Imbalance Water Deficit Improves the Seed Yield and Quality of Soybean

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Growth Conditions and Treatments
2.3. Measurement of Agronomic Characters
2.4. Quantification of Osmoprotectants in Soybean Leaves
2.4.1. Proline
2.4.2. Sucrose and Soluble Polysaccharide
2.4.3. Starch
2.5. Quantification of Qualities in Soybean Seeds
2.5.1. Protein
2.5.2. Isoflavones
2.5.3. Fatty Acids
2.6. Statistical Analyses
3. Results
3.1. Water Consumption and Soil Water Contents
3.2. Effect of Different Split-Root Water Imbalance Treatments on Morphological Characters of Soybean Genotypes
3.3. Effect of Different Split-Root Water Imbalance Treatments on Yield and Yield Components of Soybean Genotypes
3.4. Effect of Different Split-Root Water Imbalance Treatments on Biomass Accumulation and Distribution of Soybean Genotypes
3.5. Effect of Different Split-Root Water Imbalance Treatments on Osmoprotectant of Soybean Leaves
3.6. Effect of Different Split-Root Water Imbalance Treatments on Seed Protein Contents of Soybean Genotypes
3.7. Effect of Different Split-Root Water Imbalance Treatments on Isoflavone of Soybean Genotypes
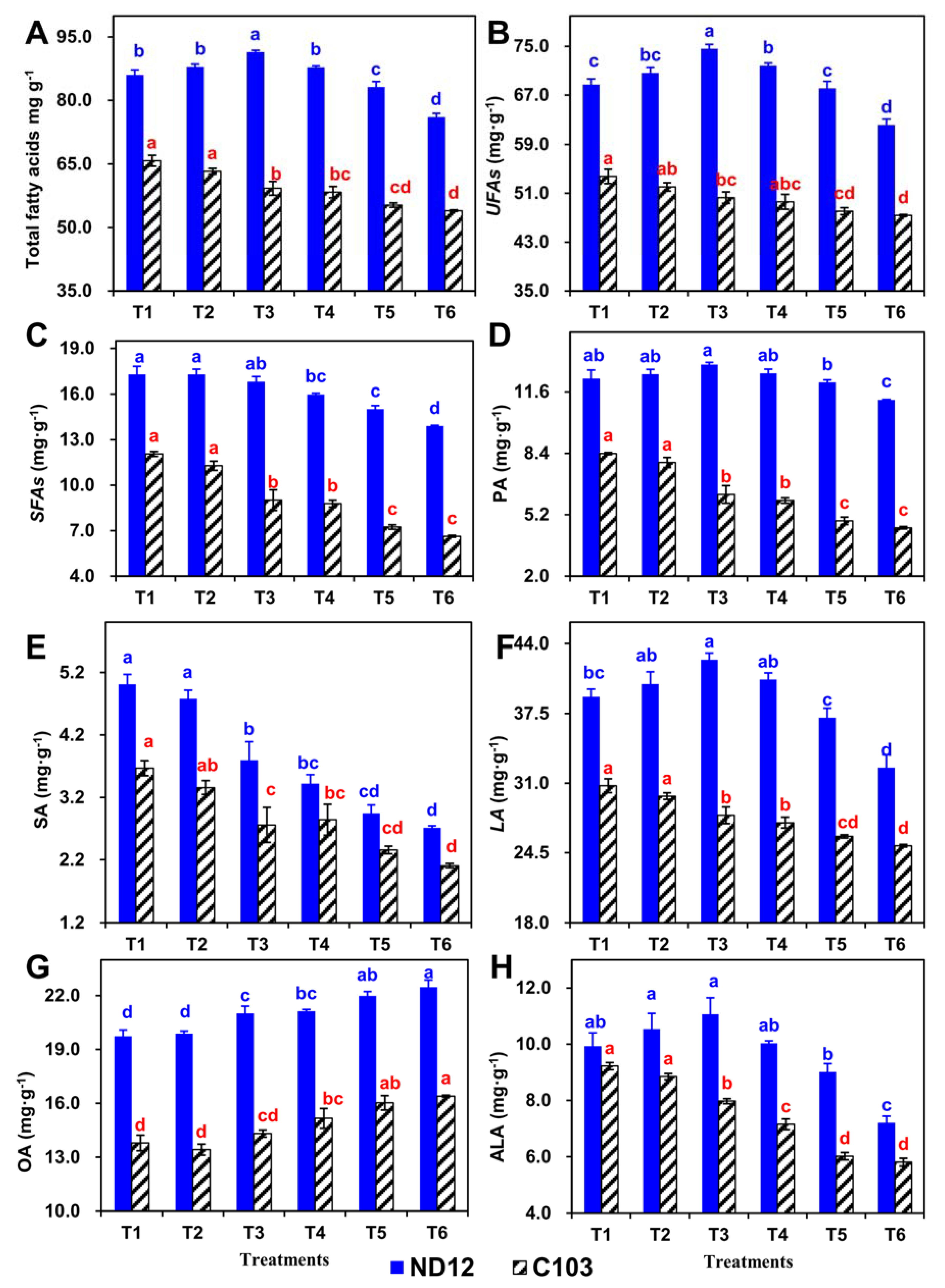
3.8. Effect of Different Split-Root Water Imbalance Treatments on Fatty Acids of Soybean Genotypes
4. Discussion
4.1. Effect of Split-Root Water Imbalance Treatments on Agro-Morphological Traits of Soybean
4.2. Effect of Split-Root Drought on Soybean Biomass Accumulation and Distribution
4.3. Effect of Split-Root Water Imbalance Treatments on Soybean Osmoprotectant
4.4. Soybean Metabolism and Split-Root Water Imbalance Treatments
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Yan, Y.; Gong, W.; Yang, W.; Wan, Y.; Chen, X.; Chen, Z.; Wang, L. Seed treatment with uniconazole powder improves soybean seedling growth under shading by corn in relay strip intercropping system. Plant Prod. Sci. 2010, 13, 367–374. [Google Scholar] [CrossRef]
- Rahman, T.; Ye, L.; Liu, X.; Iqbal, N.; Du, J.; Gao, R.; Liu, W.; Yang, F.; Yang, W. Water use efficiency and water distribution response to different planting patterns in maize–soybean relay strip intercropping systems. Exp. Agric. 2017, 53, 159–177. [Google Scholar] [CrossRef]
- Yang, F.; Cui, L.; Huang, S.; Liu, W.; Yong, T.; Yang, W. Soybean growth environment and group yield in soybean relay intercropped with different leaf type maize. Soybean Sci. 2015, 34, 402–407. [Google Scholar]
- Liu, X.; Rahman, T.; Song, C.; Su, B.; Yang, F.; Yong, T.; Wu, Y.; Zhang, C.; Yang, W. Changes in light environment, morphology, growth and yield of soybean in maize-soybean intercropping systems. Field Crops Res. 2017, 200, 38–46. [Google Scholar] [CrossRef]
- Lin, Y.; Feng, Y.; Ying, S.B.; Jing, Z.; Guo, L.W.; Wen, Y.Y. Effects of different field patterns on water distribution and water use efficiency in maize-soybean relay strip intercropping systems. Agric. Res. Arid Areas 2015, 33, 41–48. [Google Scholar]
- Bellaloui, N.; Mengistu, A. Seed composition is influenced by irrigation regimes and cultivar differences in soybean. Irrig. Sci. 2008, 26, 261–268. [Google Scholar] [CrossRef]
- Devi, M.A.; Giridhar, P. Variations in physiological response, lipid peroxidation, antioxidant enzyme activities, proline and isoflavones content in soybean varieties subjected to drought stress. PNAS India Sect. B Biol. Sci. 2015, 85, 35–44. [Google Scholar] [CrossRef]
- Gutierrez-Gonzalez, J.J.; Guttikonda, S.K.; Tran, L.-S.P.; Aldrich, D.L.; Zhong, R.; Yu, O.; Nguyen, H.T.; Sleper, D.A. Differential expression of isoflavone biosynthetic genes in soybean during water deficits. Plant Cell Physiol. 2010, 51, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Lozovaya, V.V.; Lygin, A.V.; Ulanov, A.V.; Nelson, R.L.; Daydé, J.; Widholm, J.M. Effect of temperature and soil moisture status during seed development on soybean seed isoflavone concentration and composition. Crop Sci. 2005, 45, 1934–1940. [Google Scholar] [CrossRef]
- Bellaloui, N.; Mengistu, A.; Kassem, M.A. Effects of genetics and environment on fatty acid stability in soybean seed. Food Nutr. Sci. 2013, 4, 165. [Google Scholar] [CrossRef]
- Specht, J.; Chase, K.; Macrander, M.; Graef, G.; Chung, J.; Markwell, J.; Germann, M.; Orf, J.; Lark, K. Soybean response to water. Crop Sci. 2001, 41, 493–509. [Google Scholar] [CrossRef]
- Lee, J.; Oliva, M.; Sleper, D.; Shannon, J. Irrigation has little effect on unsaturated fatty acid content in soya bean seed oil within genotypes differing in fatty acid profile. J. Agron. Crop Sci. 2008, 194, 320–324. [Google Scholar] [CrossRef]
- Desclaux, D.; Huynh, T.-T.; Roumet, P. Identification of soybean plant characteristics that indicate the timing of drought stress. Crop Sci. 2000, 40, 716–722. [Google Scholar] [CrossRef]
- Stolf-Moreira, R.; Medri, M.; Neumaier, N.; Lemos, N.; Pimenta, J.; Tobita, S.; Brogin, R.; Marcelino-Guimarães, F.; Oliveira, M.; Farias, J. Soybean physiology and gene expression during drought. Genet. Mol. Res. 2010, 9, 1946–1956. [Google Scholar] [CrossRef] [PubMed]
- Sadeghipour, O.; Abbasi, S. Soybean response to drought and seed inoculation. World Appl. Sci. J 2012, 17, 55–60. [Google Scholar]
- Bates, L.; Waldren, R.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Riazi, A.; Matsuda, K.; Arslan, A. Water-stress induced changes in concentrations of proline and other solutes in growing regions of young barley leaves. J. Exp. Bot. 1985, 36, 1716–1725. [Google Scholar] [CrossRef]
- Liu, J.; Qin, W.-T.; Wu, H.-J.; Yang, C.-Q.; Deng, J.-C.; Iqbal, N.; Liu, W.-G.; Du, J.-B.; Shu, K.; Yang, F. Metabolism variation and better storability of dark-versus light-coloured soybean (Glycine max L. Merr.) seeds. Food Chem. 2017, 223, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-Q.; Iqbal, N.; Hu, B.-Y.; Zhang, Q.-H.; Wu, H.-J.; Liu, X.; Zhang, J.; Liu, W.-G.; Yang, W.-Y.; Liu, J. Targeted metabolomics analysis of fatty acids in soybean seeds using GC-MS to reveal the metabolic manipulation of shading in the intercropping system. Anal. Methods 2017, 9, 2144–2152. [Google Scholar] [CrossRef]
- Desclaux, D.; Roumet, P. Impact of drought stress on the phenology of two soybean (Glycine max L. Merr) cultivars. Field Crops Res. 1996, 46, 61–70. [Google Scholar] [CrossRef]
- Liu, F.; Andersen, M.N.; Jensen, C.R. Loss of pod set caused by drought stress is associated with water status and aba content of reproductive structures in soybean. Funct. Plant Biol. 2003, 30, 271–280. [Google Scholar] [CrossRef]
- Zhang, M.; Zhai, Z.; Tian, X.; Duan, L.; Li, Z. Brassinolide alleviated the adverse effect of water deficits on photosynthesis and the antioxidant of soybean (Glycine max L.). Plant Growth Regul. 2008, 56, 257–264. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Siddique, K.H. Drought stress in wheat during flowering and grain-filling periods. Crit. Rev. Plant Sci. 2014, 33, 331–349. [Google Scholar] [CrossRef]
- Senapati, N.; Stratonovitch, P.; Paul, M.J.; Semenov, M.A. Drought Tolerance during Reproductive Development Is Important for Increasing Wheat Yield Potential under Climate Change in Europe. J. Exp. Bot. 2018. Available online: https://academic.oup.com/jxb/advance-article/doi/10.1093/jxb/ery226/5036557?searchresult=1 (accessed on 20 July 2018). [CrossRef] [PubMed]
- Witt, S.; Galicia, L.; Lisec, J.; Cairns, J.; Tiessen, A.; Araus, J.L.; Palacios-Rojas, N.; Fernie, A.R. Metabolic and phenotypic responses of greenhouse-grown maize hybrids to experimentally controlled drought stress. Mol. Plant 2012, 5, 401–417. [Google Scholar] [CrossRef] [PubMed]
- Son, C.; Park, S.; Kim, C.; Choi, B. Effects of Water Stress on Growth Characteristics and Grain Yield of Soybean. RDA J. Agric. Sci. 1996. Available online: http://agris.fao.org/agris-search/search.do?recordID=KR9603781 (accessed on 20 July 2018).
- Sarai Tabrizi, M.; Parsinejad, M.; Babazadeh, H. Efficacy of partial root drying technique for optimizing soybean crop production in semi-arid regions. Irrig. Drain. 2012, 61, 80–88. [Google Scholar] [CrossRef]
- Masoumi, H.; Darvish, F.; Daneshian, J.; Normohammadi, G.; Habibi, D. Effects of water deficit stress on seed yield and antioxidants content in soybean (Glycine max L.) cultivars. Afr. J. Agric. Res. 2011, 6, 1209–1218. [Google Scholar]
- Heatherly, L.G. Drought stress and irrigation effects on germination of harvested soybean seed. Crop Sci. 1993, 33, 777–781. [Google Scholar] [CrossRef]
- Hufstetler, E.V.; Boerma, H.R.; Carter, T.E.; Earl, H.J. Genotypic variation for three physiological traits affecting drought tolerance in soybean. Crop Sci. 2007, 47, 25–35. [Google Scholar] [CrossRef]
- Masoumi, H.; Masoumi, M.; Darvish, F.; Daneshian, J.; Nourmohammadi, G.; Habibi, D. Change in several antioxidant enzymes activity and seed yield by water deficit stress in soybean (Glycine max L.) cultivars. Not. Bot. Horti Agrobot. Cluj-Napoca 2010, 38, 86–94. [Google Scholar]
- Mayers, J.; Lawn, R.; Byth, D. Agronomic studies on soybean (Glycine max (L.) Merrill) in the dry seasons of the tropics. I. Limits to yield imposed by phenology. Aust. J. Agric. Res. 1991, 42, 1075–1092. [Google Scholar] [CrossRef]
- Jamieson, P.; Martin, R.; Francis, G.; Wilson, D. Drought effects on biomass production and radiation-use efficiency in barley. Field Crops Res. 1995, 43, 77–86. [Google Scholar] [CrossRef]
- Grinnan, R.; Carter, T.E.; Johnson, M.T. Effects of drought, temperature, herbivory, and genotype on plant–insect interactions in soybean (Glycine max). Arthropod-Plant Interact. 2013, 7, 201–215. [Google Scholar] [CrossRef]
- Li, F.; Wei, C.; Zhang, F.; Zhang, J.; Nong, M.; Kang, S. Water-use efficiency and physiological responses of maize under partial root-zone irrigation. Agric. Water Manag. 2010, 97, 1156–1164. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Shahnazari, A.; Andersen, M.N.; Jacobsen, S.-E.; Jensen, C.R. Effects of deficit irrigation (DI) and partial root drying (PRD) on gas exchange, biomass partitioning, and water use efficiency in potato. Sci. Hortic. 2006, 109, 113–117. [Google Scholar] [CrossRef]
- Huber, S.C.; Rogers, H.H.; Mowry, F.L. Effects of water stress on photosynthesis and carbon partitioning in soybean (Glycine max [L.] Merr.) plants grown in the field at different CO2 levels. Plant Physiol. 1984, 76, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Goldschmidt, E.E.; Huber, S.C. Regulation of photosynthesis by end-product accumulation in leaves of plants storing starch, sucrose, and hexose sugars. Plant Physiol. 1992, 99, 1443–1448. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Jensen, C.R.; Andersen, M.N. Drought stress effect on carbohydrate concentration in soybean leaves and pods during early reproductive development: Its implication in altering pod set. Field Crops Res. 2004, 86, 1–13. [Google Scholar] [CrossRef]
- Lee, B.R.; Jin, Y.L.; Jung, W.J.; Avice, J.C.; Morvan-Bertrand, A.; Ourry, A.; Park, C.W.; Kim, T.H. Water-deficit accumulates sugars by starch degradation—Not by de novo synthesis—In white clover leaves (Trifolium repens). Physiol. Plant. 2008, 134, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Villadsen, D.; Rung, J.H.; Nielsen, T.H. Osmotic stress changes carbohydrate partitioning and fructose-2, 6-bisphosphate metabolism in barley leaves. Funct. Plant Biol. 2005, 32, 1033–1043. [Google Scholar] [CrossRef]
- Ahmed, Z.; Waraich, E.A.; Rashid, A.; Shahbaz, M. Morpho-physiological and biochemical responses of camelina (Camelina sativa crantz) genotypes under drought stress. Int. Agric. Biol. 2017, 19. [Google Scholar] [CrossRef]
- Dener, E.; Kacelnik, A.; Shemesh, H. Pea plants show risk sensitivity. Curr. Biol. 2016, 26, 1763–1767. [Google Scholar] [CrossRef] [PubMed]
- Larrainzar, E.; Gil-Quintana, E.; Arrese-Igor, C.; González, E.M.; Marino, D. Split-root systems applied to the study of the legume-rhizobial symbiosis: What have we learned? J. Integr. Plant Biol. 2014, 56, 1118–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cechin, I.; Corniani, N.; de Fátima Fumis, T.; Cataneo, A.C. Ultraviolet-b and water stress effects on growth, gas exchange and oxidative stress in sunflower plants. Radiat. Environ. Biophys. 2008, 47, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Zhou, Y.; Duan, L.; Li, Z.; Eneji, A.E.; Li, J. Silicon effects on photosynthesis and antioxidant parameters of soybean seedlings under drought and ultraviolet-b radiation. J. Plant Physiol. 2010, 167, 1248–1252. [Google Scholar] [CrossRef] [PubMed]
- Al-Tawaha, A.; Seguin, P.; Smith, D.; Bonnell, R. Irrigation level affects isoflavone concentrations of early maturing soya bean cultivars. J. Agron. Crop Sci. 2007, 193, 238–246. [Google Scholar] [CrossRef]
- Raza, M.A.; Feng, L.Y.; Manaf, A.; Wasaya, A.; Ansar, M.; Hussain, A.; Khalid, M.H.B.; Iqbal, N.; Xi, Z.J.; Chen, Y.K. Sulphur application increases seed yield and oil content in sesame seeds under rainfed conditions. Field Crops Res. 2018, 218, 51–58. [Google Scholar] [CrossRef]
- Motzo, R.; Fois, S.; Giunta, F. Protein content and gluten quality of durum wheat (Triticum turgidum subsp. Durum) as affected by sowing date. J. Sci. Food Agric. 2007, 87, 1480–1488. [Google Scholar] [CrossRef]
- Iqbal, N.; Zhang, Q.; Wu, H.; Yang, C.; Deng, J.; Qin, W.; Zhang, J.; Yang, W.; Liu, J. Soybean (Glycine max L.) germplasm screening and geographical determination based on targeted isoflavone metabolomics. Appl. Ecol. Env. Res. 2018, 16, 3933–3953. [Google Scholar]
- Bennett, J.O.; Yu, O.; Heatherly, L.G.; Krishnan, H.B. Accumulation of genistein and daidzein, soybean isoflavones implicated in promoting human health, is significantly elevated by irrigation. J. Agric. food Chem. 2004, 52, 7574–7579. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Gonzalez, J.J.; Wu, X.; Gillman, J.D.; Lee, J.-D.; Zhong, R.; Yu, O.; Shannon, G.; Ellersieck, M.; Nguyen, H.T.; Sleper, D.A. Intricate environment-modulated genetic networks control isoflavone accumulation in soybean seeds. BMC Plant Biol. 2010, 10, 105. [Google Scholar] [CrossRef] [PubMed]
- Dhaubhadel, S.; Farhangkhoee, M.; Chapman, R. Identification and characterization of isoflavonoid specific glycosyltransferase and malonyltransferase from soybean seeds. J. Exp. Bot. 2008, 59, 981–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okubo, K.; Iijima, M.; Kobayashi, Y.; Yoshikoshi, M.; Uchida, T.; Kudou, S. Components responsible for the undesirable taste of soybean seeds. Biosci. Biotechnol. Biochem. 1992, 56, 99–103. [Google Scholar] [CrossRef]
- Gao, J.; Hao, X.; Thelen, K.D.; Robertson, G.P. Agronomic management system and precipitation effects on soybean oil and fatty acid profiles. Crop Sci. 2009, 49, 1049–1057. [Google Scholar] [CrossRef]
- Carrera, C.S.; Dardanelli, J.L. Water deficit modulates the relationship between temperature and unsaturated fatty acid profile in soybean seed oil. Crop Sci. 2017, 57, 3179–3189. [Google Scholar] [CrossRef]
- Dhakal, K.H.; Jung, K.-H.; Chae, J.-H.; Shannon, J.G.; Lee, J.-D. Variation of unsaturated fatty acids in soybean sprout of high oleic acid accessions. Food Chem. 2014, 164, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Bin, Q.; Rao, H.; Hu, J.N.; Liu, R.; Fan, Y.W.; Li, J.; Deng, Z.Y.; Zhong, X.; Du, F.L. The caspase pathway of linoelaidic acid (9t, 12t-c18: 2)-induced apoptosis in human umbilical vein endothelial cells. Lipids 2013, 48, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Raza, M.A.; Feng, L.Y.; Iqbal, N.; Manaf, A.; Khalid, M.H.B.; ur Rehman, S.; Wasaya, A.; Ansar, M.; Billah, M.; Yang, F.; et al. Effect of Sulphur Application on Photosynthesis and Biomass Accumulation of Sesame Varieties under Rainfed Conditions. Agronomy 2018, 8, 149. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Height (cm) | Stem Diameter (mm) | Number of Node (Plant−1) | Number of Branch (Plant−1) | Pod Length (cm) | ||
|---|---|---|---|---|---|---|
| Genotype | ND12 | 58.22 b | 6.02 b | 14.22 b | 5.55 b | 4.59 a |
| C103 | 86.11 a | 6.39 a | 16.66 a | 7.22 a | 3.96 b | |
| LSD (0.05) | 2.12 | 0.32 | 0.63 | 1.09 | 0.09 | |
| Treatment (T) | T1 | 72.50 a | 6.35 a | 15.83 a | 6.83 a | 4.45 a |
| T2 | 72.16 a | 6.31 a | 15.50 abc | 6.50 ab | 4.43 b | |
| T3 | 73.33 a | 6.44 a | 15.83 a | 6.50 ab | 4.30 c | |
| T4 | 73.00 a | 6.30 a | 15.66 ab | 6.33 ab | 4.23 cd | |
| T5 | 72.67 a | 6.06 ab | 15.00 bc | 6.33 ab | 4.17 d | |
| T6 | 69.33 b | 5.76 b | 14.83 c | 5.83 b | 3.95 e | |
| LSD (0.05) | 2.23 | 0.47 | 0.80 | 0.51 | 0.07 | |
| Interaction (G × T) | * | * | * | * | * |
| TB (g) | SYP (g) | SW (g) | NGP | NPP | NIPP | ||
|---|---|---|---|---|---|---|---|
| Genotype | ND12 | 43.99 a | 16.88 a | 19.57 a | 102.89 b | 52.16 b | 4.50 a |
| C103 | 44.57 a | 15.24 b | 12.53 b | 129.78 a | 60.67 a | 4.44 a | |
| LSD (0.05) | 0.92 | 0.64 | 0.80 | 13.92 | 2.89 | 1.26 | |
| Treatment (T) | T1 | 54.18 a | 21.22 a | 16.80 a | 142.17 a | 69.16 a | 2.00 d |
| T2 | 51.12 b | 19.74 b | 16.79 a | 129.83 b | 62.50 b | 3.66 c | |
| T3 | 46.12 c | 17.24 c | 16.48 a | 115.33 c | 56.16 c | 4.00 c | |
| T4 | 41.34 d | 14.42 d | 16.25 ab | 112.33 cd | 54.33 c | 5.00 b | |
| T5 | 38.31 e | 12.86 e | 15.57 b | 104.83 d | 51.16 c | 5.50 b | |
| T6 | 34.59 f | 10.88 f | 14.39 c | 93.50 e | 45.16 d | 6.67 a | |
| LSD (0.05) | 2.00 | 1.22 | 0.68 | 8.31 | 5.09 | 0.88 | |
| Interaction (G × T) | * | * | * | * | * | * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iqbal, N.; Hussain, S.; Zhang, X.-W.; Yang, C.-Q.; Raza, M.A.; Deng, J.-C.; Ahmad, S.; Ashgar, M.A.; Zhang, J.; Yang, W.; et al. Imbalance Water Deficit Improves the Seed Yield and Quality of Soybean. Agronomy 2018, 8, 168. https://doi.org/10.3390/agronomy8090168
Iqbal N, Hussain S, Zhang X-W, Yang C-Q, Raza MA, Deng J-C, Ahmad S, Ashgar MA, Zhang J, Yang W, et al. Imbalance Water Deficit Improves the Seed Yield and Quality of Soybean. Agronomy. 2018; 8(9):168. https://doi.org/10.3390/agronomy8090168
Chicago/Turabian StyleIqbal, Nasir, Sajad Hussain, Xiao-Wen Zhang, Cai-Qiong Yang, Muhammad Ali Raza, Jun-Cai Deng, Shoaib Ahmad, Muhammad Ahsan Ashgar, Jing Zhang, Wenyu Yang, and et al. 2018. "Imbalance Water Deficit Improves the Seed Yield and Quality of Soybean" Agronomy 8, no. 9: 168. https://doi.org/10.3390/agronomy8090168