TRPV Channels in Mast Cells as a Target for Low-Level-Laser Therapy

1
Shanghai University of Traditional Chinese Medicine and Shanghai Research Center for Acupuncture and Meridians, Shanghai 201203, China
2
Department of Mechanics and Engineering Science, Fudan University Shanghai, Shanghai 201203, China
3
Institute for Biophysics, Goethe-University Frankfurt am Main, 60438 Frankfurt, Germany
*
Author to whom correspondence should be addressed.
Cells 2014, 3(3), 662-673; https://doi.org/10.3390/cells3030662
Submission received: 5 May 2014
/
Revised: 9 June 2014
/
Accepted: 17 June 2014
/
Published: 26 June 2014
(This article belongs to the Special Issue Transient Receptor Potential (TRP) Channels)
Abstract
:Low-level laser irradiation in the visible as well as infrared range is applied to skin for treatment of various diseases. Here we summarize and discuss effects of laser irradiation on mast cells that leads to degranulation of the cells. This process may contribute to initial steps in the final medical effects. We suggest that activation of TRPV channels in the mast cells forms a basis for the underlying mechanisms and that released ATP and histamine may be putative mediators for therapeutic effects.
1. Introduction
Various physical stimuli have been applied to the body surface for medical treatment. These treatments also include low-level laser therapy (LLLT) that uses irradiation in the mW/cm2 range of visible and near-infrared (NIR) light of 600 to 1000 nm (see, e.g., [1]). In this range of wavelengths, absorption within the human skin is very low [2,3,4], and the light can penetrate the tissue in the millimeter range [5]. Heat production by absorption is assumed to play a marginal role. At an output power in the mW range power densities of 1–5000 mW/cm2 were reported to be effective in modulation of inflammation [2,4,6]. Also for wound healing, in treatment of osteo- and rheumatoid arthritis and pain relief LLLT is applied (see, e.g., [4,7,8,9]. In particular red laser light had been introduced into Chinese medicine as laser acupuncture (see, e.g., [10]). Acupuncture points are characterized by a high density of mast cells, and stimulation by conventional needling leads to mast-cell degranulation, which seems to be an essential initial step in acupuncture-induced analgesia [11]. The mast-cell degranulation in acupuncture points cannot only be induced by mechanical stress, but also by high temperature which is used in Chinese medicine during application of moxibustion [12]. These results of the combined effects of mechanical stress and high temperature suggest the involvement of activation of TRPV channels that respond to both of these stimuli [13,14]. In fact, mast cells express TRPV1, TRPV2 and TRPV4 [15], and we asked the question whether these channels in general may form a target for LLLT.
2. Results and Discussion
In the following we will review and discuss effects of laser irradiation in the visible, infra-red (IR) as well as the ultra-violet (UV) range on ion channels of the TRPV family; effects of NIR have not been investigated. Since several of the medical effects, including wound healing [16] and anti-inflammatory effects [17], involve mast-cell activation, we will also consider to which extend modulation of TRPV channels are involved in this process. For the human mast-cell line HMC-1 expression of TRPV1, 2 and 4 was demonstrated [15]. We will, therefore, review the effects of laser light on these isoforms of TRPV, and discuss the role of the functional modulation in mast-cell degranulation.
2.1. Effects of Visible Light on TRPV
2.1.1. TRPV1 as Target for Irradiation
We had previously investigated the effect of laser irradiation on TRPV1 [18]. To avoid interference with laser-induced effects on other ion channels of the TRPV family, we used the model system Xenopus oocyte with heterologously expressed TRPV1. The TRPV1-mediated current exhibited in solution with 2 mM Ca2+ a reversal potential of about −30 mV; a similar value of −20 mV has been reported for oocytes with solution containing 0.5 mM Ca2+ [19].
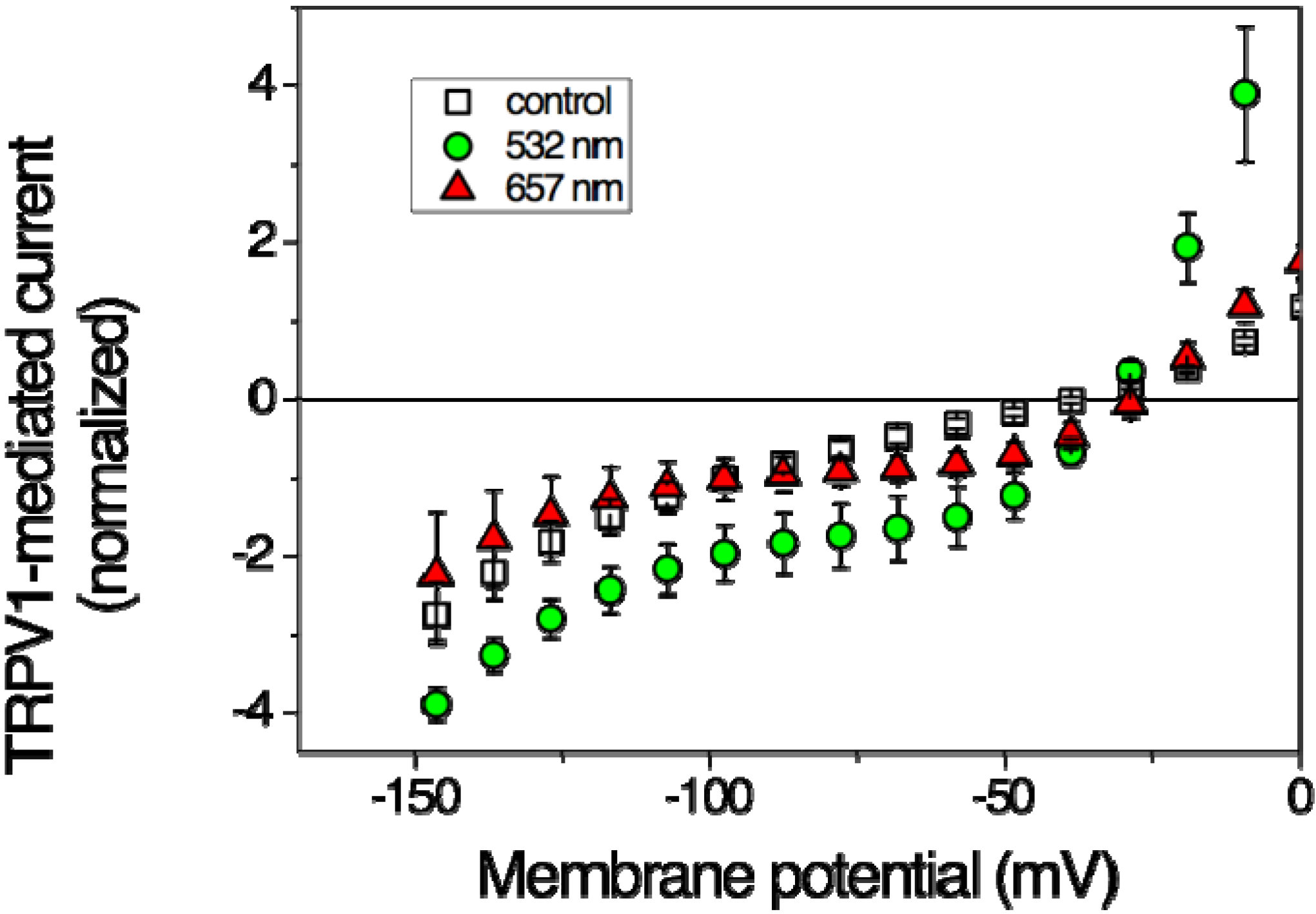
Figure 1 illustrates that TRPV1 can be activated by red laser light. With an output power of 36 mW already after 2-min irradiation a significant current increase could be detected, though in an only limited potential range; at −60 mV the current increased, nevertheless, by a factor of more than 2 (Table 1). The potential-dependent stimulation may indicate modulation of the voltage-dependent gating mechanism. Also, the effects of green laser light were investigated in the oocyte experiments [18]. Irradiation with green light of 532 nm and 40 mW output power produced a strong effect that reached steady-state after 2 min of irradiation over the entire potential range (Figure 1). With an increase by a factor of nearly 5 (at −60 mV) the effect was even more pronounced than that produced by the red light (Table 1).
Figure 1.
Effects of laser light on steady-state current-voltage curves of TRPV1-mediated current in Xenopus oocytes. Open squares represent data in the absence of irradiation (control), filled symbols at the end of a 2-min-lasting irradiation period of 637 nm (triangles up, 36 mW) and of 532 nm (circles, 40 mW), respectively at a power density of <500 mW/cm2. Data are averages of 3–5 oocytes ± SEM. (Based on data from Gu et al. 2012 [18]).
Figure 1.
Effects of laser light on steady-state current-voltage curves of TRPV1-mediated current in Xenopus oocytes. Open squares represent data in the absence of irradiation (control), filled symbols at the end of a 2-min-lasting irradiation period of 637 nm (triangles up, 36 mW) and of 532 nm (circles, 40 mW), respectively at a power density of <500 mW/cm2. Data are averages of 3–5 oocytes ± SEM. (Based on data from Gu et al. 2012 [18]).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effects of laser irradiation. Numbers in brackets refer to the respective publications, * is from unpublished work (A. Kutschireiter); + stands for stimulation, − for attenuation, n.a. for no data available.
| Red-Laser Irradiation | Green-Laser Irradiation | IR-Laser Irratiation | UV-Laser Irradiation | |
|---|---|---|---|---|
| Wavelength | 650 nm | 532 nm | 1.8–2.8 µm | ≈300 nm |
| Power density (mW/cm2) | ≤500 | ≤500 | <3 × 106 [20] <7 × 103 [21] | 15 × 103 [22] 0.3 [23] |
| TRPV1 (current increase at −60 mV) | 2.6 ± 0.5 [18] | 4.7 ± 0.2 [18] | − heat avided [20] | + [23] |
| TRPV2 (current increase at −60 mV) | 1.9 ± 0.5 [15] | n.a. | n.a. | n.a. |
| TRPV4 (current increase at −60 mV) | n.a. | ≈3.7 * | + heat induced [21] − heat avided [20] | + [22] |
2.1.2. TRPV2 as Target for Irradiation
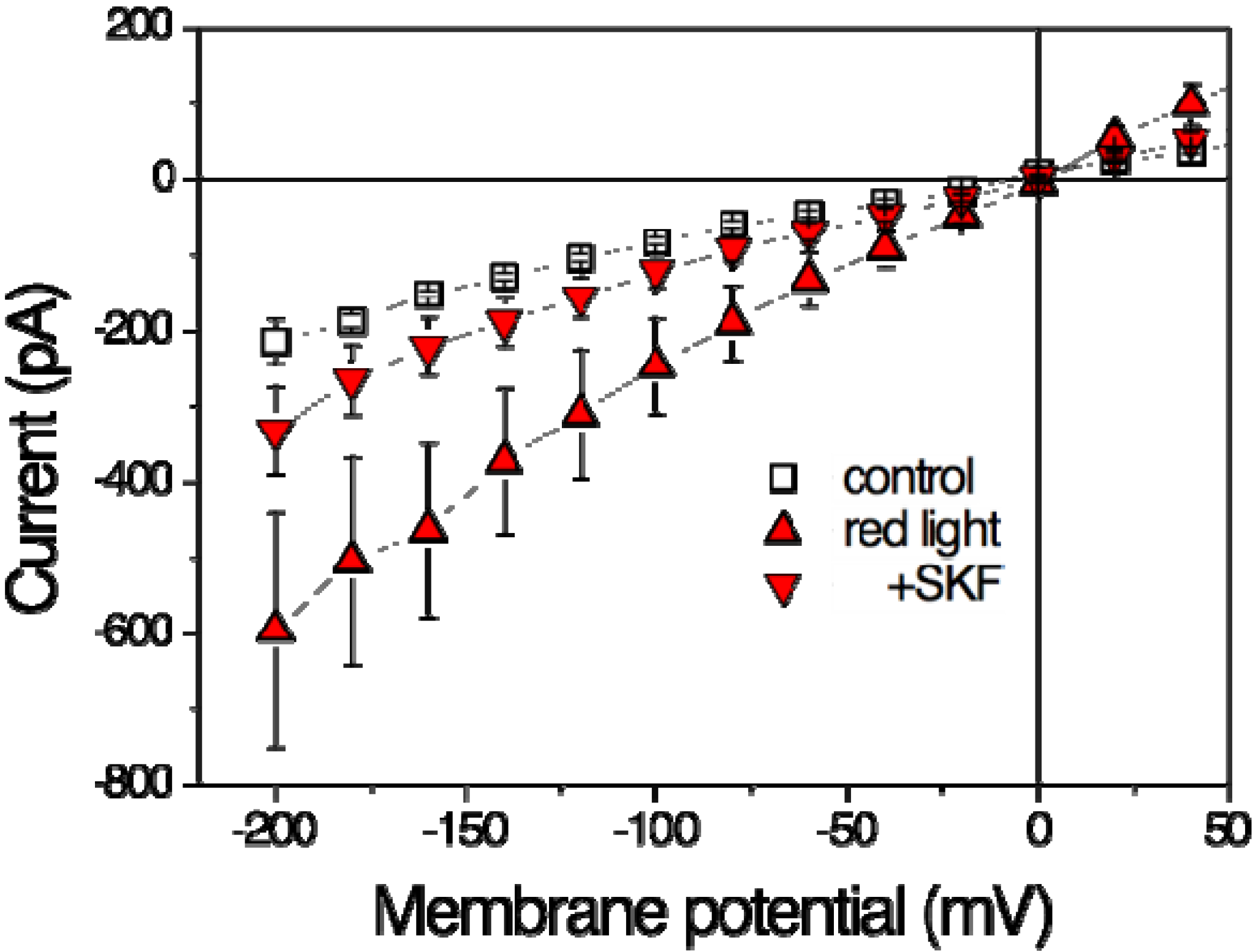
Zhang et al. [15] investigate to what extent TRPV2 could contribute to mast-cell degranulation in response to physical stimuli including laser-light irradiation. Mast cells of the human cell line HMC-1 were exposed to red-laser irradiation of 640 nm with an output power 48 mW [15]. Patch-clamp recordings of steady-state current-voltage dependencies revealed a significant increase (Figure 2) in membrane current during 20 min of irradiation by a factor of about 2 (Table 1).
This current stimulation can be completely blocked by the TRPV inhibitor Ruthenium Red (RuR) [15]. Among the TRPV family, SKF96365 is specific for TRPV2 and can block most, but not all of the current suggesting that the laser-induced increase can be attributed to a large extent to activation of TRPV2 (Figure 2). Nevertheless, there seems to exist another RuR-sensitive current that can be stimulated by red laser light. Above, we have illustrated that at least the current mediated by TRPV1 is also stimulated by red laser light.
Figure 2.
Effects of red laser light on steady-state current-voltage curves in HMC-1 cells (Human Mast Cell line). Whole-cell patch-clamp recording. Open squares represent data in the absence of irradiation (control), filled symbols at the end of a 20-min-lasting irradiation period of 640 nm (48 mW, ~500 mW/cm2) in the absence (triangles up, red light) and presence of 20 µM SKF96365 (triangle down, +SKF). Data are averages of five measurements ± SEM. (Based on data from Zhang et al. 2012 [15]).
Figure 2.
Effects of red laser light on steady-state current-voltage curves in HMC-1 cells (Human Mast Cell line). Whole-cell patch-clamp recording. Open squares represent data in the absence of irradiation (control), filled symbols at the end of a 20-min-lasting irradiation period of 640 nm (48 mW, ~500 mW/cm2) in the absence (triangles up, red light) and presence of 20 µM SKF96365 (triangle down, +SKF). Data are averages of five measurements ± SEM. (Based on data from Zhang et al. 2012 [15]).

2.1.3. TRPV4 as Target for Irradiation
Blue laser (405 nm) or green laser (532 nm) irradiation at about 150 mW/cm2 induced histamine release from rat basophilic leukemia (RBL-2H3) cells [24]. The non-specific inhibitor RuR for TRPV channels significantly blocked the irradiation-induced histamine release. Furthermore, immunocytochemical staining of TRPV4 increased after laser irradiation. The authors, therefore, conclude that TRPV4 stimulation was involved in the process of histamine release.
In Xenopus oocytes activation of TRPV4-mediated current by green laser light could be confirmed (A. Kutschireiter, unpublished); in orientating experiments with green laser light (532 nm, 40 mW, about 500 mW/cm2) a pronounced stimulation to a constant value within less than a minute was observed (Table 1).
2.2. Effects of IR Light on TRPV
In contrast to visible red light, IR light is absorbed by the tissue and converted to heat. Since the ion channels of the TRPV family are also sensors for high temperature [25], it can be expected that IR light activates TRPV channels. The effect of heat on mast-cell degranulation had been investigated [15], and it could be demonstrated that indeed heat application resulted in degranulation associated with increase intracellular Ca2+ and ATP release [26] (see Section 2.5). In addition, IR laser light (1.875 µm, 320 mW, 2.7 × 106 mW/cm2, 20–60 × 103 mJ/cm2) triggered TRPV4-mediated transient membrane potential variations in retinal ganglion cells of mice and in vestibular cells of rats [21]. An increase of the temperature by about 10 °C occurred during the irradiation and can account for TRPV4 stimulation [21].
Application of IR irradiation (2.780 µm at 260–6500 mW/cm2, 3.98–33.17 × 103 mJ/cm2) could significantly relieve capsaicin- and hypotonicity-induced nociception [20]. Furthermore, capsaicin- or hypotonicity-induced Ca2+ influx in TRPV1 and TRPV4 over-expressing HeLa cells were attenuated by this laser irradiation. These results suggest that TRPV1 and TRPV4 are inhibited by IR irradiation [20]. In this study, water was sprayed during irradiation to avoid skin burn, and the authors claimed no obvious temperature changes in their in vitro tests because of the perfusion of bath solution [20]. Therefore, the modulation of TRPV1 and TRPV4 by laser irradiation cannot be attributed to heating.
2.3. Effects of UV Light on TRPV
Solar ultraviolet (UV) irradiation is a major cause of premature aging of skin. Lee et al. [23] confirmed that UV irradiation (300–320 nm, 0.3 mW/cm2, 36 mJ/cm2) induced slow and persistent membrane current and Ca2+ influx in HaCaT cells, a human keratinocyte cell line. This current was inhibited by TRPV1 inhibitors (capsazepine and RuR). The UV-induced matrix metalloproteinase-1 (MMP-1) was decreased by TRPV1 inhibitors and was facilitated by capsaicin. Knock-down of TRPV1 using siRNA transfection also decreased MMP-1 expression as well as UV-induced Ca2+ influx in HaCaT cells. Another non-selective cation channel highly expressed in epithelial skin cell is TRPV4. Moore et al. [22] showed that Ca2+ influx into keratinocytes in response to UVB (295 nm, 15 × 103 mW/cm2, 600 mJ/cm2) depends on TRPV4. External topical application of a TRPV4-selective inhibitor (GSK205) attenuates UVB-evoked nocifensive behaviour and endothelin-1 expression, and keratinocyte-derived TRPV4 was suggested as a therapeutic target for UVB-induced sunburn.
2.4. TRPV Modulation and Mast-Cell Activation
In the above sections we illustrated and discussed that laser irradiation in the visible as well as IR and UV range results in the activation of TRPV channels, in particular of TRPV1, 2 and 4 seemed to be effected.
The activation of membrane current by red laser light is accompanied by degranulation of the mast cells [15,18,27]. Patch-clamp experiments revealed that the degranulation becomes also apparent by an increase in membrane capacitance [15]. Nevertheless, the observed increase in membrane surface by about 40% cannot account for the increase in membrane current by a factor of 2 (see Table 1) indicating that the red laser light stimulates TRPV2 channel activity.
2.5. Effect of Laser Irradiation on Intracellular Ca2+ Activity and ATP Release Mobilisation
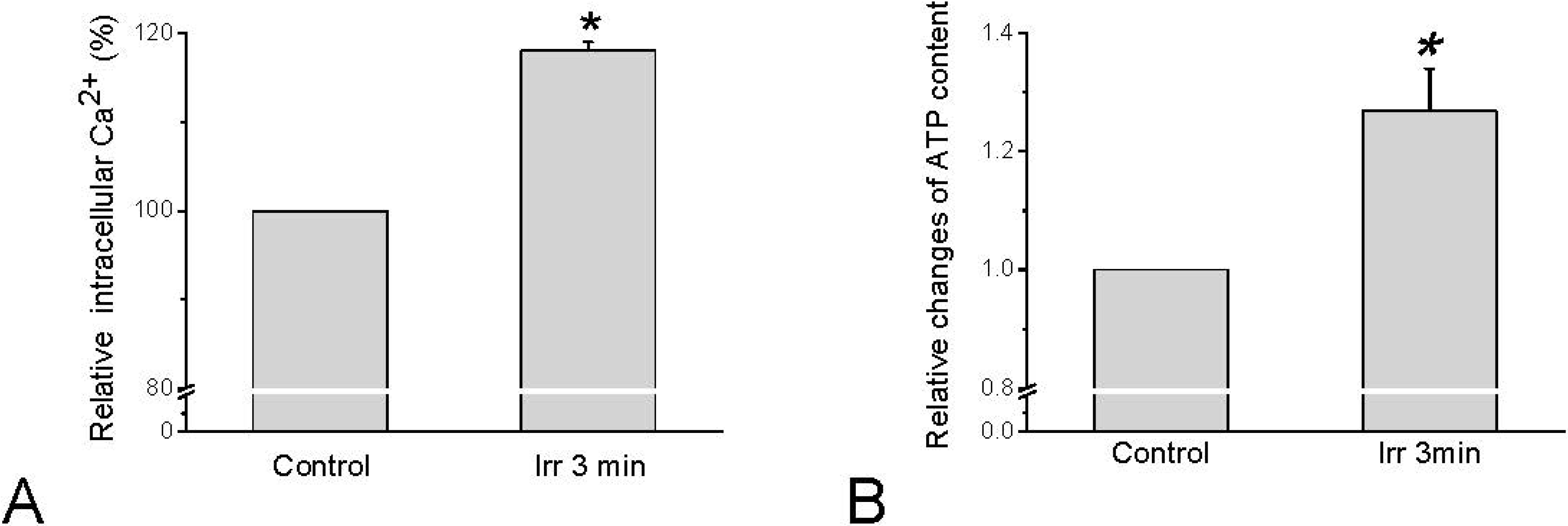
Since TRPV channels are permeable for Ca2+, Ca2+ influx can be expected to occur in response to TRPV channel activation. Concerning HMC-1 cells, intracellular calcium activity ([Ca2+]i) indeed increased in response to irradiation by red laser light of 657 nm (280 mW/cm2, 54 × 103 mJ/cm2) (Figure 3A). Interestingly, this elevation of [Ca2+]i was independent on extracellular Ca2+. With the same irradiation parameters that stimulate [Ca2+]i increase an elevation of extracellular ATP could be detected (Figure 3B), and the [Ca2+]i was demonstrated to trigger exocytotic release of ATP [32].
Figure 3.
Changes in [Ca2+]i and extracellular ATP in HMC-1 cells in response to 657 nm and 280 mW/cm2 laser irradiation for 3 min. (A) Quantitative analysis of several sets of cells normalized to basal [Ca2+]i-dependent fluorescence before irradiation (=100%). Data represent averages ± SEM (n = 15 cells, * p < 0.01 compared to control); (B) Relative changes of extracellular ATP averaged from all sample. The data represent averages ± SEM (n = 27, * p < 0.01 compared to control, and are based on Wang et al. 2013 [33]).
Figure 3.
Changes in [Ca2+]i and extracellular ATP in HMC-1 cells in response to 657 nm and 280 mW/cm2 laser irradiation for 3 min. (A) Quantitative analysis of several sets of cells normalized to basal [Ca2+]i-dependent fluorescence before irradiation (=100%). Data represent averages ± SEM (n = 15 cells, * p < 0.01 compared to control); (B) Relative changes of extracellular ATP averaged from all sample. The data represent averages ± SEM (n = 27, * p < 0.01 compared to control, and are based on Wang et al. 2013 [33]).

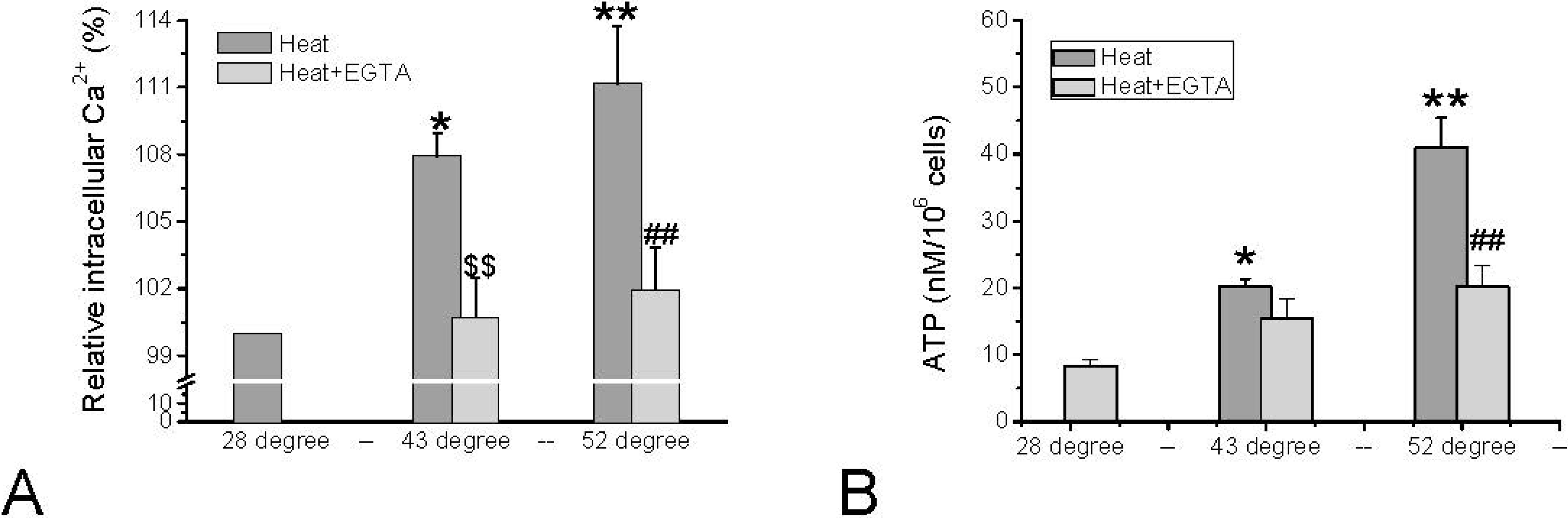
The thermal activation of TRPV1 and TRPV2 is characterized by threshold temperatures of 43 °C and 52 °C, respectively. These noxious temperatures resulted in TRPV1- and TRPV2-dependent influx of Ca2+ in HMC-1 leading to elevated [Ca2+]i (Figure 4A). Only exceeding a temperature of 43 °C increased [Ca2+]i, and a further significant increase was observed when exceeding 52 °C. The relative changes in intracellular calcium activity were monitored by measuring the [Ca2+]i–dependent fluorescence of Calcium Green. The fluorescence signal of Calcium Green was reported to be temperature-dependent [34,35], but at the temperatures exceeding 28 °C the signal is hardly affected, and the changes illustrated in Figure 4A will not significantly be altered. As was illustrated for stimulation of mast cells by red laser light (see above), the temperature-induced elevation of [Ca2+]i triggers release of ATP, which becomes apparent in elevated extracellular ATP content (Figure 4B).
The increase in [Ca2+]i as well as in extracellular ATP induced by the noxious temperatures can partially be blocked by the Ca2+ chelator EGTA suggesting that influx of Ca2+ via TRPV1 and TRPV2 is involved (Figure 4B). In fact, inhibition of TRPV2 by 20 µM SKF96365 could partially abolish the increase of the extracellular ATP level induced by 52 °C by 27.9% ± 10.0% (N = 3 independent experiments (unpublished)).
Figure 4.
Changes in [Ca2+]i and extracellular ATP in HMC-1 cells in response to noxious heat that activates TRPV1 (43 °C) and TRPV2 (52 °C), and either in the absence (dark grey bars) or presence (light grey bars) of 5 mM EGTA. (A) Quantitative analysis from several sets of experiments normalized to basal [Ca2+]i–dependent fluorescence at 28 °C (=100% of Calcium Green-1 loaded HMC-1 cells). Data represent averages ± SEM from N = 3–4 independent experiments (* p < 0.05, ** p < 0.01, compared to 28 °C; $$ p < 0.01 compare to 43 °C, ## p < 0.01, compared to 52 °C); (B) ATP content in cell suspension after 3 min of incubation at the respective temperatures. Data represent averages ±SEM from n = 6–42 samples of N = 3–11 independent experiments (* p = < 0.05 and ** p < 0.0001, compared to control; ## p < 0.0001, compare to 52 °C). Data in (A) are unpublished, data in (B) are based on Hu et al. 2014 [26].
Figure 4.
Changes in [Ca2+]i and extracellular ATP in HMC-1 cells in response to noxious heat that activates TRPV1 (43 °C) and TRPV2 (52 °C), and either in the absence (dark grey bars) or presence (light grey bars) of 5 mM EGTA. (A) Quantitative analysis from several sets of experiments normalized to basal [Ca2+]i–dependent fluorescence at 28 °C (=100% of Calcium Green-1 loaded HMC-1 cells). Data represent averages ± SEM from N = 3–4 independent experiments (* p < 0.05, ** p < 0.01, compared to 28 °C; $$ p < 0.01 compare to 43 °C, ## p < 0.01, compared to 52 °C); (B) ATP content in cell suspension after 3 min of incubation at the respective temperatures. Data represent averages ±SEM from n = 6–42 samples of N = 3–11 independent experiments (* p = < 0.05 and ** p < 0.0001, compared to control; ## p < 0.0001, compare to 52 °C). Data in (A) are unpublished, data in (B) are based on Hu et al. 2014 [26].

2.6. Underlying Mechanisms for Laser-Light-Induced TRPV Modulation
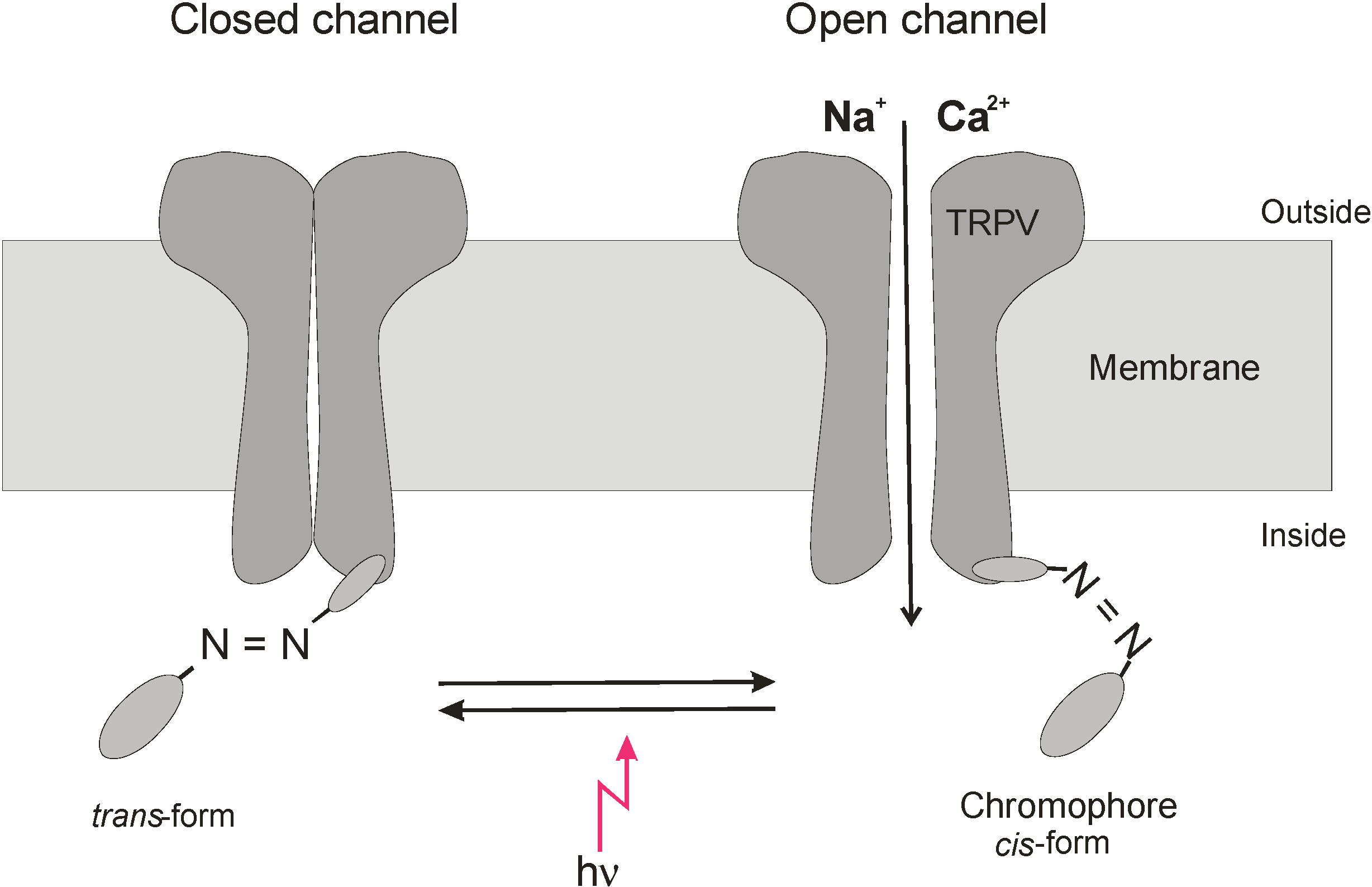
Direct modulation of TRPV channels by absorption of visible and NIR light can hardly be expected, but the existence of various chromophores has been reported that may act as switches to modulate the function of membrane proteins [1,36,37,38,39,40,41]. Laser irradiation could induce the histamine release in RBL-2H3 mast cells dependent on TRPV4 activation [20], and its wavelength for activation of the histamine release was consistent with the absorption bands of cytochrome c oxidase [42]. Activation of cytochrome c oxidase has been demonstrated to be involved in light-induced stimulation of the Na, K-ATPase [36] and tissue healing [43,44]. The light-induced stimulation of TRPV ion channels we observed in our experiments developed within a few minutes (see, e.g., [18]); therefore, we consider the involvement of cytochrome c oxidase not as a dominating process. It could be demonstrated that light can modulate in channel activity of TRPs by isomerising photochromic antagonists between trans state and cis form [37]. In particular, azobenzenes were reported to act as photoswitches for a variety of biomolecules including membrane proteins [41]. The azo-benzene-derivative AC4 of the TRPV1 antagonist capsazepine, for example, exists at 440 nm in its trans and at 360 nm in its cis configuration, and depending on the stimulus for TRPV1 activation the trans or cis configuration can reversibly act as antagonist (comp. Figure 5). Also, other members of the TRP family are reported to be involved in phototransduction. For example, TRPA1 was demonstrated to be essential for extraocular pathway in human melanocytes when exposed to UV radiation. The photocurrent is mediated by G protein-coupled receptors and leads to rapid Ca2+ mobilization [38]; also, in this process, photosensitive agents are involved. TRPA1was also reported to be activated by UV light in HEK293 cells [39]. [Ca2+]i as well as whole-cell currents and open probability of TRPA1 were potentiated by 350 nm UVA light at power density of 580 mW/cm2 and doses of 34.8–104.4 × 103 mJ/cm2. It is interesting to note that photosensitising agents could shift light activation of TRPA1 to longer wavelengths of 490 nm and 590 nm [39]. Whether a light-switchable physiological chromophore can account for the observed stimulation of the ion channel activity of TRPV needs further investigation.
Figure 5.
Modulation of TRPV channel gating by light-switched ligand. Putative modulation of an azo-chromophore between cis- and trans-form by light leading to activation of TRPV channel opening. As an example TRPV activation by the cis-form is cartooned.
Figure 5.
Modulation of TRPV channel gating by light-switched ligand. Putative modulation of an azo-chromophore between cis- and trans-form by light leading to activation of TRPV channel opening. As an example TRPV activation by the cis-form is cartooned.

The possible mechanism for TRP gating may involve direct binding by lipids such as diacylglycerol and polyunsaturated fatty acids at specific sites on the channels proteins [40]. Single-channel currents could be evoked by light in rhabodomere-attached patches from intact isolated photoreceptors of wt Drosophila, while mutants lacking TRP and TRP-like proteins are devoid of light-inducible channel activity. For TRPV4, it could be demonstrated that activation by hypotonicity or heat requires interaction with phosphatidylinositol-biphosphate [45]; in general, ligand-dependent activation of thermo-sensitive TRP channels has been discussed [43].
The effects of irradiation we have summarized in this review and that led to activation of TRPVs and/or mast cell degranulation included results mainly obtained with laser irradiation (see also [36]), but also light from light-emitting diodes (e.g., [23]), filtered light or even broad-band irradiation (e.g., [30,31,38]). Since similar effects were observed, coherence of light seems not to be essential for the observed effects.
3. Conclusions
We have shown in this review that laser irradiation in the visible and IR as well as UV range can modulate the function and expression of TRPV ion channels, and in particular TRPV1, TRPV2, and TRPV4. This may form the basis for effect of LLLT. As Ca2+-permeable ion channels, their activation may contribute to the laser-induced increase in intracellular Ca2+ that triggers degranulation and endocytotic release of ATP. Such light-induced mechanism may contribute to the basis of the medical effects of LLLT. This hypothesis still needs confirmation in animal tests and clinical trials.
Acknowledgments
The work was supported by the National Basic Research Program of China (973 Program, 2012CB518502) and the National Natural Science Foundation of China (No. 81102635 and No. 81102630). We also gratefully acknowledge the support from Green Valley Holding Co, Shanghai.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Karu, T.I.; Kolyakov, S.F. Exact action spectra for cellular responses relevant to phototherapy. Photomed. Laser Surg. 2005, 23, 355–361. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Chen, A.C.H.; Carroll, J.D.; Hamblin, M.R. Biphasic dose response in ow level lightherapy. Dose-Response 2009, 7, 358–383. [Google Scholar] [CrossRef]
- Peng, Q.; Juzeniene, A.; Chen, J.; Svaasand, L.O.; Warloe, T.; Giercksky, K.-E.; Moran, J. Lasers in medicine. Rep. Prog. Phys. 2008, 71, 1–28. [Google Scholar]
- Avci, P.; Gupta, A.; Sadasivam, M.; Vecchio, D.; Pam, Z.; Hamblin, M.R. Low-level laser (light) lherapy (LLLT) in Skin: Stimulating, lealing, restoring. Semin. Cutan. Med. Surg. 2013, 32, 41–52. [Google Scholar]
- Whittaker, P. Laser acupuncture: Past, present, and future. Lasers Med. Sci. 2004, 19, 69–80. [Google Scholar] [CrossRef]
- Bjordal, J.M.; Johnson, M.I.; Iverson, L.E.; Aimbire, F.; Lopes-Marins, R.A.B. Low-level laser therapy in acute pain: A systematic review of possible mechanisms of action and clinical effects in randomized placebo-controlled trials. Photomed. Laser Surg. 2006, 24, 158–168. [Google Scholar] [CrossRef]
- Ay, S.; Doğan, S.K.; Evcik, D. Is low-level laser therapy effective in acute or chronic low back pain? Clin. Rheumatol. 2010, 29, 905–910. [Google Scholar]
- Shen, X.; Zhao, L.; Ding, G.; Tan, M.; Gao, J.; Wang, L.; Lao, L. Effect of combined laser acupuncture on knee osteoatthitis: A pilot study. Laser Med. Sci. 2013, 24, 129–136. [Google Scholar]
- Alves, A.C.; de Carvalho, P.T.; Parente, M.; Xavier, M.; Frigo, L.; Aimbire, F.; Leal Junior, E.C.; Albertini, R. Low-level laser therapy in different stages of rheumatoid arthritis: A histological study. Lasers Med. Sci. 2013, 28, 529–536. [Google Scholar] [CrossRef]
- Litscher, G.; Gao, X.Y.; Wang, L.; Zhu, B. (Eds.) High-Tech Acupuncture and Integrative Laser Medicine; Pabst Science Publishers: Lengerich, Germany, 2012.
- Zhang, D.; Ding, G.; Shen, X.; Yao, W.; Zhang, Z.; Zhang, Y.; Gu, Q. Role of mast cells in acupuncture effects: A pilot study. Explore 2008, 4, 170–177. [Google Scholar] [CrossRef]
- Wang, Y.S.; Zhang, J.B.; Jiang, J.F.; Wang, L.L. Research on effects of the thermal stimulation by moxibustion at different temperatures on cardiac function in rats and on mast cells in the local site of moxibustion. Evid.-Based Complement. Altern. Med. 2013, 2013, 545707. [Google Scholar] [CrossRef]
- Wu, L.J.; Sweet, T.B.; Clapham, D.E. International Union of Basic and Clinical Pharmacology. LXXVI. Current progress in the mammalian TRP ion channel family. Pharmacol. Rev. 2010, 62, 381–404. [Google Scholar] [CrossRef]
- Vennekens, R.; Owsianik, G.; Nilius, B. Vanilloid transient receptor potential cation channels: An overview. Curr. Pharm. Des. 2008, 14, 18–31. [Google Scholar] [CrossRef]
- Zhang, D.; Spielmann, A.; Wang, L.; Ding, G.; Huang, F.; Gu, Q.; Schwarz, W. Mast-cell degranulation induced by physicsl stimuli involves the activation of transient-receptor-potential channel TRPV2. Physiol. Res. 2012, 61, 113–124. [Google Scholar]
- Pereira, M.C.; de Pinho, C.B.; Medrado, A.R.; Andrade, Z.A.; Reis, S.R. Influence of 670 nm low-level laser therapy on mast cells and vascular response of cutaneous injuries. J. Photochem. Photobiol. B 2010, 98, 188–192. [Google Scholar] [CrossRef]
- Marinho, R.R.; Matos, R.M.; Santos, J.S.; Ribeiro, M.A.; Smaniotto, S.; Barreto, E.O.; Ribeiro, R.A.; Lima, R.C.; Albuquerque, R.L.; Thomazzi, S.M. Potentiated anti-inflammatory effect of combined 780 nm and 660 nm low level laser therapy on the experimental laryngitis. J. Photochem. Photobiol. B 2013, 121, 86–93. [Google Scholar] [CrossRef]
- Gu, Q.B.; Wang, L.N.; Huang, F.; Schwarz, W. Stimulation of TRPV1 by green laser light. Evid.-Based Complement. Altern. Med. 2012. [Google Scholar] [CrossRef]
- Rivera-Acevedo, R.E.; Pless, S.A.; Schwarz, S.K.W.; Ahern, C.A. Extracellular quaternary ammonium blockade of transient receptor potential vanilloid subtype 1 channels xxpressed in Xenopus laevis oocytes. Mol. Pharmacol. 2012, 82, 1129–1135. [Google Scholar] [CrossRef]
- Ryu, J.J.; Yoo, S.; Kim, K.Y.; Park, J.S.; Bang, S.; Lee, S.H.; Yang, T.J.; Cho, H.; Hwang, S.W. Laser modulation of heat and capsaicin receptor TRPV1 leads to thermal antinociception. J. Dent. Res. 2010, 89, 1455–1460. [Google Scholar] [CrossRef]
- Albert, E.S.; Bec, J.M.; Desmadryl, G.; Chekroud, K.; Travo, C.; Gaboyard, S.; Bardin, F.; Marc, I.; Dumas, M.; Lenaers, G.; et al. TRPV4 channels mediate the infrared laser-evoked response in sensory neurons. J. Neurophysiol. 2012, 107, 3227–3234. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.; Cevikbas, F.; Pasolli, H.A.; Chen, Y.; Kong, W.; Kempkes, C.; Parekh, P.; Lee, S.H.; Kontchou, N.A.; Yeh, I.; et al. UVB radiation generates sunburn pain and affects skin by activating epidermal TRPV4 ion channels and triggering endothelin-1 signaling. Proc. Natl. Acad. Sci. USA 2013, 110, E3225–E3234. [Google Scholar]
- Lee, Y.M.; Kim, Y.K.; Kim, K.H.; Park, S.J.; Kim, S.J.; Chung, J.H. A novel role for the TRPV1 channel in UV-induced matrix metalloproteinase (MMP)-1 expression in HaCaT cells. J. Cell. Physiol. 2009, 219, 766–775. [Google Scholar] [CrossRef]
- Yang, W.-Z.; Chen, Y.; Yu, J.-T.; Zhou, L.-W. Effects of lower power laser irradiation on intracellular calcium and histamine release in RBL-2H3 mast cells. Photochem. Photobiol. 2007, 83, 979–984. [Google Scholar] [CrossRef]
- Benham, C.D.; Gunthorpe, M.J.; Davis, J.B. TRPV channels as temperature sensors. Cell Calcium 2003, 33, 479–487. [Google Scholar] [CrossRef]
- Hu, L.; Wang, L.N.; Grygorczyk, R.; Shen, X.Y.; Schwarz, W. Heat induces ATP release from mast cells in vitro: A putative mechnism for moxibustion. J. Tradit. Chin. Med. 2014. accepted. [Google Scholar]
- Wang, L.; Schwarz, W. Activation of mast cells by acupuncture stimuli. Forum Immunopathol. Dis. Ther. 2012, 3, 41–50. [Google Scholar]
- Berbert, F.L.; Sivieri-Araujo, G.; Ramalho, L.T.; Pereira, S.A.; Rodrigues, D.B.; de Araujo, M.S. Quantification of fibrosis and mast cells in the tissue response of endodontic sealer irradiated by low-level laser therapy. Lasers Med. Sci. 2011, 26, 741–747. [Google Scholar] [CrossRef]
- Silveira, L.B.; Prates, R.A.; Novelli, M.D.; Marigo, H.A.; Garrocho, A.A.; Amorim, J.C.; Sousa, G.R.; Pinotti, M.; Ribeiro, M.S. Investigation of mast cells in human gingiva following low-intensity laser irradiation. Photomed. Laser Surg. 2008, 26, 315–321. [Google Scholar] [CrossRef]
- Zhou, Y.D.; Fang, X.F.; Cui, Z.J. UVA-induced calcium oscillations in rat mast cells. Cell Calcium 2009, 45, 18–28. [Google Scholar] [CrossRef]
- Sarchio, S.N.; Kok, L.F.; O’Sullivan, C.; Halliday, G.M.; Byrne, S.N. Dermal mast cells affect the development of sunlight-induced skin tumours. Exp. Dermatol. 2012, 21, 241–248. [Google Scholar] [CrossRef]
- Boudreault, F.; Grygorczyk, R. Cell swelling-induced ATP release is tightly dependent on intracellular calcium elevations. J. Physiol. 2004, 561, 499–513. [Google Scholar]
- Wang, L.; Sikora, J.; Hu, L.; Shen, X.; Grygorczyk, R.; Schwarz, W. ATP release from mast cells by physical stimulation: A putative early step in activation of acupuncture points. Evid.-Based Complement. Altern. Med. 2013, 2013, 350949. [Google Scholar]
- Eberhard, M.; Erne, P. Calcium binding to fluorescent calcium indicators: Calcium green, calcium orange and calcium crimson. Biochem. Biophys. Res. Commun. 1991, 180, 209–215. [Google Scholar] [CrossRef]
- Oliver, A.E.; Baker, G.A.; Fugate, R.D.; Tablin, F.; Crowe, J.H. Effects of temperature on calcium-sensitive fluorescent probes. Biophys. J. 2000, 78, 2116–2126. [Google Scholar] [CrossRef]
- Karu, T.I. Mitochondrial signaling in mammalian cells activated by red and near-IR radiation. Photochem. Photobiol. 2008, 84, 1091–1099. [Google Scholar] [CrossRef]
- Stein, M.; Breit, A.; Fehrentz, T.; Gudermann, T.; Trauner, D. Optical control of TRPV1 channels. Angew. Chem. Int. Ed. 2013, 52, 9845–9848. [Google Scholar] [CrossRef]
- Bellono, N.W.; Kammel, L.G.; Zimmerman, A.L.; Oancea, E. UV light phototransduction activates transient receptor potential A1 ion channels in human melanocytes. Proc. Natl. Acad. Sci. USA 2013, 110, 2383–2388. [Google Scholar] [CrossRef]
- Hill, K.; Schaefer, M. Ultraviolet light and photosensitising agents activate TRPA1 via generation of oxidative stress. Cell Calcium 2009, 45, 155–164. [Google Scholar] [CrossRef]
- Delgado, R.; Bacigalupo, J. Unitary recordings of TRP and TRPL channels from isolated Drosophila retinal photoreceptor rhabdomeres: Activation by light and lipids. J. Neurophysiol. 2009, 101, 2372–2379. [Google Scholar] [CrossRef]
- Beharry, A.A.; Woolley, G.A. Azobenzene photoswitches for biomolecules. Chem. Soc. Rev. 2011, 40, 4422–4437. [Google Scholar]
- Wu, Z.H.; Zhou, Y.; Chen, J.Y.; Zhou, L.W. Mitochondrial signaling for histamine releases in laser-irradiated RBL-2H3 mast cells. Lasers Surg. Med. 2010, 42, 503–509. [Google Scholar] [CrossRef]
- Steinberg, X.; Lespay-Rebolledo, E.; Brauchi, S. A structural view of ligand-dependent activation in thermoTRP channels. Front. Physiol. 2014, 5, 171. [Google Scholar] [CrossRef]
- Wong-Riley, M.T.T.; Liang, H.L.; Eells, J.T.; Chance, B.; Henry, M.M.; Buchmann, E.; Kane, M.; Whelan, H.T. Photobiomodulation Directly Benefits Primary Neurons Functionally Inactivated by Toxins: Role of Cytochrome c Oxidase. J. Biol. Chem. 2005, 280, 4761–4771. [Google Scholar] [CrossRef]
- Garcia-Elias, A.; Mrkonjic, S.; Pardo-Pastor, C.; Inada, H.; Hellmich, U.A.; Rubio-Moscardo, F.; Plata, C.; Gaudet, R.; Vicente, R.; Valverde, M.A. Phosphatidylinositol-4,5-biphosphate-dependent rearrangement of TRPV4 cytosolic tails enables channel activation by physiological stimuli. Proc. Natl. Acad. Sci. USA 2013, 110, 9553–9558. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Wang, L.; Zhang, D.; Schwarz, W. TRPV Channels in Mast Cells as a Target for Low-Level-Laser Therapy. Cells 2014, 3, 662-673. https://doi.org/10.3390/cells3030662
AMA Style
Wang L, Zhang D, Schwarz W. TRPV Channels in Mast Cells as a Target for Low-Level-Laser Therapy. Cells. 2014; 3(3):662-673. https://doi.org/10.3390/cells3030662
Chicago/Turabian StyleWang, Lina, Di Zhang, and Wolfgang Schwarz. 2014. "TRPV Channels in Mast Cells as a Target for Low-Level-Laser Therapy" Cells 3, no. 3: 662-673. https://doi.org/10.3390/cells3030662