of the oxidized active site of [NiFe] hydrogenase from Desulfovibrio vulgaris Miyazaki F. J. Biol. Inorg. Chem. 2000, 5, 36–44. [Google Scholar] [CrossRef]
- Morra, S.; Arizzi, M.; Allegra, P.; la Licata, B.; Sagnelli, F.; Zitella, P.; Gilardi, G.; Valetti, F. Expression of different types of [FeFe]-hydrogenase genes in bacteria isolated from a population of a bio-hydrogen pilot-scale plant. Int. J. Hydrog. Energy 2014, 39, 9018–9027. [Google Scholar] [CrossRef]
- Peters, J.W.; Schut, G.J.; Boyd, E.S.; Mulder, D.W.; Shepard, E.M.; Broderick, J.B.; King, P.W.; Adams, M.W.W. [FeFe]- and [NiFe]-hydrogenase diversity, mechanism, and maturation. Bba-Mol. Cell Res. 2015, 1853, 1350–1369. [Google Scholar] [CrossRef]
- Grunwald, P. Biocatalysis: Biochemical Fundamentals and Applications; Wold Scientific Publishing Co. Pte. Ltd.: London, UK, 2018. [Google Scholar]
- Fontecilla-Camps, J.C.; Volbeda, A.; Cavazza, C.; Nicolet, Y. Structure/function relationships of [NiFe]- and [FeFe]-hydrogenases. Chem. Rev. 2007, 107, 4273–4303. [Google Scholar] [CrossRef]
- Volbeda, A.; Fontecilla-Camps, J.C. The active site and catalytic mechanism of NiFe hydrogenases. Dalton Trans. 2003, 4030–4038. [Google Scholar] [CrossRef]
- Kothari, R.; Singh, D.P.; Tyagi, V.V.; Tyagi, S.K. Fermentative hydrogen production—An alternative clean energy source. Renew. Sust. Energy Rev. 2012, 16, 2337–2346. [Google Scholar] [CrossRef]
- Elreedy, A.; Ibrahim, E.; Hassan, N.; El-Dissouky, A.; Fujii, M.; Yoshimura, C.; Tawfik, A. Nickel-graphene nanocomposite as a novel supplement for enhancement of biohydrogen production from industrial wastewater containing mono-ethylene glycol. Energy Convers. Manag. 2017, 140, 133–144. [Google Scholar] [CrossRef]
- Nagarajan, D.; Lee, D.J.; Kondo, A.; Chang, J.S. Recent insights into biohydrogen production by microalgae - From biophotolysis to dark fermentation. Bioresour. Technol. 2017, 227, 373–387. [Google Scholar] [CrossRef]
- Azwar, M.Y.; Hussain, M.A.; Abdul-Wahab, A.K. Development of biohydrogen production by photobiological, fermentation and electrochemical processes: A review. Renew. Sust. Energy Rev. 2014, 31, 158–173. [Google Scholar] [CrossRef]
- Happe, T.; Hemschemeier, A.; Winkler, M.; Kaminski, A. Hydrogenases in green algae: Do they save the algae’s life and solve our energy problems? Trends Plant Sci. 2002, 7, 246–250. [Google Scholar] [CrossRef]
- Eroglu, E.; Melis, A. Photobiological hydrogen production: Recent advances and state of the art. Bioresour. Technol. 2011, 102, 8403–8413. [Google Scholar] [CrossRef]
- Melis, A.; Neidhardt, J.; Benemann, J. Dunaliella salina (Chlorophyta) with small chlorophyll antenna sizes exhibit higher photosynthetic productivities and photon use efficiencies than normally pigmented cells. J. Appl. Phycol. 1998, 10, 515–525. [Google Scholar] [CrossRef]
- Williams, C.R.; Bees, M.A. Mechanistic modeling of sulfur-deprived photosynthesis and hydrogen production in suspensions of Chlamydomonas reinhardtii. Biotechnol. Bioeng. 2014, 111, 1–16. [Google Scholar] [CrossRef]
- Das, D.; Veziroglu, T.N. Advances in biological hydrogen production processes. Int. J. Hydrog. Energy 2008, 33, 6046–6057. [Google Scholar] [CrossRef]
- Liu, X.M.; Ren, N.Q.; Song, F.N.; Yang, C.P.; Wang, A.J. Recent advances in fermentative biohydrogen production. Prog. Nat. Sci. 2008, 18, 253–258. [Google Scholar] [CrossRef]
- Hallenbeck, P.C.; Ghosh, D. Advances in fermentative biohydrogen production: The way forward? Trends Biotechnol. 2009, 27, 287–297. [Google Scholar] [CrossRef]
- Cheng, H.L.J.; Zhang, J.; Ding, L.; Lin, R. Enhanced dark hydrogen fermentation of Enterobacter aerogenes/HoxEFUYH with carbon cloth. Int. J. Hydrog. Energy 2019, in press. [Google Scholar] [CrossRef]
- Kovacs, K.L.; Maroti, G.; Rakhely, G. A novel approach for biohydrogen production. Int. J. Hydrog. Energy 2006, 31, 1460–1468. [Google Scholar] [CrossRef]
- Sarma, S.J.; Brar, S.K.; le Bihan, Y.; Buelna, G.; Soccol, C.R. Mitigation of the inhibitory effect of soap by magnesium salt treatment of crude glycerol—A novel approach for enhanced biohydrogen production from the biodiesel industry waste. Bioresour. Technol. 2014, 151, 49–53. [Google Scholar] [CrossRef]
- Boboescu, I.Z.; Gherman, V.D.; Lakatos, G.; Pap, B.; Biro, T.; Maroti, G. Surpassing the current limitations of biohydrogen production systems: The case for a novel hybrid approach. Bioresour. Technol. 2016, 204, 192–201. [Google Scholar] [CrossRef]
- Basak, N.; Das, D. The prospect of purple non-sulfur (PNS) photosynthetic bacteria for hydrogen production: The present state of the art. World J. Microb. Biot. 2007, 23, 31–42. [Google Scholar] [CrossRef]
- Willey, J.M.; Sherwood, L.M.; Woolverton, C.J. Prescott’s Microbiology; McGraw Hill Education: New York, NY, USA, 2017. [Google Scholar]
- Ozturk, Y.; Yucel, M.; Daldal, F.; Mandaci, S.; Gunduz, U.; Turker, L.; Eroglu, I. Hydrogen production by using Rhodobacter capsulatus mutants with genetically modified electron transfer chains. Int. J. Hydrog. Energy 2006, 31, 1545–1552. [Google Scholar] [CrossRef]
- de la Cueva, S.C.; Guzman, C.L.A.; Hernandez, V.E.B.; Rodriguez, A.D. Optimization of biohydrogen production by the novel psychrophilic strain N92 collected from the Antarctica. Int. J. Hydrog. Energy 2018, 43, 13798–13809. [Google Scholar] [CrossRef]
- Kumari, S.; Das, D. Improvement of biohydrogen production using acidogenic culture. Int. J. Hydrog. Energy 2017, 42, 4083–4094. [Google Scholar] [CrossRef]
- Ergal, I.; Fuchs, W.; Hasibar, B.; Thallinger, B.; Bochmann, G.; Rittmann, S.K.M.R. The physiology and biotechnology of dark fermentative biohydrogen production. Biotechnol. Adv. 2018, 36, 2165–2186. [Google Scholar] [CrossRef]
- Buckel, W.; Thauer, R.K. Energy conservation via electron bifurcating ferredoxin reduction and proton/Na+ translocating ferredoxin oxidation. Bba-Bioenerg. 2013, 1827, 94–113. [Google Scholar] [CrossRef]
- Schafer, T.; Selig, M.; Schonheit, P. Acetyl-Coa Synthetase (Adp Forming) in Archaea, a Novel Enzyme Involved in Acetate Formation and Atp Synthesis. Arch. Microbiol. 1993, 159, 72–83. [Google Scholar] [CrossRef]
- Miura, Y. Hydrogen-Production by Biophotolysis Based on Microalgal Photosynthesis. Process. Biochem. 1995, 30, 1–7. [Google Scholar] [CrossRef]
- Show, K.Y.; Yan, Y.G.; Ling, M.; Ye, G.X.; Li, T.; Lee, D.J. Hydrogen production from algal biomass—Advances, challenges and prospects. Bioresour. Technol. 2018, 257, 290–300. [Google Scholar] [CrossRef]
- Holladay, J.D.; Hu, J.; King, D.L.; Wang, Y. An overview of hydrogen production technologies. Catal Today 2009, 139, 244–260. [Google Scholar] [CrossRef]
- Khetkorn, W.; Rastogi, R.P.; Incharoensakdi, A.; Lindblad, P.; Madamwar, D.; Pandey, A.; Larroche, C. Microalgal hydrogen production—A review. Bioresour. Technol. 2017, 243, 1194–1206. [Google Scholar] [CrossRef]
- Esper, B.; Badura, A.; Rogner, M. Photosynthesis as a power supply for (bio-)hydrogen production. Trends Plant Sci. 2006, 11, 543–549. [Google Scholar] [CrossRef]
- Aslam, M.; Ahmad, R.; Yasin, M.; Khan, A.L.; Shahid, M.K.; Hossain, S.; Khan, Z.; Jamil, F.; Rafiq, S.; Bilad, M.R.; et al. Anaerobic membrane bioreactors for biohydrogen production: Recent developments, challenges and perspectives. Bioresour. Technol. 2018, 269, 452–464. [Google Scholar] [CrossRef]
- Kapdan, I.K.; Kargi, F. Bio-hydrogen production from waste materials. Enzym. Microb. Technol. 2006, 38, 569–582. [Google Scholar] [CrossRef]
- Laurinavichene, T.V.; Kosourov, S.N.; Ghirardi, M.L.; Seibert, M.; Tsygankov, A.A. Prolongation of H(2) photoproduction by immobilized, sulfur-limited Chlamydomonas reinhardtii cultures. J. Biotechnol. 2008, 134, 275–277. [Google Scholar] [CrossRef]
- Abo-Hashesh, M.; Desaunay, N.; Hallenbeck, P.C. High yield single stage conversion of glucose to hydrogen by photofermentation with continuous cultures of Rhodobacter capsulatus JP91. Bioresour. Technol. 2013, 128, 513–517. [Google Scholar] [CrossRef]
- Xia, A.; Cheng, J.; Ding, L.K.; Lin, R.C.; Huang, R.; Zhou, J.H.; Cen, K.F. Improvement of the energy conversion efficiency of Chlorella pyrenoidosa biomass by a three-stage process comprising dark fermentation, photofermentation, and methanogenesis. Bioresour. Technol. 2013, 146, 436–443. [Google Scholar] [CrossRef]
- Larimer, F.W.; Chain, P.; Hauser, L.; Lamerdin, J.; Malfatti, S.; Do, L.; Land, M.L.; Pelletier, D.A.; Beatty, J.T.; Lang, A.S.; et al. Complete genome sequence of the metabolically versatile photosynthetic bacterium Rhodopseudomonas palustris. Nat. Biotechnol. 2004, 22, 55–61. [Google Scholar] [CrossRef]
- Machado, R.G.; Moreira, F.S.; Batista, F.R.X.; Ferreira, J.S.; Cardoso, V.L. Repeated batch cycles as an alternative for hydrogen production by co-culture photofermentation. Energy 2018, 153, 861–869. [Google Scholar] [CrossRef]
- Kars, G.; Gunduz, U.; Yucel, M.; Rakhely, G.; Kovacs, K.L.; Eroglu, I. Evaluation of hydrogen production by Rhodobacter sphaeroides OU001 and its hupSL deficient mutant using acetate and malate as carbon sources. Int. J. Hydrog. Energy 2009, 34, 2184–2190. [Google Scholar] [CrossRef]
- Ren, N.Q.; Liu, B.F.; Ding, J.; Xie, G.J. Hydrogen production with R. faecalis RLD-53 isolated from freshwater pond sludge. Bioresour. Technol. 2009, 100, 484–487. [Google Scholar] [CrossRef]
- Budiman, P.M.; Wu, T.Y. Role of chemicals addition in affecting biohydrogen production through photofermentation. Energy Convers. Manag. 2018, 165, 509–527. [Google Scholar] [CrossRef]
- Keskin, T.; Hallenbeck, P.C. Hydrogen production from sugar industry wastes using single-stage photofermentation. Bioresour. Technol. 2012, 112, 131–136. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, G.; Zhang, J.P.; Wen, C.; Sun, Z. Optimization and kinetic modeling of an enhanced bio-hydrogen fermentation with the addition of synergistic biochar and nickel nanoparticle. Int. J. Energy Res. 2019, 43, 983–999. [Google Scholar] [CrossRef]
- Kim, M.S.; Fitriana, H.N.; Kim, T.W.; Kang, S.G.; Jeon, S.G.; Chung, S.H.; Park, G.W.; Na, J.G. Enhancement of the hydrogen productivity in microbial water gas shift reaction by Thermococcus onnurineus NA1 using a pressurized bioreactor. Int. J. Hydrog. Energy 2017, 42, 27593–27599. [Google Scholar] [CrossRef]
- Lee, J.; Jung, N.; Shin, J.H.; Park, L.H.; Sung, Y.E.; Park, T.H. Enhancement of hydrogen production and power density in a bio-reformed formic acid fuel cell (BrFAFC) using genetically modified Enterobacter asburiae SNU-1. Int. J. Hydrog. Energy 2014, 39, 11731–11737. [Google Scholar] [CrossRef]
- Zagrodnik, R.; Laniecki, M. Hydrogen production from starch by co-culture of Clostridium acetobutylicum and Rhodobacter sphaeroides in one step hybrid dark-and photofermentation in repeated fed-batch reactor. Bioresour. Technol. 2017, 224, 298–306. [Google Scholar] [CrossRef]
- Shen, N.; Dai, K.; Xia, X.Y.; Zeng, R.J.; Zhang, F. Conversion of syngas (CO and H-2) to biochemicals by mixed culture fermentation in mesophilic and thermophilic hollow-fiber membrane biofilm reactors. J. Clean. Prod. 2018, 202, 536–542. [Google Scholar] [CrossRef]
- Hwang, P.S.Y.; Lee, M.-K.; Yun, Y.-M.; Kim, D.-H. Enhanced hydrogen fermentation by zero valent iron addition. Int. J. Hydrog. Energy 2019, 44, 3387–3394. [Google Scholar] [CrossRef]
- Pugazhendhi, S.S.A.; Nguyen, D.D.; Banu, J.R.; Kumar, G. Application of nanotechnology (nanoparticles) in dark fermentative hydrogen production. Int. J. Hydrog. Energy 2019, 44, 1431–1440. [Google Scholar] [CrossRef]
- Ren, H.Y.; Kong, F.Y.; Ma, J.; Zhao, L.; Xie, G.J.; Xing, D.F.; Guo, W.Q.; Liu, B.F.; Ren, N.Q. Continuous energy recovery and nutrients removal from molasses wastewater by synergistic system of dark fermentation and algal culture under various fermentation types. Bioresour. Technol. 2018, 252, 110–117. [Google Scholar] [CrossRef]
- Khan, S.Y.Z.; Ahmad, M.M.; Chok, V.S.; Uemura, Y.; Sabil, K.M. Review on Hydrogen Production Technologies in Malaysia. Int. J. Eng. Technol. 2010, 10, 1–8. [Google Scholar]
- Sun, Y.; Zhang, J.P.; Yang, G.; Li, Z.H. Analysis of trace elements in corncob by microwave Digestion-ICP-AES. Spectrosc. Spect. Anal. 2007, 27, 1424–1427. [Google Scholar]
- Taherdanak, M.; Zilouei, H.; Karimi, K. The effects of Fe-0 and Ni-0 nanoparticles versus Fe2+ and Ni2+ ions on dark hydrogen fermentation. Int. J. Hydrog. Energy 2016, 41, 167–173. [Google Scholar] [CrossRef]
- Rittmann, S.; Herwig, C. A comprehensive and quantitative review of dark fermentative biohydrogen production. Microb. Cell Fact. 2012, 11, 115. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Shen, J.Q. Enhancement effect of gold nanoparticles on biohydrogen production from artificial wastewater. Int. J. Hydrog. Energy 2007, 32, 17–23. [Google Scholar] [CrossRef]
- Patel, S.K.S.; Kalia, V.C.; Choi, J.H.; Haw, J.R.; Kim, I.W.; Lee, J.K. Immobilization of Laccase on SiO2 Nanocarriers Improves Its Stability and Reusability. J. Microbiol. Biotechnol. 2014, 24, 639–647. [Google Scholar] [CrossRef]
- Camacho, C.E.; Romano, F.I.; Ruggeri, B. Macro approach analysis of dark biohydrogen production in the presence of zero valent powered Fe degrees. Energy 2018, 159, 525–533. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, L.X.; Li, D.P. Enhanced dark fermentative hydrogen production by zero-valent iron activated carbon micro-electrolysis. Int. J. Hydrog. Energy 2015, 40, 12201–12208. [Google Scholar] [CrossRef]
- Yu, L.; Yu, Y.; Jiang, W.T.; Wei, H.Z.; Sun, C.L. Integrated treatment of municipal sewage sludge by deep dewatering and anaerobic fermentation for biohydrogen production. Environ. Sci. Pollut. Res. 2015, 22, 2599–2609. [Google Scholar] [CrossRef]
- Yu, L.; Jiang, W.T.; Yu, Y.; Sun, C.L. Effects of dilution ratio and Fe-0 dosing on biohydrogen production from dewatered sludge by hydrothermal pretreatment. Environ. Technol. 2014, 35, 3092–3104. [Google Scholar] [CrossRef]
- Yang, G.; Wang, J.L. Improving mechanisms of biohydrogen production from grass using zero-valent iron nanoparticles. Bioresour. Technol. 2018, 266, 413–420. [Google Scholar] [CrossRef]
- Khan, M.M.; Lee, J.; Cho, M.H. Electrochemically active biofilm mediated bio-hydrogen production catalyzed by positively charged gold nanoparticles. Int. J. Hydrog. Energy 2013, 38, 5243–5250. [Google Scholar] [CrossRef]
- Reddy, K.; Nasr, M.; Kumari, S.; Kumar, S.; Gupta, S.K.; Enitan, A.M.; Bux, F. Biohydrogen production from sugarcane bagasse hydrolysate: Effects of pH, S/X, Fe2+, and magnetite nanoparticles. Environ. Sci. Pollut. Res. 2017, 24, 8790–8804. [Google Scholar] [CrossRef]
- Zhang, X.K.; Tabita, F.R.; Vanbaalen, C. Nickel Control of Hydrogen-Production and Uptake in Anabaena Spp Strain-Ca and Strain-1f. J. Gen. Microbiol. 1984, 130, 1815–1818. [Google Scholar]
- Zhang, X.K.; Haskell, J.B.; Tabita, F.R.; Vanbaalen, C. Aerobic Hydrogen-Production by the Heterocystous Cyanobacteria Anabaena Spp Strain-Ca and Strain-1f. J. Bacteriol. 1983, 156, 1118–1122. [Google Scholar]
- Alshiyab, H.; Kalil, M.S.; Hamid, A.A.; Yusoff, W.M.W. Trace Metal Effect on Hydrogen Production Using C.acetobutylicum. J. Biol. Sci. 2008, 8, 1–9. [Google Scholar] [CrossRef]
- Lee, Y.J.; Miyahara, T.; Noike, T. Effect of iron concentration on hydrogen fermentation. Bioresour. Technol. 2001, 80, 227–231. [Google Scholar] [CrossRef]
- Taikhao, S.; Phunpruch, S. Effect of Metal Cofactors of Key Enzymes on Biohydrogen Production by Nitrogen Fixing Cyanobacterium Anabaena siamensis TISIR 8012. In Proceedings of the 2017 International Conference on Alternative Energy in Developing Countries and Emerging Economies, Thailand, Bangkok, 25–26 May 2017; pp. 360–365. [Google Scholar]
- Wang, J.L.; Wan, W. Effect of Fe2+ concentration on fermentative hydrogen production by mixed cultures. Int. J. Hydrog. Energy 2008, 33, 1215–1220. [Google Scholar] [CrossRef]
- Chong, M.L.; Rahman, N.A.; Yee, P.L.; Aziz, S.A.; Rahim, R.A.; Shirai, Y.; Hassan, M.A. Effects of pH, glucose and iron sulfate concentration on the yield of biohydrogen by Clostridium butyricum EB6. Int. J. Hydrog. Energy 2009, 34, 8859–8865. [Google Scholar] [CrossRef]
- Vi, L.V.T.; Salakkam, A.; Reungsang, A. Optimization of key factors affecting bio-hydrogen production from sweet potato starch. Energy Procedia 2017, 138, 973–978. [Google Scholar]
- Mullai, P.; Rene, E.R.; Sridevi1, K. Biohydrogen Production and Kinetic Modeling Using Sediment Microorganisms of Pichavaram Mangroves, India. BioMed Res. Int. 2013, 1, 1–9. [Google Scholar] [CrossRef]
- Dhar, B.R.; Elbeshbishy, E.; Nakhla, G. Influence of iron on sulfide inhibition in dark biohydrogen fermentation. Bioresour. Technol. 2012, 126, 123–130. [Google Scholar] [CrossRef]
- Karthic, P.; Joseph, S.; Arun, N. Optimization of Process Variables for Biohydrogen Production from Glucose by Enterobacter aerogenes. Open Access Sci. Rep. 2012, 1, 1–6. [Google Scholar]
- Gou, C.Y.; Guo, J.B.; Lian, J.; Guo, Y.K.; Jiang, Z.S.; Yue, L.; Yang, J.L. Characteristics and kinetics of biohydrogen production with Ni2+ using hydrogen-producing bacteria. Int. J. Hydrog. Energy 2015, 40, 161–167. [Google Scholar] [CrossRef]
- Wang, J.; Wan, W. Influence of Ni2+ concentration on biohydrogen production. Bioresour. Technol. 2008, 99, 8864–8868. [Google Scholar] [CrossRef]
- Srikanth, S.; Mohan, S.V. Regulatory function of divalent cations in controlling the acidogenic biohydrogen production process. RSC Adv. 2012, 2, 6576–6589. [Google Scholar] [CrossRef]
- Calli, B.; Boënne, W.; Vanbroekhoven, K. Bio-Hydrogen Potential of Easily Biodegradable Substrate Through Dark Fermentation; WHEC 16: Lyon, France, 2006. [Google Scholar]
- Xiaolong, H.; Minghua, Z.; Hanqing, Y.; Qinqin, S.; Lecheng, L. Effect of Sodium Ion Concentration on Hydrogen Production from Sucrose by Anaerobic Hydrogen-producing Granular Sludge. Chin. J. Chem. Eng. 2006, 14, 511–517. [Google Scholar]
- Alshiyab, H.; Kalil, M.S.; Hamid, A.A.; Yusoff, W.M.W. Effects of salt addition on hydrogen production by C.acetobutylicum. Pak. J. Biol. Sci. 2008, 11, 2193–2200. [Google Scholar] [CrossRef]
- HongLe, D.T.; Nitisoravut, R. Ni-Mg-Al Hydrotalcite for Improvement of Dark Fermentative Hydrogen Production. Energy Procedia 2015, 79, 301–306. [Google Scholar]
- Karadag, D.; Puhakka, J.A. Enhancement of anaerobic hydrogen production by iron and nickel. Int. J. Hydrog. Energy 2010, 35, 8554–8560. [Google Scholar] [CrossRef]
- Sekoai, P.T.; Ouma, C.N.M.; Preez, S.P.D.; Modisha, P.; Engelbrecht, N.; Bessarabov, D.G.; Ghimire, A. Application of nanoparticles in biofuels: An overview. Fuel 2019, 237, 380–397. [Google Scholar] [CrossRef]
- Patel, S.K.S.; Lee, J.K.; Kalia, V.C. Nanoparticles in Biological Hydrogen Production: An Overview. Indian J. Microbiol. 2018, 58, 8–18. [Google Scholar] [CrossRef]
- Zhao, Y.X.; Chen, Y.G. Nano-TiO2 Enhanced Photofermentative Hydrogen Produced from the Dark Fermentation Liquid of Waste Activated Sludge. Environ. Sci. Technol. 2011, 45, 8589–8595. [Google Scholar] [CrossRef]
- Mishra, P.; Thakur, S.; Mahapatra, D.M.; Wahid, Z.A.; Liu, H.; Singh, L. Impacts of nano-metal oxides on hydrogen production in anaerobic digestion of palm oil mill effluent—A novel approach. Int. J. Hydrog. Energy 2018, 43, 2666–2676. [Google Scholar] [CrossRef]
- Pandey, A.; Gupta, K.; Pandey, A. Effect of nanosized TiO2 on photofermentation by Rhodobacter sphaeroides NMBL-02. Biomass Bioenerg. 2015, 72, 273–279. [Google Scholar] [CrossRef]
- Nasr, M.; Tawfik, A.; Ookawara, S.; Suzuki, M.; Kumari, S.; Bux, F. Continuous biohydrogen production from starch wastewater via sequential dark-photo fermentation with emphasize on maghemite nanoparticles. J. Ind. Eng. Chem. 2015, 21, 500–506. [Google Scholar] [CrossRef]
- Han, H.L.; Cui, M.J.; Wei, L.L.; Yang, H.J.; Shen, J.Q. Enhancement effect of hematite nanoparticles on fermentative hydrogen production. Bioresour. Technol. 2011, 102, 7903–7909. [Google Scholar] [CrossRef]
- Mohanraj, S.; Kodhaiyolii, S.; Rengasamy, M.; Pugalenthi, V. Green Synthesized Iron Oxide Nanoparticles Effect on Fermentative Hydrogen Production by Clostridium acetobutylicum. Appl. Biochem. Biotechnol. 2014, 173, 318–331. [Google Scholar] [CrossRef]
- Gadhe, A.; Sonawane, S.S.; Varma, M.N. Enhancement effect of hematite and nickel nanoparticles on biohydrogen production from dairy wastewater. Int J. Hydrogen Energ 2015, 40, 4502–4511. [Google Scholar] [CrossRef]
- Engliman, N.S.; Abdul, P.M.; Wu, S.Y.; Jahim, J.M. Influence of iron (II) oxide nanoparticle on biohydrogen production in thermophilic mixed fermentation. Int. J. Hydrog. Energy 2017, 42, 27482–27493. [Google Scholar] [CrossRef]
- Gadhe, A.; Sonawane, S.S.; Varma, M.N. Influence of nickel and hematite nanoparticle powder on the production of biohydrogen from complex distillery wastewater in batch fermentation. Int. J. Hydrog. Energy 2015, 40, 10734–10743. [Google Scholar] [CrossRef]
- Sivagurunathan, P.; Pugazhendhi, A.; Kumar, G.; Park, J.H.; Kim, S.H. Biohydrogen fermentation of galactose at various substrate concentrations in an immobilized system and its microbial correspondence. J. Biosci. Bioeng. 2018, 125, 559–564. [Google Scholar] [CrossRef]
- Tanisho, S.; Ishiwata, Y. Continuous hydrogen production from molasses by fermentation using urethane foam as a support of flocks. Int. J. Hydrog. Energy 1995, 20, 541–545. [Google Scholar] [CrossRef]
- Muri, P.; Marinsek-Logar, R.; Djinovic, P.; Pintar, A. Influence of support materials on continuous hydrogen production in anaerobic packed-bed reactor with immobilized hydrogen producing bacteria at acidic conditions. Enzym. Microb. Technol. 2018, 111, 87–96. [Google Scholar] [CrossRef]
- Chookaew, T.; O-Thong, S.; Prasertsan, P. Biohydrogen production from crude glycerol by immobilized Klebsiella sp TR17 in a UASB reactor and bacterial quantification under non-sterile conditions. Int. J. Hydrog. Energy 2014, 39, 9580–9587. [Google Scholar] [CrossRef]
- Yokoi, H.; Maki, R.; Hirose, J.; Hayashi, S. Microbial production of hydrogen from starch-manufacturing wastes. Biomass Bioenerg 2002, 22, 389–395. [Google Scholar] [CrossRef]
- Jamali, N.S.; Jamaliah, M.J.; Isahak, W.N.R.W. Biofilm formation on granular activated carbon in xylose and glucose mixture for thermophilic biohydrogen production. Int. J. Hydrog. Energy 2016, 41, 21617–21627. [Google Scholar] [CrossRef]
- Sunyoto, N.; Zhu, M.; Zhang, Z.; Zhang, D. Effect of biochar addition and initial pH on hydrogen production from the first phase of two-phase anaerobic digestion of carbohydrates food waste. Energy Procedia 2017, 105, 379–384. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J.; Zang, L. Thermophilic bio-hydrogen production from corn-bran residue pretreated by calcined-lime mud from papermaking process. Bioresour. Technol. 2015, 198, 564–570. [Google Scholar] [CrossRef]
- Sekoai, P.T.; Daramola, M.O. Effect of metal ions on dark fermentative biohydrogen production using suspended and immobilized cells of mixed bacteria. Chem. Eng. Commun. 2018, 205, 1011–1022. [Google Scholar] [CrossRef]
- Satar, I.; Ghasemi, M.; Aljlil, S.A.; Isahak, W.N.R.W.; Abdalla, M.; Alam, J.; Daud, W.R.W.; Yarmo, M.A.; Akbarzadeh, O. Production of hydrogen by Enterobacter aerogenes in an immobilized cell reactor. Int. J. Hydrog. Energy 2017, 42, 9024–9030. [Google Scholar] [CrossRef]
- Nakatani, H.; Ding, N.; Ohara, Y.; Hori, K. Immobilization of Enterobacter aerogenes by a Trimeric Autotransporter Adhesin, AtaA, and Its Application to Biohydrogen Production. Catalysts 2018, 8, 159. [Google Scholar] [CrossRef]
- Palazzi, E.; Fabiano, B.; Perego, P. Process development of continuous hydrogen production by Enterobacter aerogenes in a packed column reactor. Bioprocess. Eng. 2000, 22, 205–213. [Google Scholar] [CrossRef]
- Zhang, J.S.; Fan, C.F.; Zang, L.H. Improvement of hydrogen production from glucose by ferrous iron and biochar. Bioresour. Technol. 2017, 245, 98–105. [Google Scholar] [CrossRef]
- Rami, H.Ö.; El-Emam, S. Comprehensive Review on the Techno-Economics of Sustainable Large-Scale Clean Hydrogen Production. J. Clean. Prod. 2019, in press. [Google Scholar]
- Sun, Y.; Yao, M.S.; Zhang, J.P.; Yang, G. Indirect CO2 mineral sequestration by steelmaking slag with NH4Cl as leaching solution. Chem. Eng. J. 2011, 173, 437–445. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, J.P.; Zhang, L. NH4Cl selective leaching of basic oxygen furnace slag: Optimization study using response surface methodology. Environ. Prog. Sustain. 2016, 35, 1387–1394. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, G.; Wen, C.; Zhang, L.; Sun, Z. Artificial neural networks with response surface methodology for optimization of selective CO2 hydrogenation using K-promoted iron catalyst in a microchannel reactor. J. CO2 Util. 2018, 23, 10–21. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, G.; Zhang, L.; Sun, Z. Fischer-Tropsch synthesis in a microchannel reactor using mesoporous silica supported bimetallic Co-Ni catalyst: Process optimization and kinetic modeling. Chem. Eng. Process. 2017, 119, 44–61. [Google Scholar] [CrossRef]
- Sun, Y.; Lin, Z.; Peng, S.H.; Sage, V.; Sun, Z. A Critical Perspective on CO2 Conversions into Chemicals and Fuels. J. Nanosci. Nanotechnol. 2019, 19, 1–13. [Google Scholar] [CrossRef]
- Schmidt, O.; Gambhir, A.; Staffell, I.; Hawkes, A.; Nelson, J.; Few, S. Future cost and performance of water electrolysis: An expert elicitation study. Int. J. Hydrog. Energy 2017, 42, 30470–30492. [Google Scholar] [CrossRef]
- Trchounian, A. Mechanisms for hydrogen production by different bacteria during mixed-acid and photo-fermentation and perspectives of hydrogen production biotechnology. Crit. Rev. Biotechnol. 2015, 35, 103–113. [Google Scholar] [CrossRef]
- Arizzi, M.; Morra, S.; Pugliese, M.; Guilin, M.L.; Gilardi, G.; Valetti, F. Biohydrogen and biomethane production sustained by untreated matrices and alternative application of compost waste. Waste Manag. 2016, 56, 151–157. [Google Scholar] [CrossRef]
- Hawkes, F.R.; Hussy, I.; Kyazze, G.; Dinsdale, R.; Hawkes, D.L. Continuous dark fermentative hydrogen production by mesophilic microflora: Principles and progress. Int. J. Hydrog. Energy 2007, 32, 172–184. [Google Scholar] [CrossRef]
- Hamilton, C.; Hiligsmann, S.; Calusinska, M.; Beckers, L.; Masset, J.; Wilmotte, A.; Thonart, P. Investigation of hydrogenase molecular marker to optimize hydrogen production from organic wastes and effluents of agro-food industries. Biotechnol. Agron. Soc. Environ. 2010, 14, 574–575. [Google Scholar]
- Quéméneur, M.; Hamelin, J.; Marie-Thérèse, S.B.; EricLatrille, G.-O.; EricTrably, J.-P. Changes in hydrogenase genetic diversity and proteomic patterns in mixed-culture dark fermentation of mono-, di- and tri-saccharides. Int. J. Hydrog. Energy 2011, 36, 11654–11665. [Google Scholar] [CrossRef]
- Hamilton, C.; Calusinska, M.; Baptiste, S.; Masset, J.; Beckers, L.; Thonart, P.; Hiligsmann, S. Effect of the nitrogen source on the hydrogen production metabolism and hydrogenases of Clostridium butyricum CWBI1009. Int. J. Hydrog. Energy 2018, 43, 5451–5462. [Google Scholar] [CrossRef]
- Masset, J.; Calusinska, M.; Hamilton, C.; Hiligsmann, S.; Joris, B.; Wilmotte, A.; Thonart, P. Fermentative hydrogen production from glucose and starch using pure strains and artificial co-cultures of Clostridium spp. Biotechnol. Biofuels 2012, 5, 35. [Google Scholar] [CrossRef]
Figure 1.
Schematic structure of nitrogenase, where R and R* are the ligands. The figure was rearranged from Seefeldt et al. [23].
Figure 1.
Schematic structure of nitrogenase, where R and R* are the ligands. The figure was rearranged from Seefeldt et al. [23].

Figure 2.
Schematic structure of different types of hydrogenase: (a) [NiFe] hydrogenase; (b) [FeFe] hydrogenase; and (c) [Fe] hydrogenase. The figure was adapted and rearranged from Fontecilla-Camps et al. [29].
Figure 2.
Schematic structure of different types of hydrogenase: (a) [NiFe] hydrogenase; (b) [FeFe] hydrogenase; and (c) [Fe] hydrogenase. The figure was adapted and rearranged from Fontecilla-Camps et al. [29].

Figure 3.
Conceptual illustration of the bio-hydrogen generation pathways: (a) biophotolysis, (b) photofermentation. (c) DF (dark fermentation), PSI represents photosynthesis system 1, PSII is photosynthesis system 2, Fdox is the oxidized Ferredoxin, and Fdred is the reduced Ferredoxin.
Figure 3.
Conceptual illustration of the bio-hydrogen generation pathways: (a) biophotolysis, (b) photofermentation. (c) DF (dark fermentation), PSI represents photosynthesis system 1, PSII is photosynthesis system 2, Fdox is the oxidized Ferredoxin, and Fdred is the reduced Ferredoxin.

Figure 4.
Data illustrating the mean and standard deviation of different metal additions, such as NPs, mental monomers, metal ions, metal oxides, and other metals on BHP during DF: (a) HER as a function of YH2/S; (b) qH2 as a function of YH2/S. Note: Others exclude reports using immobilized supporters.
Figure 4.
Data illustrating the mean and standard deviation of different metal additions, such as NPs, mental monomers, metal ions, metal oxides, and other metals on BHP during DF: (a) HER as a function of YH2/S; (b) qH2 as a function of YH2/S. Note: Others exclude reports using immobilized supporters.

Figure 5.
Simplified conceptual illustration diagram of an integrated and multi-coupling process of BHP (biohydrogen production) with photocatalysis using biomass as the substrate.
Figure 5.
Simplified conceptual illustration diagram of an integrated and multi-coupling process of BHP (biohydrogen production) with photocatalysis using biomass as the substrate.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of bio-hydrogen pathways catalyzed by hydrogenase.
| Pathways | Challenges | Microbes Strains | Hydrogen Enhancement |
|---|---|---|---|
| BP |
| ||
| PF | PNS bacteria, such as Rhodopseudomonas genus [64] | Process optimization, such as: | |
| DF |
|
Table 2.
Summary and comparisons of bio-hydrogen production with addition of metal monomers and nanoparticles.
Table 2.
Summary and comparisons of bio-hydrogen production with addition of metal monomers and nanoparticles.
| Metal | Conc/mg L−1 | Feed | Organism | Process | Temp/°C | Yielda/ | Reference |
|---|---|---|---|---|---|---|---|
| Au (NPs) | 10 nM | Sucrose | MC | Batch | 35 | 4.47 (+61.7%) | [80] |
| Ag (NPs) | 20 nM | Glucose | MC | Batch | 37 | 2.48 (+67.6%) | [81] |
| Ni0 | 2.5 | Glucose | AS | Batch | 37 | 57 c (+79.8%) | [82] |
| Ni (NPs) | 5.7 | Glucose | AS | Batch | 33 | 2.54 (+22.7%) | [76] |
| Ni (NPs) | 60 | Wastewater | AS | Batch | 55 | 24.7 b (+22%) | [32] |
| Fe0 | 2000 | OWM | AS | Batch | 30 | 102 b (+46%) | [83] |
| Fe0 | 400 | Sucrose | AS | Batch | 30 | 1.2 (+37%) | [84] |
| Fe0 | 550 | Sludge | AS | CSTR | 37 | 650 d (+10%) | [85] |
| Fe0 | 100 | DS | AS | Batch | 37 | 26 c (+16%) | [86] |
| Fe (NPs) | 400 | Grass | CB | Batch | 37 | 65 c (+44%) | [87] |
| Cu (NPs) | 2.5 | Hexose | CA | Batch | 30 | 1.7 (−3.5%) | [88] |
| Fe (NPs) | 200 | SJ | AS | Batch | 30 | 1.15 (+62%) | [89] |
| Ni + Fe (NPs) | 37.5 + 37.5 | Starch | AS | Batch | 37 | 250 b (110%) | [82] |
| Ni (NPs) | 35 | Glucose | CB | Batch | 35 | 212 b (+32%) | [70] |
| Ni (NPs) + BC | 35 | Glucose | CB | Batch | 35 | 238 b (+49%) | [70] |
OMW: organic market waste: DS: dewatered sludge; SJ: sugarcane juice. MC: mixed consortia; AS: Anaerobic sludge; CB: Clostridium. Butyricum; CA: Clostridium acetobutylicum; a mol/mol of hexose; b L/kg TSS or COD or VSS (TSS: total suspended solids, COD chemical oxygen demand, VSS volatile suspended solids); c mL/g-dry grass; d ml/L.d.
Table 3.
Comparisons of bio-hydrogen production with addition of nickel and other metal nanoparticles.
Table 3.
Comparisons of bio-hydrogen production with addition of nickel and other metal nanoparticles.
| Metal Ion | Opt/mg L−1 | Organism | Feed | Process | Temp/°C | Yielda | Reference |
|---|---|---|---|---|---|---|---|
| FeCl3 | 60 uM | Cyanobacteria | BG | Batch | 30 | 0.06 b (+25%) | [95] |
| FeSO4 | 300 | AS | Glucose | Batch | 35 | 302 (+56%) | [96] |
| FeSO4 | 300 | CB | Glucose | Batch | 37 | 2.4 c (+30%) | [97] |
| FeSO4 | 25 | CB | Glucose | Batch | 30 | 408 (+4.3%) | [93] |
| FeSO4 | 63 | AS | PS | Batch | 30 | 226 (52%) | [98] |
| FeSO4 | 100 | HTS | Glucose | Batch | 35 | 2.6 c (+13%) | [99] |
| FeCl2 | 353 | AS | Sucrose | Batch | 37 | 132 (+650%) | [94] |
| FeCl2 | 50 | AS | Glucose | Batch | 37 | 216 (+23.4%) | [100] |
| FeCl3 | 213 | EA | Glucose | Batch | 30 | 1.7 c (+55%) | [101] |
| Ni2+ | 0.6 | CD | Sucrose | Batch | 35 | 2.1 c (+107%) | [102] |
| Ni2+ | 0.2 | MC | Glucose | Batch | 35 | 2.4 c (+75%) | [103] |
| NiCl2 | 0.1 | AS | Glucose | Batch | 35 | 289 (+55%) | [103] |
| NiCl2 | 16 | AS | SW | Batch | 34 | 1120 (+500%) | [104] |
| MgCl2 | 200 | MC | Glucose | FB | 35 | 1.75 (+600%) | [105] |
| Na2CO3 | 2000 | AS | Sucrose | UASB | 37 | 40 (+300%) | [106] |
| MgCl2 | 500 | CB | Glucose | Batch | 30 | 209 (−30%) | [93] |
| CaCl2 | 500 | CB | Glucose | Batch | 30 | 82 (−72%) | [93] |
| NaCl | 5000 | CA | Glucose | Batch | 30 | 2.7 c (−29%) | [107] |
| Hydrotalcite | 250 | HTS | Sucrose | UASB | 37 | 3.4 c (+80%) | [108] |
| Ni-Fe | 50 + 25 | CB | Glucose | ACSTR | 30 | 300 (+70%) | [109] |
Hydrotalcite: Ni-Mg-Al; AS: anaerobic sludge: CB: Clostridium. Butyricum; HTS: heat treated sludge; EA: Enterobacter aerogenes; MC: mixed consortia; CA: Clostridium acetobutylicum; CD: Cow dung; BG: BG110 media; PS: potato starch; SW: synthetic wastewater. FB: fed-batch; UASB: upflow anaerobic blanket; ACSTR: anaerobic continuous stirred tank reactor. a ml; b μmol.mg-1 h−1; c mol.mol−1; d imL L−1 h−1.
Table 4.
Impact of metal oxide upon the activity of nickel-contained hydrogenase for hydrogen production.
Table 4.
Impact of metal oxide upon the activity of nickel-contained hydrogenase for hydrogen production.
| Metal Ion | Opt/mg L−1 | Organism | Feed | Process | Temp/°C | Yielda | Reference |
|---|---|---|---|---|---|---|---|
| TiO2 | 100 | BA | Glucose | Batch | 30 | 160 (+46%) | [112] |
| CoO (NPs) | 1 | AS | POME | Batch | 37 | 0.5 b (+10%) | [113] |
| TiO2 | 300 | RS | SM | Batch | 32 | 1900 (+54%) | [114] |
| γ-Fe2O3 | 25 | AS | SB | Batch | 30 | 0.9 c (+62%) | [115] |
| α-Fe2O3 | 63 | MC | Inorganic salt | Batch | 30 | 3.6 c (33%) | [116] |
| Fe2O3 | 175 | CA | CL | Batch | 37 | 2.3 c (+18%) | [117] |
| NiO (NPs) | 10 | MC | CDW | Batch | 37 | 13 b (+33%) | [118] |
| NiO (NPs) | 1.5 | BA | POME | Batch | 37 | 25 b (+15%) | [113] |
| NiO (NPs) | 100 | AS | GS | Batch | 35 | 2.1 c (+107%) | [119] |
| Fe2O3/NiO | 50/10 | MC | CDW | Batch | 37 | 17 b (+45%) | [118] |
| Fe2O3/NiO (NPs) | 200/5 | AS | DW | Batch | 37 | 19 b (+25%) | [120] |
BA: Bacillus anthracis; AS: anaerobic sludge: RS: Rhodobacter sphaeroides; MC: mixed consortia; CA: Clostridium acetobutylicum. POME: Palm oil mill effluent; SM: Sistrom’s medium; SB: Sugarcane bagasse; CL: curry leaf; CDW: complex dairy wastewater; GS: glucose and starch, DW: distillery water. a mL; b μmol mg−1 h−1; c mol mol−1; d mL L−1 h−1.
Table 5.
Comparisons of bio-hydrogen production with synergistic effects of adding biomass immobilization and metals.
Table 5.
Comparisons of bio-hydrogen production with synergistic effects of adding biomass immobilization and metals.
| Additives | Opt/mg L−1 | Organism | Feed | Process | Temp/°C | Yielda | Reference |
|---|---|---|---|---|---|---|---|
| AC | 200 | AS | Glucose | Batch | 60 | 1.77 c (+106%) | [122] |
| BC | 10 | AS | Food waste | Batch | 35 | 1475 d (+41%) | [123] |
| Fe0 + AC | 100 | HTS | CBR | Batch | 30 | 83 b (+48%) | [124] |
| Fe (NPs) + CAB | 1000 | AS | PW | Batch | 38 | 298 b (+400%) | [125] |
| Gel | 0.2% (w/v) | EA | Glucose | Batch | 30 | 1.77 c (+80%) | [126] |
| PF | 2500 | AS | Glucose | Continuous | 37 | 0.6 c (+21%) | [127] |
| Sponge | - | EA | Starch | Continuous | 40 | 3.03 c (+37%) | [128] |
| Foam + Fe | 0.7 (g) | EA | Molasses | Continuous | 37 | 3.5 c (+77%) | [129] |
| Fe2 ++ BC | 300 | HTS | Glucose | Batch | 35 | 234 b (+48%) | [130] |
| Ni (NPs) + BC | 35 | CB | Glucose | Batch | 35 | 238 b (+49%) | [70] |
AC: activate carbon; BC: biochar; CBR: corn-bran residue, PF: polyurethane foam. AS: anaerobic sludge; HTS: heat treated sludge; EA: Enterobacter aerogenes; CB: Clostridium. Butyricum. CBR: corn-bran residue; PW: potato waste. a mL; b mL g−1; c mol mol−1; d mL L−1 h−1.
Table 6.
The comparisons of different technical routes for hydrogen production, and their effectiveness.
Table 6.
The comparisons of different technical routes for hydrogen production, and their effectiveness.
| Route | Feedstock | Energy Efficiency/% |
|---|---|---|
| SR | Hydrocarbons | 70–85 a |
| POX | Hydrocarbons | 60–75 a |
| ATR | Hydrocarbons | 60–75 a |
| Plasma reforming | Hydrocarbons | 9–80 b |
| Pyrolysis | Coal | 50 a |
| Co-Pyrolysis | Coal + Waste material | 80 a |
| Photolysis | Solar + water | 0.5 c |
| DF | Biomass | 60–80 d |
| Photofermentation | Biomass + Solar | 0.1 e |
| Microbial electrolysis | Biomass + Electric | 78 f |
| PWS | Water + Solar | 12.4 g |
PWS photo-electrochemical water-splitting. a Thermal efficiency based on the higher heating values; b Does not include hydrogen purification; c Conversion of solar energy to hydrogen by water-splitting excluding hydrogen purification; d Theoretical maximum of 4 mol H2 for 1 mol of glucose; e Conversion of solar energy to hydrogen by organic materials excluding hydrogen purification; f Total energy efficiency including applied voltage and energy in the substrate, excluding hydrogen purification; g Conversion of solar energy to hydrogen by water-splitting excluding hydrogen purification.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sun, Y.; He, J.; Yang, G.; Sun, G.; Sage, V. A Review of the Enhancement of Bio-Hydrogen Generation by Chemicals Addition. Catalysts 2019, 9, 353. https://doi.org/10.3390/catal9040353
AMA Style
Sun Y, He J, Yang G, Sun G, Sage V. A Review of the Enhancement of Bio-Hydrogen Generation by Chemicals Addition. Catalysts. 2019; 9(4):353. https://doi.org/10.3390/catal9040353
Chicago/Turabian StyleSun, Yong, Jun He, Gang Yang, Guangzhi Sun, and Valérie Sage. 2019. "A Review of the Enhancement of Bio-Hydrogen Generation by Chemicals Addition" Catalysts 9, no. 4: 353. https://doi.org/10.3390/catal9040353
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.