
Effect of Mild Organic Substitution on Soil Quality and Microbial Community
by
 , ,
, ,
Yijun Wang
1,†, 
Yu Xu
1,†,
Lihua Jiang
2,
Yan Yang
1,
Jing Shi
1,
Xilin Guan
1,
Tao Sun
1,
Huanyu Zhao
1,
Yafei Wang
3 and
Yumin Liu
1,* 1
State Key Laboratory of Nutrient Use and Management, Institute of Agricultural Resources and Environment, Shandong Academy of Agricultural Sciences, Jinan 250100, China
2
College of Resources and Environmental Engineering, Shandong Agriculture and Engineering University, Jinan 250100, China
3
Department of Physical and Chemical Inspection, School of Public Health, Shandong University, Jinan 250012, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Agronomy 2024, 14(5), 888; https://doi.org/10.3390/agronomy14050888
Submission received: 29 March 2024
/
Revised: 18 April 2024
/
Accepted: 22 April 2024
/
Published: 24 April 2024
(This article belongs to the Special Issue Effect of Fertilization on Soil Quality)
Abstract
:Mild organic substitution is advantageous for sustainable agricultural development. In order to determine the proper fertilization strategy, it is essential to investigate the impact of substituting chemical fertilizers with varying levels of organic manure on soil nutrients, microbial communities, and crop productivity. Four treatments were implemented: no fertilizer, sole chemical fertilizer, 20% organic manure substitution, and 40% organic manure substitution. Bacterial and fungal communities were characterized through high-throughput sequencing of the 16S rRNA gene V3–V4 region and the V4 region, respectively. The 20% and 40% organic manure substitutions increased soil organic matter (SOM) content, total nitrogen (TN) content, and reduced soil pH compared to the control (CK). The 20% organic manure substitution showed the most significant improvements in soil alkaline phosphatase, urease, and invertase activities. Soil nutrient enhancement increased bacterial alpha diversity, with a milder impact on fungal alpha diversity compared to bacteria. Different fertilization treatments elevated the relative abundance of bacterial Bacteroidetes (8.11%, 21.25%, and 1.88%), Actinomycetes (12.65%, 26.36%, and 15.33%), and fungal Ascomycota (16.19%, 10.44%, and 12.69%), known for degrading recalcitrant organic matter. The sole chemical fertilizer treatment increased the pathogenic Cheatotryiales. Shared species, primarily from bacterial Actinomycetes, Firmicutes, Proteobacteria, and fungal Ascomycota phyla, were found at 20% and 40% organic manure substitution levels. Specifically, the 20% organic manure substitution level promoted the relative abundance of beneficial plant growth-promoting taxa, Oxalobacteraceae and Massilia, and suppressed pathogens, with an increase in the relative abundance of the Purpureocillium genus and Mortierellomycota. These findings suggest that a 20% OF substitution can maintain crop yield, enhance soil nutrients and enzyme activities by fostering beneficial soil bacteria, inhibiting soil-borne pathogens, and refining microbial community structure.
1. Introduction
The application of both mineral and organic fertilizers (OF) is a recognized strategy for enhancing soil quality and crop yields. While mineral fertilization can marginally increase soil organic carbon content and crop yields [1], over-reliance or sole use can disrupt matter and energy cycling in the agroecosystem, reducing soil quality and degrading agricultural product standards. A transition from chemical fertilizers to OF is emerging as a sustainable agricultural practice. Multiple studies have demonstrated that combining organic and chemical fertilizers can improve soil fertility and structure by increasing soil organic matter (SOM) and nutrient content, thereby ensuring agricultural sustainability and crop yield [2,3,4,5]. For instance, Zhou et al. [4] found that blending organic and inorganic nitrogen fertilizers in a 1:1 and 2:1 ratio notably increased vegetable yields and N uptake. A meta-analysis by Xia et al. [5] indicated a 6.8% increase in crop productivity when substituting chemical fertilizers with organic manure.
In agroecosystems, soil microorganisms, the most dynamic components, play pivotal roles in soil structure formation, organic matter decomposition, toxin elimination, and the biogeochemical cycling of elements like carbon, nitrogen, phosphorus, and sulfur [6,7]. Agricultural interventions often compromise the diversity and abundance of these microbial communities, which are essential for soil ecosystem health and sustainability [8]. The addition of exogenous organic manure can affect microbial community structure [9,10,11]. Chemical fertilizers alone induce soil acidification and reduce microbial diversity [12,13]. In contrast, integrating OF at varying levels can modulate microbial community structure and diversity differently, depending on specific organic-inorganic combinations. Under the same chemical fertilizer application, Cui et al. [14] observed that bacterial community diversity significantly increased with organic manure addition. However, Sarathchandra et al. [15] found nitrogen and phosphorus fertilizers to be inconsequential for soil microbial attributes in rangelands. Cui et al. [16] found that prolonged co-application of organic and OF limits on microbial nitrogen shifts microbial communities from copiotrophic to oligotrophic dominance, thereby influencing organic carbon decomposition. The addition of OF can influence microbial communities by shifting oligotrophic microorganisms (which thrive in soils with pure mineral fertilizers or no fertilization) towards microorganisms capable of decomposing complex organic compounds, as well as changes in key species involved in nutrient cycling, thereby affecting soil fertility [17]. Therefore, it is necessary to further investigate the impact of appropriate organic-inorganic fertilizer ratios on the beneficial development of microbial community structure and the enhancement of soil quality.
In extensive investigations of SOM, distinct bacterial and fungal communities have been extensively studied, but most research has focused on nitrogen-centric OF substitutions, neglecting the potential impacts of excessive phosphorus and potash on microbial diversity. This oversight highlights the necessity for a more comprehensive approach. Previous studies examining the effects of OF substitution on soil microorganisms have employed varying substitution rates, often without considering crop yield implications [18]. For instance, yields significantly declined with over 50% OF substitution [19]. To prioritize crop yield while addressing this gap, our study implemented four treatments: no fertilizer, sole chemical fertilizer, 20% OF substitution, and 40% OF substitution. The main objectives of this study were to assess whether substituting chemical fertilizers with organic fertilizers could enhance soil quality while simultaneously sustaining yield. We hypothesized that an appropriate proportion of organic fertilizer substitution can modulate the microbial community structure towards beneficial microbial populations, thereby promoting the enhancement of soil quality while stabilizing yield.
2. Materials and Methods
2.1. Field Experiment Conditions
The field study in Jiyang County, Shandong Province, China (36°58′ N, 116°59′ E) commenced in 2016. A detailed study site map is provided in Supplementary Materials (Figure S1). The study area features a continental monsoon climate, with mean air temperature, relative humidity, and annual precipitation at 12.8 °C, 65%, and 583.3 mm, respectively. The soil is classified as a typical calcareous alluvial soil based on the World Reference Base classification [20]. The soil, initially sampled, was calcareous and moist, with the following basic characteristics: pH 8.7, 1:2.5 w/v ratio of soil: water, organic matter 14.2 g kg−1, total nitrogen 1.2 g kg−1, Olsen phosphorus 8.8 mg kg−1, and available potassium 94.5 mg kg−1.
2.2. Design of the Experiment
A crop rotation system of winter wheat (Triticum aestivum L.) and summer maize (Zea mays L.) was observed in the field experiment since the experiment was established. In each rotation year, wheat is sown in early October and harvested in early June of the following year. The wheat cultivar ‘Luyuan 502’ was machine-sown with a row spacing of 25 cm at a seeding rate of approximately 225 kg ha−1 in 2016–2021. The maize is planted after the wheat harvest and harvested in early October. The summer maize cultivar ‘Denghai 618’ was planted with a density of 75,000 plants ha−1 in all maize cropping years. The maize seeds were sown with a row spacing of 60 cm and a plant spacing of 22.2 cm. Four treatments were applied to triplicate 40 m2 plots:
- No N fertilizer (CK)
- Chemical N fertilizer (MF)
- 20% organic manure replacing chemical fertilizer (20% OF)
- 40% organic manure replacing chemical fertilizer (40% OF)
In each wheat cropping season, all treatments except CK received total N, P2O5, and K2O fertilizers at rates of 195 kg ha−1, 105 kg ha−1, and 75 kg ha−1, respectively. Urea, superphosphate, and potassium sulfate were used as chemical N, P2O5, and K2O fertilizers in the experiment. A commercial OF derived from cow dung, containing 55.9% SOM, 1.30% N, 1.55% P2O5, and 1.22% K2O, was applied in the experiment. The application amount of OF used to substitute chemical fertilizer was calculated according to N. Chemical phosphorus and potassium application rates were adjusted based on the content of OF. In this study, 50% N fertilizer (including organic and chemical N), P fertilizer, and K fertilizer were applied basally. And an additional 50% N was applied as urea at the jointing stage of wheat. During each maize cropping season, each plot, excluding CK, received equal amounts of urea (225 kg N ha−1), superphosphate (120 kg P2O5 ha−1), and potassium sulfate (135 kg K2O ha−1). All phosphate and potassium fertilizers were applied before the planting. Nitrogen fertilizer was evenly split between basal and top-dressing applications. Details of fertilizer application rates are provided in Table 1. In this study, both wheat and maize residues were returned to the field. The biomass data of wheat and maize grains and residues for each season are presented in Tables S1 and S2, respectively. Tables S3 and S4 provide the accumulations of N, P, and K in both the grain and straw of wheat and maize, respectively.
2.3. Sampling and Analysis
2.3.1. Sample Collection
The soil samples were collected during the wheat crop harvesting stage in June 2021 using a five-point sampling method, each plot yielding 500 g of soil at a depth of 0–20 cm. Within each plot (40 m2), soils at depths of 0–20 cm were collected at five sampling points, designated as X type, employing a sterile blade. These samples were combined to create a composite sample. Subsequently, visible plant roots and organic residues were meticulously removed. The samples were then divided into two parts. One subsample was placed in plastic containers and transported to the laboratory in an ice box. These plastic containers were subsequently stored at −80 °C to facilitate microbial community analysis. The second subsample was stored at 4 °C for the analysis of soil enzyme activities and soil chemical properties.
2.3.2. Soil Chemical Property
The pH, Olsen-P, and available K were determined by air-drying and sieving soil samples through an 8-mesh sieve. Total N and SOM concentrations were measured by grinding the soil samples and passing them through a 100-mesh sieve. Soil pH was measured using a pH meter (SevenExcellence, Mettler-Toledo, Shanghai, China) with a 1:2.5 soil-to-water ratio. Olsen-P and available K were determined following the Murphy and Riley [21] and Walker and Barber [22] methods, respectively. Total soil N and SOM were analyzed via wet oxidation using a Vario Max CN instrument (VarioMax CN; Elementar, Langenselbold, Germany).
2.3.3. Soil Enzyme
Urease activity was assessed by incubating samples for 2 h at 37 °C with a 10% urea solution. Next, the culture solution was shaken thoroughly for a few seconds, and ammonium in the soil suspension was quantified using a colorimetric method [23]. Alkaline phosphatase activity was determined by measuring p-nitrophenol (pNP) released from 1 g of fresh soil incubated with 0.2 mL of toluene, 4 mL of buffer solution (pH 11.0), and 1 mL of p-nitrophenyl phosphate at 37 °C for 1 h [24]. Invertase activity was determined by measuring glucose using the 3,5-dinitrosalicylic acid method with a saccharose solution as the substrate [25]. Catalase activity was measured by back titration of residual H2O2 with 0.1 M KMnO4 added to the soil [26].
2.3.4. DNA Extraction and High-Throughput Sequencing Analysis
Soil DNA extraction was performed from 0.5 g of freeze-dried soil using the E.Z.N.A. Soil DNA Kit (Omega Bio-tek, Norcross, GA, USA) following the manufacturer’s protocol. The DNA concentration and purity were assessed using a 1% agarose gel and a NanoDrop 2000 UV-vis spectrophotometer (Thermo Scientific, Wilmington, NC, USA). The V3–V4 region of the 16S rRNA gene was amplified with primer pairs 338F and 806R, while the V4 region of the 18S rRNA gene was amplified using primer pairs SSU0817F and 1196R. PCR was conducted in triplicate with an ABI GeneAmp 9700 PCR thermocycler (ABI, Vernon, CA, USA) under the following conditions: initial denaturation at 95 °C for 3 min, followed by 27 cycles (95 °C for 30 s, 55 °C for 30 s, 72 °C for 30 s), followed by a single extension at 72 °C for 10 min, and finally ending at 4 °C. After amplification, 3 μL of the PCR product underwent 2.0% agarose gel electrophoresis and subsequent purification using the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA). The PCR product’s quantity was determined using a Quantus Fluorometer (Promega, Madison, WI, USA) [27].
Purified amplicons were sequenced on an Illumina MiSeq platform (Illumina, San Diego, CA, USA) at Majorbio Bio-Pharm Technology Co., Ltd., Shanghai, China. QIIME (ver. 1.17) was used for demultiplexing, quality filtering, and processing the raw FASTQ files. Sequences with 97% similarity were assigned to the same operational taxonomic units (OTUs) with UPARSE software (ver. 7.1; http://drive5.com/uparse/; accessed on 5 August 2021). The Ribosomal Database Project (RDP) classifier was used against the Silva [28] 16S rRNA database for bacteria and the 18S rRNA database for fungi [29]. On average, each sample yielded 47,912 quality 16S sequences and 65,951 quality 18S sequences, with bacterial and fungal average bp read lengths of 418 and 243, respectively.
2.3.5. Data and Statistical Analyses
Bacterial alpha diversity, encompassing Chao1 richness, Shannon diversity, and Peilou’s evenness indices, along with PCoA based on the Bray–Curtis distance and RDA, was conducted using the “vegan” package in R version 4.0.5. Indicator species analysis for different treatments was carried out using the “indicspecies” package in R version 4.0.5. A bipartite network was constructed and visualized using the “edgeR” and “igraph” packages in R version 4.0.5. The significant differences in bacterial and fungal taxa among the fertilization treatments were identified with linear discriminant analysis of effect size (LEfSe) analysis [30]. A one-way analysis of variance (ANOVA) for each trait was performed, fitting the linear mixed model with fixed effects tested for treatment and replication in JMP®Pro version 16 (SAS, Cary, NC, USA). The Tukey HSD test was applied in comparisons with each other after ANOVA. Pearson’s two-tailed correlation test was executed using SPSS software version 25.0.
3. Results
3.1. Crop Yield
The grain yields of wheat and maize over the recent five years (2016–2021) are presented in Figure 1. Crops treated with no N fertilizer exhibited the lowest average yields over all five years. Compared with CK, the MF 20% and 40% treatments significantly increased crop yields of wheat by 131.25%, 125%, and 128.13%, and maize by 50.28%, 49.58%, and 50%, respectively.
3.2. Soil Chemical Properties
The soil TN and SOM contents in the CK treatment were consistently lower than in the other treatments (Table 2). Compared to CK, soil TN and SOM content significantly increased in the 20% and 40% OF substitution treatments by 23.53% and 29.41% for TN and 18.18% for SOM in both treatments, respectively (p < 0.05). The content of AK and AP showed no significant differences under different treatments. Soil pH decreased significantly by 0.1 units in all fertilization treatments compared to CK (p < 0.05).
3.3. Soil Enzyme Activity
The impact of chemical fertilizers and different organic manure substitution ratios on soil microbial function and activity was assessed by measuring the activities of four soil extracellular enzymes: urease, alkaline phosphatase, invertase, and catalase (Figure 2). Alkaline phosphatase activity was significantly higher than CK under all fertilization treatments, with increases of 17.24%, 21.31%, and 17.95%, respectively (p < 0.05). The 20% OF substitution treatment significantly increased activities for urease, alkaline phosphatase, and invertase, with significant increases of 22.57%, 27.08%, and 22.78%, respectively, compared to CK (p < 0.05). Catalase activity did not significantly differ across treatments compared to CK.
3.4. Soil Bacterial and Fungal Community Structure and Diversity
Bacterial diversity increased significantly by 3.68%, 4.50%, and 4.07%, respectively (p < 0.05). Significant increases in bacterial richness were observed under the treatment of 40% OF substitution (9.84%) compared to CK (Figure 3). The treatment with 40% OF substitution exhibited the highest bacterial richness (679.23), while the 20% OF substitution treatment showed the highest diversity (5.0) and evenness (0.64). Fungal richness, diversity, and evenness were consistently lower than those of bacteria. Chemical fertilizer treatment and 20% OF substitution treatment led to significant increases in fungal richness (15.72% and 14.24%, respectively) compared to CK. No significant differences were observed in fungal diversity or evenness among the different fertilization treatments.
The Principal Coordinates Analysis (PCoA) accounted for 49.0% of bacterial community variation (Figure 4A) and 42.3% of fungal community variation (Figure 4C). CK’s bacterial community was distinctly separated from the bacterial and fungal communities under various fertilization treatments. In redundancy analysis (RDA), environmental factors explained 67.19% of bacterial community variation and 25.5% of fungal community variation. RDA results revealed that pH was negatively correlated with TN and SOM. Furthermore, pH, TN, and SOM primarily drove bacterial community variations between CK and different fertilization treatments (Figure 4B), while SOM and TN were key factors for fungal community variations (Figure 4D).
At the phylum level, significant variations in the relative abundance of dominant bacterial and fungal species were noted under different fertilization treatments. The dominant bacterial phyla, Actinobacteriota, Acidobacteriota, Proteobacteria, and Chloroflexi, exhibited relative abundances ranging between 21.77–27.51%, 18.16–21.43%, 10.55–11.47%, and 16.71–26.90%, respectively (Figure 5A). Under the 20% OF substitution treatment, Actinobacteriota and Proteobacteria showed the highest relative abundances, increasing by 21.0% and 15.25%, respectively, compared to CK, while Acidobacteriota decreased by 37.04% compared to CK. Ascomycota and Mortierellomycota were the dominant fungal phyla across all samples (Figure 5B), with relative abundances ranging between 70.73–82.18% and 8.33–12.55%, respectively. Compared to CK, Ascomycota’s relative abundance increased by 13.42%, 10.26%, and 11.25% under the sole chemical fertilizer, 20% OF substitution, and 40% OF substitution treatments, respectively.
3.5. Soil Bacterial Bipartite Networks
The bipartite network showed shared bacterial indicator OTUs between chemical fertilizer, 20% OF substitution, and 40% OF substitution treatments, mostly from the Proteobacteria phylum (Figure 6A). This partially explains the differences in bacterial community structures between CK and fertilized treatments. More shared bacterial indicator OTUs between 20% and 40% OF substitution treatments belonged to Actinobacteriota, Proteobacteriota, and Firmicutes phyla. Correlation analysis (Figure 6B) revealed positive correlations between Actinobacteriota, Proteobacteriota, Firmicutes, and soil TN while negatively correlating with pH.
The bipartite network showed significant fungal indicator OTU overlap among chemical fertilizer, 20% OF substitution, and 40% OF substitution treatments, mainly from the Ascomycota phylum (Figure 7A). Correlation analysis indicated positive correlations between Ascomycota and Basidiomycota phyla indicator OTUs with AP, SOM, and TN, respectively (Figure 7B). However, relative abundance differences among treatments were not significant.
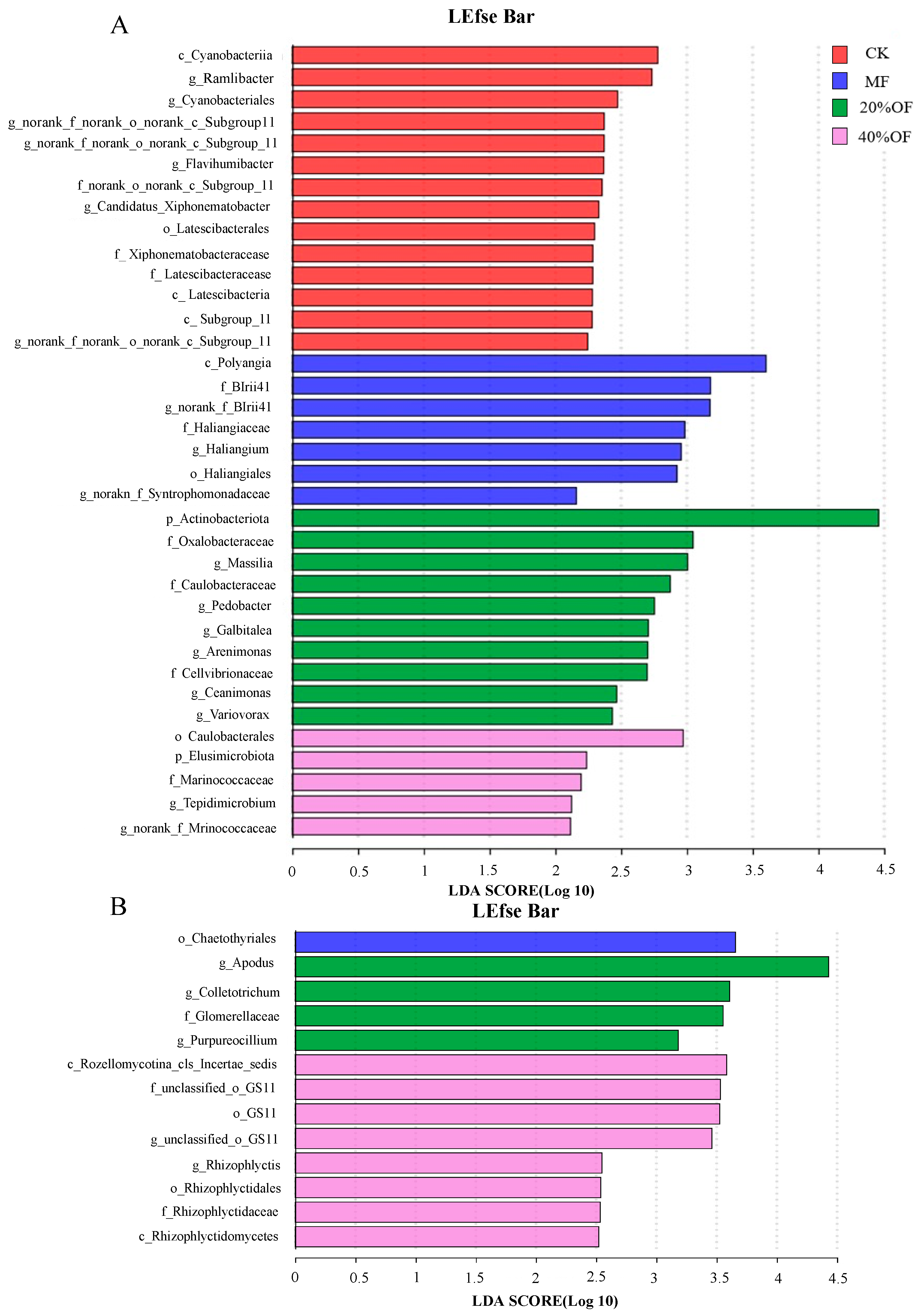
3.6. LEfSe
Using the linear discriminant analysis (LDA) effect size method, differentiation of bacterial and fungal taxa was achieved across the treatments: no fertilization, chemical fertilizer application, 20% OF substitution, and 40% OF substitution (Figure 8A). Within the four treatments at various taxonomic levels, six bacterial biomarkers with an LDA effect size > 3 were identified. Chemical fertilizer treatment exhibited distinctive taxa, including the Polyangia class, Blrii41 family, and no-rank genus. Under the 20% OF substitution treatment, Actinobacteriota phylum, Oxalobacteraceae family, and Massilia genus were predominant biomarkers.
Nine fungal taxa with an LDA effect size > 3 were identified to differentiate among the no fertilization, chemical fertilizer application, 20% OF substitution, and 40% OF substitution treatments (Figure 8B). Chemical fertilizer treatment featured the Chaetothyriales order as the most abundant and distinctive taxon. In the 20% OF substitution treatment, the Glomerellaceae family, along with the genera Apodus, Colletotrichum, and Purpureocillium, were the most abundant. The 40% OF substitution treatment displayed the highest abundance of the Rozellomycotina_cls_Incertae_sedis class and the GS11 order.
4. Discussion
4.1. Crop Yield and Soil Chemical Properties
Organic fertilizer substitution for chemical fertilizer has demonstrated synergistic benefits in terms of crop yields, sustainability, and soil fertility [31,32]. However, due to the slow release of nutrients and inefficiency of organic fertilizers, high ratios of organic to chemical fertilizer substitution can often lead to a reduction in crop yield [33]. This study found no significant yield difference between sole chemical and organic fertilizer application treatments, indicating that up to a 40% substitution with organic fertilizer can sustain high crop yields. This can be attributed to the pH regulation of the soil [34] and the improved nutrient efficiency of organic manure. When comparing soil fertility under 20% and 40% organic manure substitution treatments with unfertilized soil, our findings demonstrate that both treatments significantly increased SOM and TN contents, as shown in Table 2. It is noteworthy that a higher proportion of organic manure substitution does not necessarily lead to improved fertilizer efficacy. Remarkably, the 20% OF substitution exhibited a pronounced increase in soil fertility, emerging as the optimal level for enhancing crop nutrient assimilation and yield optimization. OF undergoes microbial decomposition, transforming into humus, which can react with sodium carbonate to form sodium humate. Consequently, organic manure substitution helps mitigate soil alkalinity, as outlined in Table 2.
4.2. Soil Enzyme Activity
Soil enzymes, including urease and alkaline phosphatase, play pivotal roles in soil carbon (C), nitrogen (N), and phosphorus (P) cycling. They are positively correlated with organic matter content and intricately involved in SOM tr ansformation [9,35,36,37]. Compared to the control, 20% OF substitution elevates C and N levels, boosting microbial metabolic activity and providing organic matter as a substrate for soil enzymes [38]. Consequently, urease, alkaline phosphatase, and invertase activities significantly increase [39]. However, using over 40% OF substitution may reduce available N, curtailing soil enzyme substrate use efficiency [16].
4.3. Soil Bacterial and Fungal Community Diversity and Structure
Soil microbial communities serve as reliable indicators of shifts in soil quality [40,41]. A richer microbial diversity within soil is widely believed to correlate with a more resilient ecosystem [42]. The introduction of chemical fertilizers and organic manure provides the energy and substrates needed to bolster microbial activity. These interventions profoundly stimulate soil prokaryote proliferation [1] and enhance the abundance, diversity, and evenness of bacterial communities (Figure 3). This observation aligns with the findings presented by Cui et al. [14]. However, when comparing the effects of substituting chemical fertilizers with organic ones (at levels < 40%), the impact on fungal communities appears less pronounced than on bacterial communities (Figure 3). This underscores the fact that bacteria are more sensitive to environmental fluctuations than fungi.
4.4. Soil Bacterial and Fungal Community Structure
In contrast to the CK, both chemical fertilizers and organic manure significantly affect soil microbial community structures (Figure 4A,C). Changes in bacterial community composition are closely associated with soil pH, SOM, and TN (Figure 4B), while shifts in fungal communities are primarily linked to soil SOM and TN (Figure 4D). It is well-documented that soil bacterial community composition exhibits a stronger correlation with soil pH compared to fungal communities [43,44,45,46]. Conversely, soil nutrients emerge as the primary determinants shaping fungal communities [47,48]. The application of chemical fertilizers and OF substitution releases nutrients, notably organic matter and effective phosphorus, diminishing the diversity of oligotrophic bacterial groups like Acidobacteriota (within the Acidobacteria phylum). Simultaneously, there is an enhancement in the diversity of eutrophic groups from the Actinobacteria and Ascomycetes phyla, which are proficient at catalyzing the decomposition of complex macromolecules, including organic matter (Figure 5) [49,50]. This finding is consistent with observations by Guo et al. [51]. In the context of organic manure substitution, both the 20% and 40% levels promote bacterial OTUs of Actinobactreiota, Proteobacteriota, and Firmicutes. Notably, Actinomycetes have the capacity to suppress soil pathogens [52]. Furthermore, the synergistic interaction between Proteobacteriota and Firmicutes facilitates the formation of multiphase biofilms in the rhizosphere, subsequently activating plant disease resistance mechanisms [53]. Therefore, in comparison to the sole use of chemical fertilizer, the 20% and 40% organic manure substitutions potentially enhance the enrichment of beneficial bacteria, thereby inhibiting soil pathogens and promoting a more balanced microbial community structure.
Compared to the CK, an increase in the relative abundances of the Polyangia phylum and the Haliangium family was observed in LDA when chemical fertilizer was applied. Haliangium, recognized as a biocontrol bacterium, produces haliangicin, inhibiting the growth of a broad spectrum of fungi and protecting plants from soil pathogens. However, it is important to note that their relative abundance tends to decrease as the cropping year progresses [54].
Conversely, in treatments with a 20% organic manure substitution in addition to chemical fertilizer, there was a significant proliferation of the Actinobateriota phylum, Oxalobacteraceae family, and Massilia genus. Many species within the Oxalobacteraceae family have been identified as effective agents for promoting plant growth and enhancing nitrogen uptake [52]. Additionally, Massilia, predominantly found in rhizosphere soil, possesses the capacity for phosphorus solubilization [55,56].
In soils treated with both chemical and organic fertilizers, a significant increase in the relative abundance of Ascomycota was observed. Ascomycota is the most widespread phylum among eukaryotes, playing a crucial role in decomposing organic substrates, including leaf litter, wood, and manure [57]. Notably, most fungal communities shared between soils treated with chemical fertilizers and those with organic manure substitutions were found to belong to Ascomycota, which was consistent with prior studies [51,55]. However, it is worth noting that a higher relative abundance of Ascomycota does not necessarily indicate improved soil health. On a positive note, in soils with a 20% organic manure substitution, there was a significant rise in the abundance of Mortierellomycota. Mortierellomycota are recognized as phosphate-solubilizing fungi, and an increase in their relative abundance may serve as an indicator of enhanced soil health [58].
From the LDA, an increase in the relative abundance of Cheatothyriales was observed due to the application of chemical fertilizers. Some fungi within Cheatothyriales are known pathogens that adhere to leaf and stem surfaces, obtaining nutrients from sugary exudates. Long-term mineral fertilizer application without an additional carbon source has been shown to alter fungal communities [59] and weaken the plant-microbial network [60]. Higher network connectivity and key microbial taxa abundance may be associated with increased soil suppressiveness [61], potentially increasing the relative abundance of certain plant pathogenic genera [62,63]. Application of OF can enhance microbe-eukaryote connections more than chemical fertilizers [64] and produce potential biocontrol agents [65]. OF can serve as an alternative carbon source for antagonists [66,67]. Notably, a 20% OF substitution increased the abundance of Purpureocillium, which inhibits various plant pathogens and viruses [68], as well as the relative abundance of Colltotrichum in the Glomerellaceae family, often associated with crop residue decay in the soil. Although some fungi within this genus are plant pathogens [69], their relative abundance remains very low (all < 0.1%). The antagonistic activity against plant pathogens depends on specific ecological conditions and may not occur simultaneously but rather be observed over time through frequent sampling [70]. In the 40% OF substitution treatment, the most abundant class was Rozellomycotina_cls_Incertae_sedis. Studies have indicated a positive correlation between Rozellomycotina_cls_Incertae_sedis and soil nitrate and ammonium content [71]. Differential fungal species analysis under 20% and 40% OF substitution levels also suggests that the impact of OF on fungal communities is less pronounced compared to bacterial communities [72,73].
5. Conclusions
In this study, the impact of varying organic-to-inorganic fertilizer ratios on essential microbial functions and structural characteristics of calcareous alluvial soil was compared. Up to a 40% substitution with organic fertilizer can sustain high crop yields. While some research suggests that OF application can enhance soil structure, higher proportions may not always yield better results. Notably, at the 20% OF ratio, the chemical properties of the soil, soil enzyme activity, and microbial community structure were significantly improved when compared to a 40% OF ratio. Different levels of OF substitution significantly increased the α-diversity of soil bacterial communities. Compared to the exclusive application of chemical fertilizers, both the 20% and 40% OF substitution levels enriched the beneficial soil microbes, inhibited soil-borne pathogens, and rebalanced the microbial community structure, thereby fostering plant growth and development. Future studies could explore the suitability of different organic-to-inorganic fertilizer ratios across different soil types. Additionally, assessing the environmental impact and economic feasibility of implementing such fertilizer ratios on a larger scale could provide valuable insights for sustainable agricultural practices.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14050888/s1. Table S1: Effect of organic fertilizer substitution on grain biomass, straw biomass, and harvest indexes of wheat from 2017 to 2021. Values are means of triplicate samples and are not significantly different at p < 0.05 when followed by the same lowercase letters. Table S2: Effect of organic fertilizer substitution on grain biomass, straw biomass, and harvest indexes of maize from 2017 to 2020. Values are means of triplicate samples and are not significantly different at p < 0.05 when followed by the same lowercase letters. Table S3: Effect of organic fertilizer substitution on the N, P, and K accumulation of wheat from 2017 to 2021. Values are means of triplicate samples and are not significantly different at p < 0.05 when followed by the same lowercase letters. Table S4: Effect of organic fertilizer substitution on the N, P, and K accumulation of maize from 2017 to 2020. Values are means of triplicate samples and are not significantly different at p < 0.05 when followed by the same lowercase letters. Figure S1: Map of the study site.
Author Contributions
Conceptualization, Y.L.; formal analysis, J.S. and H.Z.; investigation, Y.X., Y.Y., J.S., X.G., T.S., H.Z. and Y.W. (Yafei Wang); methodology, L.J. and Y.L.; resources, Y.X., Y.Y., X.G., H.Z. and Y.W. (Yafei Wang); supervision, L.J. and Y.L.; validation, Y.L.; writing—original draft, Y.W. (Yijun Wang) and Y.X.; writing—review and editing, Y.W. (Yijun Wang) and Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Science Foundation of Shandong Province of China (ZR202103050385), the National Natural Science Foundation of China (32102486), the National Key Research and Development Projects (2021YFD1900903), the Taishan Industrial Leading Talents Project of Shandong Province (LJNY202124), and the Agricultural Science and Technology Innovation Project of Shandong Academy of Agricultural Sciences (GXGC2023F03).
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhong, W.H.; Gu, T.; Wang, W.; Zhang, B.; Lin, X.G.; Huang, Q.R.; Shen, W.S. The effects of mineral fertilizer and organic manure on soil microbial community and diversity. Plant Soil 2010, 326, 511–522. [Google Scholar] [CrossRef]
- Francioli, D.; Schulz, E.; Lentendu, G.; Wubet, T.; Buscot, F.; Reitz, T. Mineral vs. organic amendments: Microbial community structure, activity and abundance of agriculturally relevant microbes are driven by long-term fertilization strategies. Front. Microbiol. 2016, 7, 1446. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Jeong, S.T.; Das, S.; Kim, P.J. Composted cattle manure increases microbial activity and soil fertility more than composted swine manure in a submerged rice paddy. Front. Microbiol. 2017, 8, 1702. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, B.; Xia, L.L.; Fan, C.H.; Xiong, Z.Q. Organic-substitute strategies reduced carbon and reactive nitrogen footprints and gained net ecosystem economic benefit for intensive vegetable production. J. Clean. Prod. 2019, 225, 984–994. [Google Scholar] [CrossRef]
- Xia, L.L.; Lam, S.K.; Chen, D.L.; Wang, J.Y.; Tang, Q.; Yan, X.Y. Can knowledge-based N management produce more staple grain with lower greenhouse gas emission and reactive nitrogen pollution? A meta-analysis. Glob. Chang. Biol. 2017, 23, 1917–1925. [Google Scholar] [CrossRef] [PubMed]
- Karlen, D.L.; Mausbach, M.J.; Doran, J.W.; Cline, R.G.; Harris, R.F.; Schuman, G.E. Soil quality: A concept, definition, and framework for evaluation. Soil Sci. Soc. Am. J. 1997, 61, 4–10. [Google Scholar] [CrossRef]
- Doran, J.W.; Zeiss, M.R. Soil health and sustainability: Managing the biotic component of soil quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, R.F.; Xue, C.; Xun, W.B.; Sun, L.; Xu, Y.C.; Shen, Q.R. Pyrosequencing reveals contrasting soil bacterial diversity and community structure of two main winter wheat cropping systems in China. Microb. Ecol. 2014, 67, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.C.; Li, K.J.; Zhou, W.; Qiu, S.J.; Huang, S.W.; He, P. Changes in soil microbial community, enzyme activities and organic matter fractions under long-term straw return in north-central China. Agric. Ecosyst. Environ. 2016, 216, 82–88. [Google Scholar] [CrossRef]
- Paterson, E.; Sim, A.; Osborne, S.M.; Murray, P.J. Long-term exclusion of plant-inputs to soil reduces the functional capacity of microbial communities to mineralise recalcitrant root-derived carbon sources. Soil Biol. Biochem. 2011, 43, 1873–1880. [Google Scholar] [CrossRef]
- Wu, G.H.; Chen, Z.H.; Jiang, D.Q.; Jiang, N.; Jiang, H.; Chen, L.J. Oxidases and hydrolases mediate soil organic matter accumulation in chernozem of northeastern China. Geoderma 2021, 403, 115206. [Google Scholar] [CrossRef]
- Zhou, J.; Guan, D.W.; Zhou, B.K.; Zhao, B.S.; Ma, M.C.; Qin, J.; Jiang, X.; Chen, S.F.; Cao, F.M.; Shen, D.L.; et al. Influence of 34-years of fertilization on bacterial communities in an intensively cultivated black soil in northeast China. Soil Biol. Biochem. 2015, 90, 42–51. [Google Scholar] [CrossRef]
- Pan, H.; Chen, M.M.; Feng, H.J.; Wei, M.; Song, F.P.; Lou, Y.H.; Cui, X.M.; Wang, H.; Zhuge, Y.P. Organic and inorganic fertilizers respectively drive bacterial and fungal community compositions in a fluvo-aquic soil in northern China. Soil Till. Res. 2020, 198, 104540. [Google Scholar] [CrossRef]
- Cui, X.W.; Zhang, Y.Z.; Gao, J.S.; Peng, F.Y.; Gao, P. Long-term combined application of manure and chemical fertilizer sustained higher nutrient status and rhizospheric bacterial diversity in reddish paddy soil of Central South China. Sci. Rep. 2018, 8, 16554. [Google Scholar] [CrossRef] [PubMed]
- Sarathchandra, S.U.; Ghani, A.; Yeates, G.W.; Burch, G.; Cox, N.R. Effect of nitrogen and phosphate fertilisers on microbial and nematode diversity in pasture soils. Soil Biol. Biochem. 2001, 33, 952–964. [Google Scholar] [CrossRef]
- Cui, J.W.; Zhu, R.L.; Wang, X.Y.; Xu, X.P.; Ai, C.; He, P.; Liang, G.Q.; Zhou, W.; Zhu, P. Effect of high soil C/N ratio and nitrogen limitation caused by the long-term combined organic-inorganic fertilization on the soil microbial community structure and its dominated SOC decomposition. J. Environ. Manag. 2022, 303, 114155. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.M.; Li, C.; Xiao, X.P.; Shi, L.H.; Cheng, K.K.; Wen, L.; Li, W.Y. Effect of short-term manure nitrogen input on soil microbial community structure and diversity in a double-cropping paddy field. Sci. Rep. 2020, 10, 13540. [Google Scholar] [CrossRef]
- Yang, Y.J.; Lei, T.; Du, W.; Liang, C.L.; Li, H.D.; Lv, J.L. Substituting chemical fertilizer nitrogen with organic manure and comparing their nitrogen use efficiency and winter wheat yield. J. Agric. Sci. 2020, 158, 260–268. [Google Scholar] [CrossRef]
- Chesworth, W.; Camps, M.A.; Macías, F.; Spaargaren, O.; Spaargaren, O.; Mualem, Y.; Micheli, E. Classification of soils: World Reference Base (WRB) for soil resources. In Encyclopedia of Soil Science, 2008th ed.; Chesworth, W., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 120–122. [Google Scholar]
- Murphy, J.; Riley, J. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Walker, J.M.; Barber, S.A. Absorption of potassium and rubidium from the soil by corn roots. Plant Soil 1962, 17, 243–259. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Liu, S.B.; Wang, J.Y.; Pu, S.Y.; Blagodatskaya, E.; Kuzyakov, Y.; Razavi, B.S. Impact of manure on soil biochemical properties: A global synthesis. Sci. Total Environ. 2020, 745, 141003. [Google Scholar] [CrossRef] [PubMed]
- Gopal, M.; Gupta, A.; Arunachalam, V.; Magu, S.P. Impact of azadirachtin, an insecticidal allelochemical from neem on soil microflora, enzyme and respiratory activities. Bioresour. Technol. 2007, 98, 3154–3158. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, G.; Xue, S.; Song, Z. Rhizosphere soil microbial activity under different vegetation types on the Loess Plateau, China. Geoderma 2011, 161, 115–125. [Google Scholar] [CrossRef]
- Wang, B.; Chen, C.; Xiao, Y.M.; Chen, K.Y.; Wang, J.; Zhao, S.; Liu, N.; Li, J.N.; Zhou, G.Y. Trophic relationships between protists and bacteria and fungi drive the biogeography of rhizosphere soil microbial community and impact plant physiological and ecological functions. Microb. Res. 2024, 280, 127603. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Amato, K.R.; Yeoman, C.J.; Kent, A.; Righini, N.; Carbonero, F.; Estrada, A.; Gaskins, H.R.; Stumpf, R.M.; Yildirim, S.; Torralba, M.; et al. Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes. ISME J. 2013, 7, 1344–1353. [Google Scholar] [CrossRef]
- Fan, T.L.; Stewart, B.A.; Wang, Y.; Luo, J.J.; Zhou, G.Y. Long-term fertilization effects on grain yield, water-use efficiency and soil fertility in the dryland of Loess Plateau in China. Agric. Ecosyst. Environ. 2005, 106, 313–329. [Google Scholar] [CrossRef]
- Garcíamantrana, I.; Selmaroyo, M.; Alcántara-Baena, C.; Collado, M.C. Shifts on gut microbiota associated to mediterranean diet adherence and specific dietary intakes on general adult population. Front. Microbiol. 2018, 9, 890. [Google Scholar]
- Urkurkar, J.S.; Tiwari, A.; Chitale, S.; Bajpai, R.K. Influence of longterm use of inorganic and organic manures on soil fertility and sustainable productivity of rice (Oryza sativa) and wheat (Triticum aestivum) in inceptisols. Indian J. Agric. Sci. 2010, 80, 208–212. [Google Scholar]
- Li, Y.; Wu, X.P.; He Gang Wang, Z.H. Benefits of yield, environment and economy from substituting fertilizer by manure for wheat production of China. Sci. Agric. Sin. 2020, 53, 4879–4890. (In Chinese) [Google Scholar]
- Liu, X.; Chen, Q.; Zhang, H.C.; Zhang, J.; Chen, Y.T.; Yao, F.C.; Chen, Y.T. Effects of exogenous organic matter addition on agricultural soil microbial communities and relevant enzyme activities in southern China. Sci. Rep. 2023, 13, 8045. [Google Scholar] [CrossRef]
- Caravaca, F.; Masciandaro, G.; Ceccanti, B. Land use in relation to soil chemical and biochemical properties in a semiarid Mediterranean environment. Soil Till. Res. 2002, 68, 22–30. [Google Scholar] [CrossRef]
- Saha, S.; Mina, B.L.; Gopinath, K.A.; Kundu, S.; Gupta, H.S. Organic amendments affect biochemical properties of a subtemperate soil of the Indian Himalayas. Nutr. Cycl. Agroecosyst. 2008, 80, 233–242. [Google Scholar] [CrossRef]
- Mendham, D.S.; O’Connell, A.M.; Grove, T.S.; Rance, S.J. Residue management effects on soil carbon and nutrient contents and growth of second rotation eucalypts. Forest Ecol. Manag. 2003, 181, 357–372. [Google Scholar] [CrossRef]
- Liu, Y.M.; Cao, W.Q.; Chen, X.X.; Yu, B.G.; Lang, M.; Chen, X.P.; Zou, C.Q. The responses of soil enzyme activities, microbial biomass and microbial community structure to nine years of varied zinc application rates. Sci. Total Environ. 2020, 737, 140245. [Google Scholar] [CrossRef]
- Jin, K.; Sleutel, S.; Buchan, D.; Neve, S.D.; Cai, D.X.; Gabriels, D.; Jin, J.Y. Changes of soil enzyme activities under different tillage practices in the Chinese Loess plateau. Soil Till. Res. 2009, 104, 115–120. [Google Scholar] [CrossRef]
- Gregorich, E.G.; Carter, M.R.; Angers, D.A.; Monreal, C.M.; Ellert, B.H. Towards a minimum data set to assess soil organic matter quality in agricultural soils. Can. J. Soil Sci. 1994, 74, 367–385. [Google Scholar] [CrossRef]
- Souza, E.D.; Costa, S.E.V.G.A.; Anghinoni, I.; Carneiro, M.A.C.; Martins, A.P.; Bayer, C. Soil quality indicators in a Rhodic Paleudult under long term tillage systems. Soil Till. Res. 2014, 139, 28–36. [Google Scholar] [CrossRef]
- Chaer, G.; Fernandes, M.; Myrold, D.; Bottomley, P. Comparative resistance and resilience of soil microbial communities and enzyme activities in adjacent native forest and agricultural soils. Microb. Ecol. 2009, 58, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, S.N.; Waite, I.S.; Blackburn, A.; Husband, R.; Rushton, S.P.; Manning, D.C.; O’Donnell, A.G. Actinobacterial community dynamics in long term managed grasslands. Antonie Van Leeuwenhoek 2009, 95, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community composition at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Guo, Q.K.; Liu, D.H.; Hu, C.; Sun, J.W.; Wang, X.B.; Liang, G.Q.; Zhou, W. Composition, predicted functions, and co-occurrence networks of fungal and bacterial communities_links to soil organic carbon under long-term fertilization in a rice-wheat cropping system. Eur. J. Soil Biol. 2020, 100, 103226. [Google Scholar] [CrossRef]
- Gong, W.; Yan, X.Y.; Wang, J.Y.; Hu, T.X.; Gong, Y.B. Long-term manure and fertilizer effects on soil organic matter fractions and microbes under a wheat–maize cropping system in northern China. Geoderma 2009, 149, 318–324. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Jacquiod, S.; Franqueville, L.; Cecillon, S.; Vogel, T.M.; Simonet, P. Soil bacterial community shifts after chitin enrichment: An integrative metagenomic approach. PLoS ONE 2013, 8, e79699. [Google Scholar] [CrossRef]
- Guo, J.J.; Liu, W.B.; Zhu, C.; Luo, G.W.; Kong, Y.L.; Ling, N.; Wang, M.; Dai, J.Y.; Shen, Q.R.; Guo, S.W. Bacterial rather than fungal community composition is associated with microbial activities and nutrient-use efficiencies in a paddy soil with short-term organic amendments. Plant Soil 2018, 424, 335–349. [Google Scholar] [CrossRef]
- Pang, G.; Cai, F.; Li, R.X.; Zhao, Z.; Li, R.; Gu, X.L.; Shen, Q.R.; Chen, W. Trichoderma-enriched organic fertilizer can mitigate microbiome degeneration of monocropped soil to maintain better plant growth. Plant Soil 2017, 416, 181–192. [Google Scholar] [CrossRef]
- Lee, S.M.; Kong, H.G.; Song, G.C.; Ryu, C.M. Disruption of Firmicutes and Actinobacteria abundance in tomato rhizosphere causes the incidence of bacterial wilt disease. ISME J. 2021, 15, 330–347. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.C.; Fu, W.J.; Hu, C.W.; Chen, G.Q.; Xiao, Z.W.; Chen, Y.R.; Wang, Z.J.; Cheng, H.Y. Variation of rhizosphere microbial community in continuous mono-maize seed production. Sci. Rep. 2021, 11, 1544. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.X.; Bi, Q.F.; Hao, X.L.; Zhou, G.W.; Yang, X.R. Massilia phosphatilytica sp. nov., a phosphate solubilizing bacteria isolated from a long-term fertilized soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 2514–2519. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, M.; Suarez, C.; Steffens, D.; Ratering, S.; Schnell, S. Effect of different soil phosphate sources on the active bacterial microbiota is greater in the rhizosphere than in the endorhiza of barley (Hordeum vulgare L.). Microb. Ecol. 2019, 77, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Leite, M.F.A.; Pan, Y.; Bloem, J.; ten Berge, H.; Kuramae, E.E. Organic nitrogen rearranges both structure and activity of the soil-borne microbial seedbank. Sci. Rep. 2017, 7, 42634. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Wen, T.; Zhang, H.; Zhao, M.L.; Penton, C.R.; Thomashow, L.S.; Shen, Q.R. Predicting disease occurrence with high accuracy based on soil macroecological patterns of Fusarium wilt. ISME J. 2020, 14, 2936–2950. [Google Scholar] [CrossRef] [PubMed]
- Paungfoo-Lonhienne, C.; Yeoh, Y.K.; Kasinadhuni, N.R.P.; Lonhienne, T.G.; Robinson, N.; Hugenholtz, P.; Ragan, M.A.; Schmidt, S. Nitrogen fertilizer dose alters fungal communities in sugarcane soil and rhizosphere. Sci. Rep. 2015, 5, 8678. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.L.; Hu, W.J.; Tao, J.; Liu, Y.Z.; Kong, Z.Y.; Wu, L. Soil bacterial community structure and extracellular enzyme activities under different land use types in a long-term reclaimed wetland. J. Soils Sediments 2019, 19, 2543–2557. [Google Scholar] [CrossRef]
- Banerjee, S.; Walder, F.; Büchi, L.; Meyer, M.; Held, A.Y.; Gattinger, A.; Keller, T.; Charles, R.; van der Heijden, M.G.A. Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa in roots. ISME J. 2019, 13, 1722–1736. [Google Scholar] [CrossRef]
- van Bruggen, A.H.C.; Finckh, M.R. Plant diseases and management approaches in organic farming systems. Annu. Rev. Phytopathol. 2016, 54, 25–54. [Google Scholar] [CrossRef]
- Berg, M.; Koskella, B. Nutrient-and dose-dependent microbiome-mediated protection against a plant pathogen. Curr. Biol. 2018, 28, 2487–2492. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, A.K.A.; Harkes, P.; van den Elsen, S.; Holterman, M.; Korthals, G.W.; Helder, J.; Kuramae, E.E. Organic amendment strengthens interkingdom associations in the soil and rhizosphere of barley (Hordeum vulgare). Sci. Total Environ. 2019, 695, 133885. [Google Scholar] [CrossRef]
- Lewis, J.A.; Fravel, D.R.; Papavizas, G.C. Cladorrhinum foecundissimum- a potential biological control agent for the reduction of Rhizoctonia solani. Soil Biol. Biochem. 1995, 27, 863–869. [Google Scholar] [CrossRef]
- Bonanomi, G.; Antignani, V.; Capodilupo, M.; Scala, F. Identifying the characteristics of organic soil amendments that suppress soilborne plant diseases. Soil Biol. Biochem. 2010, 42, 136–144. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Abdelnabby, H.; Xiao, Y. The role of a phospholipase (PLD) in virulence of Purpureocillium lilacinum (Paecilomyces lilacinum). Microb. Pathog. 2015, 85, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Talhinhas, P.; Baroncelli, R. Colletotrichum species and complexes: Geographic distribution, host range and conservation status. Fungal Divers. 2021, 110, 109–198. [Google Scholar] [CrossRef]
- Semenov, M.V.; Krasnov, G.S.; Semenov, V.M.; van Bruggen, A. Mineral and organic fertilizers distinctly affect fungal communities in the crop rhizosphere. J. Fungi 2022, 8, 251. [Google Scholar] [CrossRef]
- Wang, J.Q.; Shi, X.Z.; Zheng, C.Y.; Suter, H.; Huang, Z.Q. Different responses of soil bacterial and fungal communities to nitrogendeposition in a subtropical forest. Sci. Total Environ. 2021, 755, 142449. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, J.N.; Lu, M.; Qin, C.; Chen, Y.H.; Yang, L.; Huang, Q.W.; Wang, J.C.; Shen, Z.G.; Shen, Q.R. Microbial communities of an arable soil treated for 8 years with organic and inorganic fertilizers. Biol. Fertil. Soils 2016, 52, 455–467. [Google Scholar] [CrossRef]
- van Diepeningen, A.D.; de Vos, O.J.; Zelenev, W.; Semenov, A.M.; van Bruggen, A.H.C. DGGE fragments oscillate with or counter to fluctuations in cultivable bacteria along wheat roots. Microb. Ecol. 2005, 50, 506–517. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of OF substitution on the average grain yield of wheat (A) and maize (B) during the growing seasons. Values are means of triplicate samples and are not significantly different at p < 0.05 when followed by the same lowercase letters. The error bars represent standard deviations.
Figure 1.
Effect of OF substitution on the average grain yield of wheat (A) and maize (B) during the growing seasons. Values are means of triplicate samples and are not significantly different at p < 0.05 when followed by the same lowercase letters. The error bars represent standard deviations.
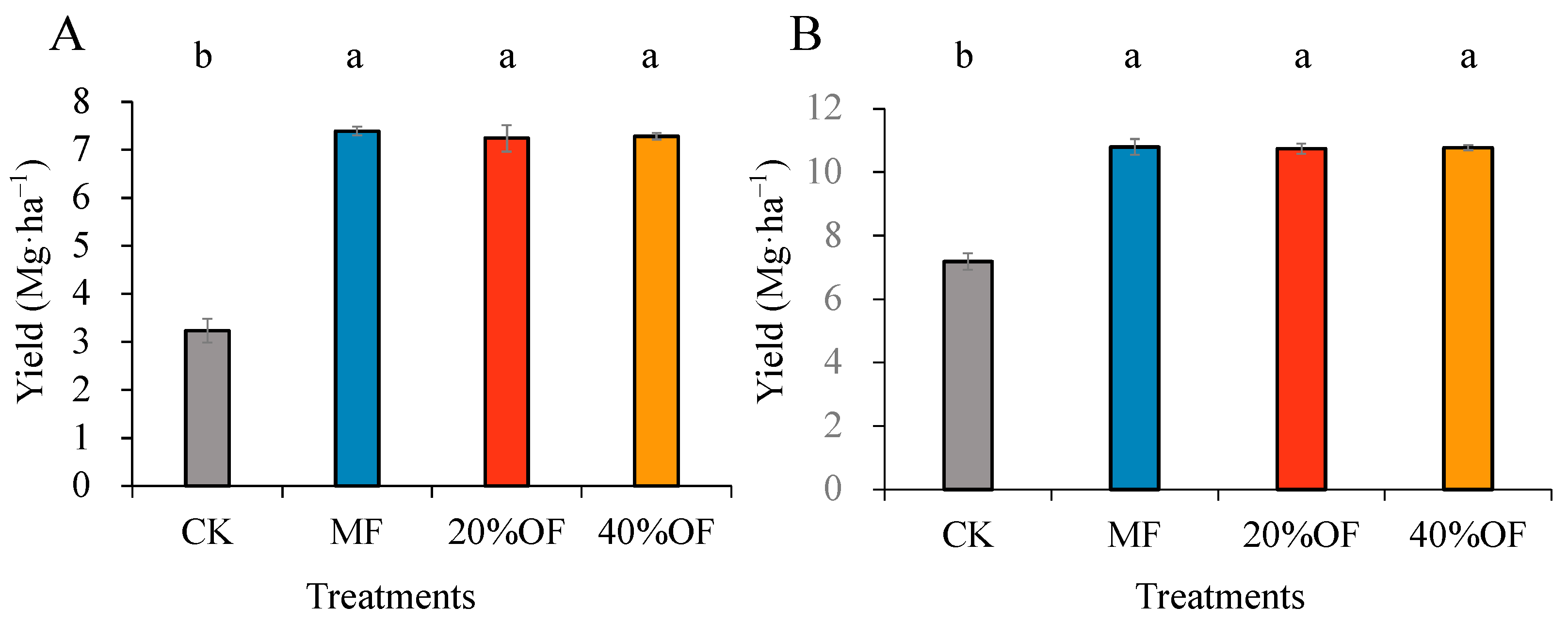
Figure 2.
Soil enzyme activities under different organic substitution ratios. The same lowercase letters are not significantly different at p < 0.05. The error bars represent standard deviations.
Figure 2.
Soil enzyme activities under different organic substitution ratios. The same lowercase letters are not significantly different at p < 0.05. The error bars represent standard deviations.

Figure 3.
Soil bacterial (A) and fungal (B) alpha-diversity under different organic substitution ratios. The same lowercase letters are not significantly different at p < 0.05. The error bars represent standard deviations.
Figure 3.
Soil bacterial (A) and fungal (B) alpha-diversity under different organic substitution ratios. The same lowercase letters are not significantly different at p < 0.05. The error bars represent standard deviations.
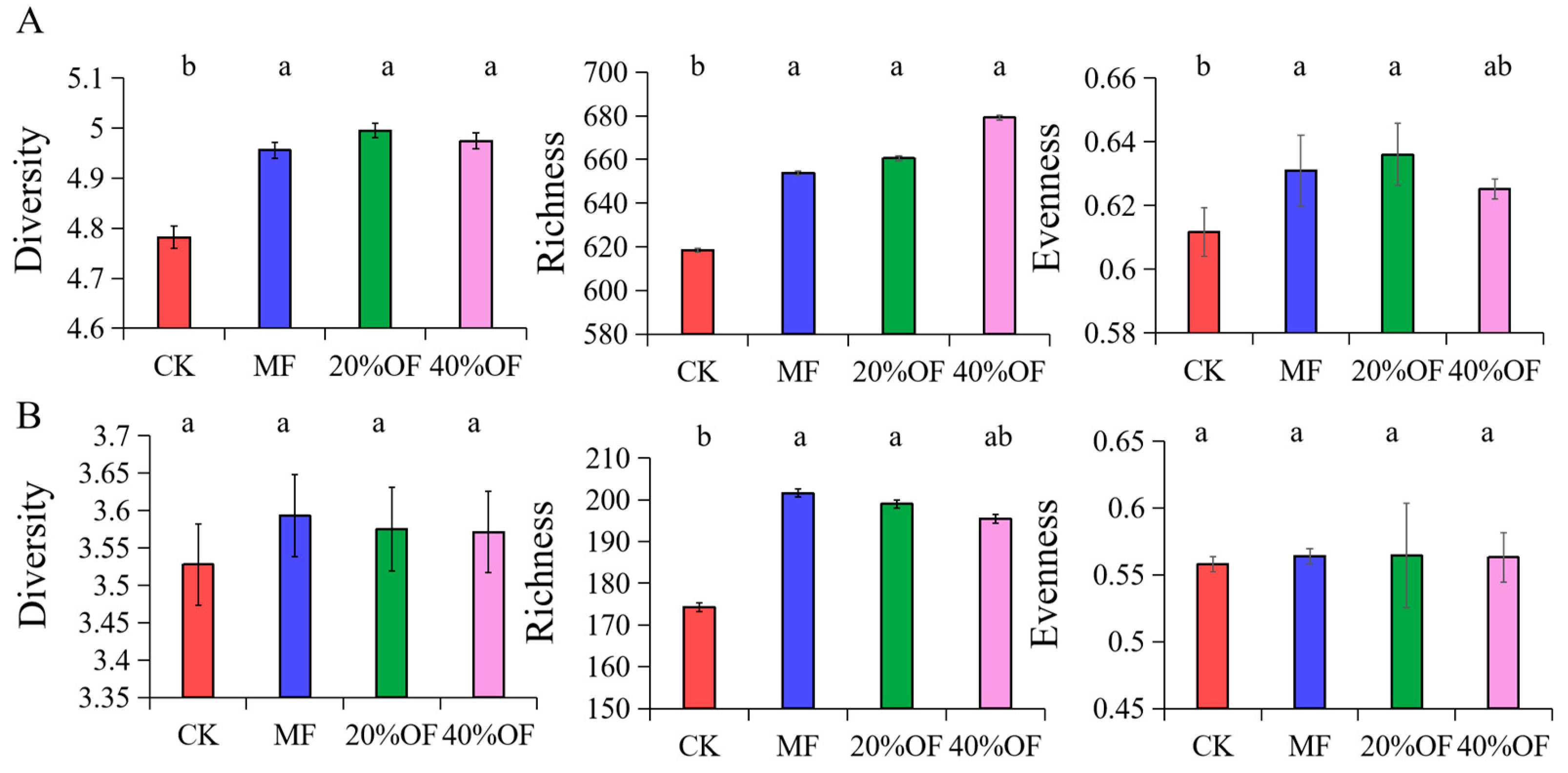
Figure 4.
Soil bacterial community structure under different organic substitution ratios. Principal coordinate analysis of Bray–Curtis distance for soil bacteria (A) and fungi (C). Redundancy analysis of Bray–Curtis distance for soil bacteria (B) and fungi (D).
Figure 4.
Soil bacterial community structure under different organic substitution ratios. Principal coordinate analysis of Bray–Curtis distance for soil bacteria (A) and fungi (C). Redundancy analysis of Bray–Curtis distance for soil bacteria (B) and fungi (D).
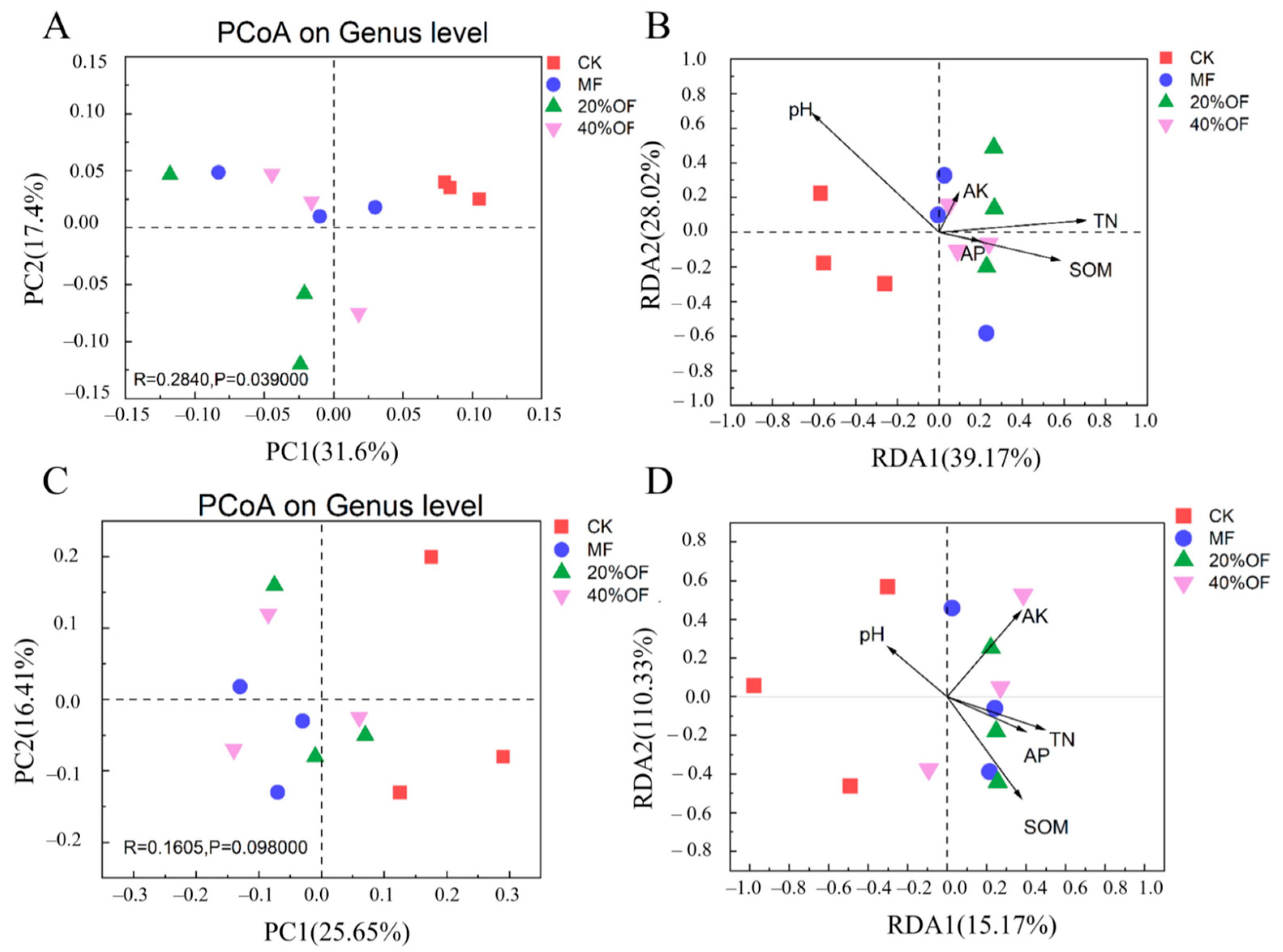
Figure 5.
Principal coordinate analysis of Bray–Curtis distance for soil bacteria (A) and fungi (B).
Figure 5.
Principal coordinate analysis of Bray–Curtis distance for soil bacteria (A) and fungi (B).

Figure 6.
Soil bacterial bipartite networks under different organic substitution ratios. (A) Bipartite networks display cropping system-specific OTUs in soil bacterial communities as determined using indicator species analysis. (B) Correlation analysis between indicator species and soil biochemical properties. SOM—soil organic matter; TN—total nitrogen; AP—available phosphorus; AK—available potassium.
Figure 6.
Soil bacterial bipartite networks under different organic substitution ratios. (A) Bipartite networks display cropping system-specific OTUs in soil bacterial communities as determined using indicator species analysis. (B) Correlation analysis between indicator species and soil biochemical properties. SOM—soil organic matter; TN—total nitrogen; AP—available phosphorus; AK—available potassium.

Figure 7.
Soil fungal bipartite networks under different organic substitution ratios. (A) Bipartite networks display cropping system-specific OTUs in soil fungal communities as determined using indicator species analysis. (B) Correlation analysis between indicator species and soil biochemical properties. SOM—soil organic matter; TN—total nitrogen; AP—available phosphorus; AK—available potassium.
Figure 7.
Soil fungal bipartite networks under different organic substitution ratios. (A) Bipartite networks display cropping system-specific OTUs in soil fungal communities as determined using indicator species analysis. (B) Correlation analysis between indicator species and soil biochemical properties. SOM—soil organic matter; TN—total nitrogen; AP—available phosphorus; AK—available potassium.
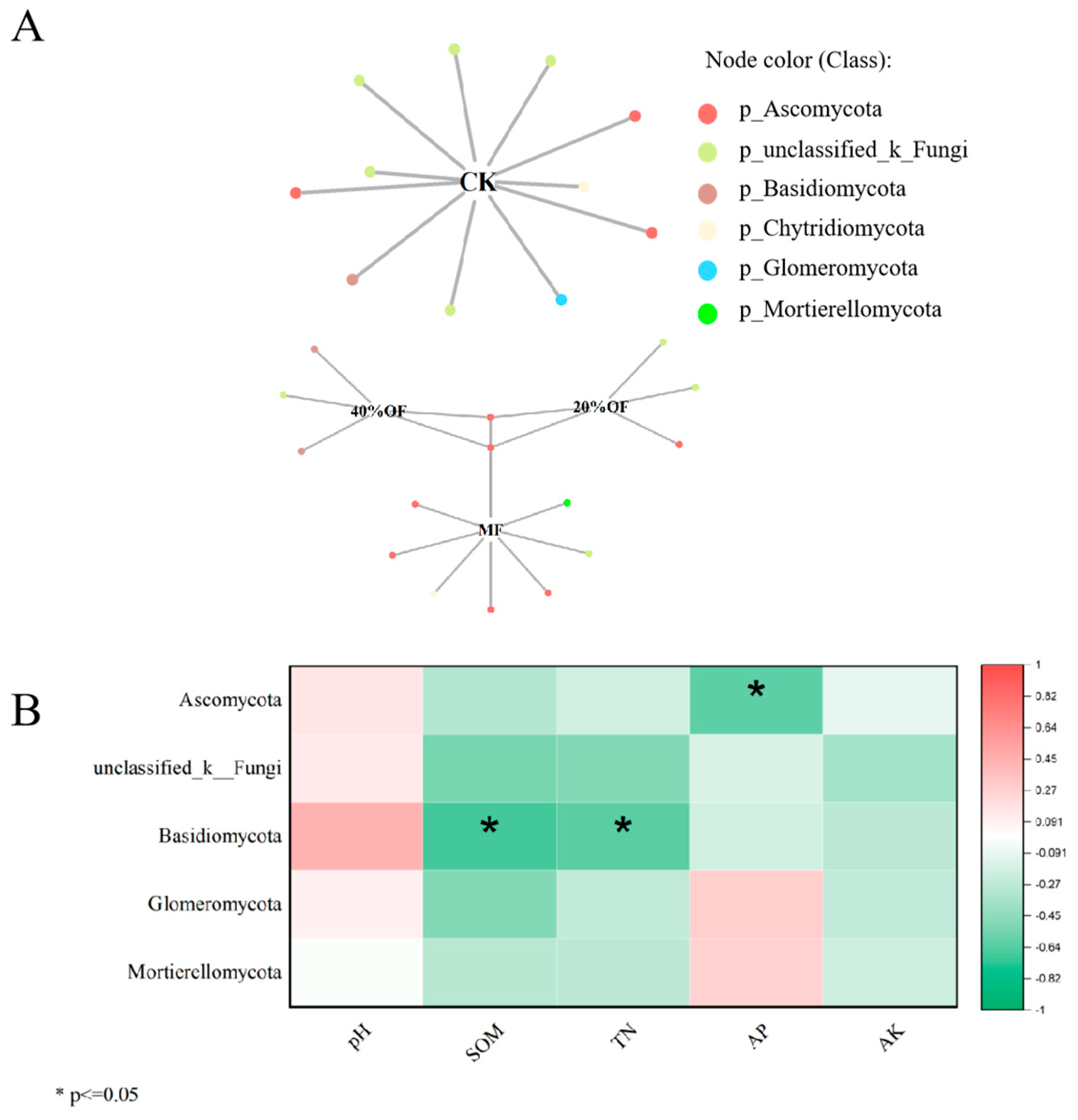
Figure 8.
A linear discriminant analysis (LDA) score computed for the significantly different abundant taxa of bacteria (A) and fungi (B).
Figure 8.
A linear discriminant analysis (LDA) score computed for the significantly different abundant taxa of bacteria (A) and fungi (B).

{kind=link}
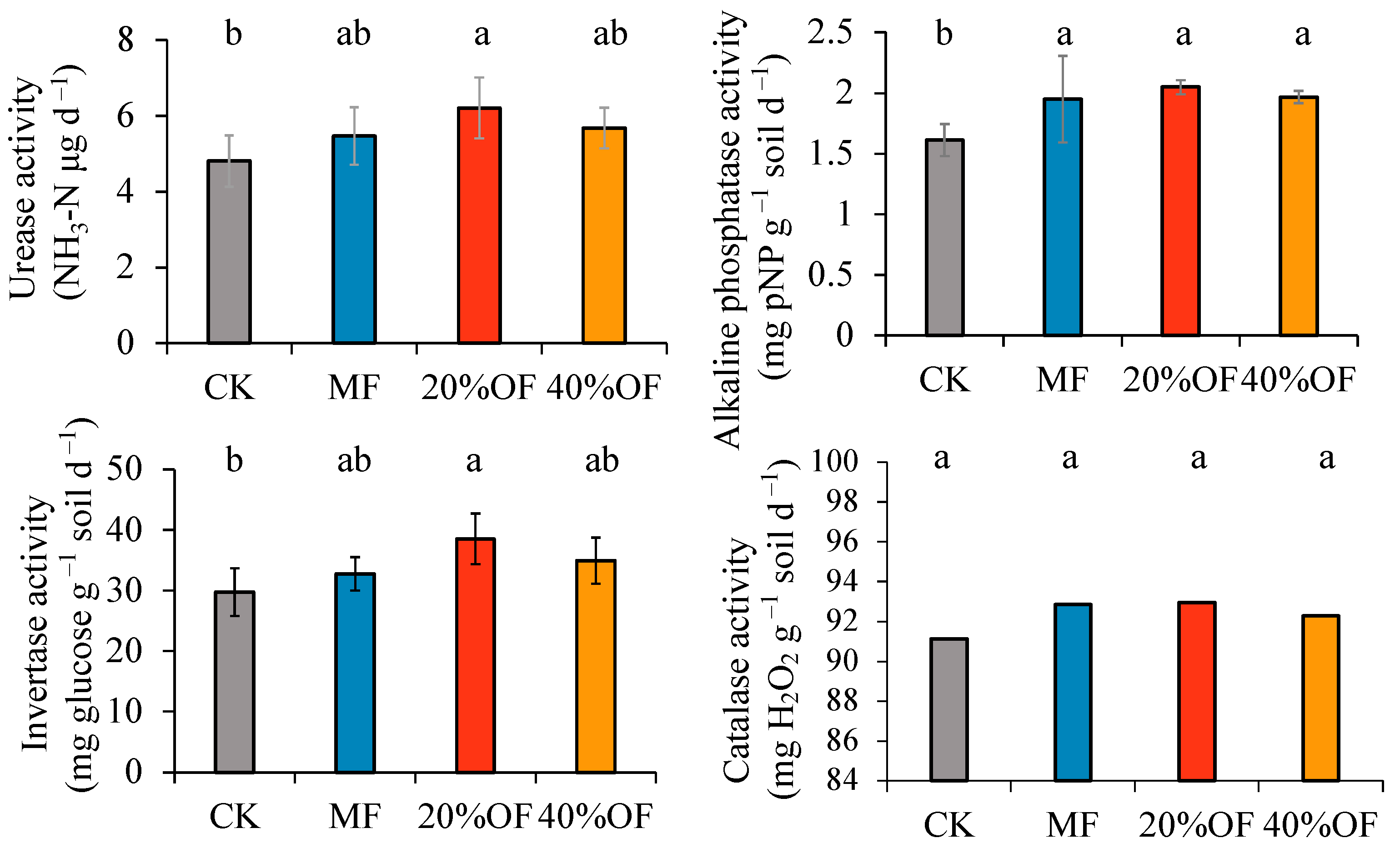{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Description of fertilizer treatments in this study.
| Cropping Season and Treatments | Fertilizer Source | N Rate | P2O5 Rate | K2O Rate | |
|---|---|---|---|---|---|
| Basal N | Topdressing N | ||||
| (kg ha−1) | |||||
| Wheat cropping season | |||||
| CK | CF | 0 | 0 | 105 | 75 |
| MF | CF | 97.5 | 97.5 | 105 | 75 |
| 20% OF | CF | 58.5 | 97.5 | 58.5 | 38.4 |
| OF | 39 | 0 | 46.5 | 36.6 | |
| 40% OF | CF | 19.5 | 97.5 | 12 | 1.8 |
| OF | 78 | 0 | 93 | 73.2 | |
| Maize cropping season | |||||
| CK | CF | 0 | 0 | 120 | 135 |
| MF | CF | 112.5 | 112.5 | 120 | 135 |
| 20% OF | CF | 112.5 | 112.5 | 120 | 135 |
| 40% OF | CF | 112.5 | 112.5 | 120 | 135 |
Note: CF—chemical fertilizer; OF—organic fertilizer; CK—no N fertilization control; MF—chemical fertilizer treatment; 20% OF—OF substitution for 20% of chemical fertilizer; 40% OF—OF substitution for 40% of chemical fertilizer.
Table 2.
Chemical properties of soil under different organic substitution ratios.
| Treatment | pH | Organic Matter (%) | Total N (g kg−1) | Available K (mg kg−1) | Available P (mg kg−1) |
|---|---|---|---|---|---|
| CK | 9.0 ± 0.18 a | 1.7 ± 0.38 b | 1.1 ± 0.08 b | 199 ± 13.11 a | 17.9 ± 2.73 a |
| MF | 8.9 ± 0.18 b | 2.0 ± 0.44 ab | 1.2 ± 0.21 ab | 200.3 ± 16.52 a | 18.9 ± 6.11 a |
| 20% OF | 8.8 ± 0.15 b | 2.1 ± 0.27 a | 1.3 ± 0.11 a | 202.0 ± 8.21 a | 20.9 ± 4.23 a |
| 40% OF | 8.9 ± 0.13 b | 2.2 ± 0.14 a | 1.3 ± 0.12 a | 201.7 ± 12.42 a | 19.5 ± 5.41 a |
Note: Data are shown as mean ± standard deviation (n = 3). Values followed by different lowercase letters indicated a significant difference among treatments (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Y.; Xu, Y.; Jiang, L.; Yang, Y.; Shi, J.; Guan, X.; Sun, T.; Zhao, H.; Wang, Y.; Liu, Y. Effect of Mild Organic Substitution on Soil Quality and Microbial Community. Agronomy 2024, 14, 888. https://doi.org/10.3390/agronomy14050888
AMA Style
Wang Y, Xu Y, Jiang L, Yang Y, Shi J, Guan X, Sun T, Zhao H, Wang Y, Liu Y. Effect of Mild Organic Substitution on Soil Quality and Microbial Community. Agronomy. 2024; 14(5):888. https://doi.org/10.3390/agronomy14050888
Chicago/Turabian StyleWang, Yijun, Yu Xu, Lihua Jiang, Yan Yang, Jing Shi, Xilin Guan, Tao Sun, Huanyu Zhao, Yafei Wang, and Yumin Liu. 2024. "Effect of Mild Organic Substitution on Soil Quality and Microbial Community" Agronomy 14, no. 5: 888. https://doi.org/10.3390/agronomy14050888
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.