
Expression of PDLIM5 Spliceosomes and Regulatory Functions on Myogenesis in Pigs

1
National Engineering Laboratory for Livestock and Poultry Breeding, Beijing Key Laboratory of Animal Genetic Engineering, China Agricultural University, Beijing 100193, China
2
College of Animal Science and Technology, Yunnan Agricultural University, Kunming 650201, China
*
Authors to whom correspondence should be addressed.
Cells 2024, 13(8), 720; https://doi.org/10.3390/cells13080720
Submission received: 14 March 2024
/
Revised: 18 April 2024
/
Accepted: 19 April 2024
/
Published: 21 April 2024
Abstract
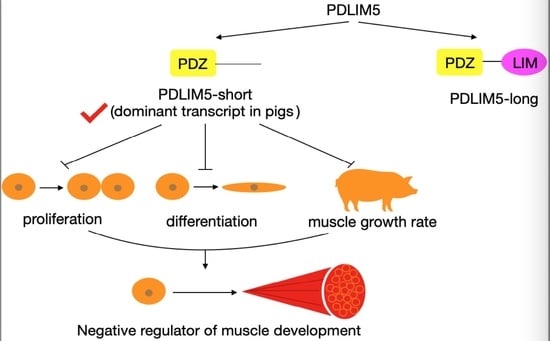
:Meat yield, determined by muscle growth and development, is an important economic trait for the swine industry and a focus of research in animal genetics and breeding. PDZ and LIM domain 5 (PDLIM5) are cytoskeleton-related proteins that play key roles in various tissues and cells. These proteins have multiple isoforms, primarily categorized as short (PDLIM5-short) and long (PDLIM5-long) types, distinguished by the absence and presence of an LIM domain, respectively. However, the expression patterns of swine PDLIM5 isoforms and their regulation during porcine skeletal muscle development remain largely unexplored. We observed that PDLIM5-long was expressed at very low levels in pig muscles and that PDLIM5-short and total PDLIM5 were highly expressed in the muscles of slow-growing pigs, suggesting that PDLIM5-short, the dominant transcript in pigs, is associated with a slow rate of muscle growth. PDLIM5-short suppressed myoblast proliferation and myogenic differentiation in vitro. We also identified two single nucleotide polymorphisms (−258 A > T and −191 T > G) in the 5′ flanking region of PDLIM5, which influenced the activity of the promoter and were associated with muscle growth rate in pigs. In summary, we demonstrated that PDLIM5-short negatively regulates myoblast proliferation and differentiation, providing a theoretical basis for improving pig breeding programs.
1. Introduction
During embryogenesis, skeletal muscles in vertebrate limbs develop from muscle progenitor cells derived from somites [1]. Postnatal muscle growth primarily relies on the expansion of muscle fiber cell volume. Muscle development involves a series of biological processes, including myoblast proliferation, differentiation, and fusion, leading to the formation of multinuclear myotubes, which mature into functional muscle fibers [2]. The growth of these muscle fibers is primarily regulated by a range of specific muscle-related transcription factors [3] and protein kinases [4]. Myogenic regulatory factors serve as pivotal hubs within the gene transcription network linked to muscle development, directly orchestrating gene expression to promote skeletal muscle development [5]. Cyclin-dependent kinases regulate the muscle cell cycle progression [6,7,8]. The precise timing and spatial location of the expression of these regulators are strictly controlled to ensure the accurate and orderly progression of muscle development [9,10].
The cytoskeleton plays a key role in maintaining the internal structure of cells, bearing external forces, and preserving cell morphology. Cytoskeleton-related proteins play an indispensable role in cytoskeletal changes. PDZ and LIM domain 5 (PDLIM5) are cytoskeleton-associated proteins belonging to the PDZ and LIM domain families, originally discovered using yeast two-hybrid screening methods [11]. PDLIM5, a scaffolding protein, comprises one N-terminal PDZ domain and three C-terminal LIM domains, which facilitate protein–protein interactions [12]. PDLIM5 generates multiple splice variants conserved across humans [13], rats [14], and mice [15]. Human PDLIM5 contains 9 isoforms (PDLIM5 isoform a–i). The long PDLIM5 containing LIM (PDLIM5-long) is widely expressed in various tissues, whereas short PDLIM5 without the LIM domain (PDLIM5-short) is specifically expressed in cardiac and skeletal muscles [16]. The expression of different spliceosomes occurs at distinct temporal points [17,18,19]. PDLIM5-long is predominant in the embryonic heart of mice, and the short isoforms are abundant in the adult heart [17] and skeletal muscles [18]. Comparable findings were also reported in the human myocardium [19], indicating that PDLIM5 fulfills distinct roles in different tissues. The porcine PDLIM5 encodes 11 isoforms; Among these, isoforms X1-9 possess a long LIM domain and are classified as PDLIM5-long, and X10 and X11 less LIM domain are classified as PDLIM5-short.
PDLIM5 binds to cytoskeletal and membrane proteins via its PDZ domain and interacts with various signaling molecules, including protein kinases and transcription factors, through its LIM domain [20]. Consequently, PDLIM5 plays diverse roles in various tissues and cells, including those of the heart, skeletal muscle, kidney [21], and nervous system [22,23,24]. PDLIM5 interacts with CREB to promote cardiomyocyte growth [25], and PDLIM5-knockout mice exhibit defects in heart contraction [15]. Additionally, PDLIM5 can play a role in the development of various diseases, including cancer [26,27,28], pulmonary hypertension [29], depressive disorders [30], and type 2 diabetes and hypertension [31], highlighting the significance of exploring the function of PDLIM5. The short and long isoforms of PDLIM5 seem to play distinct roles and have distinct functions in the development of striated muscles [18]. The expression of the long isoform of PDLIM5 gradually decreases during early myoblast differentiation and postnatal heart development [18], and it may facilitate the activation of satellite cells and the myogenic differentiation of C2C12 cells by targeting the inhibitor of DNA binding 1 or 2 (Id1 or Id2) [32,33]. The promotion of long splicing variants during myogenesis is also conserved in chicken skeletal muscle satellite cells [34]. Conversely, the expression of PDLIM5-short increases in the late postnatal period and restricts cardiomyocyte cytokinesis, hypertrophy, and myogenic differentiation [18], indicating a potential negative regulatory effect of PDLIM5-short in striated muscle development. However, the function and expression patterns of PDLIM5 with different splicing isoforms in pigs remain largely unknown. Our previous study based on transcriptome and proteome profiles demonstrated that PDLIM5 is differentially expressed in pigs exhibiting divergent growth phenotypes [35]. Therefore, we hypothesized that PDLIM5, a candidate gene for growth traits, exhibits differential expression in pigs and regulates muscle growth.
This study investigated the tissue expression profiles of swine PDLIM5 isoforms and identified their functions in myoblasts proliferation and differentiation. Additionally, we screened for single nucleotide polymorphisms (SNPs) across multiple pig breeds and analyzed the association between PDLIM5 and growth traits in these pigs.
2. Materials and Methods
2.1. Experimental Materials
Tibetan (TP) and Wujin (WJ) pigs, native Chinese pigs with slow growth characteristics, and large white (LW) pigs, fast-growing commercial pigs introduced from abroad, were reared at the Tibet Agriculture and Animal Husbandry College. For RNA extraction, at least six embryos from each group (TP, WJ and LW) were sampled from two pregnant sows after 60 days of insemination. The heart, back fat (BF), lung, liver, leg and intestinal embryonic tissues were used, all of which were used for semi-quantitative reverse transcriptional polymerase chain reaction (sqRT-PCR). Additionally, embryonic longissimus dorsi (LD) muscle tissues from the 12th rib were sampled for both sqRT-PCR and quantitative real-time polymerase chain reaction (qRT-PCR). Diannan small-ear (DSE) and New Huai populations (NHP), which are native Chinese pigs, were reared in Xishuangbanna, Yunnan and Dingyuan, Anhui, respectively. Ear samples were collected from four breeds (DSE, n = 16; TP, n = 34; LW, n = 24; NPH, n = 97) for DNA extraction. The animal feeding and tests were conducted based on the National Research Council Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee (approval number: AW80203202-1-1).
2.2. RNA Extraction and Expression Analysis
RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Subsequently, the RNA was reverse-transcribed into cDNA by using the FastKing One Step Reverse Transcription Kit (TIANGEN, Beijing, China). Target gene expression was evaluated using sqRT-PCR and qRT-PCR. qRT-PCR was performed using SYBR Green Master Mix (TIANGEN) and specific primers (Supplementary Table S1). Gene expression was analyzed using the 2−ΔΔCt method [36]. Target genes expression was normalized to GAPDH expression, which was used as a control.
2.3. SNP Screening, Genotyping, and Correlation Analysis
The primers for SNP screening of PDLIM5 (NC_010450.4) are listed in Supplementary Table S2. In general, the sequence approximately 2000 bp upstream of the gene is commonly cloned as the promoter of the gene. Thus, the amplification sequences covered approximately 2000 bp upstream of the PDLIM5 transcription start site. SNPs among different pig populations were screened and identified using PCR and Sanger sequencing (ABI3030XL DNA, Thermo Fisher Scientific, Wilmington, DE, USA). In brief, the PCR products amplified from ten individuals of each breed were pooled, and the purified products were sequenced to identify the SNPs by using Sanger sequencing. Chromas Pro (Technelysium, South Brisbane, Australia) and DNAMAN6.0 (Lynnon, San Ramon, CA, USA) were used to analyze and interpret the sequencing results. For determining the genotypes of the identified SNPs, individual samples were separately used as templates for PCR amplification, and the purified products were sequenced by using Sanger sequencing. Chromas Pro and DNAMAN6.0 software were used to analyze the peak patterns to determine the genotype. Ninety-seven boars, individuals of age weighing 30 and 90 kgs, were obtained from the Anhui Kexin Pig Breeding Farm.
2.4. Dual-Luciferase Reporter Assay
The PDLIM5 promoter region was cloned into the PGL3 luciferase reporter vector (Addgene, Cambridge, MA, USA). Then, the PDLIM5 promoter luciferase construct was transfected into cells in a 24-well plate for 48 h. Subsequently, the cells were lysed using 100 μL lysis buffer, and promoter activity was assayed using a dual luciferase reporter assay system (Promega, Madison, WI, USA). The luciferase activities were measured using a PerkinElmer 2030 Multilabel Reader (PerkinElmer, Waltham, MA, USA). The primers used are listed in Supplementary Table S3.
2.5. Primary Myoblasts Isolation and Culture
Primary myoblasts were isolated from the LD muscle of pigs. In brief, the minced muscles were digested using 0.2% type II collagenase (Sigma-Aldrich, St. Louis, MO, USA) and 2.5 U/mL dispase (Roche Applied Science, Nutley, NJ, USA) for 1 h. The filtered cell suspension was centrifuged at 1000× g for 5 min and cultured in Ham’s F10 nutrient mixture medium (Gibco, Waltham, MA USA), which was supplemented with 20% fetal bovine serum (Gibco), 1% penicillin-streptomycin (Gibco), and 5 ng/mL basic fibroblast growth factor (PeproTech, Cranbury, NJ, USA). The cells were incubated in an atmosphere of 5% CO2 atmosphere at 37 °C. DMEM supplemented with 2% horse serum (HS; Gibco) was used to induce myogenic differentiation.
2.6. Plasmid Construction and Transfection
The full-length sequence of porcine PDLIM5 was amplified using full-length-F/R primers (F: 5′-tagcgtttaaacttaATGAGCAACTACAGTGTGTC-3′; R: 5′-atccgagctcggtacCTGTACGTTAAGAGCACGTG-3′) and cloned into the pcDNA3.1 plasmid. For functional study, myoblasts were transfected with PDLIM5 overexpression plasmid using lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions.
2.7. Cell Counting Kit-8 (CCK8) and EdU Assays
After transfection for 24 h, cells were incubated with CCK8 (10 µL reagent dissolved in 100 µL DMEM, Beyotime Biotechnology, Shanghai, China) at 37 °C for 1–4 h in the dark. The absorbance at 450 nm was measured using a microplate reader (Biotek, Winooski, VT, USA) to determine the proliferation ability. For 5-ethynyl-2-deoxyuridine (EdU, Ribobio, Guangzhou, China) assays, cells were seeded in 96-well plates and transfected with control or overexpression plasmid for 24 h. Subsequently, transfected cells were incubated with 50 μM EdU at 37 °C for 2 h. Finally, the proliferated cells labeled using EdU were imaged under a microscope (Leica, Heidelberg, Germany).
2.8. Immunofluorescence
Immunofluorescence was performed according to our published method [37]. In brief, differentiated cells were fixed using 4% paraformaldehyde and permeabilized with 0.5% Triton X-100 for 15 min. After blocking for 1–2 h, the cells were incubated overnight with a mouse anti-myosin heavy chain antibody (cat. no. M4276, Sigma-Aldrich, St. Louis, MO, USA; 1:500) at 4 °C. Subsequently, the cells were washed thrice with PBS and incubated with a fluorescently labeled secondary antibody (A11032; Thermo Fisher Scientific, Wilmington, DE, USA, 1:400) for 1 h in the dark. The nuclei were stained with DAPI (Roche Applied Science, Nutley, NJ, USA) for 5 min. Digital images were captured using a fluorescence microscope (Leica image analysis system, Model Q500MC, Leica, Heidelberg, Germany).
2.9. Statistical Analysis
Statistical analyses were performed using one-way ANOVA and Student’s t-test. Results are presented as mean ± standard deviation (SD). Differences were considered statistically significant at p < 0.05. A χ2 test was used to analyze the distribution of genotypes and compare the differences in genotype distribution.
3. Results
3.1. Expression and Identification of Main PDLIM5 Isoforms in Pigs
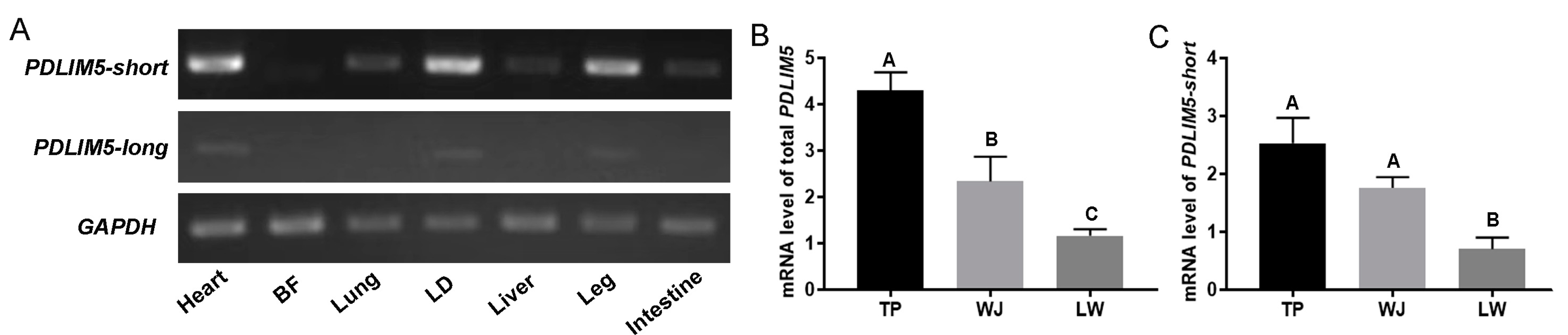
PDLIM5 has diverse splice variants, which may display different expression patterns and functions. PDLIM5-short was highly expressed in the heart, LD, and leg muscles in pigs, and PDLIM5-long was expressed at extremely low levels in various porcine tissues, including muscles (Figure 1A). In addition, total PDLIM5 (encoding all splice variants) was detected in the LD tissues of pigs with different growth rates. Slow-growing TP and WJ pigs exhibited significantly higher PDLIM5 expression in the LD tissues than fast-growing LW pigs (Figure 1B), which was consistent with our previous transcriptomic results [35]. Similarly, PDLIM5-short exhibited the same expression trend as total PDLIM5 (Figure 1C). These results indicate that PDLIM5-short is the dominant transcript in pigs and is likely associated with muscle growth rate. Consequently, we further focused on PDLIM5-short and conductedstudies on its potential role.
3.2. Inhibition of Inhibited Myoblast Proliferation by PDLIM5
As PDLIM5-short is the main splice variant in pigs, a PDLIM5-short overexpression plasmid was constructed and transfected into porcine primary myoblasts to explore its biological functions during myogenesis (Figure 2A). PDLIM5 overexpression markedly reduced the absorbance value after CCK8 treatment (Figure 2B) and EdU positivity, compared with that of the control (Figure 2C). Overexpression of PDLIM5 effectively downregulated proliferative genes, including cyclin D1 (CCND1), Ki67, proliferating cell nuclear antigen (PCNA), and cyclin-dependent kinase 4 (CDK4) (Figure 2D–G), further highlighting the negative role of PDLIM5 in myoblast proliferation.
3.3. PDLIM5 Suppressed Myogenic Differentiation
We investigated the role of PDLIM5 in porcine myoblasts during various stages of differentiation. Overexpression of PDLIM5 significantly inhibited myoblast differentiation, as demonstrated by decreased transcript levels of myogenin (MyoG), myogenic differentiation (MyoD), and myosin heavy chain (MyHC) on the second and fourth day of differentiation (Figure 3A–C). To confirm these results and visually observed myotube differentiation, we performed myosin immunofluorescence staining. Consistent with our expectations, fewer myotubes were formed by myoblasts overexpressing PDLIM5 than by those transfected with a control vector (Figure 3D,E). This findingindicates that PDLIM5 suppresses the myogenic differentiation of porcine myoblasts.
3.4. Screening of SNP Sites in the PDLIM5 Gene
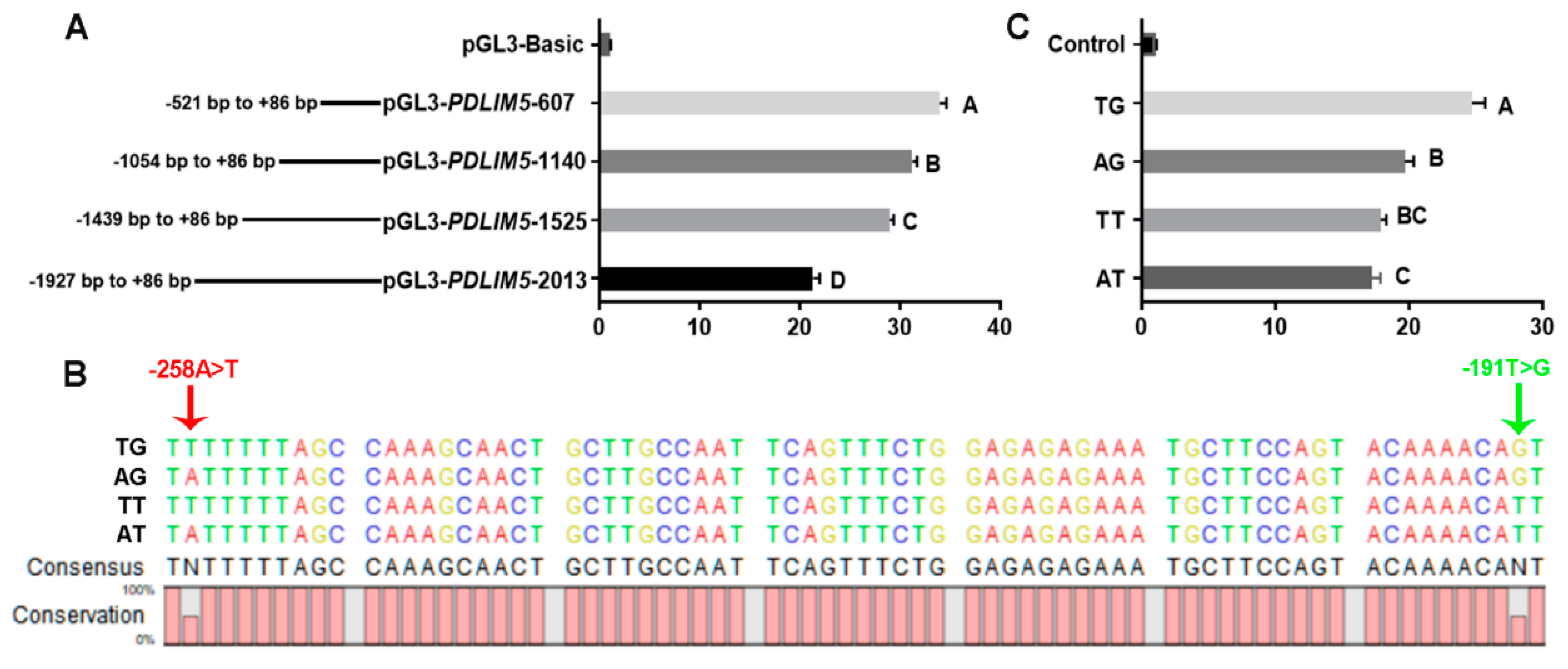
To identify promoter regions and SNPs that may affect transcription, we transfected C2C12 cells with different lengths of the PDLIM5 promoter region. The results showed that the region from −521 to +86 bp upstream of the translation start site (+1) of the promoter regions exhibited the strongest relative luciferase activity, indicating that this region served as the core promoter of PDLIM5 (Figure 4A).
Two SNPs, −258 A > T, and −191 T > G, were identified in the core promoter region of PDLIM5. The genotype and allele frequencies of these two SNPs in different breeds are listed in Table 1. In LW, the locus of −258 A > T was the AA genotype, and the site in mini-type indigenous breeds (DSE and TP) exhibited three genotypes (AA, AT, and TT) in Hardy–Weinberg Equilibrium (p > 0.05). The −191 T > G site showed higher frequencies of allele T in the introduced breeds (LW, frequencies of 0.938) than in indigenous pigs (DSE and TP; frequencies of 0.844 and 0.265 respectively). Collectively, the genotypes of the LW populations at both loci were almost all AT/AT, and different mutated genotypes were observed in the DSE and TP populations. To verify the effects of different genotypes at these two sites on promoter activity, we amplified and cloned four haplotype sequences formed by the combination of the two sites into dual-luciferase vectors (Figure 4B). The transcriptional activity of the promoters was significantly different, (the order being TG > AG > TT > AT) (Figure 4C).
The NHP population exhibited three genotypes at each SNP locus (Table 2). We compared the ages of the 30 and 90 kg body weights among the three genotypes. At the −258 A > T site, the AA genotype was associated with significantly faster growth than the AT and TT genotypes (p < 0.01). The TT genotype at the −191 T > G site showed faster growth than the TG and GG genotypes (p < 0.01). A further joint analysis of these two loci revealed that six combined genotypes were present in the NHP population (TG/TG = 3, AT/AG = 9, AT/AT =69, AT/TT = 5, AT/TG = 10, and AG/TG = 1), and the growth rate of AT/AT-type pigs was significantly higher than that of the other groups (Table 3).
4. Discussion
PDLIM5 contains the PDZ and LIM domains, which are associated with the cytoskeleton [38] and kinases [39]. We found that PDLIM5 was highly expressed in the muscles of slow-growing pig breeds, consistent with our prior study [35], suggesting its potential role in muscle development. PDLIM5 can be categorized into long isoforms with LIM domains, and short isoforms without LIM domains [20]. Long- and short-splice isoforms exist in mammals, including humans [16] and mice [14]. The splicing transition from LIM-containing PDLIM5-long to LIM-free PDLIM5-short was reported during postnatal heart development and in the early period of C2C12 differentiation [18]. In this study, we explored the role of two isoforms of the PDLIM5 gene in the regulation of skeletal muscle development in pigs. PDLIM5-long showed very low expression in pig muscles. PDLIM5-short, as the main isoform, was highly expressed in the muscles of slow-growing pigs, and its expression pattern was similar to that of total PDLIM5. These findings correlate the relative presence of the different isoforms with muscle growth rate, suggesting that the role of alternative splicing variants of the same gene has different effect on skeletal muscle biology. Especially, PDLIM5 short isoforms without LIM may have important but unknown functions in mature striated muscles. Therefore, we explored the function of PDLIM5-short in porcine muscle cells by using gene overexpression.
PDLIM5 acts as a scaffold that controls diverse cellular functions by interacting with various molecules [26]. PDLIM5 prevents proliferation and cell cycle progression in the nervous system [40]. Gan et al. reported that the RNA binding protein with multiple splicing represses PDLIM5-short isoforms by splicing exon 8 of PDLIM5, maintaining the expression of PDLIM5-long variants in embryonic cardiomyocytes. The absence of RBPMS leads to the accumulation of PDLIM5-short variants, which arrest the cell cycle and disrupt cardiomyocyte cytokinesis [41]. This results are consistent with our finding that PDLIM5-short inhibits myoblast proliferation. PDLIM5-short overexpression effectively downregulated the level of proliferative genes, including Ki67, PCNA and CDK4, but there was no significant decrease observed in the CCND1 level. This phenomenon may involve multiple complex factors. First, the expression of CCND1 is not solely controlled by a single gene but is part of a complex network involving multiple upstream genes and transcription factors. The overexpression effect of PDLIM5-short may be counteracted or modified by other regulatory factors. Second, gene expression is regulated at multiple levels, including transcriptional regulation, post-transcriptional modification, protein translation, and modification. Therefore, even if a gene is overexpressed at the transcriptional level, its ultimate impact on CCND1 expression may be limited by other regulatory mechanisms. Furthermore, intracellular gene expression is a dynamic equilibrium process with intricate interactions and feedback mechanisms among various genes and proteins. Overexpression of PDLIM5-short may trigger a cascade of reactions that may not be directly correlated with CCND1 expression.
The different functions of PDLIM5-long and PDLIM5-short in myoblast differentiation and hypertrophy were also identified. A study proved that PDLIM5-long promoted mice satellite cell activation during myogenesis [32]. However, PDLIM5-short disrupted normal muscle differentiation [42]. In rat neonatal cardiomyocytes, overexpression of the PDLIM5-long isoform increased the expression of hypertrophy markers and cell size, and the overexpression of the PDLIM5-short isoform prevented these hypertrophic changes [17]. Similarly, we observed that overexpression of LIM-free PDLIM5 significantly inhibited the expression of differentiation markers and myotube formation in porcine myoblasts, revealing a negative regulatory effect of PDLIM5-short isoform on muscle differentiation in pigs. This results also indicates that PDLIM5 is functionally conserved in mammalian muscle development.
These results show that PDLIM5-short may inhibit the growth and development of muscles by inhibiting myoblast proliferation and differentiation. This phenomenon is consistent with earlier results showing high expression of PDLIM5-short in native pig breeds with slow growth rates. Yamazaki et al. reported that PDLIM5-long promoted the C2C12 myogenic differentiation through the repression of Id2 translocation from the cytoplasm to the nucleus [25]. PDLIM5-short variants act as antagonists of PDLIM5-long variants in postnatal mouse hearts [17]. Regarding whether PDLIM5-short can also act as an antagonist of the PDLIM5-long variant in muscle cells to affect protein nuclear translocation, or whether PDLIM5-short, the main splicing variant in pigs, has a unique but unknown mechanism of regulating myoblast proliferation and differentiation, these questions require further experimental verification.
A study reported that polymorphisms in PDLIM5 are associated with an increased risk of dilated cardiomyopathy [43]. In this study, we performed segmented amplification of 2000 bp upstream of the transcription start site of the PDLIM5 gene and found that the promoter activity showed an increasing trend with the truncation of the fragment length. Thus, we speculated that the promoter region of PDLIM5 contains transcription suppression sites. The promoter activity in the −521 to +86 bp region was the highest, and we identified two SNPs (−258 A > T and −191 T > G) that formed four haplotypes (AT, AG, TT, and TG) in this region. The promoter activity of haplotype TG was significantly higher than that of AT, and TG haplotypes were mainly distributed in TP pigs, and almost all of haplotypes were AT in LW pigs. This results seemed to explain why the expression level of PDLIM5 was higher in TP pigs than in LW pigs. Additionally, correlation analysis of gene polymorphisms and growth rate showed that genotype AA of the −258 A > T site, genotype TT of the −191 T > G locus, and combined genotype AT/AT of the two loci were associated with fast body growth within NHP populations. Therefore, our results suggest that these two SNPs regulate the expression of PDLIM5 and postnatal body growth in pigs and are potential molecular markers for use in pig breeding. The regulation of gene expression and muscle development by SNPs requires further research in porcine primary myoblasts.
5. Conclusions
Our study revealed that PDLIM5-short was the dominant isoform in the muscle tissues of pigs and was highly expressed in the muscles of slow-growing pigs and that PDLIM5 inhibited myoblast proliferation and myogenic differentiation. We explored the biological functions of PDLIM5 and identified SNPs associated with the muscle growth rate that were used for genetic improvement. This findings provide a basis for further research on the mechanisms underlying muscle development and molecular breeding in pigs.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cells13080720/s1. Table S1: The primer sequences for SqRT-PCR and qPCR. Table S2: The primer sequences for SNP identification of the PDLIM5 gene. Table S3: The primer sequences for promoter region segmentation.
Author Contributions
Conceptualization, Y.F. and H.Z.; methodology, Y.F. and B.Z.; software, J.N.; validation, Y.F. and S.L.; formal analysis, S.L.; investigation, S.L. and D.Y.; resources, D.Y. and B.Z.; data curation, Y.F. and S.L.; writing—original draft preparation, Y.F.; writing—review and editing, H.Z.; visualization, X.H.; supervision, X.H.; project administration, B.Z; funding acquisition, Y.F., D.Y. and H.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Yunnan Major Science and Technology Project (202302AE090015), China Postdoctoral Science Foundation (2022M713404), National Natural Science Foundation of China (32060736).
Institutional Review Board Statement
The animal study protocol was approved by the China Agricultural University Animal Care and Use Committee on 8 February 2023 (permit number: AW80203202-1-1).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of the study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Buckingham, M.; Vincent, S.D. Distinct and dynamic myogenic populations in the vertebrate embryo. Curr. Opin. Genet. Dev. 2009, 19, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Wigmore, P.M.; Stickland, N.C. Muscle development in large and small pig fetuses. J. Anat. 1983, 137, 235–245. [Google Scholar] [PubMed]
- Reis, E.P.; Paixão, D.M.; Brustolini, O.J.; Silva, F.F.; Silva, W.; Araújo, F.M.; Salim, A.C.; Oliveira, G.; Guimarães, S.E. Expression of myogenes in longissimus dorsi muscle during prenatal development in commercial and local Piau pigs. Genet. Mol. Biol. 2016, 39, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Knight, J.D.; Kothary, R. The myogenic kinome: Protein kinases critical to mammalian skeletal myogenesis. Skelet. Muscle. 2011, 1, 29–47. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Zhang, J.; Raza, S.H.A.; Song, Y.; Jiang, C.; Song, X.; Wu, H.; Alotaibi, M.A.; Albiheyri, R.; Al-Zahrani, M.; et al. Interaction of MyoD and MyoG with Myoz2 gene in bovine myoblast differentiation. Res. Vet. Sci. 2022, 152, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Skapek, S.X.; Rhee, J.; Kim, P.S.; Novitch, B.G.; Lassar, A.B. Cyclin-mediated inhibition of muscle gene expression via a mechanism that is independent of pRB hyperphosphorylation. Mol. Cell. Biol. 1996, 16, 7043–7053. [Google Scholar] [CrossRef]
- Saab, R.; Bills, J.L.; Miceli, A.P.; Anderson, C.M.; Khoury, J.D.; Fry, D.W.; Navid, F.; Houghton, P.J.; Skapek, S.X. Pharmacologic inhibition of cyclin-dependent kinase 4/6 activity arrests proliferation in myoblasts and rhabdomyosarcoma-derived cells. Mol. Cancer Ther. 2006, 5, 1299–1308. [Google Scholar] [CrossRef]
- Skapek, S.X.; Rhee, J.; Spicer, D.B.; Lassar, A.B. Inhibition of myogenic differentiation in proliferating myoblasts by cyclin D1-dependent kinase. Science 1995, 267, 1022–1024. [Google Scholar] [CrossRef]
- Karimi Majd, S.; Gholami, M.; Bazgir, B. PAX7 and MyoD proteins expression in response to eccentric and concentric resistance exercise in active young men. Cell J. 2023, 25, 135–142. [Google Scholar] [CrossRef]
- Mohammadabadi, M.; Bordbar, F.; Jensen, J.; Du, M.; Guo, W. Key genes regulating skeletal muscle development and growth in farm animals. Animals 2021, 11, 835. [Google Scholar] [CrossRef]
- Kuroda, S.; Tokunaga, C.; Kiyohara, Y.; Higuchi, O.; Konishi, H.; Mizuno, K.; Gill, G.N.; Kikkawa, U. Protein-protein interaction of zinc finger LIM domains with protein kinase C. J. Biol. Chem. 1996, 271, 31029–31032. [Google Scholar] [CrossRef] [PubMed]
- Te Velthuis, A.J.; Bagowski, C.P. PDZ and LIM domain-encoding genes: Molecular interactions and their role in development. Sci. World J. 2007, 7, 1470–1492. [Google Scholar] [CrossRef] [PubMed]
- Niederländer, N.; Fayein, N.A.; Auffray, C.; Pomiès, P. Characterization of a new human isoform of the enigma homolog family specifically expressed in skeletal muscle. Biochem. Biophys. Res. Commun. 2004, 325, 1304–1311. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, N.; Hoshijima, M.; Oyasu, M.; Saito, N.; Tanizawa, K.; Kuroda, S. ENH, containing PDZ and LIM domains, heart/skeletal muscle-specific protein, associates with cytoskeletal proteins through the PDZ domain. Biochem. Biophys. Res. Commun. 2000, 272, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Kimura, K.; Peter, A.K.; Cui, L.; Ouyang, K.; Shen, T.; Liu, Y.; Gu, Y.; Dalton, N.D.; Evans, S.M.; et al. Loss of enigma homolog protein results in dilated cardiomyopathy. Circ. Res. 2010, 107, 348–356. [Google Scholar] [CrossRef]
- Ueki, N.; Seki, N.; Yano, K.; Masuho, Y.; Saito, T.; Muramatsu, M. Isolation, tissue expression, and chromosomal assignment of a human LIM protein gene, showing homology to rat enigma homologue (ENH). J. Hum. Genet. 1999, 44, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Wälchli, S.; Fujita, T.; Ryser, S.; Hoshijima, M.; Schlegel, W.; Kuroda, S.; Maturana, A.D. Splice variants of enigma homolog, differentially expressed during heart development, promote or prevent hypertrophy. Cardiovasc. Res. 2010, 86, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Ito, J.; Hashimoto, T.; Nakamura, S.; Aita, Y.; Yamazaki, T.; Schlegel, W.; Takimoto, K.; Maturana, A.D. Splicing transitions of the anchoring protein ENH during striated muscle development. Biochem. Biophys. Res. Commun. 2012, 421, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Verdonschot, J.A.J.; Robinson, E.L.; James, K.N.; Mohamed, M.W.; Claes, G.R.F.; Casas, K.; Vanhoutte, E.K.; Hazebroek, M.R.; Kringlen, G.; Pasierb, M.M.; et al. Mutations in PDLIM5 are rare in dilated cardiomyopathy but are emerging as potential disease modifiers. Mol. Genet. Genomic. Med. 2020, 8, e1049–e1063. [Google Scholar] [CrossRef]
- Huang, X.; Qu, R.; Ouyang, J.; Zhong, S.; Dai, J. An Overview of the Cytoskeleton-Associated Role of PDLIM5. Front. Physiol. 2020, 11, 975–983. [Google Scholar] [CrossRef]
- Su, Y.; Hiemstra, T.F.; Yan, Y.; Li, J.; Karet, H.I.; Rosen, L.; Moreno, P.; Karet Frankl, F.E. PDLIM5 links kidney anion exchanger 1 (kAE1) to ILK and is required for membrane targeting of kAE1. Sci. Rep. 2017, 7, 39701–39712. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.L.; Gou, C.Y.; Wang, W.H.; Li, Y.; Cui, Y.; Duan, J.J.; Chen, Y. A novel effect of PDLIM5 in α7 nicotinic acetylcholine receptor upregulation and surface expression. Cell. Mol. Life Sci. 2022, 79, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, Y.; Donta, M.; Mireles, L.L.; Paulucci-Holthauzen, A.; Waxham, M.N.; McCrea, P.D. Role of a PDLIM5: PalmD complex in directing dendrite morphology. bioRxiv 2023, 1, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, M.W.; Sun, Y.Y.; Hu, X.Y.; Geng, P.P.; Shu, H.; Wang, X.N.; Wang, H.; Zhang, J.F.; Cheng, H.Q.; et al. Nicotine pretreatment alleviates MK-801-induced behavioral and cognitive deficits in mice by regulating PDLIM5/CRTC1 in the PFC. Acta. Pharmacol. Sin. 2023, 44, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Ito, J.; Iijima, M.; Yoshimoto, N.; Niimi, T.; Kuroda, S.; Maturana, A.D. Scaffold protein enigma homolog activates CREB whereas a short splice variant prevents CREB activation in cardiomyocytes. Cell. Signal. 2015, 27, 2425–2433. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, L.; Huang, H.; Lv, J.M.; Chen, M.; Qu, F.J.; Pan, X.W.; Li, L.; Yin, L.; Cui, X.G.; et al. High expression of PDLIM5 facilitates cell tumorigenesis and migration by maintaining AMPK activation in prostate cancer. Oncotarget 2017, 8, 98117–98134. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, X.; Xu, Z.; He, Y.; Guo, C.; He, L.; Huan, C.; Cai, C.; Huang, J.; Zhang, J.; et al. PDLIM5 inhibits STUB1-mediated degradation of SMAD3 and promotes the migration and invasion of lung cancer cells. J. Biol. Chem. 2020, 295, 13798–13811. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhang, Y.; Yu, S.; Li, S.; Jiang, W.; Zhu, Y.; Xu, Y.; Yang, C.; Tian, G.; Mi, J.; et al. PDLIM5 identified by label-free quantitative proteomics as a potential novel biomarker of papillary thyroid carcinoma. Biochem. Biophys. Res. Commun. 2018, 499, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zhou, G.; Zhou, Q.; Tang, H.; Ibe, J.C.; Cheng, H.; Gou, D.; Chen, J.; Yuan, J.X.; Raj, J.U. Loss of microRNA-17-92 in smooth muscle cells attenuates experimental pulmonary hypertension via induction of PDZ and LIM domain 5. Am. J. Respir. Crit. Care. Med. 2015, 191, 678–692. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, W.; Xiao, Z.; Wang, G.; Yin, S.; Zhu, F.; Wang, H.; Cheng, J.; Wang, X.; He, X.; et al. A major single nucleotide polymorphism of the PDLIM5 gene associated with recurrent major depressive disorder. J. Psychiatry. Neurosci. 2008, 33, 43–46. [Google Scholar]
- Owusu, D.; Pan, Y.; Xie, C.; Harirforoosh, S.; Wang, K.S. Polymorphisms in PDLIM5 gene are associated with alcohol dependence, type 2 diabetes, and hypertension. J. Psychiatr. Res. 2017, 84, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, M.; Ito, J.; Koyama, R.; Iijima, M.; Yoshimoto, N.; Niimi, T.; Kuroda, S.; Maturana, A.D. Scaffold protein enigma homolog 1 overcomes the repression of myogenesis activation by inhibitor of DNA binding 2. Biochem. Biophys. Res. Commun. 2016, 474, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Liu, N.; Luo, L.; Zhong, J.; Tang, Z.; Kang, K.; Qu, J.; Peng, W.; Liu, L.; Li, L.; et al. MicroRNA-17-92 regulates myoblast proliferation and differentiation by targeting the ENH1/Id1 signaling axis. Cell Death Differ. 2016, 23, 1658–1669. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Yin, H.; Yu, X.; Zhang, Y.; Ma, M.; Li, D.; Zhu, Q. PDLIM5 affects chicken skeletal muscle satellite cell proliferation and differentiation via the p38-MAPK Pathway. Animals 2021, 11, 1016. [Google Scholar] [CrossRef] [PubMed]
- Shang, P.; Wang, Z.; Chamba, Y.; Zhang, B.; Zhang, H.; Wu, C. A comparison of prenatal muscle transcriptome and proteome profiles between pigs with divergent growth phenotypes. J. Cell Biochem. 2019, 120, 5277–5286. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Shang, P.; Zhang, B.; Tian, X.; Nie, R.; Zhang, R.; Zhang, H. Function of the porcine TRPC1 gene in myogenesis and muscle growth. Cells 2021, 10, 147. [Google Scholar] [CrossRef] [PubMed]
- Guy, P.M.; Kenny, D.A.; Gill, G.N. The PDZ domain of the LIM protein enigma binds to beta-tropomyosin. Mol. Biol. Cell 1999, 10, 1973–1984. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Ruiz-Lozano, P.; Martone, M.E.; Chen, J. Cypher, a striated muscle-restricted PDZ and LIM domain-containing protein, binds to alpha-actinin-2 and protein kinase C. J. Biol. Chem. 1999, 274, 19807–19813. [Google Scholar] [CrossRef]
- Lasorella, A.; Iavarone, A. The protein ENH is a cytoplasmic sequestration factor for Id2 in normal and tumor cells from the nervous system. Proc. Natl. Acad. Sci. USA 2006, 103, 4976–4981. [Google Scholar] [CrossRef]
- Gan, P.; Wang, Z.; Morales, M.G.; Zhang, Y.; Bassel-Duby, R.; Liu, N.; Olson, E.N. RBPMS is an RNA-binding protein that mediates cardiomyocyte binucleation and cardiovascular development. Dev. Cell 2022, 57, 959–973. [Google Scholar] [CrossRef] [PubMed]
- Ito, J.; Takita, M.; Takimoto, K.; Maturana, A.D. Enigma homolog 1 promotes myogenic gene expression and differentiation of C2C12 cells. Biochem. Biophys. Res. Commun. 2013, 435, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Fang, J.; Lv, J.; Pan, Z.; Yin, X.; Cheng, H.; Guo, X. Novel polymorphisms in PDLIM3 and PDLIM5 gene encoding Z-line proteins increase risk of idiopathic dilated cardiomyopathy. J. Cell. Mol. Med. 2019, 23, 7054–7062. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Characterization of PDLIM5 expression in pigs. (A) Determination of PDLIM5 expression in different tissues of TP pigs at the embryonic stage using sqRT-PCR. Expression of total PDLIM5 (B) and PDLIM5-short (C) at the transcript level in the LD of three pig breeds. Total PDLIM5 contains all splice variants; PDLIM5-short, short isoform of PDLIM5 without the LIM domain, and PDLIM5-long, long isoform PDLIM5 with the LIM domain. LD for longissimus dorsi; BF for back fat. TP for tibetan pig (n = 6), WJ for wujin pig (n = 6), LW for large white (n = 6). Each bar represents the means ± SD. Different letters indicate significant differences between groups.
Figure 1.
Characterization of PDLIM5 expression in pigs. (A) Determination of PDLIM5 expression in different tissues of TP pigs at the embryonic stage using sqRT-PCR. Expression of total PDLIM5 (B) and PDLIM5-short (C) at the transcript level in the LD of three pig breeds. Total PDLIM5 contains all splice variants; PDLIM5-short, short isoform of PDLIM5 without the LIM domain, and PDLIM5-long, long isoform PDLIM5 with the LIM domain. LD for longissimus dorsi; BF for back fat. TP for tibetan pig (n = 6), WJ for wujin pig (n = 6), LW for large white (n = 6). Each bar represents the means ± SD. Different letters indicate significant differences between groups.
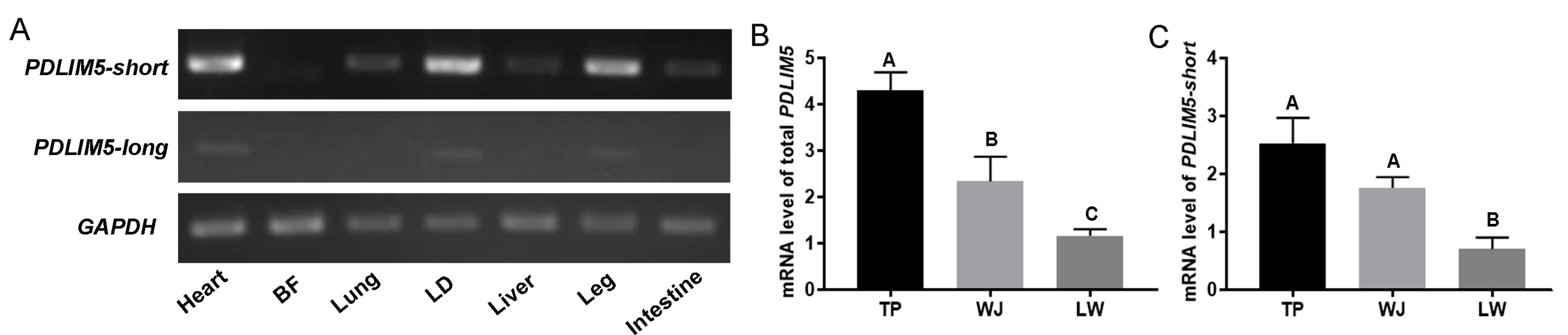
Figure 2.
PDLIM5 inhibits myoblast proliferation. (A) Efficiency of the detection of PDLIM5-short overexpression plasmid. (B) CCK8 assay of proliferating myoblasts transfected with overexpression constructs. (C) Representative images of EdU staining for proliferated cells after pcDNA3.1-PDLIM5 transfection. Blue indicates nuclei stained with DAPI, red indicates EdU-positive proliferating cells, scale bar = 130 µm. (D–G) The mRNA expression levels of proliferative genes. GAPDH is used as a reference gene. The data represent the mean ± SD of three independent experiments, ** p < 0.01, *** p < 0.001, N.S., not significant.
Figure 2.
PDLIM5 inhibits myoblast proliferation. (A) Efficiency of the detection of PDLIM5-short overexpression plasmid. (B) CCK8 assay of proliferating myoblasts transfected with overexpression constructs. (C) Representative images of EdU staining for proliferated cells after pcDNA3.1-PDLIM5 transfection. Blue indicates nuclei stained with DAPI, red indicates EdU-positive proliferating cells, scale bar = 130 µm. (D–G) The mRNA expression levels of proliferative genes. GAPDH is used as a reference gene. The data represent the mean ± SD of three independent experiments, ** p < 0.01, *** p < 0.001, N.S., not significant.
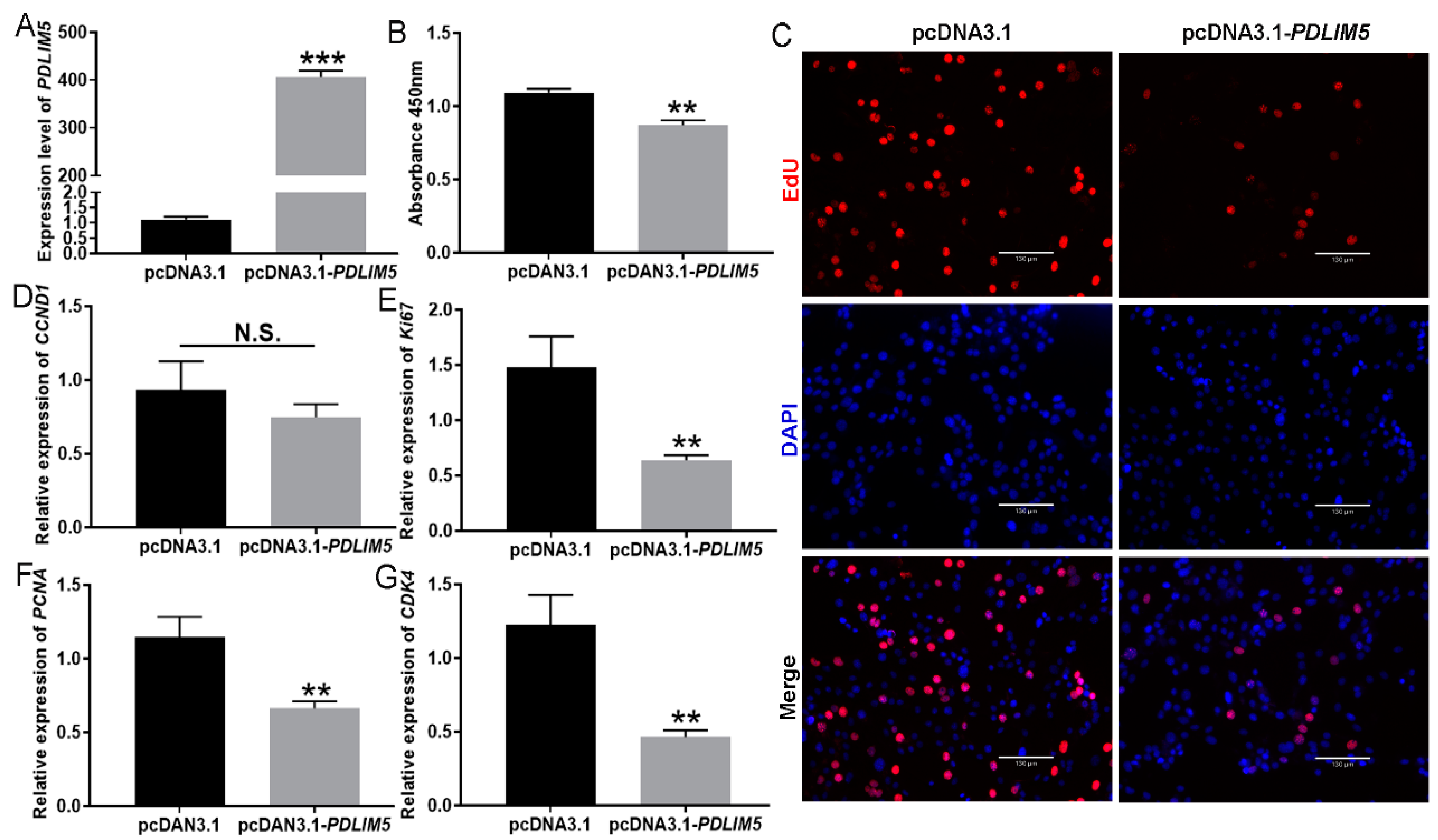
Figure 3.
PDLIM5 suppresses myogenic differentiation. (A–C) Expression of myogenic genes (MyoG, MyoD, and MyHC) at the transcript level. Porcine myoblasts were transfected with pcDNA3.1 or PDLIM5-short overexpression plasmid, and then induced to differentiate for 0, 2, and 4 d (D0, D2, and D4). (D,E) Representative images of MyHC staining for myoblasts after pcDNA3.1-PDLIM5 transfection. Transfected cells cultured with differentiation medium for 3 and 6 d. Blue indicates nuclei stained with DAPI; red indicates MyHC-positive myotubes; scale bar = 130 µm. The data represent the mean ± SD of three independent experiments. GAPDH has been used as a reference gene. * p < 0.05, ** p < 0.01.
Figure 3.
PDLIM5 suppresses myogenic differentiation. (A–C) Expression of myogenic genes (MyoG, MyoD, and MyHC) at the transcript level. Porcine myoblasts were transfected with pcDNA3.1 or PDLIM5-short overexpression plasmid, and then induced to differentiate for 0, 2, and 4 d (D0, D2, and D4). (D,E) Representative images of MyHC staining for myoblasts after pcDNA3.1-PDLIM5 transfection. Transfected cells cultured with differentiation medium for 3 and 6 d. Blue indicates nuclei stained with DAPI; red indicates MyHC-positive myotubes; scale bar = 130 µm. The data represent the mean ± SD of three independent experiments. GAPDH has been used as a reference gene. * p < 0.05, ** p < 0.01.
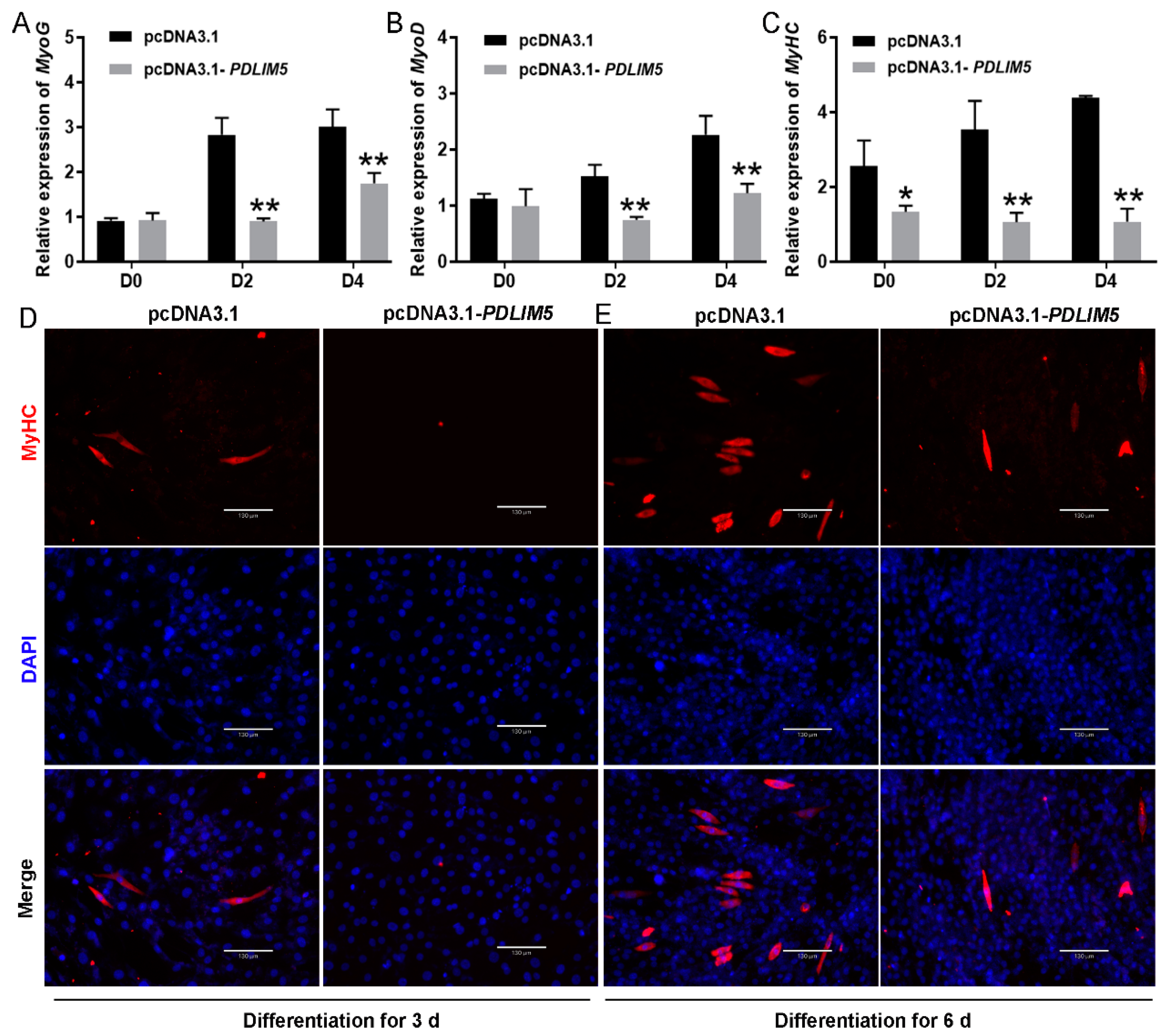
Figure 4.
SNP sites and promoter activity analysis. (A) Comparison of the activities of double luciferase vectors expressed after transfection of C2C12 cells with different fragment lengths. (B) Sequencing results of −521 bp ± 86 bp double luciferase active vectors constructed with four different haplotypes (TG, AG, TT, and AT). Arrows indicate the sites of two SNPs, −258 A > T (red) and −191 T > G (green). (C) Dual-luciferase analysis for promoter activity of four haplotype double luciferase vectors in −521 bp ± 86 bp. Each bar represents the mean ± SD of six independent experiments. Different capital letters represent significant differences, p < 0.01.
Figure 4.
SNP sites and promoter activity analysis. (A) Comparison of the activities of double luciferase vectors expressed after transfection of C2C12 cells with different fragment lengths. (B) Sequencing results of −521 bp ± 86 bp double luciferase active vectors constructed with four different haplotypes (TG, AG, TT, and AT). Arrows indicate the sites of two SNPs, −258 A > T (red) and −191 T > G (green). (C) Dual-luciferase analysis for promoter activity of four haplotype double luciferase vectors in −521 bp ± 86 bp. Each bar represents the mean ± SD of six independent experiments. Different capital letters represent significant differences, p < 0.01.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Gene and genotype frequencies in −258 A > T and −191 T > G in different pig breeds.
| Loci | Breed | Sample Size | Genotype Frequency | Allele Frequency | χ2 Value (p-Value) | |||
|---|---|---|---|---|---|---|---|---|
| −258 A > T | AA | AT | TT | A | T | |||
| DSE | 16 | 0.688 | 0.187 | 0.125 | 0.781 | 0.219 | 0.881 (0.664) | |
| TP | 34 | 0.471 | 0.353 | 0.176 | 0.647 | 0.353 | 1.105 (0.576) | |
| LW | 24 | 1 | 0 | 0 | 1 | 0 | / | |
| −191 T > G | TT | TG | GG | T | G | |||
| DSE | 16 | 0.750 | 0.188 | 0.062 | 0.844 | 0.156 | 1.155 (0.561) | |
| TP | 34 | 0.059 | 0.412 | 0.529 | 0.265 | 0.735 | 0.022 (0.989) | |
| LW | 24 | 0.916 | 0.042 | 0.042 | 0.938 | 0.063 | 2.023 (0.364) | |
Table 2.
Analysis of the effects of the different genotypes of the PDLIM5 gene in NHP pigs.
| Loci | Genotype (Sample Size) | Days to 30 kg | Days to 90 kg |
|---|---|---|---|
| −258 A > T | AA (n = 78) | 92.15 ± 4.91 C | 183.53 ± 6.21 C |
| AT (n = 16) | 96.36 ± 4.84 B | 190.99 ± 5.93 B | |
| TT (n = 3) | 107.07 ± 7.32 A | 202.93 ± 6.32 A | |
| −191 T > G | TT (n = 74) | 91.85 ± 5.73 C | 183.57 ± 6.55 C |
| TG (n = 19) GG (n = 4) | 96.26 ± 5.13 B 105.11 ± 7.45 A | 191.16 ± 4.62 B 201.01 ± 6.41 A |
Different capital letters represent significant differences among genotype groups (p < 0.01).
Table 3.
Joint analysis of the effects of −258 A > T and −191 T > G loci on growth traits in NHPs.
| Genotype (Sample Size) | Days to 30 kg | Days to 90 kg |
|---|---|---|
| TG/TG (n = 3) | 107.08 ± 7.73 A | 202.93 ± 6.27 A |
| AT/AG (n = 9) | 95.14 ± 5.49 B,C | 189.78 ± 6.56 A,B |
| AT/AT (n = 69) | 91.62 ± 5.89 C | 183.33 ± 5.53 B |
| AT/TT (n = 5) | 94.59 ± 4.61 B,C | 190.99 ± 7.13 A,B |
| AT/TG (n = 10) | 97.27 ± 4.25 A,B | 190.66 ± 4.82 A,B |
| AG/TG (n = 1) | 99.19 | 195.24 |
All sample data are expressed as mean ± SD. Statistical significances of AG/TG groups with only one individual were not analyzed.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fu, Y.; Li, S.; Nie, J.; Yan, D.; Zhang, B.; Hao, X.; Zhang, H. Expression of PDLIM5 Spliceosomes and Regulatory Functions on Myogenesis in Pigs. Cells 2024, 13, 720. https://doi.org/10.3390/cells13080720
AMA Style
Fu Y, Li S, Nie J, Yan D, Zhang B, Hao X, Zhang H. Expression of PDLIM5 Spliceosomes and Regulatory Functions on Myogenesis in Pigs. Cells. 2024; 13(8):720. https://doi.org/10.3390/cells13080720
Chicago/Turabian StyleFu, Yu, Shixin Li, Jingru Nie, Dawei Yan, Bo Zhang, Xin Hao, and Hao Zhang. 2024. "Expression of PDLIM5 Spliceosomes and Regulatory Functions on Myogenesis in Pigs" Cells 13, no. 8: 720. https://doi.org/10.3390/cells13080720
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.