
Breeding Soft Durum Wheat through Introgression of the T5AL·5VS Translocated Chromosome
by

Wen Li
1,†, 
Yi Wei
1,†,
Yinyu Jin
1,
Heyu Chen
1,
Lingna Kong
1,
Xiaoxue Liu
1,
Liping Xing
1,2,
Aizhong Cao
1,2 and
Ruiqi Zhang
1,2,* 1
State Key Laboratory of Crop Genetics & Germplasm Enhancement and Application/JCIC-MCP, College of Agronomy, Nanjing Agricultural University, Nanjing 210095, China
2
Zhongshan Biological Breeding Laboratory, No. 50 Zhongling Street, Nanjing 210014, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Agronomy 2024, 14(4), 848; https://doi.org/10.3390/agronomy14040848
Submission received: 28 March 2024
/
Revised: 12 April 2024
/
Accepted: 13 April 2024
/
Published: 18 April 2024
(This article belongs to the Section Crop Breeding and Genetics)
Abstract
:The limited culinary utilizations of durum wheat (Triticum turgidum ssp. durum) are partly related to its very hard kernel texture, which is due to the softness genes Puroindoline a (Pina) and Puroindoline b (Pinb) on the Hardness (Ha) locus eliminated during allopolyploid formation. A previous study has reported that the softness genes Dina/Dinb, homologous to Pina/Pinb, were located on the chromosome arm 5VS of wild species Dasypyrum villosum. In the present study, we describe the process of transferring the soft grain texture from D. villosum into durum wheat through homoeologous recombination to develop a Robertsonian translocation. A durum wheat–D. villosum T5AL·5V#5S translocation line, S1286, was developed and characterized by molecular cytogenetic analysis from BC4F2 progeny of durum cv. ZY1286/D. villosum 01I140. The translocation line S1286 exhibited a soft grain texture as evidenced by observation through an electron microscope and a Single Kernel Characterization System (SKCS) hardness value of 5.5. Additionally, a newly developed 5VS/5AS co-dominant InDel marker, LW5VS-1, facilitated the transfer of the T5AL·5V#5S translocated chromosome into diverse durum wheat backgrounds. Subsequently, the T5AL·5V#5S translocated chromosome was transferred into five high-yielding durum wheat backgrounds by backcrossing and traced using marker LW5VS-1. Compared with each recurrent parent, T5AL·5V#5S lines showed good viability, similar development, and no yield penalty. Meanwhile, a significant decrease in plant height of about 6.0% was observed when comparing T5AL·5V#5S translocation lines with their recurrent parents. Accordingly, our results provide an efficient strategy for developing soft kernel durum wheat through the combination of T5AL·5V#5S translocation and the co-dominant marker LW5VS-1, which will be crucial for meeting the future challenges of sustainable agriculture and food security.
1. Introduction
Durum wheat (Triticum turgidum subsp. durum; 2n = 28, AABB) is the second most widely grown wheat species and an agronomically competitive crop of common wheat in low-rainfall and semiarid regions [1]. However, durum wheat exhibits a very hard kernel texture, leading to the milling product having large particle size and a high level of damaged starch that limits its end-use production. Therefore, a soft kernel durum germplasm needs to be developed to expand its limited use [2].
Grain texture in common wheat is primarily conditioned by the Hardness (Ha) locus on chromosome arm 5DS, which is comprised of two linked genes: Puroindoline a (Pina) and Puroindoline b (Pinb) [2,3]. Expression of wild-type Pina and Pinb in common wheat gives a soft kernel phenotype, while specific Pina and/or Pinb alleles of deletions or mutations result in a hard kernel. The wild relatives of wheat all possess forms of the Pina and Pinb homologs [3]. However, the large deletions fixed in the Ha loci of tetraploid wheat during allopolyploid formation and all the homologs of Pina and Pinb are eliminated on the A and B genomes, which results in durum generally having a harder texture than any hexaploid wheat [3,4].
Due to the cause-and-effect relationship between the homologs of Pina and Pinb and kernel hardness, efforts have been made to alter the kernel texture of durum wheat through introduction of the genes Pina and Pinb [4]. A small portion of the distal tip of a chromosome 5D short arm (5DS) of the Chinese Spring (CS), carrying genes Pina and Pinb, has been transferred to chromosome 5B, forming a 5BL·5BS-5DS translocation by non-homologous chromosomes recombination in the ph1b mutation [2,5,6]. Subsequently, a soft durum line Svevo was developed through introgression of the 5BL·5BS-5DS translocated chromosome [7]. In addition, overexpression of the Pina gene in transgenic durum wheat led to a medium-hard kernel texture [8]. The analysis of milling and baking quality indicated that the soft grain texture of durum wheat impacted nearly every aspect of milling, baking, pasta, and noodle quality [9,10,11,12,13,14,15]. By contrast, the traits of grain hardness index, break flour yield, and flour yield of soft durum wheat were similar to commercial soft white wheat cultivars, but soft kernel durum wheat exhibited some advantages over hexaploid wheat in noodle quality, particularly in low discoloration and PPO activity and high yellow color [12,13,14]. Consequently, development of an efficient strategy for breeding soft kernel durum will facilitate planting and utilizing durum wheat globally.
The presence of functional Pina/Pinb homologous genes in wheat wild relatives also can be transferred into wheat to vary the range in grain texture [16,17,18,19]. A previous study has shown that the chromosome arm 5VS of Dasypyrum villosum (2n = 2x = 14, VV), an annual related species of wheat, possesses forms of the Pina and Pinb homologs, designated as Dina and Dinb, repetitively [20]. Wheat–D. villosum T5DL·5V#4S and T6AL·6AS-5V#4S translocation lines have been developed to introduce the softness genes into common wheat, providing successful cases of expanding wheat grain texture by alien introgression [20,21]. In addition, the chromosome arm 5V#5S from D. villosum accession 01I140 harbors the linked genes Pm5V/Yr5V (resistance to powdery mildew and stripe rust) simultaneously [22]. Moreover, the T5DL·5V#5S translocated chromosome introgressed into different bread wheat backgrounds showed no yield penalty [22,23]. Therefore, introgression of the chromosome arm 5V#5S from D. villosum accession 01I140 into durum wheat by developing Robertsonian translocation has potential value for breeding a soft grain texture.
Here, we aimed to transfer chromosome arm 5V#S from D. villosum 01I140 to durum wheat by developing the compensating translocation. A durum wheat–D. villosum T5AL·5V#S translocation was identified by a combination of cytological and molecular approaches. The T5AL·5V#S translocation line S1286 has a soft grain texture without yield penalty. Thus, the combination of the T5AL·5V#S translocated chromosome, molecular selection tool, and speed breeding provides an efficient strategy for developing soft kernel durum wheat.
2. Materials and Methods
2.1. Plant Materials
The wild species D. villosum accession 01I140 (V#5), collected in the Pisa region of Italy, was initially crossed with spring durum wheat cv. ZY1286 (introduced from CIMMYT) to generate F1 hybrids, and the F1 plants were fertilized using the pollen of ZY1286. GISH detection was performed in BC1F1 and BC2F1 seeds. From the BC4F2 progeny of ZY1286/01I140//4*ZY1286, a homozygous durum wheat–D. villosum T5AL·5V#5S translocation line, S1286, was identified by molecular cytological approaches (see flow chart in Figure 1). The other five durum wheat cultivars, ZY1728, ZY1730, ZY1731, ZY1732, and JY001, with high yield potential were used to backcross with S1286 to develop new soft durum lines. To identify and locate markers specific to wheat group 5 chromosomes and D. villosum chromosome 5V, the Chinese Spring and nulli-tetrasomic (NT) lines N5AT5D were also used in this study. All the genetic stocks used in the present study are maintained at the Cytogenetics Institute, Nanjing Agricultural University (CINAU).
2.2. Rapid Alien Introgression
To accelerate the alien introgression process, we combined a speed breeding method with chromosome engineering approaches. We selected spring durum wheat cv. ZY1286 as a recurrent parent and vernalized the backcrossed seeds at 4 °C for seven days after germination. Next, the progenies of ZY1286/01I140 were grown in custom-made growth chambers (3 m × 3 m × 3 m) illuminated with LED lights set to an 18 h photoperiod (6 h of darkness) and temperatures 22 °C and 18 °C [22]. It takes about three months to produce a generation.
2.3. Cytological Detection
The alien chromatins of D. villosum were detected by genomic in situ hybridization (GISH) and fluorescence in situ hybridization (FISH) technologies. The root tips (1.5–2.0 cm) of the materials were treated through nitrous oxide gas and 90% acetic acid. The genome DNA of D. villosum accession 01I140, labeled with fluorescein-12-dUTP (green), was used as a probe for GISH detection [24]. The combination of oligonucleotide probes Oligo-pSc119.2-1 and Oligo-AFA-1,2, synthesized by Shanghai Sangon Biotech Co., Ltd. (Shanghai, China) and labeled with 6-carboxytetramethylrhodamine (TAM), was used to identify the chromosomes of durum wheat A and B genomes by nondenaturing fluorescence in situ hybridization (NDFISH) [25]. GISH/FISH signals and cytological images were observed and captured using an Olympus BX60 microscope (Olympus, Tokyo, Japan).
2.4. Molecular Marker Analysis
The total genomic DNAs of all materials were isolated using a modified CTAB method. The 5VS-specific molecular marker CINAU5V-237 (F: GTTTATCAGGCGGTGCCATA; R: GGACTTCTTGCTCCCCTTTC) was used to screen the progenies of ZY1286/01I140 for selection of the translocations carrying chromosome arm 5VS [22]. An InDel position between orthologous genes Dv05G092600 of D. villosum 5VS (https://bigd.big.ac.cn) and TraesCS5A02G075900 of Chinese Spring 5AS (http://wheatomics.sdau.edu.cn/) was used to develop a new 5VS/5AS co-dominant molecular LW5VS-1 (F: CGGAATGTATGGGGAACAT; R: GCTAGATAGACAGATTTATGGC) for transferring T5AL·5V#5S into diverse durum backgrounds. The PCR amplification was conducted using a T100TM Thermal Cycler (Bio-RAD Laboratories, Emeryville, CA, USA) following the procedure described by Zhang et al. (2022) [22]. The amplification samples were separated on 8% nondenaturing polyacrylamide gels for CINAU5V-237 and were separated in 1% agarose gels for LW5VS-1.
2.5. Evaluation of Grain Texture
To measure the grain hardness, 50 seeds per plant or 300 seeds per line were analyzed using the Perten Single Kernel Characterization System (SKCS) 4100 (Perten Instruments, Inc., Springfield, IL, USA), following the AACC International approved method 55-31 (AACCI, 2000). In addition, the mature dry seeds of T5AL·5V#S translocation line S1286 and durum cv ZY1286 were longitudinally cut in the center, and the halves were attached to an aluminum electron microscope stub using double-sided tape with the cut side upward. The samples were then coated with gold and examined using a Hitachi SEM-600 (Hitachi High-Technologies Corp., Tokyo, Japan). Images of the grain texture were captured at magnifications of 1.5K (15 kV) and 3.0K (15 kV).
2.6. Agronomic Traits Assessment
The wheat–D. villosum T5AL·5V#5S translocation lines and their recurrent parents were planted in a natural field located in the experimental station of Jiangsu Academy of Agricultural Sciences, Nanjing. Each plot consisted of four rows 1.5 m long and 0.25 m in width between each row. Twenty seeds per row were hand-seeded. Yield trials of each line were performed in randomized plot design with three replications. The agronomic traits were evaluated in 2022 and 2023 planting seasons. Plants were sprayed with pesticides and fungicides to control any insects and diseases. At the physiology maturity stage, twenty plants located in the middle of the internal two rows of each line were randomly selected for the analysis of yield-related traits, such as plant height, spike length, spike number per plant, spikelet number per spike, kernel number per spike, and thousand-kernel weight.
2.7. Statistical Analysis
The mean values and standard errors of the data on agronomic and grain quality treatments were determined using Microsoft Excel. t-tests or Tukey’s tests were then conducted using SPSS 26.0 statistical software (SPSS, Inc., Chicago, IL, USA) to determine the significance of differences. Significant differences between the means of two treatments were determined with a probability (p) value.
3. Results
3.1. Development and Identification of a Durum–D. villosum T5AL·5VS Translocation
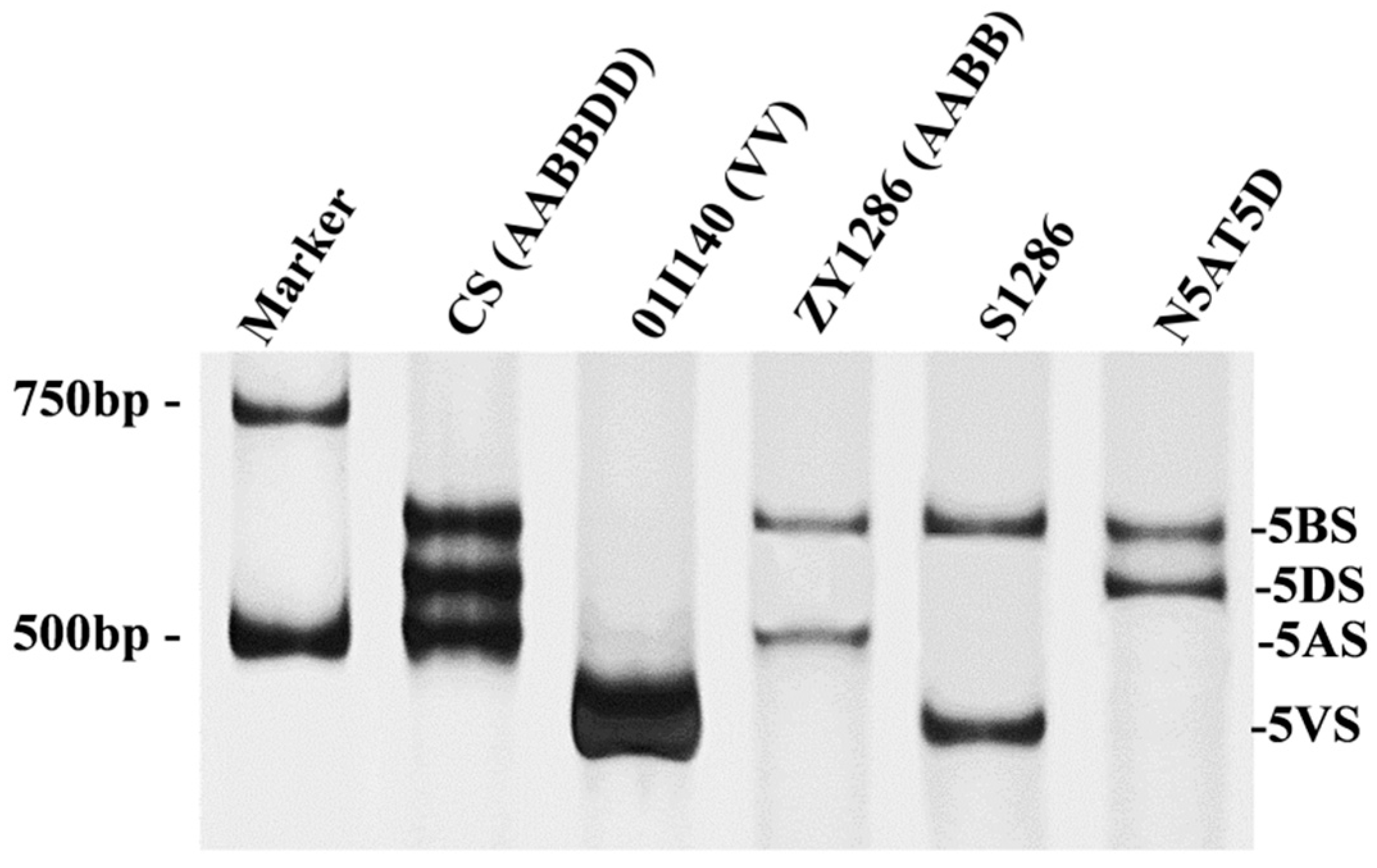
To transfer chromosome arm 5VS into durum wheat, durum cv ZY1286 was crossed with D. villosum accession 01I140. In their F1 plants, the chromosomes of A, B, and V genomes appeared as univalents at meiotic metaphase I, with a tendency to break and fuse at the centromeres, leading to the formation of Robertsonian translocations. We initially screened the Robertsonian whole-arm translocations in BC1F1 and BC2F1 progenies by GISH (Figure 1). Subsequently, the 5VS-specific molecular marker CINAU5V-237 was used to identify the translocation plants with chromosome arm 5V#5S. Plant T20-3-26 was identified to contain TW·5V#5S translocation from approximately 400 seeds of BC1F1 progeny (Figure 2a). Through backcrossing, the homozygous TW·5V#5S translocation plant S1286 was developed from the BC4F2 progeny. The cell of plant S1286 contained 28 chromosomes, pairing 14 bivalents (Figure 2b,c). To confirm the chromosome component, we conducted seed root-tip cell analysis of S1286 using GISH/FISH techniques. The patterns of GISH/FISH indicated that the opposite arm of the 5V#5S translocated chromosome in S1286 was chromosome arm 5AL (Figure 2d). The 5VS-specific molecular CINAU5V-237 identification showed that 5VS- and 5BS-specific bands were all present, while the 5AS-specific band was absent (Figure 3), confirming that S1286 is a T5AL·5V#5S translocation line.
3.2. Determination of Grain Texture by SEM and SKCS
To characterize the grain texture of S1286, we examined the degree of direct adhesion between starch granules and matrix proteins of freeze-fractured grain using scanning electron microscopy (SEM). The results showed a clear difference between ZY1286 and S1286 in terms of the degree of direct adhesion between starch granules and matrix proteins. The matrix proteins were adhered to the starch granules in ZY1286 (Figure 4a,b), whereas they cleanly separated from the surface of the starch granules in S1286 (Figure 4c,d). The grain hardness index (SKCS) showed that the average hardness of ZY1286 was 75.9, whereas for S1286 it was 5.5, a significant reduction of approximately 90%. These results indicated that introgression of D. villosum chromosome arm 5VS into durum chromosome 5A changes the fundamental grain properties of durum wheat, resulting in a soft grain texture.
3.3. Introgression of the T5AL·5V#5S Translocated Chromosome into Diverse Durum Varieties
An InDel position between orthologous genes Dv05G092600 of D. villosum 5VS and TraesCS5A02G075900 of wheat 5AS was converted into a breeding-friendly co-dominant marker, LW5VS-1, that can be used to trace the T5AL·5V#5S translocated chromosome in diverse durum wheat genetic backgrounds (Figure 5). The soft grain T5AL·5V#5S parent S1286 was transferred to elite durum cv. ZY1728, ZY1730, ZY1731, ZY1732, and JY001 by backcrossing and tracing with the co-dominant marker LW5VS-1; BC4F2 populations of each durum variety were obtained in two years by speed breeding. The segregation ratios of homozygous T5AL·5V#5S plants: heterozygous plants: non-T5AL·5V#5S plants all fitted to 1:2:1 in the five BC4F2 populations (Table 1), suggesting that the transmission of the T5AL·5V#5S translocated chromosome in diverse durum backgrounds is normal. SKCS index results showed that all the T5AL·5V#5S homozygous plants exhibited significantly lower HI (≤20) than that in each recurrent parent (>60) (Table 2). These results suggest that introgression of the T5AL·5V#5S translocated chromosome into diverse durum backgrounds leads to grain texture being changed dramatically.
3.4. Evaluation of Agronomic Traits
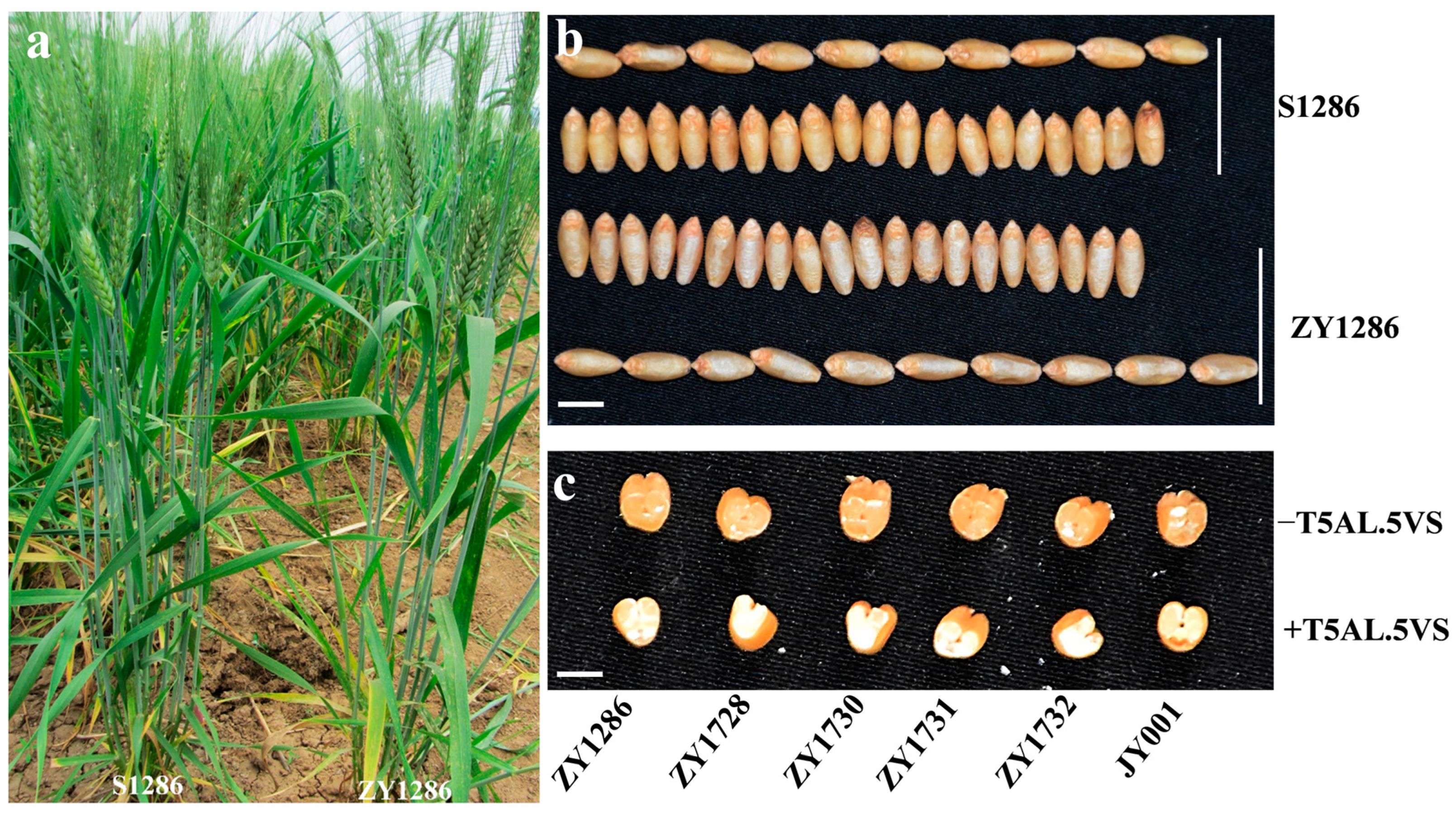
To investigate whether the T5AL·5V#5S translocated chromosome has a negative effect on the agronomic traits of durum wheat, we compared the yield-related traits between ZY1286 and S1286 under field conditions. Upon detailed characterization, S1286 was found to exhibit normal fertility and similar developmental stages to ZY1286, but it had a reduced plant height of approximately 5.0 cm compared to ZY1286 (Figure 6a, Table 2). In contrast, the seed width of S1286 was increased, but its length was decreased (Figure 6b). However, S1286 did not exhibit a significant reduction in yield-related traits such as spike length, seeds per spike, spikelet number per spike, and thousand-kernel weight, but there was a small increase in spike number per plant (Table 2).
In addition, five homozygous T5AL·5V#5S lines (BC4F3 and BC4F4) were selected in each durum background for further trait evaluation (Table 2). All T5AL·5V#5S lines exhibited a significantly lower plant height than their recurrent parents, a reduction of about 6.0%. However, T5AL·5V#5S NILs’ in-field conditions showed no significant reduction in spike length, spikelet number per spike, and kernel number per spike. The spike number per plant values for T5AL·5V#5S lines derived from ZY1728, ZY1730, ZY1731, and ZY1732 were significantly higher than those of the respective parents, whereas JY001 and its derived T5AL·5V#5S lines had no obvious differences. Thousand-kernel weight values were not consistent for different backcross derivatives. Derivatives from ZY1728, ZY1730, and ZY1731 had slightly lower thousand-kernel weight values, whereas T5AL·5V#5S lines derived from ZY1732 and JY001 produced similar values than their recurrent parents. These results indicated that thousand-kernel weight was largely influenced by genetic background. Thus, the selection of parents with acceptable thousand-kernel weight values will be essential when using T5AL·5V#5S translocation to breed soft durum. As expected, all the T5AL·5V#5S lines showed a very soft grain texture with significantly lower values in the grain hardness index (Table 2, Figure 6c). In summary, T5AL·5V#5S could have no significant negative effects on agronomic traits, providing a useful source for expanding durum wheat grain texture.
4. Discussion
Durum wheat is an underutilized crop due to it very hard kernel texture, causing its currently small range of uses, primarily pasta and couscous [26]. Development of soft kernel durum can remove this limitation [2,7]. Although the yield of common wheat is better than that of durum wheat in most production areas, the photosynthesis process in durum wheat appears to be more resilient than common wheat under heat stress [1,27]. Physiologically, durum may have advantages in heat-stressed conditions, as its yields can be equal to or slightly better than common wheat. Some field studies with large panels of cultivars in particular environments have shown good yield potential for durum wheat; for example, in the Yaqui Valley of Mexico, durum wheat has the potential to produce greater grain yields than common wheat [27]. In many areas, durum wheat appears to have a competitive yield and biotic and abiotic advantages over bread wheat [26]. Climate change is threatening durum and bread wheat production due to more severe environmental conditions and the spread of new virulent races of fungal diseases [28]. Thus, developing soft durum allows farmers to choose from more varieties, of either durum or common wheat, that are best adapted to the local environment. In the present study, we aimed to develop soft durum through genetically compensating Robertsonian translocations between durum wheat chromosome 5A or 5B and D. villosum chromosome 5V. Theoretically, all the chromosomes of A, B, and V genomes are univalents in the F1 hybrids of ZY1286/01I140, which led to the homoeologous chromosomes of 5A, 5B, and 5V having a tendency to break at the centromeres. This was followed by fusion of the broken arms, giving rise to Robertsonian whole-arm translocations of T5AL·5VS and T5BL·5VS. However, despite this tendency, only a T5AL·5VS translocation was identified among approximately 400 seeds from the BC1F1 progeny. This indicated that Robertsonian whole-arm translocations between chromosomes 5V and 5A/5B are rare in the durum background. Previously, only a soft durum germplasm, the 5BL·5BS-5DS translocation line, was developed [2,7]. The developed durum–D. villosum T5AL·5V#5S translocation provides a distinct durum germplasm with a soft grain texture.
Variation in the tryptophan-rich domain of Pina/Pinb homologs in wheat and its wild relatives is of particular importance to the structure and function of these proteins and to grain texture [29,30]. Compared with Pina and Pinb, a nonsynonymous substitution (amino acid-altering) mutation (Lys-42 to -Arg) was present in both tryptophan-rich regions of the Pina/Pinb homologous genes Dina and Dinb located on chromosome arm 5VS of D. villosum. As a result, the T5DL·5VS translocation line has a much softer grain texture than that of wild genotype Pina/Pinb in common wheat [20]. In this study, all the durum–D. villosum T5AL·5V#5S (Dina/Dinb) translocation lines in the six durum backgrounds exhibited a soft grain texture with HI values of less than 20 (Table 2). In comparison, the HI values of 5BL·5BS-5DS (Pina/Pinb) translocation lines in several durum backgrounds were between 20 and 40 [2]. It seems that durum with the T5AL·5V#5S translocated chromosome has a lower grain hardness than that with the 5BL·5BS-5DS translocated chromosome. Several studies have been undertaken to characterize the end-use quality of the initial soft durum germplasm [10,11,12,13,14,15]. These soft durum lines with the 5BL·5BS-5DS translocated chromosome exhibited versatile characteristics and highly desirable end-use quality for cookies, hearth bread, noodles, and pasta [10,11,12]. Therefore, a further analysis comparing the end-use quality between the durum–D. villosum T5AL·5V#5S translocation line and common wheat needs to be carried out through the production of products.
The present study also investigated whether the T5AL·5V#5S translocated chromosome affects the yield-related traits of durum wheat. Plant height is one of the most important traits in wheat improvement [31]. All the T5AL·5V#5S translocation lines of different backgrounds had a slightly lower plant height than that of each recurrent parent, indicating that this parameter was influenced predominantly by the introgressed chromosome arm 5VS of D. villosum. Furthermore, no significant differences were observed in the spike length, spikelet number per spike, and kernel number per spike between T5AL·5V#5S translocation lines and their background parents. The improved ability of spikes per plant, together with the decreased thousand-kernel weight under a certain genetic background, suggested that it was a promising genotype of T5AL·5V#5S translocation for use in crossing and selection programs aimed at soft durum wheat improvement. Further field trials using a larger plot area/genotype will be needed for a more exact comparison of yield parameters between the durum–D. villosum T5AL·5V#5S translocation line and the recurrent parent.
Grain hardness assays of physical methods usually require expensive equipment, they are time-consuming and damage the seeds after harvest, and they are rarely used in wheat breeding [32]. As such, a suite of new approaches that provide low-cost and accurate prediction of a soft kernel would facilitate soft durum improvement [33]. Soft-grained T5AL·5V#5S introgression in diverse, well-adapted durum wheat genetic backgrounds without a yield penalty confirmed its potential breeding utility. The co-dominant maker LW5VS-1 developed based on a conserved InDel between orthologous genes 5VS of D. villosum and 5AS of wheat provides a solution for identifying the soft plants when segregating populations early in the breeding cycle when using T5AL·5V#5S introgression in durum wheat improvement. As a result, the combination of maker LW5VS-1 and speed breeding technology will facilitate transferring T5AL·5V#5S into diverse durum wheat backgrounds speedily.
5. Conclusions
In this study, a durum wheat–D. villosum T5AL·5V#5S translocation line, S1286, with a soft grain texture was developed and characterized by detailed molecular cytogenetic analysis. A co-dominant maker, LW5VS-1, developed based on a conserved InDel between orthologous genes 5VS and 5AS facilitated transferring the T5AL·5V#5S translocated chromosome into diverse durum wheat backgrounds. The evaluation of major agronomic traits of lines with T5AL·5V#5S translocation in several high-yielding durum wheat backgrounds showed no obvious yield penalty. Therefore, the developed T5AL·5V#5S translocation together with the new molecular marker provide an efficient strategy for developing soft kernel durum wheat, which will be required to meet the future challenges of sustainable agriculture and food security.
Author Contributions
R.Z. conceived and designed the experiments. W.L. and Y.W. developed introgression lines. Y.J., H.C., L.K. and X.L. analyzed GISH/FISH and molecular markers. L.X. and A.C. analyzed agronomic traits. R.Z., Y.W. and W.L. wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32272062) and the National Key Research and Development Program of China (2023YFD1200402).
Data Availability Statement
Data are contained within this article. The plant materials and datasets generated and analyzed during the present study are available from the corresponding authors upon reasonable request.
Conflicts of Interest
All the authors have no conflicts of interest, and they agree with the publication.
References
- Dias, A.S.; Semedo, J.; Lidon, F.C. Bread and durum wheat under heat stress: A comparative study on the photosynthetic performance. J. Agron. Crop Sci. 2011, 197, 50–56. [Google Scholar] [CrossRef]
- Morris, C.F.; Simeone, M.C.; King, G.E.; Lafiandra, D. Transfer of soft kernel texture from Triticum aestivum to durum wheat, Triticum turgidum ssp. durum. Crop Sci. 2011, 51, 114–122. [Google Scholar] [CrossRef]
- Bhave, M.; Morris, C.F. Molecular genetics of puroindolines and related genes: Allelic diversity in wheat and other grasses. Plant Mol. Biol. 2008, 66, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Huang, L.; Gill, B.S. Recurrent deletions of puroindoline genes at the grain hardness locus in four independent lineages of polyploid wheat. Plant Physiol. 2008, 146, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Boehm, J.D.; Zhang, M.; Cai, X.; Morris, C.F. Molecular and cytogenetic characterization of the 5DS-5BS chromosome translocation conditioning soft kernel texture in durum wheat. Plant Genome 2017, 10, 1–11. [Google Scholar] [CrossRef]
- Ibba, M.I.; Zhang, M.; Cai, X.; Morris, C.F. Identification of a conserved ph1b-mediated 5DS-5BS crossing over site in soft-kernel durum wheat lines. Euphytica 2019, 215, 200. [Google Scholar] [CrossRef]
- Morris, C.F. Development of soft kernel durum wheat. Front. Agric. Sci. Eng. 2019, 6, 273–278. [Google Scholar] [CrossRef]
- Li, Y.; Mao, X.; Wang, Q.; Zhang, J.; Li, X.; Ma, F.; Sun, F.; Chang, J.; Chen, M.; Wang, Y.; et al. Overexpression of puroindoline a gene in transgenic durum wheat (Triticum turgidum ssp. durum) leads to a medium–hard kernel texture. Mol. Breed. 2014, 33, 545–554. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.; Sun, F.; Li, X.; Wang, P.; Yang, G.; He, G. Expression of Puroindoline a in durum wheat affects milling and pasting properties. Front. Plant. Sci. 2019, 10, 482. [Google Scholar] [CrossRef]
- Gazza, L.; Sgrulletta, D.; Cammerata, A.; Gazzelloni, G.; Perenzin, M.; Pogna, N.E. Pasta making and bread making quality of soft-textured durum wheat lines. J. Cereal Sci. 2011, 54, 481–487. [Google Scholar] [CrossRef]
- Heinze, K.; Kiszonas, A.M.; Murray, J.C.; Morris, C.F.; Lullien-Pellerin, V. Puroindoline genes introduced into durum wheat reduce milling energy and change milling behavior similar to soft common wheats. J. Cereal Sci. 2016, 71, 183–189. [Google Scholar] [CrossRef]
- Murray, C.; Kiszonas, A.K.; Morris, C.F. Influence of soft kernel texture on the flour, water absorption, rheology, and baking quality of durum wheat. Cereal Chem. 2017, 94, 215–222. [Google Scholar] [CrossRef]
- Murray, J.C.; Kiszonas, A.M.; Wilson, J.D.; Morris, C.F. Effect of soft kernel texture on the milling properties of soft durum wheat. Cereal Chem. 2016, 93, 513–517. [Google Scholar] [CrossRef]
- Boehm, J.D.; Ibba, M.I.; Kiszonas, A.M.; Morris, C.F. End-use quality of CIMMYT-derived soft-kernel durum wheat germplasm: I. Grain, milling and soft wheat quality. Crop Sci. 2017, 57, 1475–1484. [Google Scholar]
- Boehm, J.D.; Ibba, M.I.; Kiszonas, A.M.; Morris, C.F. End-use quality of CIMMYT-derived soft-kernel durum wheat germplasm: II. Dough strength and pan bread quality. Crop Sci. 2017, 57, 1485–1494. [Google Scholar] [CrossRef]
- Gasparis, S.; Orczyk, W.; Nadolska-Orczyk, A. Sina and Sinb genes in triticale do not determine grain hardness contrary to their orthologs Pina and Pinb in wheat. BMC Plant Biol. 2013, 13, 190. [Google Scholar] [CrossRef] [PubMed]
- Kalra, G.; Panozzo, J.; Bhave, M. Genetic variability of Hin genes in selected barley germplasm and potential implications for grain texture. Euphytica 2013, 215, 105. [Google Scholar] [CrossRef]
- Chen, M.; Wilkinson, M.; Tosi, P.; He, G.; Shewry, P. Novel puroindoline and grain softness protein alleles in Aegilops species with the C, D, S, M and U genomes. Theor. Appl. Genet. 2005, 111, 1159–1166. [Google Scholar] [CrossRef]
- Cuesta, S.; Guzmán, C.; Alvarez, J.B. Allelic diversity and molecular characterization of puroindoline genes in five diploid species of the Aegilops genus. J. Exp. Bot. 2013, 64, 5133–5143. [Google Scholar] [CrossRef]
- Zhang, R.Q.; Cao, Y.P.; Wang, X.E.; Feng, Y.G.; Chen, P.D. Development and characterization of a Triticum aestivum-H. villosa T5VS·5DL translocation line with soft grain texture. J. Cereal Sci. 2010, 51, 220–225. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, X.; Chen, P. Molecular and cytogenetic characterization of a small alien-segment translocation line carrying the softness genes of Haynaldia villosa. Genome 2012, 55, 639–646. [Google Scholar] [CrossRef]
- Zhang, R.; Lu, C.; Meng, X.; Fan, Y.; Du, J.; Liu, R.; Feng, Y.; Xing, L.; Cápal, P.; Holušová, K.; et al. Fine mapping of powdery mildew and stripe rust resistance genes Pm5V/Yr5V transferred from Dasypyrum villosum into wheat without yield penalty. Theor. Appl. Genet. 2022, 135, 3629–3642. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Du, J.; Chen, H.; Gong, S.; Jin, Y.; Meng, X.; Zhang, T.; Fu, B.; Molnár, I.; Holušová, K.; et al. Wheat Pm55 alleles exhibit distinct interactions with an inhibitor to cause different powdery mildew resistance. Nat. Commun. 2024, 15, 503. [Google Scholar] [CrossRef] [PubMed]
- Komuro, S.; Endo, R.; Shikata, K.; Kato, A. Genomic and chromosomal distribution patterns of various repeated DNA sequences in wheat revealed by a fluorescence in situ hybridization procedure. Genome 2013, 56, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Zhuang, L.; Wang, Y.; Yuan, L.; Wang, Q.; Wang, D.; Tan, L.; Shen, J.; Xu, H.; Zhao, H.; et al. Development of oligonucleotides and multiplex probes for quick and accurate identification of wheat and Thinopyrum bessarabicum chromosomes. Genome 2017, 60, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.F.; Kiszonas, A.M.; Murray, J.; Boehm, J.; Ibba, M.I.; Zhang, M.; Cai, X. Re-evolution of durum wheat by introducing the hardness and Glu-D1 loci. Front. Sustain. Food Syst. 2019, 3, 103. [Google Scholar] [CrossRef]
- Mastrangelo, A.M.; Cattivelli, L. What makes bread and durum wheat different? Trends Plant Sci. 2021, 26, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Curtis, T.; Halford, N.G. Food security: The challenge of increasing wheat yield and the importance of not compromising food safety. Ann. Appl. Biol. 2014, 164, 354–372. [Google Scholar] [CrossRef]
- Bonafede, M.; Kong, L.; Tranquilli, G.; Ohm, H.; Dubcovsky, J. Reduction of a Triticum monococcum chromosome segment carrying the softness genes Pina and Pinb translocated to bread wheat. Crop Sci. 2007, 47, 819–826. [Google Scholar] [CrossRef]
- Chantret, N.; Salse, J.; Sabot, F.; Rahman, S.; Bellec, A.; Laubin, B.; Dubois, I.; Dossat, C.; Sourdille, P.; Joudrier, P.; et al. Molecular basis of evolutionary events that shaped the Hardness locus in diploid and polyploid wheat species (Triticum and Aegilops). Plant Cell 2005, 17, 1033–1045. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Liu, Z.H.; Yang, B.; Chen, H.; Zhang, H.; Hou, D.B. The contribution of photosynthesis traits and plant height components to plant height in wheat at the individual quantitative trait locus level. Sci. Rep. 2020, 10, 12261. [Google Scholar] [CrossRef]
- Nucia, A.; Okoń, S.; Tomczyńska-Mleko, M.; Nawrocka, A. Molecular and physical characterization of grain hardness in European spring common wheat (Triticum aestivum L.). 3 Biotech. 2021, 11, 345. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Pradhan, S.; Shahi, D.; Khan, J.; Mcbreen, J.; Bai, G.; Murphy, J.P.; Babar, M.A. Increased prediction accuracy using combined genomic information and physiological traits in a soft wheat panel evaluated in multi-environments. Sci. Rep. 2020, 10, 7023. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Flow chart for transferring soft grain texture from D. villosum into durum wheat.

Figure 2.
Identification of the durum wheat–D. villosum 5V#5S introgression lines through genomic in situ (GISH) and fluorescent in situ (FISH) hybridization. D. villosum genomic DNA labeled with fluorescein-12-dUTP (green) as probes was used for GISH. Probes for FISH were Oligo-pSc119.2-1 and Oligo-AFA-1,2, labeled with TAM (red). Wheat chromosomes were counterstained with DAPI (blue). (a) GISH/FISH patterns of plant T20-3-26, containing a single TW·5V#5S translocation chromosome (arrow indicate). (b) GISH patterns of S1286 (2n = 28), containing a pair of TW·5V#5S translocated chromosomes. (c) GISH patterns of metaphase I in S1286 showing the translocated chromosome pair forming a ring bivalent. (d) GISH/FISH patterns of S1286, showing that the opposite arm of the 5V#5S translocated chromosome in S1286 was chromosome arm 5AL, which was a homozygous T5AL·5V#5S translocation line. Bars, 10 μm.
Figure 2.
Identification of the durum wheat–D. villosum 5V#5S introgression lines through genomic in situ (GISH) and fluorescent in situ (FISH) hybridization. D. villosum genomic DNA labeled with fluorescein-12-dUTP (green) as probes was used for GISH. Probes for FISH were Oligo-pSc119.2-1 and Oligo-AFA-1,2, labeled with TAM (red). Wheat chromosomes were counterstained with DAPI (blue). (a) GISH/FISH patterns of plant T20-3-26, containing a single TW·5V#5S translocation chromosome (arrow indicate). (b) GISH patterns of S1286 (2n = 28), containing a pair of TW·5V#5S translocated chromosomes. (c) GISH patterns of metaphase I in S1286 showing the translocated chromosome pair forming a ring bivalent. (d) GISH/FISH patterns of S1286, showing that the opposite arm of the 5V#5S translocated chromosome in S1286 was chromosome arm 5AL, which was a homozygous T5AL·5V#5S translocation line. Bars, 10 μm.

Figure 3.
PCR amplification patterns of 5VS-specific marker CINAU5V-237. The line S1286 has both 5VS- and 5BS-specific bands but lacks the bands of wheat 5AS and 5DS.
Figure 3.
PCR amplification patterns of 5VS-specific marker CINAU5V-237. The line S1286 has both 5VS- and 5BS-specific bands but lacks the bands of wheat 5AS and 5DS.

Figure 4.
Stereo-scanned electron microscopy of freeze-fractured grains of ZY1286 and S1286. The matrix protein adhered to the surfaces of the starch granules in ZY1286 (a,b) but separated in line S1286 (c,d). Magnification 1000× (a,c) and 2000× (b,d).
Figure 4.
Stereo-scanned electron microscopy of freeze-fractured grains of ZY1286 and S1286. The matrix protein adhered to the surfaces of the starch granules in ZY1286 (a,b) but separated in line S1286 (c,d). Magnification 1000× (a,c) and 2000× (b,d).

Figure 5.
PCR amplification patterns of 5VS/5AS co-dominant marker LW5VS-1 in ZY1286/S1286 F2 individuals.
Figure 5.
PCR amplification patterns of 5VS/5AS co-dominant marker LW5VS-1 in ZY1286/S1286 F2 individuals.

Figure 6.
Plant and seed morphology of T5AL·5V#5S translocation line and the recurrent parents. (a) The plants of S1286 and ZY1286, showing similar developmental stages. (b) Comparison of seeds between ZY1286 and T5AL·5V#5S translocation line S1286. Scale bar = 0.5 cm. (c) The transverse sections of seeds of different T5AL·5V#5S translocation lines and their recurrent parents. Scale bar = 0.5 cm.
Figure 6.
Plant and seed morphology of T5AL·5V#5S translocation line and the recurrent parents. (a) The plants of S1286 and ZY1286, showing similar developmental stages. (b) Comparison of seeds between ZY1286 and T5AL·5V#5S translocation line S1286. Scale bar = 0.5 cm. (c) The transverse sections of seeds of different T5AL·5V#5S translocation lines and their recurrent parents. Scale bar = 0.5 cm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of PCR marker identification for BC4F2 individuals.
| Combination | No. of Plants | Homozygous Plants/HI | Heterozygous Plants | Non-T5AL·5VS Plants/HI | χ2 (χ2 0.05,2 = 5.99) |
|---|---|---|---|---|---|
| ZY1728/S1286//4*ZY1728 (BC4F2) | 146 | 36 (8.6 ± 2.3) b | 70 | 40 (73.6 ± 9.4) a | 0.47 |
| ZY1730/S1286//4*ZY1730 (BC4F2) | 138 | 32 (15.1 ± 4.9) b | 73 | 33 (71.4 ± 8.2) a | 0.36 |
| ZY1731/S1286//4*ZY1731 (BC4F2) | 124 | 29 (9.7 ± 3.8) b | 65 | 30 (74.3 ± 6.9) a | 0.31 |
| ZY1732/S1286//4*ZY1732 (BC4F2) | 163 | 36 (13.4 ± 5.2) b | 88 | 39 (69.4 ± 5.8) a | 1.15 |
| JY0018/S1286//4* JY001 (BC4F2) | 135 | 31 (10.5 ± 4.5) b | 75 | 29 (70.6 ± 6.7) a | 1.78 |
Note: the HI values followed by different letters are significantly different at p = 0.05.
Table 2.
Comparison of the agronomic characteristics of T5AL·5V#5S lines with their recurrent parents.
Table 2.
Comparison of the agronomic characteristics of T5AL·5V#5S lines with their recurrent parents.
| Lines | Chromosome Component | Plant Height (cm) | Spike Length (cm) | Spikelet Number per Spike | Kernel Number per Spike | Spike Number per Plant | Thousand-Kernel Weight (g) | Grain Hardness Index |
|---|---|---|---|---|---|---|---|---|
| ZY1286 | 5AL·5AS | 89.6 a | 7.6 a | 18.7 a | 41.4 a | 8.2 a | 40.1 a | 75.9 a |
| S1286 | T5AL·5V#5S | 83.8 b | 7.5 a | 18.2 a | 40.3 a | 8.9 b | 39.4 a | 5.5 b |
| ZY1728 | 5AL·5AS | 93.4 a | 8.3 a | 19.3 a | 42.5 a | 8.4 a | 42.3 a | 73.2 a |
| S1728-1 | T5AL·5V#5S | 90.1 b | 8.1 a | 20.1 a | 43.4 a | 9.1 b | 40.2 b | 11.4 b |
| S1728-2 | T5AL·5V#5S | 89.4 b | 8.6 a | 19.7 a | 42.1 a | 8.7 b | 41.5 b | 10.8 b |
| S1728-3 | T5AL·5V#5S | 88.7 b | 8.4 a | 20.4 a | 43.3 a | 9.2 b | 40.9 b | 11.6 b |
| S1728-4 | T5AL·5V#5S | 87.5 b | 8.3 a | 20.6 a | 42.7 a | 9.4 b | 41.3 b | 10.5 b |
| S1728-5 | T5AL·5V#5S | 88.2 b | 8.5 a | 20.3 a | 43.1 a | 9.2 b | 40.3 b | 10.9 b |
| ZY1730 | 5AL·5AS | 92.7 a | 8.7 a | 20.3 a | 43.4 a | 9.2 a | 41.8 a | 71.4 a |
| S1730-1 | T5AL·5V#5S | 87.3 b | 8.3 a | 19.1 a | 42.8 a | 9.7 b | 40.1 b | 8.7 b |
| S1730-2 | T5AL·5V#5S | 88.2 b | 8.4 a | 19.5 a | 42.6 a | 10.2 b | 39.7 b | 6.6 b |
| S1730-3 | T5AL·5V#5S | 86.1 b | 8.7 a | 20.2 a | 43.1 a | 10.4 b | 39.9 b | 7.8 b |
| S1730-4 | T5AL·5V#5S | 86.5 b | 8.5 a | 19.3 a | 42.9 a | 9.8 b | 40.3 b | 9.5 b |
| S1730-5 | T5AL·5V#5S | 87.2 b | 8.2 a | 19.4 a | 42.7 a | 10.3 b | 39.4 b | 8.9 b |
| ZY1731 | 5AL·5AS | 87.1 a | 7.7 a | 18.8 a | 41.3 a | 9.4 a | 41.7 a | 74.6 a |
| S1731-1 | T5AL·5V#5S | 83.5 b | 7.1 a | 19.1 a | 41.6 a | 11.2 b | 38.6 b | 9.7 b |
| S1731-2 | T5AL·5V#5S | 82.5 b | 6.9 a | 19.2 a | 40.1 a | 11.9 b | 39.4 b | 11.4 b |
| S1731-3 | T5AL·5V#5S | 83.4 b | 7.5 a | 18.4 a | 39.8 a | 13.1 b | 38.7 b | 12.1 b |
| S1731-4 | T5AL·5V#5S | 85.6 b | 7.8 a | 18.5 a | 41.2 a | 12.4 b | 38.6 b | 8.5 b |
| S1731-5 | T5AL·5V#5S | 84.7 b | 7.3 a | 18.7 a | 40.5 a | 12.6 b | 39.5 b | 9.9 b |
| ZY1732 | 5AL·5AS | 88.7 a | 7.9 a | 18.7 a | 41.6 a | 8.8 a | 38.1 a | 74.8 a |
| S1732-1 | T5AL·5V#5S | 83.4 b | 7.5 a | 18.4 a | 40.7 a | 10.0 b | 37.4 a | 7.6 b |
| S1732-2 | T5AL·5V#5S | 83.9 b | 8.1 a | 19.2 a | 41.3 a | 9.6 b | 36.4 a | 8.5 b |
| S1732-3 | T5AL·5V#5S | 82.6 b | 7.7 a | 18.3 a | 39.8 a | 10.1 b | 38.0 a | 9.7 b |
| S1732-4 | T5AL·5V#5S | 84.7 b | 7.4 a | 18.1 a | 39.4 a | 9.5 b | 37.6 a | 9.4 b |
| S1732-5 | T5AL·5V#5S | 85.2 b | 8.0 a | 19.4 a | 40.2 a | 9.3 b | 37.3 a | 10.1 b |
| JY001 | 5AL·5AS | 96.1 a | 8.6 a | 20.0 a | 43.1 a | 12.3 a | 36.7 a | 69.8 a |
| E001-1 | T5AL·5V#5S | 90.7 b | 8.4 a | 19.4 a | 42.2 a | 13.4 b | 35.4 a | 5.2 b |
| E001-2 | T5AL·5V#5S | 91.4 b | 9.1 a | 20.4 a | 43.7 a | 12.9 b | 36.2 a | 6.4 b |
| E001-3 | T5AL·5V#5S | 91.7 b | 8.8 a | 20.1 a | 42.9 a | 14.5 b | 36.1 a | 7.7 b |
| E001-4 | T5AL·5V#5S | 92.3 b | 8.6 a | 19.6 a | 41.8 a | 13.7 b | 35.6 a | 5.8 b |
| E001-5 | T5AL·5V#5S | 90.3 b | 8.5 a | 19.3 a | 41.6 a | 12.4 b | 40.3 a | 5.4 b |
Note: the values followed by different letters are significantly different within each background at p = 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, W.; Wei, Y.; Jin, Y.; Chen, H.; Kong, L.; Liu, X.; Xing, L.; Cao, A.; Zhang, R. Breeding Soft Durum Wheat through Introgression of the T5AL·5VS Translocated Chromosome. Agronomy 2024, 14, 848. https://doi.org/10.3390/agronomy14040848
AMA Style
Li W, Wei Y, Jin Y, Chen H, Kong L, Liu X, Xing L, Cao A, Zhang R. Breeding Soft Durum Wheat through Introgression of the T5AL·5VS Translocated Chromosome. Agronomy. 2024; 14(4):848. https://doi.org/10.3390/agronomy14040848
Chicago/Turabian StyleLi, Wen, Yi Wei, Yinyu Jin, Heyu Chen, Lingna Kong, Xiaoxue Liu, Liping Xing, Aizhong Cao, and Ruiqi Zhang. 2024. "Breeding Soft Durum Wheat through Introgression of the T5AL·5VS Translocated Chromosome" Agronomy 14, no. 4: 848. https://doi.org/10.3390/agronomy14040848
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.