Cytomegalovirus (CMV)-Specific Perforin and Granzyme B ELISPOT Assays Detect Reactivation of CMV Infection in Inflammatory Bowel Disease

Abstract
:Abbreviations
| CMV | cytomegalovirus |
| GzB | granzyme B |
| PFN | perforin |
| IBD | inflammatory bowel disease |
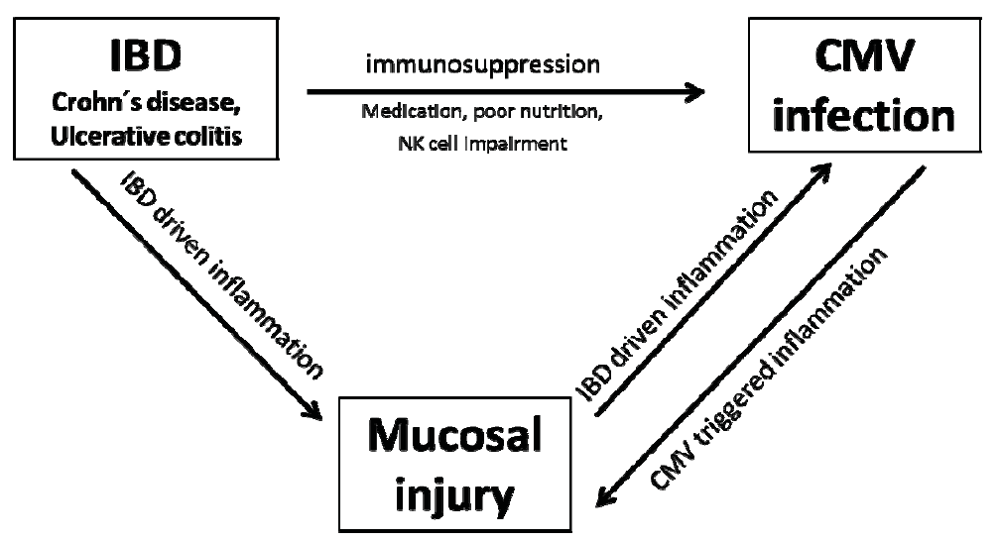
1. Introduction
2. Results
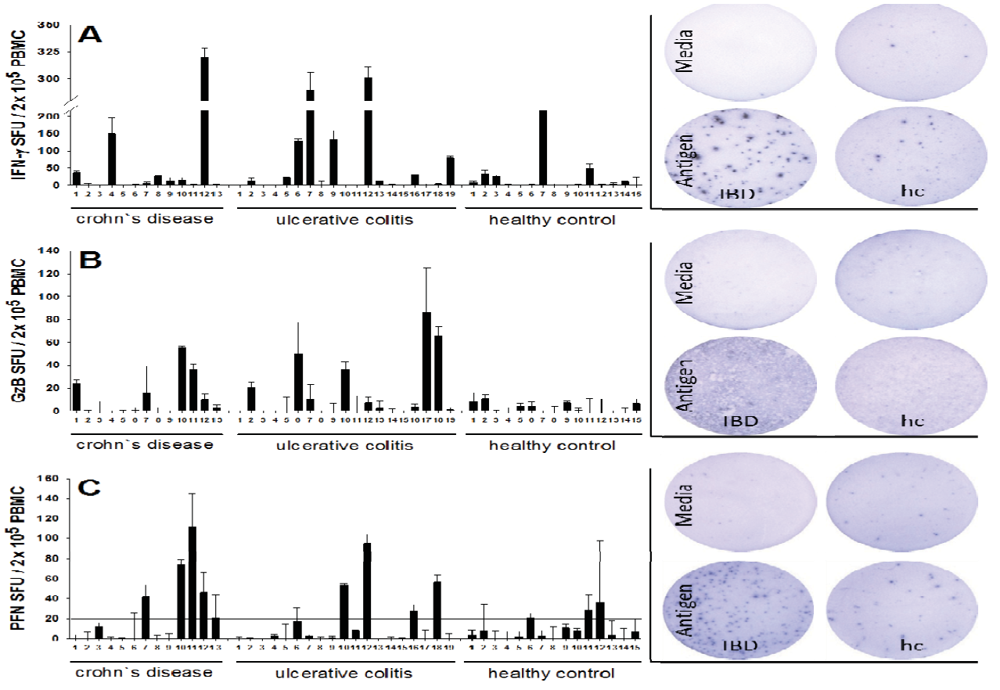
2.1. Increased Frequencies of Cytomegalovirus CMV-Reactive T Cells Producing IFN-γ, Granzyme B (GzB), and Perforin (PFN) in Inflammatory Bowel Disease (IBD) Patients
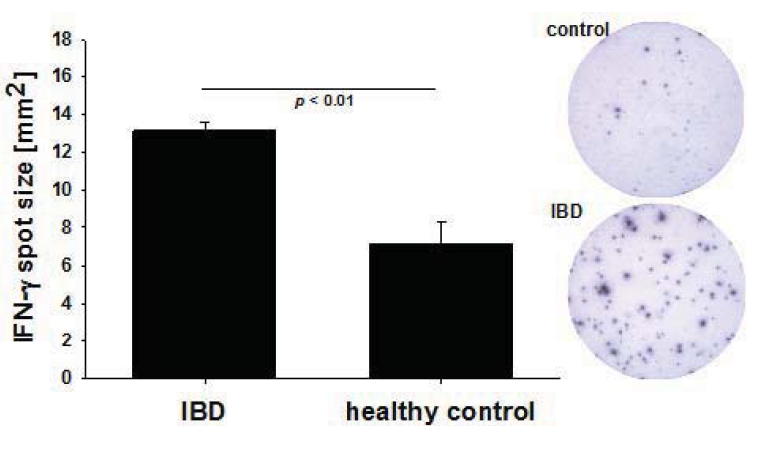
2.2. Increased Per Cell IFN-γ Productivity of CMV-Reactive T Cells in IBD Patients
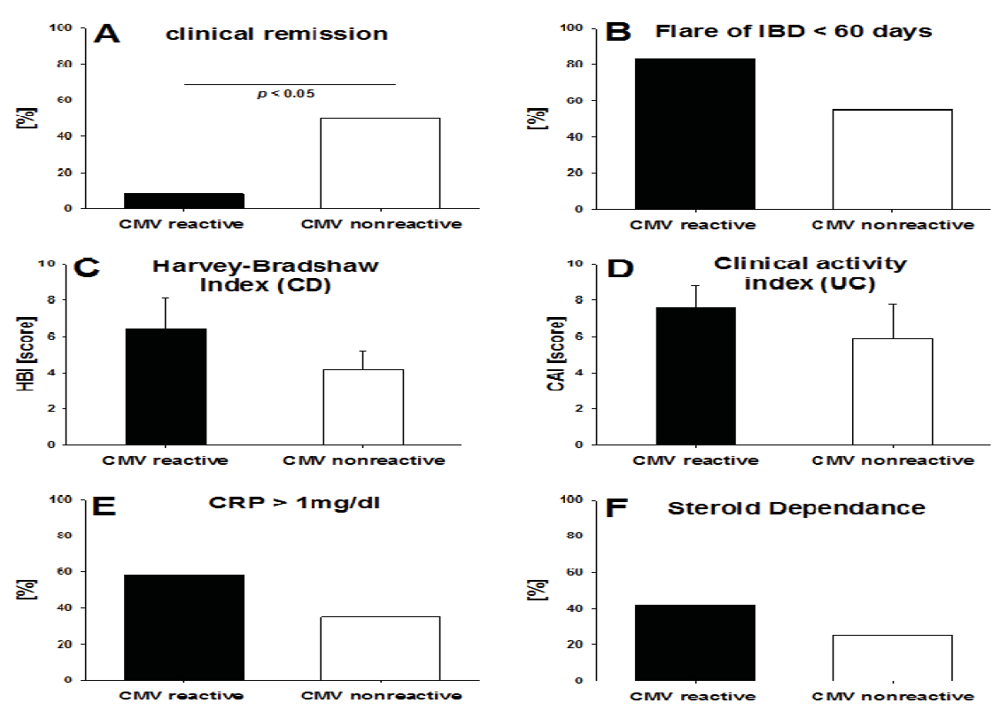
2.3. CMV-Specific T Cell Activity vs. the Clinical Activity of IBD



2.4. Detection of T Cell Activation by CMV vs. Viral Replication Measured by PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CMV DNA PCR replication measurements | |||
|---|---|---|---|
| CMV DNA PCR [copies/mL] | |||
| at ELISPOT testing | 1 month prior | ≥ 2 months prior | |
| IBD patient | |||
| all CD, all UC except 8,14,19 | neg. | not tested | not tested |
| UC 8 | neg. | 2270 | not tested |
| UC 14 | neg. | not tested | 1892 |
| UC 19 | neg. | 5403 | 31000 |
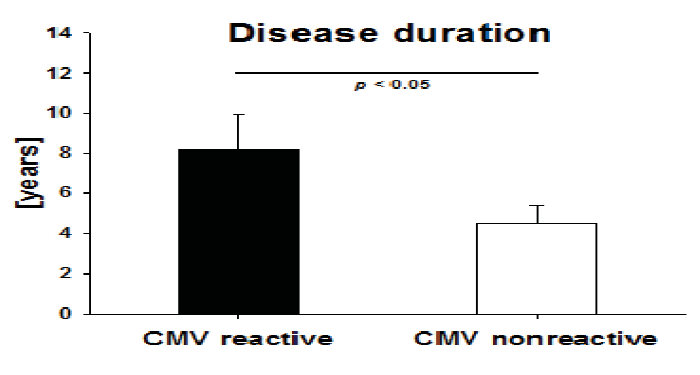
2.5. Disease Duration in IBD Patients with High CMV-Reactivity

3. Discussion

3. Experimental Section-Materials and Methods
3.1. Subjects, Cell Separation
3.2. CMV PCR Assay
3.3. Antigens
3.4. Cell Thawing and ELISPOT Assays
3.5. Statistics
4. Conclusions
Acknowledgements
Reference
- Criscuoli, V.; Rizzuto, M.R.; Cottone, M. Cytomegalovirus and inflammatory bowel disease: Is there a link? World J. Gastroenterol. 2006, 12, 4813–4818. [Google Scholar]
- Irving, P.M.; Gibson, P.R. Infections and IBD. Nat. Clin. Pract. Gastroenterol. Hepatol. 2008, 5, 18–27. [Google Scholar]
- Kishore, J.; Ghoshal, U.; Ghoshal, U.C.; Krishnani, N.; Kumar, S.; Singh, M.; Ayyagari, A. Infection with cytomegalovirus in patients with inflammatory bowel disease: Prevalence, clinical significance and outcome. J. Med. Microbiol. 2004, 53, 1155–1160. [Google Scholar] [CrossRef]
- Rahbar, A.; Bostrom, L.; Lagerstedt, U.; Magnusson, I.; Soderberg-Naucler, C.V.; Sundqvist, A. Evidence of active cytomegalovirus infection and increased production of IL-6 in tissue specimens obtained from patients with inflammatory bowel diseases. Inflamm. Bowel Dis. 2003, 9, 154–161. [Google Scholar] [CrossRef]
- Cottone, M.; Pietrosi, G.; Martorana, G.; Casa, A.; Pecoraro, G.; Oliva, L.; Orlando, A.; Rosselli, M.; Rizzo, A.; Pagliaro, L. Prevalence of cytomegalovirus infection in severe refractory ulcerative and Crohn’s colitis. Am. J. Gastroenterol. 2001, 96, 773–775. [Google Scholar]
- Kandiel, A.; Lashner, B. Cytomegalovirus colitis complicating inflammatory bowel disease. Am. J. Gastroenterol. 2006, 101, 2857–2865. [Google Scholar] [CrossRef]
- Vega, R.; Bertran, X.; Menacho, M.; Domenech, E.; Moreno de Vega, V.; Hombrados, M.; Cabre, E.; Ojanguren, I.; Gassull, M.A. Cytomegalovirus infection in patients with inflammatory bowel disease. Am. J. Gastroenterol. 1999, 94, 1053–1056. [Google Scholar] [CrossRef]
- Orvar, K.; Murray, J.; Carmen, G.; Conklin, J. Cytomegalovirus infection associated with onset of inflammatory bowel disease. Dig. Dis. Sci. 1993, 38, 2307–2310. [Google Scholar] [CrossRef]
- Pfau, P.; Kochman, M.L.; Furth, E.E.; Lichtenstein, G.R. Cytomegalovirus colitis complicating ulcerative colitis in the steroid-naive patient. Am. J. Gastroenterol. 2001, 96, 895–899. [Google Scholar]
- de la Hoz, R.E.; Stephens, G.; Sherlock, C. Diagnosis and treatment approaches of CMV infections in adult patients. J. Clin. Virol. 2002, 25, S1–S12. [Google Scholar]
- Mazzulli, T.; Drew, L.W.; Yen-Lieberman, B.; Jekic-McMullen, D.; Kohn, D.J.; Isada, C.; Moussa, G.; Chua, R.; Walmsley, S. Multicenter comparison of the digene hybrid capture CMV DNA assay (version 2.0), the pp65 antigenemia assay, and cell culture for detection of cytomegalovirus viremia. J. Clin. Microbiol. 1999, 37, 958–963. [Google Scholar]
- Rawlinson, W.D. Broadsheet Number 50: Diagnosis of human cytomegalovirus infection and disease. Pathology 1999, 31, 109–115. [Google Scholar] [CrossRef]
- Dimitroulia, E.; Spanakis, N.; Konstantinidou, A.E.; Legakis, N.J.; Tsakris, A. Frequent detection of cytomegalovirus in the intestine of patients with inflammatory bowel disease. Inflamm. Bowel Dis. 2006, 12, 879–884. [Google Scholar] [CrossRef]
- D’Ovidio, V.; Vernia, P.; Gentile, G.; Capobianchi, A.; Marcheggiano, A.; Viscido, A.; Martino, P.; Caprilli, R. Cytomegalovirus infection in inflammatory bowel disease patients undergoing anti-TNFalpha therapy. J. Clin. Virol. 2008, 43, 180–183. [Google Scholar] [CrossRef]
- Kuerten, S.; Nowacki, T.M.; Kleen, T.O.; Asaad, R.J.; Lehmann, P.V.; Tary-Lehmann, M. Dissociated production of perforin, granzyme B, and IFN-gamma by HIV-specific CD8(+) cells in HIV infection. AIDS Res. Hum. Retrovir. 2008, 24, 62–71. [Google Scholar] [CrossRef]
- Nowacki, T.M.; Kuerten, S.; Zhang, W.; Shive, C.L.; Kreher, C.R.; Boehm, B.O.; Lehmann, P.V.; Tary-Lehmann, M. Granzyme B production distinguishes recently activated CD8(+) memory cells from resting memory cells. Cell Immunol. 2007, 247, 36–48. [Google Scholar] [CrossRef]
- Harty, J.T.; Tvinnereim, A.R.; White, D.W. CD8+ T cell effector mechanisms in resistance to infection. Annu. Rev. Immunol. 2000, 18, 275–308. [Google Scholar] [CrossRef]
- Ramshaw, I.A.; Ramsay, A.J.; Karupiah, G.; Rolph, M.S.; Mahalingam, S.; Ruby, J.C. Cytokines and immunity to viral infections. Immunol. Rev. 1997, 159, 119–135. [Google Scholar] [CrossRef]
- Wong, P.; Pamer, E.G. CD8 T cell responses to infectious pathogens. Annu. Rev. Immunol. 2003, 21, 29–70. [Google Scholar] [CrossRef]
- Oehen, S.; Brduscha-Riem, K. Differentiation of naive CTL to effector and memory CTL: Correlation of effector function with phenotype and cell division. J. Immunol. 1998, 161, 5338–5346. [Google Scholar]
- Wherry, E.J.; Ahmed, R. Memory CD8 T-cell differentiation during viral infection. J. Virol. 2004, 78, 5535–5545. [Google Scholar] [CrossRef]
- Kaech, S.M.; Ahmed, R. Memory CD8+ T cell differentiation: Initial antigen encounter triggers a developmental program in naive cells. Nat. Immunol. 2001, 2, 415–422. [Google Scholar]
- Takata, H.; Takiguchi, M. Three memory subsets of human CD8+ T cells differently expressing three cytolytic effector molecules. J. Immunol. 2006, 177, 4330–4340. [Google Scholar]
- Wherry, E.J.; Teichgraber, V.; Becker, T.C.; Masopust, D.; Kaech, S.M.; Antia, R.; von Andrian, U.H.; Ahmed, R. Lineage relationship and protective immunity of memory CD8 T cell subsets. Nat. Immunol. 2003, 4, 225–234. [Google Scholar]
- Shresta, S.; Pham, C.T.; Thomas, D.A.; Graubert, T.A.; Ley, T.J. How do cytotoxic lymphocytes kill their targets? Curr. Opin. Immunol. 1998, 10, 581–587. [Google Scholar] [CrossRef]
- Trambas, C.M.; Griffiths, G.M. Delivering the kiss of death. Nat. Immunol. 2003, 4, 399–403. [Google Scholar]
- Catalfamo, M.; Henkart, P.A. Perforin and the granule exocytosis cytotoxicity pathway. Curr. Opin. Immunol. 2003, 15, 522–527. [Google Scholar] [CrossRef]
- Trapani, J.A.; Smyth, M.J. Functional significance of the perforin/granzyme cell death pathway. Nat. Rev. Immunol. 2002, 2, 735–747. [Google Scholar] [CrossRef]
- Trapani, J.A.; Sutton, V.R. Granzyme B: Pro-apoptotic, antiviral and antitumor functions. Curr. Opin. Immunol. 2003, 15, 533–543. [Google Scholar] [CrossRef]
- Henkart, P.A. Lymphocyte-mediated cytotoxicity: Two pathways and multiple effector molecules. Immunity 1994, 1, 343–346. [Google Scholar] [CrossRef]
- van den Brink, M.R.; Burakoff, S.J. Cytolytic pathways in haematopoietic stem-cell transplantation. Nat. Rev. Immunol. 2002, 2, 273–281. [Google Scholar] [CrossRef]
- Lanzavecchia, A.; Sallusto, F. Understanding the generation and function of memory T cell subsets. Curr. Opin. Immunol. 2005, 17, 326–332. [Google Scholar] [CrossRef]
- Currier, J.R.; Kuta, E.G.; Turk, E.; Earhart, L.B.; Loomis-Price, L.; Janetzki, S.; Ferrari, G.; Birx, D.L.; Cox, J.H. A panel of MHC class I restricted viral peptides for use as a quality control for vaccine trial ELISPOT assays. J. Immunol. Methods 2002, 260, 157–172. [Google Scholar] [CrossRef]
- Schlingmann, T.R.; Shive, C.L.; Targoni, O.S.; Tary-Lehmann, M.; Lehmann, P.V. Increased per cell IFN-gamma productivity indicates recent in vivo activation of T cells. Cell Immunol. 2009, 258, 131–137. [Google Scholar] [CrossRef]
- Navalpotro, D.; Gimeno, C.; Navarro, D. PCR detection of viral DNA in serum as an ancillary analysis for the diagnosis of acute mononucleosis-like syndrome due to human cytomegalovirus (HCMV) in immunocompetent patients. J. Clin. Virol. 2006, 35, 193–196. [Google Scholar] [CrossRef]
- Eyre-Brook, I.A.; Dundas, S. Incidence and clinical significance of colonic cytomegalovirus infection in idiopathic inflammatory bowel disease requiring colectomy. Gut 1986, 27, 1419–1425. [Google Scholar] [CrossRef]
- Matsuoka, K.; Iwao, Y.; Mori, T.; Sakuraba, A.; Yajima, T.; Hisamatsu, T.; Okamoto, S.; Morohoshi, Y.; Izumiya, M.; Ichikawa, H.; et al. Cytomegalovirus is frequently reactivated and disappears without antiviral agents in ulcerative colitis patients. Am. J. Gastroenterol. 2007, 102, 331–337. [Google Scholar] [CrossRef]
- Yoshino, T.; Nakase, H.; Mikami, S.; Nio, M.; Ueno, S.; Uza, N.; Ohmori, K.; Manabe, T.; Chiba, T. Importance of diagnosis of concomitant cytomegalovirus infection in patients with intestinal Behcet’s disease. Inflamm. Bowel Dis. 2008, 14, 877–878. [Google Scholar] [CrossRef]
- Hommes, D.W.; Sterringa, G.; van Deventer, S.J.; Tytgat, G.N.; Weel, J. The pathogenicity of cytomegalovirus in inflammatory bowel disease: A systematic review and evidence-based recommendations for future research. Inflamm. Bowel Dis. 2004, 10, 245–250. [Google Scholar] [CrossRef]
- Domenech, E.; Vega, R.; Ojanguren, I.; Hernandez, A.; Garcia-Planella, E.; Bernal, I.; Rosinach, M.; Boix, J.; Cabre, E.; Gassull, M.A. Cytomegalovirus infection in ulcerative colitis: A prospective, comparative study on prevalence and diagnostic strategy. Inflamm. Bowel Dis. 2008, 14, 1373–1379. [Google Scholar] [CrossRef]
- Harvey, R.F.; Bradshaw, J.M. A simple index of Crohn’s-disease activity. Lancet 1980, 1, 514. [Google Scholar]
- Rachmilewitz, D. Coated mesalazine (5-aminosalicylic acid) versus sulphasalazine in the treatment of active ulcerative colitis: A randomised trial. BMJ 1989, 298, 82–86. [Google Scholar] [CrossRef]
- Kreher, C.R.; Dittrich, M.T.; Guerkov, R.; Boehm, B.O.; Tary-Lehmann, M. CD4+ and CD8+ cells in cryopreserved human PBMC maintain full functionality in cytokine ELISPOT assays. J. Immunol. Methods 2003, 278, 79–93. [Google Scholar] [CrossRef]
- Gavin, M.A.; Gilbert, M.J.; Riddell, S.R.; Greenberg, P.D.; Bevan, M.J. Alkali hydrolysis of recombinant proteins allows for the rapid identification of class I MHC-restricted CTL epitopes. J. Immunol. 1993, 151, 3971–3980. [Google Scholar]
- Kondo, E.; Akatsuka, Y.; Kuzushima, K.; Tsujimura, K.; Asakura, S.; Tajima, K.; Kagami, Y.; Kodera, Y.; Tanimoto, M.; Morishima, Y.; Takahashi, T. Identification of novel CTL epitopes of CMV-pp65 presented by a variety of HLA alleles. Blood 2004, 103, 630–638. [Google Scholar] [CrossRef]
- McLaughlin-Taylor, E.; Pande, H.; Forman, S.J.; Tanamachi, B.; Li, C.R.; Zaia, J.A.; Greenberg, P.D.; Riddell, S.R. Identification of the major late human cytomegalovirus matrix protein pp65 as a target antigen for CD8+ virus-specific cytotoxic T lymphocytes. J. Med. Virol. 1994, 43, 103–110. [Google Scholar] [CrossRef]
- Wills, M.R.; Carmichael, A.J.; Mynard, K.; Jin, X.; Weekes, M.P.; Plachter, B.; Sissons, J.G. The human cytotoxic T-lymphocyte (CTL) response to cytomegalovirus is dominated by structural protein pp65: Frequency, specificity, and T-cell receptor usage of pp65-specific CTL. J. Virol. 1996, 70, 7569–7579. [Google Scholar]
- Guerkov, R.E.; Targoni, O.S.; Kreher, C.R.; Boehm, B.O.; Herrera, M.T.; Tary-Lehmann, M.; Lehmann, P.V.; Schwander, S.K. Detection of low-frequency antigen-specific IL-10-producing CD4(+) T cells via ELISPOT in PBMC: Cognate vs. nonspecific production of the cytokine. J. Immunol. Methods 2003, 279, 111–121. [Google Scholar] [CrossRef]
- Hawkins, N.; Self, S.; Wakefield, J. The automated counting of spots for the ELISpot assay. J. Immunol. Methods 2006, 316, 52–58. [Google Scholar] [CrossRef]
- Hesse, M.D.; Karulin, A.Y.; Boehm, B.O.; Lehmann, P.V.; Tary-Lehmann, M. A T cell clone’s avidity is a function of its activation state. J. Immunol. 2001, 167, 1353–1361. [Google Scholar]
- Karulin, A.Y.; Hesse, M.D.; Tary-Lehmann, M.; Lehmann, P.V. Single-cytokine-producing CD4 memory cells predominate in type 1 and type 2 immunity. J. Immunol. 2000, 164, 1862–1872. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nowacki, T.M.; Bettenworth, D.; Ross, M.; Heidemann, J.; Lehmann, P.V.; Lügering, A. Cytomegalovirus (CMV)-Specific Perforin and Granzyme B ELISPOT Assays Detect Reactivation of CMV Infection in Inflammatory Bowel Disease. Cells 2012, 1, 35-50. https://doi.org/10.3390/cells1020035
Nowacki TM, Bettenworth D, Ross M, Heidemann J, Lehmann PV, Lügering A. Cytomegalovirus (CMV)-Specific Perforin and Granzyme B ELISPOT Assays Detect Reactivation of CMV Infection in Inflammatory Bowel Disease. Cells. 2012; 1(2):35-50. https://doi.org/10.3390/cells1020035
Chicago/Turabian StyleNowacki, Tobias M., Dominik Bettenworth, Matthias Ross, Jan Heidemann, Paul V. Lehmann, and Andreas Lügering. 2012. "Cytomegalovirus (CMV)-Specific Perforin and Granzyme B ELISPOT Assays Detect Reactivation of CMV Infection in Inflammatory Bowel Disease" Cells 1, no. 2: 35-50. https://doi.org/10.3390/cells1020035