MicroRNAs at the Interface between Osteogenesis and Angiogenesis as Targets for Bone Regeneration


Department of Cranio-Maxillofacial Surgery, University of Münster, Albert-Schweitzer-Campus 1, 48149 Münster, Germany
Cells 2019, 8(2), 121; https://doi.org/10.3390/cells8020121
Submission received: 20 December 2018
/
Revised: 25 January 2019
/
Accepted: 30 January 2019
/
Published: 3 February 2019
(This article belongs to the Special Issue Regulatory microRNA)
Abstract
:Bone formation and regeneration is a multistep complex process crucially determined by the formation of blood vessels in the growth plate region. This is preceded by the expression of growth factors, notably the vascular endothelial growth factor (VEGF), secreted by osteogenic cells, as well as the corresponding response of endothelial cells, although the exact mechanisms remain to be clarified. Thereby, coordinated coupling between osteogenesis and angiogenesis is initiated and sustained. The precise interplay of these two fundamental processes is crucial during times of rapid bone growth or fracture repair in adults. Deviations in this balance might lead to pathologic conditions such as osteoarthritis and ectopic bone formation. Besides VEGF, the recently discovered important regulatory and modifying functions of microRNAs also support this key mechanism. These comprise two principal categories of microRNAs that were identified with specific functions in bone formation (osteomiRs) and/or angiogenesis (angiomiRs). However, as hypoxia is a major driving force behind bone angiogenesis, a third group involved in this process is represented by hypoxia-inducible microRNAs (hypoxamiRs). This review was focused on the identification of microRNAs that were found to have an active role in osteogenesis as well as angiogenesis to date that were termed “CouplingmiRs (CPLGmiRs)”. Outlined representatives therefore represent microRNAs that already have been associated with an active role in osteogenic-angiogenic coupling or are presumed to have its potential. Elucidation of the molecular mechanisms governing bone angiogenesis are of great relevance for improving therapeutic options in bone regeneration, tissue-engineering, and the treatment of bone-related diseases.
1. Introduction
The replacement of large bone defects and the availability of adequate tissue-engineered bone remains a major clinical challenge. This tremendous demand results from the high incidence of large segmental bone defects due to trauma, congenital malformations, ageing, or bone-related diseases such as osteoporosis, inflammation or tumors [1]. However, the development of gene therapy approaches in recent years demonstrated that tissue-engineered bone offers new therapeutic strategies to repair tissue defects. Thereby, one of the major disadvantages in the clinical use of engineered bone constructs so far, i.e., the inability to provide sufficient blood supply in the initial phase after implantation which leads to insufficient cell integration and cell death, could be overcome [2]. As an explanation for this shortcoming, the function of angiogenesis—the process of forming new blood vessels from pre-existing vasculature—in bone regeneration is still poorly defined, and the molecular mechanisms that regulate angiogenesis in bone are only just starting to be unraveled. Angiogenesis, a term that was coined in 1935 to describe the formation of blood vessels in the placenta, occurs during normal vertebrate embryogenesis but also as a response to pathophysiological circumstances during the processes of tumor formation, wound healing and tissue regeneration [3,4]. Elucidating the wide orchestrating variety of signal pathways and stimuli linking angiogenesis and bone formation on the molecular level is therefore of great interest. In this context, the gained information will yield improved bone replacement in fracture healing, or prevention of bone loss in osteoporosis and therapeutically-induced reparative angiogenesis.
Bone, with its main function in supporting and withstanding mechanical forces, is a mineralized mesenchymal tissue that also possesses an important role in maintaining mineral homeostasis and the energy metabolism of the organism [5]. The formation of bone, which is either developed by the endochondral or by the intramembranous ossification program, is a complex process which depends on physiological interaction with blood vessels that are simultaneously formed [6,7,8,9]. Endochondral ossification designates the process of long bone formation which results from the intermediate generation of a primordial cartilage skeleton composed of mesenchymal stem cells (MSCs) that differentiate into chondrocytes and only at a later stage gradually transform into mature bone recruited by different types of bone-forming cells [6]. Intramembranous ossification, in contrast, denominates the process of establishing flat bones where condensed MSCs directly differentiate into osteoblasts, forming an ossification center. This pathway leads to the formation of craniofacial, calvarial, and clavicle bones [10]. In either type of bone development, angiogenesis—the invasion of small blood vessels derived from preexisting blood vessels—is required [11,12,13]. Bone angiogenesis is induced by growth factors expressed by osteogenic cells such as hypertrophic chondrocytes and osteoblasts at an early stage during osteogenesis [14]. Thereby, the transport of oxygen and nutrients, as well as the further recruitment of osteoprogenitor cells (MSCs) and osteoblasts, are facilitated. In the later phase of bone formation, angiogenesis is essential for trabecular (spongy, cancellous) bone formation and for maturation of the newly-formed bone by close coordination of mineralization and vascularization in either type of bone. In addition, endochondral angiogenesis is particularly important for the replacement of cartilaginous structures at the primary ossification center which generates the bone marrow cavity, and, at a later stage, in establishing the secondary ossification center at the epiphyses (the distal end of long bones) [15,16]. Recent in vitro experiments in transgenic mice demonstrated that this task is accomplished by a specialized type of blood vessel, i.e., the so-called H-type that is present in long bones [17]. This underlines the fact that, comparable to other organs, the acquisition and maintenance of specialized properties by endothelial cells (ECs) is very important for the functional homeostasis in bone.
Bone is a tissue that has to undergo permanent remodeling and requires a counterbalanced process between the anabolic activities of bone formation (osteogenic) cells and the catabolic activities of bone resorption (osteoclast) cells. This activity enforces continuous self-renewal of bone, thereby maintaining an appropriate bone mass and calcium equilibrium. Interference with bone homeostasis could prevent tissue formation, leading to immature or abnormal bone formation [18]. Impaired blood vessel formation, as found in age-related or disease-induced bone loss, therefore, could also result in imbalanced or defective bone formation [19]. In support of this, a reduced density of blood vessels altering the microcirculation was found to be present in osteoporotic bone in mice and humans, that may lead to local abnormal bone metabolism and provoke an increased risk of fracturing [20,21]. Furthermore, the genetic program of bone angiogenesis needs reactivation during callus formation in fracture healing, which represents a complex process that has not yet been fully elucidated [22].
2. Molecular Regulation of Bone Angiogenesis
Bone angiogenesis is mainly governed by a spectrum of transcription factors and growth factors which have been mostly elaborated for endochondral ossification so far [23,24]. It involves interactive signaling between cells of the skeletal system, namely chondrocytes and osteoblasts, and cells derived from the bone vascular system, primarily ECs. Initially, the formation of blood vessels is promoted by osteogenic cells producing pro-angiogenic factors which, in turn, later support the settlement of osteoprogenitor cells [25]. One of the major driving forces behind angiogenic-osteogenic coupling in bone, where oxygen concentrations below 1% are encountered, is the necessity of supplying the tissue with oxygen [15,26,27]. Via tightly hypoxia-regulated induction of the transcriptional activator hypoxia inducible factor (HIF), a cascade of target genes which are involved in a wide variety of biological processes including energy metabolism, erythropoiesis, cell survival, apoptosis and angiogenesis including the major angiogenic regulator vascular endothelial growth factor (VEGF) are expressed in osteogenic cells [28]. The direct mediator of oxygen-dependent modifications of the HIF factors is the von Hippel-Lindau tumor suppressor protein (pVHL), an E3 ubiquitin ligase that targets HIF-1a for proteasomal degradation. During endochondral ossification, VEGF-A is produced by both chondrocytes, particularly in a later stage in which they undergo hypertrophy, and by osteoblasts [29]. As proof of principle, it has been demonstrated that in mice which produce only altered expression levels of a soluble form of VEGF (VEGF120), delayed blood vessel invasion into the primary ossification center and altered osteoblast differentiation in vitro occurs [11]. In addition to HIFs also the fibroblast growth factor (FGF) family of signaling molecules with the members FGF2 and FGF9, and their receptors FGFR1 and 2, were found to be involved in the transcriptional regulation of VEGFA and VEGFR2 expression during formation of blood vessels in bone [30]. Recently, Notch signaling has been implicated as a response to VEGF signaling in ECs in osteogenic-angiogenic coupling [31]. While in other tissues NOTCH expression generally negatively regulates angiogenesis, it seems to have the opposite role in enchondral bone formation by promoting EC proliferation and vessel growth. In a reciprocal fashion, ECs respond by an angiocrine release of NOGGIN, an antagonist of the bone morphogenetic protein (BMP) pathway that stimulates the maturation of hypertrophic chondrocytes expressing VEGF in the growth plate. This could be nicely demonstrated in mice lacking Notch specifically in ECs, which demonstrated reduced bone angiogenesis due to a loss of type-H blood vessels, as well as mutant bone formation and VEGF expression. Other major players are represented by members of matrix metalloproteases (MMPs) due to their proteolytic activity originating from osteoclasts and vascular cells [32]. MMPs also seem to mediate intracellular signaling involving extracellular matrix-integrin interactions necessary during bone angiogenesis and bone remodeling. Besides VEGFA, the placental growth factor (PlGF/PGF), a member of the VEGF family which binds to VEGFR1, has been found to play an exclusively important role in callus remodeling during fracture healing [33,34]. Other known modulating factors of angiogenesis during bone repair include transforming growth factor beta (TGF-β), BMPs, and growth differentiation factor (GDF) [35].
3. The Role of MicroRNAs
MicroRNAs (miRs/miRNAs) are a newly discovered expanding class of endogenous small, non-coding RNAs that positively or negatively regulate gene expression and cellular processes via the RNA interference pathway [5,36,37,38,39,40,41,42,43,44]. By targeting messenger RNA transcripts post-translationally, they provoke either translational repression or degradation, depending on the degree of sequence homology. Upon precursor transcription from intronic or polycistronic genomic loci by RNA polymerase II, biogenesis of the primary miRNA (pri-miRNA) transcript takes place by a two-step processing mechanism involving the RNAses Drosha and DICER (DGCR8 RNase III complex) [45,46,47]. Thereby, single or multiple miRNAs that form hairpin-like structures are exported to the cytoplasm by an exportin 5- and RAN-GTP-dependent process, and cleaved. However, in an alternative non-canonical pathway, miRNAs can be configured by direct transcription or refolded spliced introns as endo-shRNAs (endogenous short hairpin RNAs) or as mirtrons, respectively [48]. Subsequently, the targeting strand of the double-stranded mature miRNAs that are 18 to 22 nucleotides in length is integrated into the RNA-induced silencing complex (RISC) with the help of Argonaute proteins. RISC can finally bind specific target (or so-called “seed”) sequences of mRNAs represented by 2 to 8 nucleotides located mostly in their 3´-untranslated regions (UTRs) [49,50]. Up- or down- regulation of the miRNA itself by stage- and tissue-specific expression patterns during development can lead to modified expression of its target genes. Thus, miRNAs function as decisive regulatory molecules in many different cellular activities such as development, proliferation, and differentiation, metabolism, or apoptotic cell death, or even cell fate determination and maintenance (e.g., pluripotency control of embryonic stem cells) [51,52,53,54,55,56,57,58,59,60]. Moreover, relevant to bone biology, miRNAs have also been identified to acquire endocrine or paracrine functions by their secretion into the blood stream where they subsequently circulate [61]. Consistent with this finding, it was determined that a single miRNA can be involved in coordinating genetic networks by simultaneously regulating the endogenous expressions of multiple target genes. While miRNAs seem to function rather as auxiliary factors during normal physiological processes, their task seems to become more important under stress or disease-related conditions. Accordingly, disturbed miRNA expression is increasingly identified in a number of pathological conditions such as tumorigenesis or viral infection [53,62].
4. MicroRNAs in Bone Angiogenesis: OsteomiRs, AngiomiRs, and HypoxamiRs
It is now well established that miRNAs are physiologically relevant to all steps of bone as well as blood vessel formation during embryonic development and in maintenance during adulthood [63]. OsteomiRs have been identified to regulate chondrocyte, osteoblast, and osteoclast differentiation by positively targeting the principal osteogenic transcription factors and signaling molecules of osteogenesis [5,38,64,65,66,67,68,69]. In addition to regulating MSC commitment—i.e., the differentiation of precursor cells into chondrocytic and osteoblastic lineages—several studies showed that miRNAs also contribute to the maturation and function of these cells, suggesting also important roles in bone regeneration. Nevertheless, the exact mechanisms of skeletal miRNAs governing the complex interactions and signaling pathways of different bone-forming cells are only beginning to be elucidated. Deregulated miRNAs expression or even genetic variation by mutations or single nucleotide polymorphisms in miRNAs or their binding sites have been identified furthermore in bone disorders such as osteoporosis, osteosarcoma, osteopetrosis, osteogenesis imperfecta, osteoarthritis, and furthermore in bone fracture [63,70,71,72,73,74].
Increasing evidence further indicates that miRNAs act as pro- and anti-angiogenic regulators of adaptive blood vessel growth in normal cardiovascular development and in tumor angiogenesis [75,76,77,78,79,80]. The role of these so-called angiomiRs or vascular microRNAs in angiogenic development was initially discovered by the detection of severely disrupted blood vessel formation and delayed angiogenic capabilities of ECs in mid-gestational lethal mouse mutants for the miRNA precursor-processing enzyme Dicer [81]. These mutants die around embryonic day 12 to 14 of development due to vascular defects in the embryo and the yolk sac. Furthermore, a smooth muscle-specific Dicer deletion in the mouse exerted late embryonic lethality associated with extensive internal hemorrhage which could be explained by a significant loss of vascular contractile function, smooth muscle cell (SMC) differentiation, and vascular remodeling [82]. Knockdown experiments of Dicer in zebrafish moreover provoked a phenotype of pericardial edema and inadequate circulation. But also, loss-of-function of the EC-specific miR-126 in homozygous deficient mice caused defects in vascular integrity and angiogenesis [83]. These findings suggested that angiomiRs modulate crucial target genes in cells derived from angioblastic precursor cells and SMC, which are indispensable during embryonic angiogenesis. By investigating the function of Dicer in adult mice and human cells, considerable dysregulated angiogenesis related to growth factor release, ischemia, and wound healing could be revealed, reflecting important postnatal angiogenic functions [80,84,85]. To date, miRNA have been implicated in a long list of cardiovascular diseases comprising myocardial infarction, heart failure, stroke, peripheral and coronary artery disease and several more [86,87]. Nevertheless, the pathological implications of angiomiRs surfaced also with the help of endothelium-specific Dicer-deficient mice, as the ablation led to reduced tumor progression due to diminished angiogenesis, which is a prerequisite for tumor development [88]. For example, two miRNAs induced by VEGF expression (miRs-296, miRs-132) have been identified as candidates supporting the angiogenic switch during tumor formation i.e., the transition from a pre-vascular to a vascularized tumor phenotype [89,90]. In conclusion, the combination of Dicer-deficient angiogenic phenotypes suggests crucial roles for miRNAs in regulating structure and function of embryonic and postnatal blood vessel development.
In the context of angiogenesis, an additional, very important category is a specialized subset of hypoxia-inducible miRNAs, whose increasing number of representatives was also termed hypoxamiRs [91,92,93,94,95,96]. Thus, reduced oxygen supply in ossification centers of bone stimulate the expression of VEGF and other angiogenic factors that lead to the development of blood vessel structures [97]. Additionally, hypoxia-regulated pathways have been attributed to regulatory functions such as smooth muscle cell proliferation and contractility, cardiac remodeling, cardiac metabolism and ischemic cardiovascular diseases [94]. Together with a variety of other target genes which are important for physiological low oxygen adaption, their expression is initiated by upregulation of the transcription factor hypoxia-inducible factor alpha (HIF) [98]. One group of hypoxamiRs are therefore upregulated following HIF expression (HIF-dependent hypoxamiRs), with the master hypoxamiR-210 being the most prominent example [99,100]. Hypoxia-dependently expressed miRNAs that affect HIF expression itself also belong to hypoxamiRs. Thus, for the adaptation to low oxygen conditions and induction of angiogenesis, HIF displays a unique role by controlling further upregulation of hypoxamiR-424 in ECs, which promotes its own protein stabilization [101]. A last group of hypoxamiRs, moreover, influences HIF expression in the absence of hypoxia. As an example, miR-31 decreases HIF-1α expression via the “factor-inhibiting HIF (FIH)” while the miR17-92 cluster suppresses HIF-1α upon c-MYC induction [102,103].
5. Specific MicroRNAs Implicated in Angiogenic-Osteogenic Coupling
Taken together, the functions of osteomiRs, angiomiRs, and hypoxamiRs suggest the possibility that miRNAs will also have crucial roles in bone angiogenesis. Subsequently, miRNAs will be outlined that were found to have a significant function in osteogenesis as well as angiogenesis, and therefore represent miRNAs that have already been identified to have an active role in angiogenic-osteogenic coupling or are presumed to have its potential (Figure 1, Table 1). Collectively, these may also be referred to as “CouplingmiRs/CPLGmiRs”. MiRNAs with a confirmed function in this process could be employed as therapeutic targets in bone regeneration. Consequently, they could improve the coordination and enhancement of the endogenous osteogenesis and angiogenesis process. Elucidation of the molecular mechanisms governing osteoblast differentiation and angiogenesis are furthermore of great importance for improving the treatment of bone-related diseases.
5.1. MiR-9
miR-9 is a highly conserved microRNA, but exhibits a divergent expression pattern, and seems to modulate different targets in a cellular context- and developmental stage-specific manner [104]. The studies of Han et al. have demonstrated a regulatory role for miR-9 in the development and differentiation of human bone marrow derived MSCs (hBM-MSCs) and neural progenitor cells on proliferation, migration and differentiation [105]. In this regard, cell-autonomous effects have been described in vertebrates by effecting Notch, Wnt, and BAF53a expression. Furthermore, miR-9 has also been related to tissue repair processes involving human MSCs (hMSCs). Qu et al. provided evidence that increased miR-9 expression levels closely correlate with enhanced differentiation of MC3T3-E1 osteoblasts [106]. The effects of miR-9 on angiogenesis were studied by the same authors by using human umbilical vein endothelial cells (HUVECs). Here, miR-9 mimics transfection effectively increased VEGF, VE-cadherin, and FGF concentrations in the culture medium, leading to increased EC migration and capillary tube formation in vitro. The underlying molecular mechanism for both the regulation of osteoblast differentiation and angiogenesis was found in the activation of AMP-activated (AMPK) signaling. Conclusively, the results of Qu et al. imply a potential important role of miR-9 in regulating the process of bone injury repair, and therefore, a potential therapeutic target for the treatment of bone injury-related diseases.
A different study that analyzed the function of miR-9 in C2C12 mesenchymal cells further supported the role of miR-9 in osteoblast differentiation. By significantly decreasing the expression of DKK1 protein, but not of its mRNA, miR-9 stimulated alkaline phosphatase (ALP) activity and osteoblast mineralization, as well as the expression of several osteoblast marker genes, such as COL I (collagen I), OCN (osteocalcin), and BSP (bone sialoprotein) [107]. MiR-9 furthermore was detected as a tumor-secreted pro-angiogenic miRNA that promoted EC migration and tumor angiogenesis in vitro. Mechanistically, this was explained by interference with the expression levels of SOCS5, and thereby, the activation of the JAK-STAT pathway [108].
Moreover, altered expression of miR-9 was found during screening osteoarthritis cartilage involved in the control of tumor necrosis factor α (TNFα) expression. Here, miR-9 has been implicated as a key regulator in the process of endochondral ossification, since its expression varied significantly between the early and late stage of chondrocyte development [73,109]. Functional experiments in a mouse tibial plateau fracture model implicated that miR-9 and miR-181a significantly downregulated Bim concentration. Thereby, osteoclast survival was stimulated and the migration ability of osteoclasts was effected [110].
5.2. MiR-10a
The function of miR-10a during osteoblast differentiation and angiogenesis in vitro was analyzed in MC3T3-E1 and MUVEC (mouse umbilical vein endothelial cells) by the research group of Li et al., respectively [111]. Upon BMP2-induced osteoblast differentiation of MC3T3-E1 cells, miR-10a was downregulated. In contrast, when miR-10a was overexpressed, a suppressive effect on β-catenin and LEF1 expression could be demonstrated. Overexpression inhibited osteogenic differentiation, as demonstrated by reduced expression of the osteoblast-differentiation markers ALP, runt-related transcription factor 2 (RUNX2), Osterix (OSX), and distal-less homeobox 5 gene (DLX5). Moreover, it led to a decrease in MUVEC proliferation, migration and tube formation in combination with reduced concentrations of the angiogenesis-related genes VEGF, VE-cadherin, cyclin D1, and MMP2. As the canonical WNT/β-catenin signaling pathway was found to play an important role in osteogenic cell proliferation, differentiation, and bone regeneration, miR-10a offers a potential therapeutic target for the treatment of bone regeneration and bone-related diseases [112].
A further report referred to the role of miR-10a in regulating endothelial progenitor cell (EPC) senescence in the mouse [113]. Zhu et al. provided evidence that upon upregulation of miR-10A and miR-21, Hmga2 (High-mobility group AT-hook 2) expression is gradually decreased during aging in bone-marrow cells that were enriched for EPCs. Suppression of miR-10A* and miR-21 in aged EPCs, on the other hand, increased Hmga2 expression and improved EPC angiogenesis in vitro and in vivo. This could be demonstrated by rejuvenated EPCs, which resulted in decreased senescence-associated β–galactosidase expression, increased self-renewal potential and decreased p16Ink4a/p19Arf expression. In conclusion, the study demonstrates that the miR-10A*/miR-21–Hmga2–P16Ink4A/P19Arf axis controls EPC senescence and angiogenesis and may represent a potential therapeutic intervention target for improving EPC-mediated angiogenesis and vascular repair.
5.3. MiR-10a/10b
Hassel et al. investigated the function of miR-10 in zebrafish blood vessel formation. During embryogenesis, the knockdown of miR-10a/10b impaired blood vessel outgrowth due to an altered tip cell differentiation behavior, and led to defects of intersegmental vessel growth by modulating fms-related tyrosine kinase 1 (flt1) levels post-transcriptionally [114]. However, as the knockdown of flt1 did not fully rescue the angiogenic phenotypes in miR-10 mutant zebrafish, as well as in miR-10-deficient HUVECs, the authors concluded that in ECs, flt1 could not represent a direct exclusive target of miR-10. They provided evidence that miR-10a/10b regulated angiogenesis in a Notch-dependent manner by directly targeting mib1 (mindbomb E3 ubiquitin protein ligase 1) in zebrafish ECs. Inhibition of mib1 and Notch signaling partially rescued the angiogenic defects in miR-10 morphants, suggesting that the observed angiogenic defects in miR-10a/10b morphants are caused by up-regulation of Notch signaling [115].
5.4. MiR-20a
As a member of the extensively studied miR-17-92 cluster with prominent roles in tissue and organ development, the role of miR-20a was elucidated during osteogenesis by Zhang et al. [116]. They disclosed that, together with several osteoblast markers (BMP2, BMP4, RUNX2, OSX, OCN and OPN), miR-20a was upregulated during osteogenic differentiation of hBM-MSCs derived from bone marrow from differently aged persons. Adipocyte markers, however, such as PPARγ and the osteoblast antagonists, BAMBI and CRIM1, were down-modulated. By introducing miR-20a mimics and lentiviral-miR20a-expression vectors into hBM-MSCs, they verified that miR-20a enhances osteogenic differentiation. Simultaneous direct interaction with all the aforementioned positive and negative effectors of BMP/RUNX2 signaling could be confirmed.
In a report by Doebele et al., miR-20a was investigated with respect to its cell-intrinsic angiogenic activity in EC, as different members of the miR-17-92 cluster that are highly expressed in tumor cells were found to be expressed at increased levels during ischemic conditions [117]. They determined that in vitro overexpression of miR-20a (together with miR-17, -18a, -19a) rigorously inhibited EC sprout formation, whereas their inhibition using antagomiR treatment led to an increase of spheroid sprouting (irrespective of miR-19a). Interestingly, in vivo matrigel plug assays employing antagomiRs for miRs-17-20a as inhibitors demonstrated enhanced angiogenic sprouting, but in contrast to in vitro results, they were unaltered in tumor angiogenesis, indicating context-sensitive regulation. In particular, the pro-angiogenic target gene Janus kinase 1, but also the cell cycle inhibitor p21 and the S1P receptor EDG, were shown to be downregulated by miR-17/20a.
Deng et al. provide evidence that miR-20a and miR-31 serve as stimulators of angiogenesis [118]. As an underlying mechanism, they found that the expression of both miRNA molecules is upregulated via AKT and ERK signals that are themselves activated by the angiogenic factor VEGF. As target genes, miR-20a and miR-31 were found to directly associate with the 3´-UTR of the tumor necrosis factor superfamily-15 (TNFSF15) gene, thereby clarifying its by then unknown mechanistic role in vascular homeostasis. TNFSF15 is expressed in ECs of mature vasculature, and is a known inhibitor of angiogenesis. Interestingly, VEGF-stimulated downregulation of TNFSF15 could be attenuated by treatment of HUVECs with AKT inhibitor LY294002, leading to reduced miR-20a and miR-31 levels, while ERK inhibitor U0126 prevented VEGF-induced expression of miR-20a only. In contrast, inactivation of either ERK or AKT signals restored TNFSF15 gene expression and elevated miR-20a or miR-31 levels which led to enhanced capillary-like tube formation in an in vitro angiogenesis assay.
5.5. MiR-26a/b
Luzi et al. investigated and confirmed an important function of miR-26a during human adipose-derived MSCs (hADSCs) differentiation towards the osteogenic lineage induced by treatment with dexamethasone, ascorbic acid, and beta-glycerol phosphate [119]. Upon inhibition of miR-26a by antisense RNA, upregulation of the transcription factor SMAD1—which was predicted in silico—and its regulated osteogenic differentiation marker genes could be observed in treated osteoblasts. In a follow-up study of Luzi et al., these results were extended to include the interaction between menin and miR-26a as regulators of osteogenic differentiation in hADSCs [120]. Menin is a presumable transcriptional regulator that modulates mesenchymal cell commitment to the myogenic or osteogenic lineages. It is encoded by the MEN1 oncosuppressor gene which causes the multiple endocrine neoplasia type-1 syndrome. The results demonstrated orchestrated down-regulation of MEN1 mRNA and miR-26a, with a consequent up-regulation of SMAD1 protein in hADSCs.
Su et al., however, reported an opposing role of miR-26 in hBM-MSC during osteogenic differentiation, suggesting distinct post-transcriptional regulation of tissue-specific hMSC differentiation [121]. Using bioinformatics and functional assays, they confirmed that miR-26a directly regulates SMAD1, but added GSK3β as a target to regulate BMP and WNT signaling pathways. The distinct activation pattern and comparative analysis revealed that miR-26a significantly inhibited SMAD1 to suppress BMP signaling for interfering with the osteogenic differentiation of hADSCs, whereas it targeted on GSK3β to activate WNT signaling for promoting osteogenic differentiation of hBM-SCs. Overall, they concluded that the BMP pathway was more essential for promoting osteogenic differentiation of hADSCs, whereas WNT signaling was enhanced more potently and played a more important role than BMP signaling in osteogenic differentiation of hBM-SCs. In conclusion, although miR-26a enhances osteogenic differentiation in both cell types, different signaling pathways were employed in hBM-MSCs and hADSCs.
In to addition unrestricted somatic stem cells (USSC), the studies of Trompeter et al. investigated a rare population in human cord blood with respect to osteogenic differentiation. Gene expression profiling of two different USSC cell lines (SA5/73 and SA8/25) identified, among other candidates, miRs-26a/b and miR-29b to be consistently upregulated during osteogenic differentiation [122]. As osteo-inhibitory targets of these miRNAs, CDK6 and HDAC4 were evaluated that were downregulated during osteogenic differentiation of USSC, whereas SMAD1 was found as an osteo-promoting target. During osteogenic differentiation of USSC or following ectopic expression of miR-26a/b, SMAD1 exhibited an unchanged expression level, however.
Entangling of miR-26a in pathological and physiological angiogenesis was investigated by Icli et al. in ECs [123]. They studied the effects of modifying the expression of BMP/SMAD1 signaling. Upregulation of miR-26a led to EC cycle arrest, inhibited EC migration, sprouting angiogenesis, and network tube formation in matrigel. Upon inhibiting miR-26a expression, a contrasting phenotype could be detected. At the molecular level, Icli et al. demonstrated direct binding of miR-26 to the 3′-UTR of SMAD1 thereby reducing its mRNA levels, which subsequently suppressed ID1 expression and increased p21WAF/CIP and p27 protein expression.
5.6. MiR-29b
MiRNA profiling of MC3T3 preosteoblastic cells derived from fetal mouse calvaria and differentiated to osteoblasts led to the identification of miR-29b, among other members of the miR-29, miR-let-7, and miR-26 families by Li et al. [124]. Versatile effects of miR-29b were found to promote osteoblastogenesis at multiple stages as a key regulator. One mechanism pursues the silencing of negative regulators of osteogenic differentiation, such as TGF-β3, HDAC4, ACTVR2A, CTNNBIP1, and DUSP2 that involve particularly the osteogenic function of RUNX2, as well as the SMAD, ERK, p38 MAPK, and WNT signaling pathways. A second path seeks the suppression of extracellular matrix protein synthesis relevant to bone development (such as COL1A1, COL5A3, and COL4A2) to preserve the differentiated phenotype during mineralization of mature osteoblasts. This alternative mechanism seems to enhance mineral deposition and to prevent fibrosis.
Rossi et al. also disclosed a link of miR-29b to osteoclastogenesis and proposed it for the treatment of multiple myeloma-related bone disease as its expression declined increasingly during human osteoclast differentiation and affected proper bone resorption [125]. Several findings indicated that miR-29b is a negative regulator of human OCL differentiation and activity. Thus, lentiviral transduction of miR-29b into OCLs was associated with diminished tartrate acid phosphatase expression, lacunae generation, and collagen degradation. Attenuated resorptive osteoclast capabilities, due to miR-29b inhibition of proteolytic enzymes, were documented by reduced cathepsin K, MMP-9, and MMP-2 expression. Overall, downstream phenotypic effects along the M-CSF and RANK-L axes that led to impaired action of the master transcription factor NAFTc-1 were explained by miR-29b targeting of c-FOS.
Zhang et al. found inhibitory activity of miR-29b on VEGF secretion via the anti-angiogenic cytokine TNFSF15 (VEGI; TL1A) in the mouse EC line bEnd.3, which defines a new angiogenesis-related signaling pathway [126]. In contrast, down-modulation of TNFSF15 activity by a specific siRNA against its receptor DR3/TNFRSF25, or a neutralizing antibody against TNFSF15, reinstated VEGF generation but suppressed miR-29b expression. TNFSF15-enhanced activation of the JNK-GATA3 signaling pathway was furthermore able to stimulate miR-29b expression but silenced VEGF production, as demonstrated by a specific JNK inhibitor or siRNA.
Li et al. described an anti-angiogenic and anti-tumorigenic role for miR-29b by the regulation of AKT3 expression [127]. AKT is known to induce tumor vascularization via VEGF, and cancer cell activity via c-MYC arrest, in breast tumor. In vitro and in vivo ectopic expression of miR-29b therefore blocked angiogenesis, as well as tumor cell formation, evidencing it as a potential useful anti-cancer therapeutic agent.
5.7. MiR-31
Granchi et al. detected miR-31 by profiling miRNA expression during osteogenic differentiation and mineralization of hBM-MSCs that were derived from three individual donors [128]. As an identified direct target gene, miR-31 differentially modulated the expression of the bone-specific transcription factor OSX during osteogenic differentiation [129]. This could be demonstrated by an inverse miRNA-target expression ratio in osteosarcoma cell lines and an increase in OSX expression upon specific miR-31 inhibition.
Deng et al. provided evidence that the expression of miR-31 increasingly declined during the osteogenic differentiation of hBM-MSC cells [130]. This regulation coincided with increased ALP activity, mineralization of hBM-MSC cultures and expression of the osteogenic transcription factors OPN, BSP, OSX, and OCN, with the exception of RUNX2. Mechanistically, they uncovered a RUNX2, SATB2, and miR-31 regulatory feedback loop that determined hBM-MSC differentiation using inhibitors and mimics of miR-31. RUNX2 directly regulates miR-31 expression levels which itself controls the translation of SATB2 protein.
In a study of Suarez et al. that investigated TNF-mediated induction of endothelial adhesion molecules, the expression of miR-155, -31, -17, and -191 were found to be increased without a change of miR-20a, -222, and -126 levels in HUVECs [84]. By miRNA target prediction algorithms, the 3´-UTR of E-selectin was identified as target molecule, which was verified experimentally by reporter assays using miR-31 mimics. miR-31-mediated regulation of E-selectin expression not only regulates binding of neutrophil granulocytes to HUVECs, but is also involved in inhibition of angiostatin-induced angiogenesis by affecting cell migration [131].
An angiogenesis-related function of miR-31 reported by Deng et al. has already been described above in the subsection of miR-20a [118].
5.8. MiR-34a
Opposing roles for miR-34a in differentiation of hMSC towards osteoblast have been reported. Chen et al. identified miR-34a in a microarray screening, and stated that it inhibits osteoblast differentiation in hMSC and in vivo bone formation in a preclinical model of heterotopic bone formation in mice [132]. Thus, when miR-34a was overexpressed in hMSC, it inhibited early and late OB commitment, as well as differentiation and hMSC proliferation, while anti-miR oligonucleotide treatment reversed these effects. In addition to several cell cycle regulator and cell proliferation proteins (including CDK4, CDK6, and Cyclin D1), JAGGED1, a NOTCH1 receptor ligand, was elicited as a target gene of miR-34a that was regulated at both the transcriptional and translational levels, as determined by RNA interference. JAGGED1 has previously been implicated in human bone biology, as its deficiency causes skeletal abnormalities in the Alagille syndrome.
Kang et al., however, claimed that miR-34a-5p-induced activation of the Notch signaling pathway is a positive regulator of glucocorticoid-mediated osteogenic differentiation of hMSCs [133]. They demonstrated dexamethasone-inhibited osteoblastic differentiation of murine BM-MSC via miR-34a-5p-mediated gene silencing of coincidently identical target genes, as published by Chen et al. Differences in both reports were discussed to be due to different roles of the Notch signaling pathway in osteogenic differentiation of hMSCs that were derived from different species.
A more recent publication by Fan et al. found miR-34a upregulation during osteogenic differentiation of hADSCs [134]. Elevated levels of miR-34a in hADSCs promoted mineralization, ALP activity, and the expression of the key regulatory osteogenic transcription factor RUNX2 by targeting the retinoblastoma binding protein 2 (RBP2); furthermore, heterotopic bone formation was enhanced in vivo. Expression of NOTCH1 and Cyclin D1 genes, that were also involved in this coregulatory network, were found to be downregulated on the other hand, which facilitates cell cycle exit [135,136]; this is the consequence of suppressed proliferation but enforced terminal maturation of osteoblasts by RUNX2.
When investigating the role of miR-34a in glucocorticoid-induced osteonecrosis of the femoral head (GIOFH), Zha et al. verified the findings of Kang et al., but extended this knowledge [137]. By investigating dexamethasone-treated rat subjected to miR-34a-overexpressing lentiviruses, decreased blood vessel development was observed, indicating that VEGF presents a regulatory target of miR-34a. In vitro, miR-34a overexpression enhanced the inhibitory effects of dexamethasone on the viability and activity of ECs and downregulated VEGF protein expression levels.
A study by Zhao et al. investigated the angiogenic role of miR-34a in EPCs derived from adult male Spraque-Dawley rats [138]. The rationale behind this project was the previous implication of miR-34a in targeting silent information regulator 1 (SIRT1), provoking cell cycle arrest or apoptosis. The results confirmed the inhibitory effects of raised miR-34a expression levels on SIRT1 and, thus, on EPC-mediated angiogenesis by inducing senescence. Mechanistic causes could be found in increased acetylated levels of the FOXO1 transcription factor, regulated by SIRT1, which could be demonstrated by knockdown of SIRT1. The angiogenesis-promoting role of miR-34a and Sirt1 in this context seems to lie in its previously discovered function in vascular endothelial homeostasis by preventing stress-induced senescence in health and disease [139].
A negative regulatory role for miR-34a could also be documented by Kumar et al. in tumor angiogenesis in head and neck squamous cell carcinoma (HNSCC) [140]. MiR-34a expression was markedly decreased in HNSCC tumors and cell lines, and ectopic expression of the miR-34a therefore affected cell proliferation and migration in HNSCC cell lines and in a SCID mouse xenograft model. These effects seemed to be mediated by the regulation of survivin expression via the transcription factor E2F3a that is critical for cell cycle progression. Furthermore, tumor angiogenesis was found to be dysregulated by interference of miR-34a with VEGF secretion in tumor cells, as well as EC proliferation, migration, and tube formation, by downregulating a number of key proteins including E2F3, SIRT1, survivin, and CDK4.
5.9. MiR-92a
The autosomal dominant Feingold syndrome that is characterized by microcephaly, short stature, and digital anomalies was identified in individuals carrying hemizygous deletions of the miR-17-92 cluster [172]. To dissect this complex phenotype that could be partially mirrored in mice deficient of the miR-17-92 cluster, and due to postnatal lethality, a single miR-92a targeted mouse line was established by Penzkofer et al. [173]. Surviving mice exhibited reduced body weight and skeletal defects represented by reductions in body length, skull, and tibia length, as well as metacarpal bone size; these defects were presumably also the result of osteoblast proliferation and differentiation phenotypes, as found in hemizygous miR-17-92 mouse mutants [174]. In contrast, however, single miR-92a deletion does not cause low bone mineral density attributed to reduced type-I collagen mRNA, ALP activity, and mineralization ability. Phenotypic differences could be explained by the design of the genomic deletion of miR-92a that partially attenuated expression levels of miR-18a in skeletal tissue. The direct molecular targets of miR-92a responsible for regulation of osteogenesis have not yet been elicited.
A recent study of Mao et al., however, identified aggrecanase-1 and aggrecanase-2 (ADAMTS4/5) as direct miR-92a targets in chondrogenic hMSCs and human chondrocytes by reporter assays [175]. These two members of the ADAMTS family represent MMPs that are important for normal chondrocyte differentiation, but which also promote the progression of osteoarthritis by cartilage degeneration. In comparison to normal cartilage, real-time-PCR analyses also revealed higher miR-92a-3p expression levels in chondrogenic hMSCs, whereas markedly reduced miRNA expression could be detected in ostearthritis cartilage. Moreover, miR-92a-3p-modified expression of ADAMTS-4/5 could be downregulated by IL-1β transfected primary human chondrocytes. The study was preceded by previous findings about miR-92a-3p upregulation in hADSC-derived chondrocytes and chondrogenic hMSCs, as well as osteoarthritis cartilage, where histone deacetylase 2 (HDAC2) was identified as a target gene of miR-92a-3p [176,177].
MiR-92a was first reported as a part of the miR-17-92 cluster by Bonauer et al., and was found to be abundant in human ECs [178]. MiR-92 was established as a suppressor of angiogenesis which targets the expression of several proangiogenic proteins, including the integrin subunit alpha 5 (ITGA5) that is well known for severe vascular defects in gene targeted mice. Systemic administration of miR-92a antagomiR therefore improved the growth of blood vessels and functional recovery of damaged tissue in mouse models of limb ischemia and myocardial infarction, possibly by indirectly inhibiting apoptosis [58]. Thus, miR-92a may serve as a valuable therapeutic target in the setting of ischemic disease.
Daniel et al. further elaborated the function of miR-92a upon re-endothelialization and neointimal formation after wire-induced injury of the femoral artery in mice [144]. By using specific LNA (locked nucleic acids)-based antimiRs as well as miR-92a-deficient ECs in the mouse, they found enhanced re-endothelialization and inhibited neointimal formation induced by de-repression of the miR-92a targets integrin a5 and sirt1 [179]. Thus, an important role can be attributed to miR-92a in blood vessel regeneration in ischemic tissues and after vascular injury.
In a subsequent study, Zhang et al. investigated the ability of miR-92a to influence apoptosis and angiogenesis in ECs in the presence of oxidative stress [180]. It was previously determined that senescent ECs that undergo apoptosis, and which are frequent in ageing and atherosclerosis, have diminished miR-92a levels [181]. The results provided evidence that pre-miR-92a treatment of HUVECs prevented oxidative stress-induced apoptosis in EC, whereas capillary tube formation i.e., angiogenesis, was maintained. Mechanistically, it was determined that pre-miR-92 directly repressed PTEN, and thereby, activated the AKT signaling pathway that regulates EC apoptosis and angiogenesis.
In a study of Kalinina et al., paracrine effects of miR-92a on hMSCs were studied [143]. Conditioned medium of hMSCs transfected with pre-miR-92a prevented tube formation by HUVECs, which could be attributed to significantly lower secretion of hepatocyte growth factor (HGF) and angiopoetin-1 independent of VEGF secretion. HGF is a factor required for stimulation of proliferation of ECs, whereas angiopoietin-1 regulates vessel stabilization and maturation. As neither gene was predicted as direct miR-92a targets, these still have to be identified. Restoration of tube formation was achieved by replenishment of HGF, but not with anti-miR-92a treatment of hMSCs. This led to the conclusion that miR-92a suppresses hMSC-induced angiogenesis by downregulating the secretion of HGF and, therefore, is involved in the control of anti-angiogenic activities in hMSCs.
5.10. MiR-125b
MiR-125b is an early discovered miRNA that plays a key role in cellular functions. Experiments by Mizuno et al. using BMP-4-induced or exogenous miRNA-transfected murine MSC ST2 cells found miR-125b to inhibit osteoblastic (and adipogenic) differentiation through modulating cell proliferation [146]. Further experiments with transfection of siRNA identified ErbB2 as a target gene. miR125b-gene targeted mice, generated by Lee et al., identified further supported the important function of miR-125b in regulating stem cell directional differentiation, but did not reveal its molecular role [182].
Decreasing levels of miR-125b were also identified by Huang et al. during the differentiation of C3H10T1/2 cells, and reporter gene assays led to the identification of a putative target binding site in the 3´-UTR of the Cbfβ gene, a master regulatory gene of osteogenesis [148]. Therefore, silencing of miR-125b increased the mRNA levels of Cbfβ and of osteoblastic marker genes ALP, OCN, and OPN. Conclusively, RUNX2 is considered as an indirect target of miR-125b as well.
Transient expression of miR-125b expression in hypoxic VEGF-or or bFGF stimulated ECs was moreover shown by Muramatsu et al. to directly bind and block the translation of vascular endothelial (VE)-cadherin mRNA and therefore inhibit in vitro tube formation by ECs [150]. Thus, miR-125b may be tested in tumor therapy for inducing disruption of blood vessel formation.
5.11. MiR-135b
During the osteogenic differentiation of several USSC, miR-135b was identified as the most consistently down-regulated candidate by Schaap-Oziemlak et al. [151]. Retroviral overexpression resulting in decreased mineralization confirmed a function of miR-135b in osteogenesis of USSC. Furthermore, quantitative RT-PCR analysis of USSC that overexpressed miR-135b showed decreased expression of the bone mineralization markers IBSP and OSX.
In a profiling study by Xu et al. that investigated the exosomal content of hBM-MSCs during osteogenic differentiation, miR-135b was found to be significantly increased [183]. Bioinformatic analysis led to the conclusion that several important pathways related to osteoblastic differentiation were engaged, and that exosomal miRNA is a regulator thereof.
By establishing hypoxia-resistant multiple myeloma (MM) cells through exposure to chronic hypoxia, Umezu et al. presumably identified a new mechanism of hypoxia-induced angiogenesis targeting the FIH-1/HIF-1 signaling pathway via exosome-contained miRNAs [152,184]. In contrast to the transiently hypoxia-upregulated miR-210 transcriptional levels which decline gradually under normoxic conditions, miR-135b levels were maintained high, even under normoxic conditions. By delivering exosomes to HUVECs, miR-135b is enabled to target the factor-inhibiting hypoxia-inducible factor-1 gene (FIH-1) that encodes an asparaginyl hydroxylase enzyme which inhibits HIF-1a. The positive correlation between miR-135b, HIF-1a, and microvessel density was initially identified by Zhang et al. in a HNSCC model [185]. As a result, endothelial tube formation could be promoted under hypoxic as well as normoxic environments.
5.12. MiR-181a
Among other functions, the miR-181 family has been implicated in genetic regulation of early hematopoiesis and lymphangiogenesis [60,186,187]. Microarray analysis of human HCS-2/8 cells led to the characterization of miR-181a. Sumiyoshi et al. accredited miR-181a with an important function in the maintenance of cartilaginous metabolism by a negative feedback system employing repression of the CCN family member 1 (CCN1) and aggrecan (ACAN) genes, which are both known to be involved in chondrocyte differentiation [154].
In another study, the examination of synovial fluid cells in a mouse model of tibial plateau fractures led to the detection of two downregulated miRNAs, miR-9 and miR-181a [110]. Cbl, an important E3 ubiquitin ligase for bone resorption that was tested as a putative target gene of miR-9 and miR-181a, elicited increased amounts of ubiquitinated Bim, a pro-apoptotic gene in mouse primary osteoclast cells. Therefore, Wang et al. concluded that upregulated Cbl might regulate the survival rate of primary mouse osteoclast cells, as previously Cbl-dependent apoptosis via ubiquitinated Bim was reported [188].
Sun et al. implicated miR-181a as a potential oncomiR (cancer-associated miRNA) in the angiogenesis and metastasis of chondrosarcoma in a xenograft mouse model [156]. They found that miR-181 increases VEGF and MMP1 expression, as well as CXCR4 signaling, via negatively regulating RGS16 (regulator of G-protein signaling 16) under hypoxic cell culture conditions. RGS16 is an inhibitor of CXCR4 which regulates, upon amplified signaling, angiogenesis, invasion, and metastasis in chondrosarcoma. The therapeutic usefulness of this mechanism was proven by miR-181a antagomiR treatment, which decreased proangiogenic gene expression as well as tumor growth and metastasis in a xenograft mouse model.
5.13. MiR-195
Together with miR-497, miR-195 was downregulated in a microarray screen of primary hMSCs performed by Almeida et al. [158]. hMSCs underwent induced osteogenic differentiation with the aim of identifying candidates that are capable of contributing to bone fracture repair. Osteogenic markers were therefore found to be diminished upon overexpression or increased upon inhibition of miR-195 in hMSCs. Using the chicken CAM assay, studying the paracrine effects of hMSCs, the authors furthermore demonstrated decreased blood vessel formation in vivo. VEGF was identified as the target gene that mediates this phenotype, at least in part. MiR-195 interacted with the VEGF 3´-UTR in bone cancer cells and also regulated mRNA and protein expression levels.
Reduced expression of miR-195 furthermore resulted in increased angiogenesis and metastasis in HCC tissues, whereas either loss-of-function or gain-of-function abrogated the ability of HCC cells to migrate and induce capillary tube formation of ECs, as reported by Wang et al. [159]. In xenograft tumors, upregulated miR-195 expression provoked reduced microvessel densities and the formation of metastases. Detailed molecular investigations disclosed VEGF and the prometastatic factors VAV2 and CDC42 as direct targets of miR-195, which could be proven by mirroring the phenotype either by knockdown or overexpression of these target genes. The group also demonstrated that higher VEGF levels due to miR-195 down-regulation promoted EC-mediated tumor angiogenesis by the involvement of VEGF receptor 2 signaling.
5.14. MiR-200b
miR-200b was identified by microarray screening as a miRNA that exhibits downregulated expression upon exposure of osteoblasts to collagen and to silicate-based periodontal grafting material (PerioGlas, P-15) that is used to promote bone formation [189]. Rønbjerg et al. demonstrated miR-200b as a potent regulator of the target gene ZFHX1B via direct interaction [190]. ZFHX1B is a transcriptional repressor involved in the regulation of the TGFβ signaling pathway and epithelial-mesenchymal transitions by E-cadherin, a mediator of cell-cell adhesion, in mesenchymal cells.
In addition to the known communication of interacting cells by the exosomal release of miRNAs (e.g., mir-135b [153,191]), Fan et al. discovered angiogenic-osteogenic cell coupling and reciprocal interactions via gap junctions [161,192]. They provided evidence that in a direct co-culture, miR-200b was transferred in a TGF-β-stimulated process from rat BM-MSCs to HUVECs through gap junctions formed of connexin 43. By this transfer and decrease of miR-200b, VEGF-A-induced expression enhanced osteogenic differentiation in BM-MSCs. In HUVECs, increasing miR-200b levels down-modulated ZEB2, ETS1, KDR and GATA2 transcription factors, which led to a decline of the angiogenic potential of HUVECs, in contrast. In vitro angiogenesis in this co-culture could therefore be partially rescued by employing the TGF-β inhibitor SB431542 or TGF-β-neutralizing antibody. These findings could provide a new strategy for cell-based bone regeneration.
In lung epithelial carcinoma cells (A549 cells), direct negative regulation of VEGF, VEGFR1 (Flt-1) and VEGFR2 (KDR) by binding of miR-200b to the corresponding 3´-UTR could be demonstrated by Liu et al. [193]. This interaction could be furthermore confirmed in an in vitro angiogenesis assay by transfection of HUVECs, which resulted in reduced capillary tube formation and significantly reduced VEGF-induced phosphorylation of ERK1/2. In addition, miR-200b targets the Ets-1 transcription factor, that might concomitantly downregulate VEGFR2, as found in human mammalian epithelial cells by Chan et al. [194]. Thus, miR-200b may be used as a therapeutic angiogenesis inhibitor.
5.15. MiR-210
Upregulated expression of miR-210 was detected in BMP-4-induced osteoblastic differentiation of murine stromal BM-MSC ST2 cells by Mizuno et al. [163]. Transfection experiments of sense and antisense miR-210 therefore promoted or repressed osteogenesis, respectively. As a target gene mediating the positive regulation, the activin A receptor type-1B (AcvR1b) gene was elicited that effects the TGF-β/activin signaling pathway negatively.
Studies of Fasanaro et al. proved that the expression of miR-210 progressively increases upon exposure to hypoxia. The overexpression of miR-210 in HUVECs using miRNA mimics stimulated the formation of capillary-like structures and enhanced VEGF-induced cell chemotaxis [165,195,196]. Thus, miR-210 up-regulation is a crucial hypoxia response element of ECs, affecting cell survival, migration, and differentiation and might participate in the modulation of the angiogenic response to ischemia [99,197,198]. As a consistently reported target gene mediating these effects, Ephrin-A3 receptor tyrosine kinase ligand (EFNA3) was validated. EFNA3 was previously shown to play a role in the regulation of angiogenesis and VEGF signaling during vascular development and remodeling via EFNA1/EphA2 interaction [199,200]. Overexpression of an Ephrin-A3 allele that is not targeted by miR-210 therefore prevented miR-210-mediated stimulation of tube formation and EC migration.
In its function of promoting osteoblast differentiation by increased VEGF, ALP and OSX expression in rat MSCs and suppression of adipocyte differentiation, due to decreased PPAR-γ in vitro, miR-210 was also implicated in the regulation of postmenopausal osteoporosis [164]. Although the exact mechanism still needs to be elaborated, Liu et al. also found that HIF-1α and VEGF expression was increased in 17β-estradiol (E2)-treated osteoblasts.
5.16. MiR-222
Yan et al. identified miR-222-3p as a negative regulator involved in osteogenic differentiation of hBM-MSCs [166]. Enhancement of miR-222-3p function in hBM-MSCs was blocking protein levels of SMAD5 and RUNX2. miR-222-3p-specific inhibition via lentivirus infection, in contrast, led to their enhanced expression and increased phosphorylation of Smad proteins (1, 5, 8) that are responsible for expression of osteogenic genes. Thus, in addition to the Smad5-RUNX2 signaling pathway elevated levels of the osteoblast markers OSX, ALP, and OC, as well as increased matrix mineralization, could be detected.
The role of miR-222-3p in c-Src-mediated regulation of osteoclastogenesis was proven by Takigawa et al. [167]. Depending on the use of miR-222-3p inhibitor or mimics in RAW264.7 pre-osteoclastic cells, either upregulation, or downregulation of the mRNA expression levels of osteoclast marker genes NFATc1 or TRAP, were observed, respectively. Inhibition of of c-Src activity and activation of osteoclastogenesis via miR-222-3p was implemented by increased amounts of RANKL-induced expression of TRAP and cathepsin K protein levels. Thereby, the number of multi-nucleated osteoclasts and their pit formation was reduced.
Poliseno et al. investigated the role of miR-221 and miR-222 during in vitro angiogenesis [168]. Both miRNAs were identified among 15 upregulated miRNAs that allegedly target receptors of angiogenic factors in a large-scale screen of HUVECs. Mechanistically, they were found to post-translationally modify the angiogenic effects of stem cell factor (SCF) by targeting its receptor, c-KIT. Together with other angiogenic growth factors, such as VEGF, bFGF, or HGF, SCF is able to stimulate proliferation and migration of ECs and induce capillary-like tube formation when performing in vitro angiogenesis assays [201]. Consistently, tube formation, wound healing, or cell migration was suppressed in miR-221/miR-222– transfected and SCF-treated HUVECs.
5.17. MiR-424
miR-424 was identified by differential screening in a miRNA microarray of isolated primary hMSCs from four individuals that were, or were not, osteo-differentiated using osteogenic differentiation medium. In combination with miR-31, miR-106a, and miR-148a, Gao et al. found that miR-424 was suppressed, and predicted it to target RUNX2, CBFB (core-binding factor, beta subunit), and BMPs [169]. In a similar experimental setting of Vimalraj et al., miR-424, together with miR-106a, miR-148a, let-7i and miR-99a, were detected as hMSC-specific miRNAs that were found to be expressed only in undifferentiated hMSCs [170]. Here, bioinformatics analysis mostly predicted the MAPK, WNT, and insulin signaling pathways as targets.
A recent study of Li et al. elucidated further specific functional mechanisms of miR-424 during bone formation and oxidative stress [171]. It was reported that miR-424 was downregulated by the transcription factor FOXO1 using consensus binding sites in the promoter. Subsequently, via miR-424 mediated upregulation of FGF2 under oxidative stress, proliferation and osteogenic differentiation of hMSCs was accomplished. Uncovering the miR-424/FGF2 pathway revealed a new mechanism how FOXO1 promotes bone formation and could potentially enhance bone repair.
miR-424 was furthermore recognized by Gosh et al. as an important hypoxamiR in human ECs by revealing a novel pathway for HIF regulation [101]. Under hypoxic conditions levels miR-424 or its rodent homolog, mu-miR-322, were found to be elevated in ECs and in ischemic tissues undergoing vascular remodeling in an experimental myocardial infarction rat model. It was elicited that the target of miR-424 is represented by cullin 2 (CUL2), which serves the purpose of stabilizing the hypoxic transcription factor HIF-1α. CUL2 is an essential component for assembling the ubiquitin ligase system, normally leading to its continuous degradation. MiR-424 expression was experimentally evaluated to be regulated by the transcription factor PU.1 that itself was found to be regulated by RUNX-1 and C/EBPα transcription factors. Ectopic expression of miR-424 in retrovirally-transduced HUVECs therefore stimulated angiogenesis in vivo in athymic mice that were subcutaneously implanted.
6. Outlook and Future Directions: MiRNAs in Therapeutic Applications
Bone is a highly vascularized tissue, and is thus reliant on the coordinated interaction between osteogenesis and angiogenesis which occurs between osteoblastic and ECs during development, remodeling and regeneration of the skeletal system [22,27,202]. Deciphering the molecular nature of the mechanisms that couple bone formation to blood vessel formation should therefore be of great interest to enhance bone formation capabilities in vitro and in vivo [203]. These specific underlying mechanisms are gaining growing attention under physiologic and pathologic conditions such as fracture healing, prevention of osteoporosis, tissue engineering or bone regeneration. Thus far, mostly a combination of different growth factors controlling osteogenesis and angiogenesis (e.g., such VEGF, angiopoietins, BMPS, RUNX2) have succeeded in achieving bone formation to a certain degree [33,204,205,206]. Nevertheless, each of these factors has individual roles at certain stages of development, and may disturb the subtle orchestration of required regulation steps.
MicroRNAs provide potent modifiers which coordinate a broad spectrum of biological processes. In contrast to transcription factors, single miRNAs may only modestly affect individual mRNA target expression. However, on the one hand a specific target mRNA can experience increased repression if it has multiple binding sites in the 3′-UTR. On the other hand, the same 3′-UTR of a gene can be targeted by many different miRNAs simultaneously that intervene with its regulation of expression [207]. Bioinformatic analysis of target prediction databases, such as TargetScan, miRanda and Pictar, suggest that a single average miRNA is capable of modulating up to several hundred target genes by perfect or imperfect base-pairing in combination with tissue-specific expression [208,209,210,211]. Additionally, by feedforward or feedback loops that form a regulatory network, miRNA effects can be amplified [212]. And it is estimated that approximately one-third of genome-encoded proteins are effected by miRNA regulation [213]. Therefore, these regulatory RNAs which control multiple endogenous signaling processes simultaneously possess a unique capacity to interfere with all cellular processes and have become an important tool in biological and medical research. As the application of miRNA-based methods for the treatment and monitoring of different pathological conditions is constantly becoming more prevalent, their therapeutic engagement could establish a refined method to stimulate inartificial bone development.
There is increasing evidence that miRNAs play important roles in controlling osteogenesis and angiogenesis. One study identified mutations in miR-2861 in two related adolescents that likely contributed to primary osteoporosis [214]. miR-2861 regulates osteoblast differentiation by targeting HDAC5, which enhances RUNX2 degradation. Moreover, a recent study showed accelerated osteoblast differentiation of hMSCs in a three-dimensional scaffold in vitro through manipulation of miR-148b and miR-489 expression [215]. Several studies also explored the use of miRNA mimicking or inhibitory agents for bone regeneration in animal studies. However, only a few studies have found that miRNAs are positive regulators of these processes. As outlined, miR-29b has been reported to promote osteoblast differentiation by targeting several well characterized inhibitors of bone formation in vitro [124]. Single site-specific delivery of miR-29b into a two-week post fracture callus by Lee et al. significantly improved mouse femoral fracture healing [216]. This was documented radiographically by a decrease of callus width and area; histomorphometrical and micro-computed tomographical analyses demonstrated increased bone volume fraction and bone mineral density of the callus. A single report further delineated the use of miRNAs as a therapeutic strategy to modulate angiogenesis-osteogenesis coupling during bone regeneration or repair in a subcutaneous assay in the mouse. The studies of Li et al. preferred miR-26a over miR-21 and miR-29b, which were all identified as the most potent candidates to mediate both angiogenesis and osteogenesis by microarray profiling of primary osteoblasts [217]. miR-26a was found to be a factor that is upregulated in newly-formed bone, and itself stimulates the expression of osteoblast-specific makers RUNX2, ALP, and OCN, mineralization of osteoblasts as well as VEGF secretion in murine primary BM-MSCs and in MC3T3-E1 cells in vitro. The enhancement of miR-26a expression in vivo by transfection of BM-MSCs with miR-26a mimicking agents led to complete repair of a critical-size calvarial bone defect, mainly due to simultaneously regulating endogenous angiogenesis-osteogenesis coupling. To date, its ability to integrate multiple signaling pathways thus makes miR-26a an ideal candidate for regenerating bone by miRNA-based therapy. Murata et al. detected significantly decreased levels of miR-92a in patients with trochanteric fractures, and investigated its significance in a mouse femoral fracture model [142]. Systemic as well as local administration of antimir-92a via LNA-stabilized oligonucleotide increased callus volume and enhanced fracture healing in the early phase by promoting neovascularization in the mouse femur. Nevertheless, as outlined previously, the direct osteogenesis-related molecular targets for this mechanism, which offers therapeutic potential for repairing bone, still remain to be clarified. In addition to the basic osteogenic functions of miR-31, Deng et al. investigated the therapeutic potential of hAD-MSC combined with beta-calcium triphosphate scaffolds in repairing a rat critical-sized calvarial defect [130,217]. The group reported that a knockdown of miR-31 significantly enhanced the repair of the defect, as could be noticed by an increased bone volume and mineral density in combination with a decreased scaffold. As the molecular basis of the osteogenic differentiation and bone regeneration program, in vitro results using lentiviral expression constructs revealed a BMP-2-inducible regulatory loop between Runx2, miR-31, as well as the miR-31 target gene, AT-rich sequence-binding protein 2 (Satb2). As a negative modulator of angiogenesis, Yoshizuka et al. locally administered miR-222 inhibitor mixed with atelocollagen to a rat femoral transverse fracture with the purpose of enhancing bone healing by stimulating osteogenesis, chondrogenesis, as well as angiogenesis [218]. Bone union at the fracture site with increased capillary density could be confirmed by radiographic, μCT and histological evaluation at 8 weeks after administration. Inhibition of miR-222 promoted osteogenic (RUNX2, COL1A1, OCN) or chondrogenic (COL2A1, aggrican, SOX9) differentiation in hMSCs, as determined by expression of osteogenic or chondrogenic markers, respectively.
In this review, miRNAs or “CouplingmiRs/CPLGmiRs” were identified that are known so far to regulate, either positively or negatively, angiogenesis as well as osteogenesis function—as potent molecular managers—that may simultaneously regulate multiple endogenous signaling cascades. This has been summarized in Figure 2, where the described miRNAs have also been depicted with regard to their allocation during the developmental fate of osteogenic or angiogenic cells. As for many miRNAs in general, also for coupling miRs/CPLGmiRs, most mechanisms of action are negative/inhibitory in nature for the regulation of osteogenesis as well as angiogenesis. Also noteworthy, but not surprising, is the finding that most miRNAs potentially involved in angiogenic-osteogenic coupling are encountered in the osteoblastic lineage rather than in chondroblasts or osteoclasts. And miRNAs are particularly present during the differentiation of osteoblasts, while they exert their modulatory effects during angiogenesis mostly in mature endothelial cells. Delivery of miRNAs may provide a way to maximally mimic the native bone development environment, and thus possess the therapeutic potential to enhance bone regeneration and repair. In contrast, miR inhibitors, i.e., antisense oligonucleotides directed against miRNAs, can be applied in the form of antagomiRs or LNA-antimiRs [58,179,219]. RNA/DNA hybrids modified by this locked nucleic acid-technology represent single stranded modified antisense oligonucleotides with increased stability and affinity and also facilitate cell penetration. Furthermore, they are biocompatible, as they cause no cellular immune response, are highly soluble, and have already been tested by various in vitro and in vivo studies as well as clinical trials (e.g., miR-122 LNA anti-miRNA oligonucleotides for Hepatitis C virus or miR-34 for solid cancers) [220,221]. Additionally, any new findings in this field of research might additionally yield considerable anti-angiogenic targets for tumor therapies, as tumor angiogenesis—i.e., the formation of tumor-associated angiogenic vessels—is a key requirement in tumor growth, progression, and metastasis. As an example, it would be very favorable to identify angiomiRs that negatively regulate VEGF expression in combination with current anti-VEGF therapies. Inversely, the identification of novel oncomiRs could also pave the way and lead to the description of unknown miRNAs with a function in bone angiogenesis [222,223,224]. Currently, microRNA-targeting therapy is still in development due to new challenges compared to conventional drugs, and results of experimental studies in animal models need to be transferred to clinical applications. Nevertheless, in addition to their role as potential angiogenic or bone regenerative therapeutic targets, miRNAs are emerging as distinguished disease biomarkers relevant to specific physiologic or pathologic conditions which could serve for immediate use in cardiovascular and bone diseases.
Author Contributions
L.F.F. was responsible for collecting literature, drafting, writing, reviewing, and illustration of the manuscript.
Funding
We acknowledge support by Open Access Publication Fund of University of Münster.
Conflicts of Interest
The author declares no conflict of interest.
References
- Crane, G.; Ishaug, S.; Mikos, A. Bone tissue engineering. Nat. Med. 1995, 1, 1322–1324. [Google Scholar] [CrossRef] [PubMed]
- Rouwkema, J.; Rivron, N.C.; Blitterswijk, C.A. Vascularization in tissue engineering. Trends Biotechnol. 2008, 26, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Hertig, A. Angiogenesis in the early human chorion and the primary placenta of the macaque monkey. Contrib. Embryol. 1935, 25, 37–81. [Google Scholar]
- Chung, A.S.; Ferrara, N. Developmental and pathological angiogenesis. Annu. Rev. Cell Dev. Biol. 2011, 27, 563–584. [Google Scholar] [CrossRef]
- Lian, J.B.; Stein, G.S.; van Wijnen, A.J.; Stein, J.L.; Hassan, M.Q.; Gaur, T.; Zhang, Y. MicroRNA control of bone formation and homeostasis. Nat. Rev. Endocrinol. 2013, 8, 212–227. [Google Scholar] [CrossRef] [PubMed]
- Kronenberg HM Developmental regulation of the growth plate. Nature 2003, 423, 332–336. [CrossRef] [PubMed]
- Berendesen, A.; Olsen, B. Bone development. Bone 2015, 80, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Olsen, B.; Reginato, A.; Wang, W. Bone development. Annu. Rev. Cell Dev. Biol. 2000, 16, 191–220. [Google Scholar] [CrossRef]
- Karsenty, G. The complexities of skeletal biology. Nature 2003, 423, 316–318. [Google Scholar] [CrossRef]
- Helms, J.; Schneider, R. Cranial skeletal biology. Nature 2003, 423, 326–331. [Google Scholar] [CrossRef]
- Maes, C.; Carmeliet, P.; Moermans, K.; Stockmans, I.; Smets, N. Impaired angiogenesis and endochondral bone formation in mice lacking the vascular endothelial growth factor isoforms VEGF 164 and VEGF 188. Mech Dev. 2002, 111, 61–73. [Google Scholar] [CrossRef]
- Gerber, H.; Ferrara, N. Angiogenesis and Bone Growth. TCM 2000, 10, 223–228. [Google Scholar] [CrossRef]
- Brandi, M.; Collin-Osdoby, P. Vascular biology and the skeleton. J. Bone Miner. Res. 2006, 21, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Sivaraj, K.K.; Adams, R.H. Blood vessel formation and function in bone. Development 2016, 143, 2706–2715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schipani, E.; Maes, C.; Carmeliet, G.; Semenza, G.L. Regulation of osteogenesis-angiogenesis coupling by HIFs and VEGF. J. Bone Miner. Res. 2009, 24, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Schipani, E.; Wu, C.; Rankin, E.B.; Giaccia, A.J. Regulation of Bone Marrow Angiogenesis by Osteoblasts during Bone Development and Homeostasis. Front. Endocrinol. 2013, 4, 85. [Google Scholar] [CrossRef]
- Kusumbe, A.P.; Ramasamy, S.K.; Adams, R.H. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nature 2014, 507, 323–328. [Google Scholar] [CrossRef] [Green Version]
- Carmeliet, P.; Ferreira, V.; Breier, G.; Pollefeyt, S.; Kieckens, L.; Gertsenstein, M.; Fahrig, M.; Vandenhoeck, A.; Harpal, K.; Eerhardt, C.; et al. Abnormal blood vessel development and lethality in embryos lacking a single VEGF allele. Nature 1996, 380, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Carulli, C.; Innocenti, M.; Brandi, M.L. Bone vascularization in normal and disease conditions. Front. Endocrinol. 2013, 4, 1–10. [Google Scholar] [CrossRef]
- Ding, W.-G.; Yan, W.; Wei, Z.-X.; Liu, J.-B. Difference in intraosseous blood vessel volume and number in osteoporotic model mice induced by spinal cord injury and sciatic nerve resection. J. Bone Miner. Metab. 2012, 30, 400–407. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, F.; Zhang, P.; Wang, H.; Qu, Z.; Jia, P.; Yao, Z.; Shen, G.; Li, G.; Zhao, G.; et al. Human type H vessels are a sensitive biomarker of bone mass. Cell Death Dis. 2017, 8, e2760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stegen, S.; Van Gastel, N.; Carmeliet, G. Bringing new life to damaged bone: The importance of angiogenesis in bone repair and regeneration. Bone 2014, 70, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Olsen, B.R. Osteoblast-derived VEGF regulates osteoblast differentiation and bone formation during bone repair. J. Clin. Invest. 2016, 126, 509–526. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.; Olsen, B.R. The roles of vascular endothelial growth factor in bone repair and regeneration. Bone 2016, 91, 30–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maes, C.; Kobayashi, T.; Selig, M.; Torrekens, S.; Roth, S.; Mackem, S.; Carmeliet, G.; Kronenberg, H. Osteoblast precursors, but not mature osteoblasts, move into developing and fractured bones along with invading blood vessels. Dev. Cell 2010, 19, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Maes, C.; Carmeliet, G.; Schipani, E. Hypoxia-driven pathways in bone development, regeneration and disease. Nat. Rev. Rheumatol. 2012, 8, 358–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wan, C.; Deng, L.; Liu, X.; Cao, X.; Gilbert, S.R.; Bouxsein, M.L.; Faugere, M.; Guldberg, R.E.; Gerstenfeld, L.C.; et al. The hypoxia-inducible factor α pathway couples angiogenesis to osteogenesis during skeletal development. J. Clin. Invest. 2007, 117, 1616–1626. [Google Scholar] [CrossRef] [Green Version]
- Pugh, C.W.; Ratcliffe, P.J. Regulation of angiogenesis by hypoxia: Role of the HIF system. Nat. Med. 2003, 9, 677–684. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.; Lecouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef]
- Ornitz, D.; Marie, P. Fibroblast growth factor signaling in skeletal development and disease. Genes Dev. 2015, 29, 1463–1468. [Google Scholar] [CrossRef]
- Ramasamy, S.K.; Kusumbe, A.P.; Wang, L.; Adams, R.H. Endothelial Notch activity promotes angiogenesis and osteogenesis in bone. Nature 2014, 507, 376–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortega, N.; Behonick, D.; Werb, Z. Matrix remodeling during endochondral ossification. Trends Cell Biol. 2004, 14, 86–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinheinz, J.; Stratmann, U.; Joos, U.; Wiesmann, H.-P. VEGF-Activated Angiogenesis During Bone Regeneration. J. Oral Maxillofac. Surg. 2005, 63, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- Maes, C.; Coenegrachts, L.; Stockmans, I.; Daci, E.; Luttun, A.; Petryk, A.; Gopalakrishnan, R.; Moermans, K.; Smets, N.; Verfaillie, C.M.; et al. Placental growth factor mediates mesenchymal cell development, cartilage turnover, and bone remodeling during fracture repair. J. Clin. Invest. 2006, 116, 16–18. [Google Scholar] [CrossRef]
- Kingsley, D. What do BMPs do in mammals? Clues from the mouse short-ear mutation. Trends Genet. 1994, 10, 16–21. [Google Scholar] [CrossRef]
- Hassan, M.Q.; Tye, C.E.; Stein, G.S.; Lian, J.B. Non-coding RNAs: Epigenetic regulators of bone development and homeostasis. Bone 2015, 81, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Papaioannou, G.; Mirzamohammadi, F.; Kobayashi, T. MicroRNAs involved in bone formation. Cell Mol. Life Sci. 2014, 71, 4747–4761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papaioannou, G. miRNAs in Bone Development. Curr. Genom. 2015, 16, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. MicroRNAs: Tiny regulators with great potential. Cell 2001, 107, 823–826. [Google Scholar] [CrossRef]
- Lau, N.C.; Lim, L.P.; Weinstein, E.G.; Bartel, D.P. An abundant class of tiny RNAs with probable regulatory roles in Caenorhabditis elegans. Science 2001, 294, 858–862. [Google Scholar] [CrossRef]
- Bartel, D.P.; Lee, R.; Feinbaum, R. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function Genomics: The miRNA Genes. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.C.; Ambros, V. An extensive class of small RNAs in Caenorhabditis elegans. Science 2001, 294, 858–862. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, L.A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [PubMed]
- Hutvagner, G.; McLachlan, J.; Pasquinelli, A.E.; Balint, E.; Tuschl, T.; Zamore, P.D. A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNA. Science 2001, 293, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Grishok, A.; Pasquinelli, A.E.; Conte, D.; Li, N.; Parrish, S.; Ha, I.; Baillie, D.L.; Fire, A.; Ruvkun, G.; Mello, C. Genes and mechanisms related to RNA interference regulate expression of the small temporal RNAs that control C. elegans developmental timing. Cell 2001, 106, 23–34. [Google Scholar] [CrossRef]
- Knight, S.W.; Bass, B.L. A role for the RNase III enzyme DCR-1 in RNA interference and germ line development in Caenorhabditis elegans. Science 2001, 293, 2269–2271. [Google Scholar] [CrossRef]
- Yang, J.S.; Lai, E.C. Alternative miRNA biogenesis pathways and the interpretation of core miRNA pathway mutants. Mol. Cell 2011, 43, 892–903. [Google Scholar] [CrossRef]
- Lai, E.C. Micro RNAs are complementary to 3´UTR sequence motifs that mediate negative post-transcriptional regulation. Nat. Genet. 2002, 30, 363–364. [Google Scholar] [CrossRef]
- Bernstein, E.; Caudy, A.A.; Hammond, S.M.; Hannon, G.J. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 2001, 409, 363–366. [Google Scholar] [CrossRef]
- Wang, Y.; Medvid, C.; Melton, R.; Jaenisch, R.; Blelloch, R. DGCR8 is essential for microRNA biogenesis and silencing of embryonic stem cell self-renewal. Nat. Genet. 2007, 39, 380–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Förstemann, K.; Tomari, Y.; Du, T.; Vagin, V.V.; Denli, A.M.; Bratu, D.P.; Klattenhoff, C.; Theurkauf, W.E.; Zamore, P.D. Normal microRNA maturation and germ-line stem cell maintenance requires Loquacious, a double stranded RNA-binding domain protein. PLoS Biol. 2005, 3, e236. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Shen, X.J.; Zou, Q.; Wang, S.P.; Tang, S.M.; Zhang, G.Z. Biological functions of microRNAs: A review. J. Physiol. Biochem. 2011, 67, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Melton, C.; Judson, R.L.; Blelloch, R. Opposing micro-RNA families regulate self-renewal in mouse embryonic stem cells. Nature 2010, 463, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Brennecke, J.; Hipfner, D.R.; Stark, A.; Russell, R.B.; Cohen, S.M. bantam encodes a developmentally regulated microRNA that controls cell proliferation and regulates the proapoptotic gene hid in Drosophila. Cell 2003, 113, 25–36. [Google Scholar] [CrossRef]
- Hipfner, D.R.; Weigmann, K.; Cohen, S.M. The bantam gene regulates Drosophila growth. Genetics 2002, 161, 1527–1537. [Google Scholar] [PubMed]
- Esau, C.; Davis, S.; Murray, S.F.; Yu, X.X.; Pandey, S.K.; Pear, M.; Watts, L.; Booten, S.L.; Graham, M.; McKay, R.; et al. miR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab. 2006, 3, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Krützfeldt, J.; Rajewsky, N.; Braich, R.; Rajeev, K.G.; Tuschl, T.; Manoharan, M.; Stoffel, M. Silencing of microRNAs in vivo with ´antagomirs´. Nature 2005, 438, 685–689. [Google Scholar] [CrossRef]
- Miska, E.A. How microRNAs control cell division, differentiation and death. Curr. Opin. Genet. Dev. 2005, 15, 563–568. [Google Scholar] [CrossRef]
- Chen, C.Z.; Li, L.; Lodish, L.F.; Bartel, D. MicroRNAs modulate hematopoietic lineage differentiation. Science 2004, 303, 83–86. [Google Scholar] [CrossRef]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez-Garcia, I.; Miska, E.A. MicroRNA functions in animal development and human disease. Development 2005, 132, 4653–4662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gennari, L.; Bianciardi, S.; Merlotti, D. MicroRNAs in bone diseases. Osteoporos. Int. 2017, 28, 1191–1213. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.; Kalomoiris, S.; Nolta, J.; Fierro, F. Concise Review: MicroRNA Function in Multipotent Mesenchymal Stromal Cells. Stem Cells 2014, 32, 1074–1082. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.; Gao, D.; Gao, C.; Wei, P.; Niu, M.; Shuai, C. MicroRNAs regulate signaling pathways in osteogenic differentiation of mesenchymal stem cells (Review). Mol. Med. Rep. 2016, 14, 623–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, S.; Deng, Y.; Gu, P.; Fan, X. MicroRNAs Regulate Bone Development and Regeneration. Int. J. Mol. Sci. 2015, 16, 8227–8253. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.; Chen, X.; Yu, X. MicroRNAs in Osteoclastogenesis and Function: Potential Therapeutic Targets for Osteoporosis. Int. J. Mol. Sci. 2016, 17, 349. [Google Scholar] [CrossRef]
- Dong, S.; Yang, B.; Guo, H.; Kang, F. MicroRNAs regulate osteogenesis and chondrogenesis. Biochem. Biophys. Res. Commun. 2012, 418, 587–591. [Google Scholar] [CrossRef]
- Kiga, K.; Mimuro, H.; Suzuki, M.; Shinozaki-Ushiku, A.; Kobayashi, T.; Sanada, T.; Kim, M.; Ogawa, M.; Iwasaki, Y.W.; Kayo, H.; et al. Epigenetic silencing of miR-210 increases the proliferation of gastric epithelium during chronic Helicobacter pylori infection. Nat. Commun. 2014, 5, 4497. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Qiu, M.; Dou, C.; Cao, Z.; Dong, S. MicroRNAs in Bone Balance and Osteoporosis. Drug Dev. Res. 2015, 76, 235–245. [Google Scholar] [CrossRef]
- Nugent, M. MicroRNAs and Fracture Healing. Calcif. Tissue Int. 2017, 101, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Tian, B.O.; Qu, X.; Liu, F.; Tang, T.; Qin, A.N.; Zhu, Z.; Dai, K. MicroRNAs play a role in chondrogenesis and osteoarthritis (Review). Int. J. Mol. Med. 2014, 34, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Min, Z.; Zhang, R.; Yao, J.; Jiang, C.; Guo, Y.; Cong, F.; Wang, W.; Tian, J.; Zhong, N.; Sun, J.; et al. MicroRNAs associated with osteoarthritis differently expressed in bone matrix gelatin (BMG) rat model. Int. J. Clin. Exp. Med. 2015, 8, 1009–1017. [Google Scholar] [PubMed]
- Seeliger, C.; Balmayor, E.; van Griensven, M. miRNAs Related to Skeletal Diseases. Stem Cells Dev. 2016, 25, 1261–1281. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Cheresh, D.A. Emerging Role of Micro-RNAs in the Regulation of Angiogenesis. Genes Cancer 2011, 2, 1134–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Olson, E.N. AngiomiRs—Key Regulators of Angiogenesis. Curr. Opin. Genet. Dev. 2009, 19, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Small, E.M.; Olson, E.N. Pervasive roles of microRNAs in cardiovascular biology. Nature 2011, 469, 336–342. [Google Scholar] [CrossRef] [Green Version]
- Weis, S.M.; Caheresh, D.A. Tumor angiogenesis: Molecular pathways and therapeutic targets. Nat. Med. 2011, 17, 1359–1370. [Google Scholar] [CrossRef]
- Salinas-Vera, Y.; Marchat, L.; Gallardo-Rincon, D.; Ruiz-Garcia, E.; Astudillo- De La Vega, H.; Echavarria-Zepeda, R.; Lopez-Camarillo, C. AngiomiRs: MicroRNAs driving angiogenesis in cancer (Review). Int. J. Mol. Med. 2018, 2018. [Google Scholar] [CrossRef]
- Suarez, Y.; Sessa, W.C. MicroRNAs As Novel Regulators of Angiogenesis. Circ. Res. 2009, 104, 442–454. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Yang, D.; Na, S.; Sandusky, G.; Zhang, Q.; Zhao, G. Dicer is required for embryonic angiogenesis during mouse development. J. Biol. Chem. 2005, 280, 9330–9335. [Google Scholar] [CrossRef] [PubMed]
- Albinsson, S.; Suarez, Y.; Skoura, A.; Offermann, S.; Miano, J.M.; Sessa, W.C. MicroRNAs are necessary for vascular smooth muscle growth, differentiation, and function. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnnaly, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The endothelial-specific microRNA miR-126 governs vascular integrity and angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Suarez, Y.; Wang, C.; Manes, T.D.; Pober, J.S. Cutting edge: TNF-induced microRNAs regulate TNF-induced expression of E-selectin and intercellular adhesion molecule-1 on human endothelial cells: Feedback control of inflammation. J. Immunol. 2010, 184, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Suarez, Y.; Fernandez-Hernando, C.; Pober, J.; Sessa, W. Dicer dependent microRNAs regulate gene expression and functions in human endothelial cells. Circ. Res. 2007, 100, 1164–1173. [Google Scholar] [CrossRef] [PubMed]
- Greco, S.; Gaetano, C.; Martelli, F. HypoxamiR Regulation and Function in Ischemic Cardiovascular Diseases. Antioxid. Redox Signal. 2014, 21, 1202–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samanta, S.; Balasubramanian, S.; Rajasingh, S.; Patel, U.; Dhanasekaran, A.; Dawn, B.; Rajasingh, J. MicroRNA: A new therapeutic strategy for cardiovascular diseases. Trends Cardiovasc. Med. 2016, 26, 407–419. [Google Scholar] [CrossRef] [Green Version]
- Suarez, Y.; Fernandez-Hernando, C.; Yu, J.; Gerber, S.A.; Harrison, K.D.; Pober, J.S.; Iruela-Arispe, M.L.; Merkenschlager, M.; Sessa, W.C. Dicer-dependent endothelial microRNAs are necessary for postnatal angiogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 14082–14087. [Google Scholar] [CrossRef] [Green Version]
- Würdinger, T.; Tannous, B.A.; Saydam, O.; Skog, J.; Grau, S.; Soutschek, J.; Weissleder, R.; Breakefield, X.O.; Krichevsky, A.M. miR-296 regulates growth factor receptor overexpression in angiogenic endothelial cells. Cancer Cell 2008, 14, 382–393. [Google Scholar] [CrossRef]
- Anand, S.; Cheresh, D.A. MicroRNA-mediated Regulation of the Angiogenic Switch. Curr. Opin. Hematol. 2011, 18, 171–176. [Google Scholar] [CrossRef]
- Nallamshetty, S.; Chan, S.Y.; Loscalzo, J. Hypoxia: A master regulator of microRNA biogenesis and activity. Free Radic. Biol. Med. 2013, 64, 20–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madanecki, P.; Kapoor, N.; Bebok, Z.; Ochocka, R.; Collawn, J.F.; Bartoszewski, R. Regulation of angiogenesis by hypoxia: The role of microRNA. Cell. Mol. Biol. Lett. 2013, 18, 47–57. [Google Scholar] [CrossRef] [PubMed]
- el Azzouzi, H.; Leptidis, S.; Doevendans, P.A.; De Windt, L.J. HypoxamiRs: Regulators of cardiac hypoxia and energy metabolism. Trends Endocrinol. Metab. 2015, 26, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Greco, S.; Martelli, F. MicroRNAs in Hypoxia Response. Antioxid. Redox Signal. 2014, 21, 1164–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collet, G.; Skrzypek, K.; Grillon, C.; Matejuk, A.; El Hafni-Rahbi, B.; Fayel, N.L.; Kieda, C. Hypoxia control to normalize pathologic angiogenesis: Potential role for endothelial precursor cells and miRNAs regulation. Vascul. Pharmacol. 2012, 56, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Bertero, T.; Rezzonico, R.; Pottier, N.; Mari, B. Impact of MicroRNAs in the Cellular Response to Hypoxia. Int. Rev. Cell Mol. Biol. 2017, 333, 91–158. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z.; Lv, Q.; Ye, W.; Wong, A.C.-K.; Cai, G.; Gu, D.; Ji, Y.; Zhao, C.; Wang, J.; Yang, B.B.; et al. MiRNA-Directed Regulation of VEGF and Other Angiogenic Factors under Hypoxia. PloS ONE 2006, 1, e116. [Google Scholar] [CrossRef]
- Loscalzo, J. The cellular response to hypoxia: Tuning the system with microRNAs. J. Clin. Invest. 2010, 120, 3815–3817. [Google Scholar] [CrossRef]
- Devlin, C.; Greco, S.; Martelli, F.; Ivan, M. MiR-210: More than a silent player in hypoxia. IUBMB Life 2011, 63, 94–100. [Google Scholar] [CrossRef]
- Chan, S.; Loscalzo, J. MicroRNA-210: A unique and pleiotropic hypoxamir. Cell Cycle 2010, 9, 1072–1083. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, G.; Subramanian, I.V.; Adhikari, N.; Zhang, X.; Joshi, H.P.; Basi, D.; Chandrashekhar, Y.S.; Hall, J.L.; Roy, S.; Zeng, Y.; et al. Hypoxia-induced microRNA-424 expression in human endothelial cells regulates HIF- α isoforms and promotes angiogenesis. J. Clin. Invest. 2010, 120, 4141–4154. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, A.; Yanagisawa, K.; Tanaka, M.; Cao, K.; Matsuyama, Y.; Goto, H.; Takahashi, T. Identification of hypoxia-inducible factor-1alpha as a novel target for miR-17-92 microRNA cluster. Cancer Res. 2008, 68, 5540–5545. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Tsai, M.; Hung, P.; Kao, S.; Liu, T.; Wu, K.; Chiou, S.; Lin, S.; Chang, K. miR31 ablates expression of the HIF regulatory factor FIH to activate the HIF pathway in head and neck carcinoma. Cancer Res. 2010, 70, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Yuva-Aydemir, Y.; Simkin, A.; Gascon, E.; Gao, F.-B. MicroRNA-9: Functional evolution of a conserved small regulatory RNA. RNA Biol. 2011, 8, 557–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, R.; Kan, Q.; Sun, Y.; Wang, S.; Zhang, G.; Peng, T.; Jia, Y. MiR-9 promotes the neural differentiation of mouse bone marrow mesenchymal stem cells via targeting zinc finger protein 521. Neurosci. Lett. 2012, 515, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Lu, D.; Guo, H.; Miao, W.; Wu, G. MicroRNA-9 regulates osteoblast differentiation and angiogenesis via the AMPK signaling pathway. Mol. Cell Biochem. 2016, 411, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xu, H.; Kou, J.; Wang, Q.; Zheng, X.; Yu, T. MiR-9 promotes osteoblast differentiation of mesenchymal stem cells by inhibiting DKK1 gene expression. Mol. Biol. Rep. 2016, 43, 939–946. [Google Scholar] [CrossRef]
- Zhuang, G.; Wu, X.; Jiang, Z.; Kasman, I.; Yao, J.; Guan, Y.; Oeh, J.; Modrusan, Z.; Bais, C.; Sampath, D.; et al. Tumour-secreted miR-9 promotes endothelial cell migration and angiogenesis by activating the JAK-STAT pathway. EMBO J. 2012, 31, 3513–3523. [Google Scholar] [CrossRef] [Green Version]
- Jones, S.; Watkins, G.; Le Good, N.; Roberts, S.; Murphy, C.; Brockbank, S.; Needham, M.; Read, S.; Newham, P. The identification of differentially expressed microRNA in osteoarthritic tissue that modulate the production of TNF-alpha and MMP13. Osteoarthr. Cartil. 2009, 17, 464–472. [Google Scholar] [CrossRef]
- Wang, S.; Tang, C.; Zhang, Q.; Chen, W. Reduced miR-9 and miR-181a expression down-regulates Bim concentration and promote osteoclasts survival. Int. J. Clin. Exp. Pathol. 2014, 7, 2209–2218. [Google Scholar]
- Li, J.; Zhang, Y.; Zhao, Q.; Wang, J.; He, X. MicroRNA-10a Influences Osteoblast Differentiation and Angiogenesis by Regulating ß-Catenin Expression. Cell. Physiol. Biochem. 2015, 37, 2194–2208. [Google Scholar] [CrossRef]
- Day, T.F.; Guo, X.; Garrett-Beal, L.; Yang, Y. Wnt/beta-catenin signaling in mesenchymal progenitors controls osteoblast and chondrocyte differentiation during vertebrate skeletogenesis. Dev. Cell 2005, 8, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Deng, S.; Ma, Q.; Zhang, T.; Jia, C.; Zhuo, D.; Yang, F.; Wei, J.; Wang, L.; Dykxhoorn, D.M.; et al. MicroRNA-10A* and MicroRNA-21 Modulate Endothelial Progenitor Cell Senescence Via Suppressing High-Mobility Group A2. Circ. Res. 2013, 112, 152–164. [Google Scholar] [CrossRef]
- Hassel, D.; Cheng, P.; White, M.P.; Ivey, K.N.; Kroll, J.; Augustin, H.G.; Katus, H.A.; Stainier, D.Y.R.; Srivastava, D. MicroRNA-10 Regulates the Angiogenic Behavior of Zebrafish and Human Endothelial Cells by Promoting Vascular Endothelial Growth Factor Signaling. Circ. Res. 2012, 111, 1421–1433. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ling, C.C.; Li, L.; Qin, Y.; Qi, J.; Liu, X.; You, B.; Shi, Y.; Zhang, J.; Xu, Q.J.H.; et al. MicroRNA-10a/10b represses a novel target gene mib1 to regulate angiogenesis. Cardiovasc. Res. 2016, 110, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Fu, W.; He, M.; Xie, W.; Lv, Q.; Li, G.; Wang, H.; Lu, G.; Hu, X.; Jiang, S.; et al. MiRNA-20a promotes osteogenic differentiation of human mesenchymal stem cells by co-regulating BMP signaling. RNA Biol. 2011, 8, 829–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doebele, C.; Bonauer, A.; Fischer, A.; Scholz, A.; Ress, Y.; Urbich, C.; Hofmann, W.-K.; Zeiher, A.M.; Dimmeler, S. Members of the microRNA-17-92 cluster exhibit a cell-intrinsic antiangiogenic function in endothelial cells. Blood 2010, 115, 4944–4950. [Google Scholar] [CrossRef] [Green Version]
- Deng, H.-T.; Liu, H.-L.; Zhai, B.-B.; Zhang, K.; Xu, G.-C.; Peng, X.-M. Vascular endothelial growth factor suppresses TNFSF15 production in endothelial cells by stimulating miR-31 and miR-20a expression via activation of Akt and Erk signals. FEBS Open Bio. 2017, 7, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Luzi, E.; Marini, F.; Sala, S.C.; Tognarini, I.; Galli, G.; Brandi, M.L. Osteogenic Differentiation of Human Adipose Tissue–Derived Stem Cells Is Modulated by the miR-26a Targeting of the SMAD1 Transcription Factor. J. Bone Miner. Res. 2008, 23, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Luzi, E.; Marini, F.; Tognarini, I.; Galli, G.; Falchetti, A.; Brandi, M.L. The regulatory network menin-microRNA 26a as a possible target for RNA-based therapy of bone diseases. Nucleic. Acid Ther. 2012, 22, 103–108. [Google Scholar] [CrossRef]
- Su, X.; Liao, L.; Shuai, Y.; Jing, H.; Liu, S.; Zhou, H.; Liu, Y.; Jin, Y. MiR-26a functions oppositely in osteogenic differentiation of BMSCs and ADSCs depending on distinct activation and roles of Wnt and BMP signaling pathway. Cell Death Dis. 2015, 6, e1851. [Google Scholar] [CrossRef] [PubMed]
- Trompeter, H.-I.; Dreesen, J.; Hermann, E.; Iwaniuk, K.M.; Hafner, M.; Renwick, N.; Tuschl, T.; Wernet, P. MicroRNAs miR-26a, miR-26b, and miR-29b accelerate osteogenic differentiation of unrestricted somatic stem cells from human cord blood. BMC Genom. 2013, 14, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Icli, B.; Wara, A.K.M.; Moslehi, J.; Sun, X.; Plovie, E.; Cahill, M.; Marchini, J.F.; Schissler, A.; Padera, R.F.; Shi, J.; et al. MicroRNA-26a regulates pathological and physiological angiogenesis by targeting BMP/SMAD1 signaling. Circ. Res. 2013, 113, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hassan, M.Q.; Jafferji, M.; Aqeilan, R.I.; Garzon, R.; Croce, C.M.; Van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Lian, J.B. Biological Functions of miR-29b Contribute to Positive Regulation of Osteoblast Differentiation. J. Biol. Chem. 2009, 284, 15676–15684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, M.; Pitari, M.R.; Amodio, N.; Di Martino, T.M.; Conforti, F.; Leone, E.; Botta, C.; Paolino, F.M.; Giudice, T.D.E.L.; Iuliano, E.; et al. miR-29b Negatively Regulates Human Osteoclastic Cell Differentiation and Function: Implications for the Treatment of Multiple Myeloma-Related Bone Disease. J. Cell. Physiol. 2013, 228, 1506–1515. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Cai, H.-X.; Gao, S.; Yang, G.-L.; Deng, H.-T.; Xu, G.-C.; Han, J.; Zhang, Q.-Z.; Li, L.-Y. TNSF15 suppresses VEGF production in endothelial cells by stimulating miR-29b expression via activation of JNK-GATA3 Signals. Oncotarget 2016, 7, 69436–69449. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cai, B.; Shen, L.; Dong, Y.; Lu, Q.; Sun, S.; Liu, S.; Ma, S.; Ma, P.X.; Chen, J. MiRNA-29b suppresses tumor growth through simultaneously inhibiting angiogenesis and tumorigenesis by targeting Akt3. Cancer Lett. 2017, 397, 111–119. [Google Scholar] [CrossRef]
- Granchi, D.; Ochoa, G.; Leonardi, E.; Devescovi, V.; Baglìo, S.R.; Osaba, L.; Baldini, N.; Ciapetti, G. Gene expression patterns related to osteogenic differentiation of bone marrow-derived mesenchymal stem cells during ex vivo expansion. Tissue Eng. Part. C Methods 2010, 16, 511–523. [Google Scholar] [CrossRef]
- Baglìo, S.R.; Devescovi, V.; Granchi, D.; Baldini, N. MicroRNA expression profiling of human bone marrow mesenchymal stem cells during osteogenic differentiation reveals Osterix regulation by miR-31. Gene 2013, 527, 321–331. [Google Scholar] [CrossRef]
- Deng, Y.; Wu, S.; Zhou, H.; Bi, X.; Wang, Y.; Hu, Y.; Gu, P.; Fan, X. Effects of a miR-31, Runx2, and Satb2 regulatory loop on the osteogenic differentiation of bone mesenchymal stem cells. Stem Cells Dev. 2013, 22, 2278–2286. [Google Scholar] [CrossRef]
- Luo, J.; Lin, J.; Paranya, G.; Bischoff, J. Angiostatin Upregulates E-Selectin in Proliferating Endothelial Cells. Biochem. Biophys. Res. Commun. 1998, 911, 906–911. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Holmstrom, K.; Qiu, W.; Ditzel, N.; Shi, K.; Hokland, L.; Kassem, M. MicroRNA-34a Inhibits Osteoblast Differentiation and In Vivo Bone Formation of Human Stromal Stem Cells. Stem Cells 2014, 32, 902–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.; Chen, H.; Huang, P.; Qi, J.; Qian, N.; Deng, L.; Guo, L. Glucocorticoids impair bone formation of bone marrow stromal stem cells by reciprocally regulating microRNA-34a-5p. Osteoporos. Int. 2016, 27, 1493–1505. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Jia, L.; Zheng, Y.; Jin, C.; Liu, Y.; Liu, H.; Zhou, Y. Mir-34a Promotes Osteogenic Differentiation of Human Adipose-Derived Stem Cells via the RBP2/NOTCH I/CYCLIN DI Coregulatory Network. Stem Cell Rep. 2016, 7, 236–248. [Google Scholar] [CrossRef] [PubMed]
- Engin, F.; Yao, Z.; Yang, T.; Zhou, G.; Bertin, T.; Jiang, M.M.; Chen, Y.; Wang, L.; Zheng, H.; Sutton, R.E.; et al. Dimorphic effects of Notch signaling in bone homeostasis. Nat. Med. 2008, 14, 299–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galindo, M.; Pratap, J.; Young, D.W.; Hovhannisyan, H.; Im, H.J.; Choi, J.Y.; Lian, J.B.; Stein, J.L.; Stein, G.S.; van Wijnen, A.J. The bone-specific expression of Runx2 oscillates during the cell cycle to support a G1-related antiproliferative function in osteoblasts. J. Biol. Chem. 2005, 280, 20274–20285. [Google Scholar] [CrossRef] [PubMed]
- Zha, X.; Sun, B.; Zhang, R.; Li, C.; Yan, Z.; Chen, J. Regulatory Effect of MicroRNA-34a on Osteogenesis and Angiogenesis in Glucocorticoid-Induced Osteonecrosis of the Femoral Head. J. Orthop. Res. 2018, 36, 417–424. [Google Scholar] [CrossRef]
- Zhao, T.; Li, J.; Chen, A.F. MicroRNA-34a induces endothelial progenitor cell senescence and impedes its angiogenesis via suppressing silent information regulator 1. Am. J. Endocrinol. Metab. 2010, 299, E110–E116. [Google Scholar] [CrossRef] [Green Version]
- Mattagajasingh, I.; Kim, C.; Naqvi, A.; Yamamori, T.; Hoffman, T.; Jung, S.; DeRicco, J.; Kasuno, K.; Irani, K. SIRT1 promotes endothelium-dependent vascular relaxation by activating endothelial nitric oxide synthase. Proc. Natl. Acad. Sci. USA 2007, 104, 14855–14860. [Google Scholar] [CrossRef] [Green Version]
- Kumar, B.; Yadav, A.; Lang, J.; Teknos, T.N.; Kumar, P. Dysregulation of MicroRNA-34a Expression in Head and Neck Squamous Cell Carcinoma Promotes Tumor Growth and Tumor Angiogenesis. PLoS ONE 2012, 7, e37601. [Google Scholar] [CrossRef]
- Chai, Z.T.; Kong, J.; Zhu, X.D.; Zhang, Y.Y.; Lu, L.; Zhou, J.M.; Wang, L.R.; Zhang, K.Z.; Zhang, Q.B.; Ao, J.Y.; et al. MicroRNA-26a Inhibits Angiogenesis by Down-Regulating VEGFA through the PIK3C2α/Akt/HIF-1α Pathway in Hepatocellular Carcinoma. PLoS ONE 2013, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Ito, H.; Yoshitomi, H.; Yamamoto, K.; Fukuda, A.; Yoshikawa, J.; Furu, M.; Ishikawa, M.; Shibuya, H.; Matsuda, S. Inhibition of miR-92a enhances fracture healing via promoting angiogenesis in a model of stabilized fracture in young mice. J. Bone Miner. Res. 2014, 29, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, N.; Klink, G.; Glukhanyuk, E.; Lopatina, T.; Anastassia, E.; Akopyan, Z.; Tkachuk, V. miR-92a regulates angiogenic activity of adipose-derived mesenchymal stromal cells. Exp. Cell Res. 2015, 339, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Daniel, J.-M.; Penzkofer, D.; Teske, R.; Dutzmann, J.; Koch, A.; Bielenberg, W.; Bonauer, A.; Boon, R.A.; Fischer, A.; Bauersachs, J.; et al. Inhibition of miR-92a improves re-endothelialization and prevents neointima formation following vascular injury. Cardiovasc. Res. 2014, 103, 564–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goettsch, C.; Rauner, M.; Pacyna, N.; Hempel, U.; Bornstein, S.R.; Hofbauer, L.C. MiR-125b regulates calcification of vascular smooth muscle cells. Am. J. Pathol. 2011, 179, 1594–1600. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, Y.; Yagi, K.; Tokuzawa, Y.; Kanesaki-Yatsuka, Y.; Suda, T.; Katagiri, T.; Fukuda, T.; Maruyama, M.; Okuda, A.; Amemiya, T.; et al. miR-125b inhibits osteoblastic differentiation by down-regulation of cell proliferation. Biochem. Biophys. Res. Commun. 2008, 368, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhang, P.; Liang, P.; Huang, X. The expression of miR-125b regulates angiogenesis during the recovery of heat-denatured HUVECs. Burns 2015, 41, 803–811. [Google Scholar] [CrossRef]
- Huang, K.; Fu, J.; Zhou, W.; Li, W.; Dong, S.; Yu, S.; Hu, Z.; Wang, H.; Xie, Z. MicroRNA-125b regulates osteogenic differentiation of mesenchymal stem cells by targeting Cbfb in vitro. Biochimie 2014, 102, 47–55. [Google Scholar] [CrossRef]
- Xihong, L.U.; Min, D.; Honghui, H.E.; Dehui, Z.; Wei, Z. miR-125b regulates osteogenic differentiation of human bone marrow mesenchymal stem cells by targeting Smad4. J. Cent. South. Univ. (Med. Sci.) 2013, 38, 341–346. [Google Scholar] [CrossRef]
- Muramatsu, F.; Kidoya, H.; Naito, H.; Sakimoto, S.; Takakura, N. microRNA-125b inhibits tube formation of blood vessels through translational suppression of VE-cadherin. Oncogene 2013, 32, 414–421. [Google Scholar] [CrossRef]
- Schaap-Oziemlak, A.M.; Raymakers, R.A.; Bergevoet, S.M.; Gilissen, C.; Jansen, B.J.H.; Adema, G.J.; Kögler, G.; le Sage, C.; Agami, R.; van der Reijden, B.A.; et al. MicroRNA hsa-miR-135b Regulates Mineralization in Osteogenic Differentiation of Human Unrestricted Somatic Stem Cells. Stem Cells Dev. 2010, 19, 877–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umezu, T.; Tadokoro, H.; Azuma, K.; Yoshizawa, S.; Ohyashiki, K.; Ohyashiki, J.H. Exosomal miR-135b shed from hypoxic multiple myeloma cells enhances angiogenesis by targeting factor-inhibiting HIF-1. Blood 2014, 124, 3748–3757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Santini, G.C.; De Veirman, K. ; Broek, I.V.; Leleu, X.; De, A.; Van Camp, B.; Vanderkerken, K.; Van Riet, I. Upregulation of miR-135b Is Involved in the Impaired Osteogenic Differentiation of Mesenchymal Stem Cells Derived from Multiple Myeloma Patients. PLoS ONE 2013, 8, e79752. [Google Scholar] [CrossRef]
- Sumiyoshi, K.; Kubota, S.; Ohgawara, T.; Kawata, K.; Abd El Kader, T.; Nishida, T.; Ikeda, N.; Shimo, T.; Yamashiro, T.; Takigawa, M. Novel Role of miR-181a in Cartilage Metabolism. J. Cell. Biochem. 2013, 114, 2094–2100. [Google Scholar] [CrossRef] [PubMed]
- Gabler, J.; Ruetze, M.; Kynast, K.L.; Grossner, T.; Diederichs, S.; Richter, W. Stage-Specific miRs in Chondrocyte Maturation: Differentiation-Dependent and Hypertrophy-Related miR Clusters and the miR-181 Family. Tissue Eng. Part. A 2015, 21, 2840–2851. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Charbonneau, C.; Wei, L.; Chen, Q.; Terek, R.M. miR-181a Targets RGS16 to Promote Chondrosarcoma Growth, Angiogenesis, and Metastasis. Mol. Cancer Res. 2015, 13, 1347–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Wei, L.; Chen, Q.; Terek, R.M. MicroRNA Regulates Vascular Endothelial Growth Factor Expression in Chondrosarcoma Cells. Clin. Orthop. Relat. Res. 2015, 473, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.I.; Silva, A.M.; Vasconcelos, D.M.; Almeida, C.R.; Caires, H.; Pinto, M.T.; Calin, A.; Santos, S.G.; Barbosa, M.A. miR-195 in human primary mesenchymal stromal/stem cells regulates proliferation, osteogenesis and paracrine effect on angiogenesis. Oncotarget 2015, 7, 7–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Zhao, N.; Li, S.; Fang, J.; Chen, M.; Yang, J.; Jia, W.; Yuan, Y.; Zhuang, S. MicroRNA-195 Suppresses Angiogenesis and Metastasis of Hepatocellular Carcinoma by Inhibiting the Expression of VEGF, VAV2, and CDC42. Hepatology 2013, 58, 642–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zeng, C.; Tu, M.; Jiang, W.; Dai, Z.; Hu, Y.; Deng, Z.; Xiao, W. MicroRNA-200b acts as a tumor suppressor in osteosarcoma via targeting ZEB1. Onco Targets Ther. 2016, 9, 3101–3111. [Google Scholar]
- Fan, X.; Teng, Y.; Ye, Z.; Zhou, Y.; Tan, W.-S. The effect of gap junction-mediated transfer of miR-200b on osteogenesis and angiogenesis in a co-culture of MSCs and HUVECs. J. Cell Sci. 2018, 131, jcs216135. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Yoon, S.; Jeong, Y.; Yoon, J.; Baek, K. Regulation of Vascular Endothelial Growth Factor Signaling by miR-200b. Mol. Cells 2011, 32, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, Y.; Tokuzawa, Y.; Ninomiya, Y.; Yagi, K.; Yatsuka-Kanesaki, Y.; Suda, T.; Fukuda, T.; Katagiri, T.; Kondoh, Y.; Amemiya, T.; et al. miR-210 promotes osteoblastic differentiation through inhibition of AcvR1b. FEBS Lett. 2009, 583, 2263–2268. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-D.; Cai, F.; Liu, L.; Zhang, Y.; Yang, A.-L. microRNA-210 is involved in the regulation of postmenopausal osteoporosis through promotion of VEGF expression and osteoblast differentiation. Biol. Chem. 2015, 396, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Fasanaro, P.; D’Alessandra, Y.; Di Stefano, V.; Melchionna, R.; Romani, S.; Pompilio, G.; Capogrossi, M.C.; Martelli, F. MicroRNA-210 modulates endothelial cell response to hypoxia and inhibits the receptor tyrosine kinase ligand ephrin-A3. J. Biol. Chem. 2008, 283, 15878–15883. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Guo, D.; Yang, S.; Sun, H.; Wu, B.; Zhou, D. Inhibition of miR-222-3p activity promoted osteogenic differentiation of hBMSCs by regulating Smad5-RUNX2 signal axis. Biochem. Biophys. Res. Commun. 2016, 470, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Takigawa, S.; Chen, A.; Wan, Q.; Na, S.; Sudo, A.; Yokota, H.; Hamamura, K. Role of miR-222-3p in c-Src-Mediated Regulation of Osteoclastogenesis. Int. J. Mol. Sci. 2016, 17, 240. [Google Scholar] [CrossRef] [PubMed]
- Poliseno, L.; Tuccoli, A.; Mariani, L.; Evangelista, M.; Citti, L.; Woods, K.; Mercatanti, A.; Hammond, S.; Rainaldi, G. MicroRNAs modulate the angiogenic properties of HUVECs. Blood 2006, 108, 3068–3071. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Yang, T.; Han, J.; Yan, K.; Qiu, X.; Zhou, Y.; Fan, Q.; Ma, B. MicroRNA Expression During Osteogenic Differentiation of Human Multipotent Mesenchymal Stromal Cells From Bone Marrow. J. Cell. Biochem. 2011, 112, 1844–1856. [Google Scholar] [CrossRef]
- Vimalraj, S.; Selvamurugan, N. MicroRNAs expression and their regulatory networks during mesenchymal stem cells differentiation toward osteoblasts. Int. J. Biol. Macromol. 2014, 66, 194–202. [Google Scholar] [CrossRef]
- Li, L.; Qi, Q.; Luo, J.; Huang, S.; Ling, Z.; Gao, M.; Zhou, Z.; Stiehler, M.; Zou, X. FOXO1-suppressed miR-424 regulates the proliferation and osteogenic differentiation of MSCs by targeting FGF2 under oxidative stress. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- de Pontual, L.; Yao, E.; Callier, P.; Faivre, L.; Drouin, V.; Cariou, S.; Van Haeringen, A.; Geneviève, D.; Goldenberg, A.; Oufadem, M.; Manouvrier, S.; Munnich, A.; et al. Germline deletion of the miR-17 ~ 92 cluster causes skeletal and growth defects in humans. Nat. Genet. 2011, 43, 1026–1030. [Google Scholar] [CrossRef] [PubMed]
- Penzkofer, D.; Bonauer, A.; Fischer, A.; Tups, A.; Brandes, R.P.; Zeiher, A.M.; Dimmeler, S. Phenotypic Characterization of miR-92a - /- Mice Reveals an Important Function of miR-92a in Skeletal Development. PLoS ONE 2014, 9, e101153. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Ma, J.; Chen, S.; Chen, X.; Yu, X. MicroRNA-17-92 cluster regulates osteoblast proliferation and differentiation. Endocrine 2014, 45, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Wu, P.; Zhang, Z.; Zhang, Z.; Liao, W.; Li, Y.; Kang, Y. MicroRNA-92a-3p Regulates Aggrecanase-1 and Aggrecanase-2 Expression in Chondrogenesis and IL-1β- Induced Catabolism in Human Articular Chondrocytes. Cell. Physiol. Biochem. 2017, 44, 38–52. [Google Scholar] [CrossRef]
- Zhang, Z.; Kang, Y.; Zhang, Z.; Zhang, H.; Duan, X.; Liu, J.; Li, X.; Liao, W. Expression of microRNAs during chondrogenesis of human adipose-derived stem cells. Osteoarthr. Cartil. 2012, 20, 1638–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, G.; Zhang, Z.; Huang, Z.; Chen, W.; Huang, G.; Meng, F.; Zhang, Z.; Kang, Y. MicroRNA-92a-3p regulates the expression of cartilage-specific genes by directly targeting histone deacetylase 2 in chondrogenesis and degradation. Osteoarthr. Cartil. 2017, 25, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Bonauer, A.; Carmona, G.; Iwasaki, M.; Mione, M.; Koyanagi, M.; Fischer, A.; Burchfield, J.; Fox, H.; Doebele, C.; Ohtani, K.; et al. MicroRNA-92a Controls Angiogenesis and Functional Recovery of Ischemic Tissues in Mice. Science 2009, 324, 1710–1713. [Google Scholar] [CrossRef] [PubMed]
- Elmén, J.; Lindow, M.; Schütz, S.; Lawrence, M.; Petri, A.; Obad, S.; Lindholm, M.; Hedtjärn, M.; Hansen, H.; Berger, U.; et al. LNA-mediated microRNA silencing in non-human primates. Nature 2008, 452, 896–899. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, M.; Qin, G.; Weintraub, N.L.; Tang, Y. MiR-92a regulates viability and angiogenesis of endothelial cells under oxidative stress. Biochem. Biophys. Res. Commun. 2015, 446, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Rippe, C.; Blimline, M.; Magerko, K.A.; Lawson, B.R.; LaRocca, T.; Donato, A.; Seals, D.R. MicroRNA Changes in Human Arterial Endothelial Cells with Senescence: Relation to Apoptosis, eNOS and Inflammation Catarina. Exp. Gerontol. 2012, 47, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kim, H.K.; Chung, S.; Kim, K.S.; Dutta, A. Depletion of human micro-RNA miR-125b reveals that it is critical for the proliferation of differentiated cells but not for the down-regulation of putative targets during differentiation. J. Biol. Chem. 2005, 280, 16635–16641. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.-F.; Yang, G.-H.; Pan, X.-H.; Zhang, S.-J.; Zhao, C.; Qiu, B.-S.; Gu, H.-F.; Hong, J.-F.; Cao, L.; Chen, Y.; et al. Altered MicroRNA Expression Profile in Exosomes during Osteogenic Differentiation of Human Bone Marrow- Derived Mesenchymal Stem Cells. PLoS ONE 2014, 9, e114627. [Google Scholar] [CrossRef] [PubMed]
- Fan, G. Hypoxic exosomes promote angiogenesis Platelets: Balancing the septic triad. Blood 2014, 124, 3669–3670. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Sun, Z.-J.; Bian, Y.; Kulkarni, A.B. MicroRNA-135b acts as a tumor promoter by targeting the hypoxia-inducible factor pathway in genetically defined mouse model of head and neck squamous cell carcinoma. Cancer Lett. 2013, 331, 230–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazenwadel, J.; Michael, M.Z.; Harvey, N.L. Prox1 expression is negatively regulated by miR-181 in endothelial cells. Blood 2010, 116, 2395–2401. [Google Scholar] [CrossRef] [Green Version]
- Naguibneva, I.; Ameyar-Zazoua, M.; Polesskaya, A.; Ait-Si-Ali, S.; Groisman, R.; Souidi, M.; Cuvellier, S.; Harel-Bellan, A. The microRNA miR-181 targets the homeobox protein Hox-A11 during mammalian myoblast differentiation. Nat. Cell Biol. 2006, 8, 278–284. [Google Scholar] [CrossRef]
- Akiyama, T.; Bouillet, P.; Miyazaki, T.; Kadono, Y.; Chikuda, H.; Chung, U.; Fukuda, A.; Hikita, A.; Seto, H.; Okada, T.; et al. Regulation of osteoclast apoptosis by ubiquitination of proapoptotic BH3-only Bcl-2 family member Bim. EMBO J. 2003, 22, 6653–6664. [Google Scholar] [CrossRef]
- Palmieri, A.; Pezzetti, F.; Brunelli, G.; Zollino, I.; Scapoli, L.; Martinelli, M.; Arlotti, M.; Carinci, F. Differences in osteoblast miRNA induced by cell binding domain of collagen and silicate-based synthetic bone. J. Biomed. Sci. 2007, 14, 777–782. [Google Scholar] [CrossRef]
- Christoffersen, N.R.; Silahtaroglu, A.; Ørom, U.L.F.A.; Kauppinen, S.; Lund, A.H. miR-200b mediates post-transcriptional repression of ZFHX1B. RNA 2007, 13, 1172–1178. [Google Scholar] [CrossRef]
- Baglio, S.R.; Rooijers, K.; Koppers-lalic, D.; Verweij, F.J.; Lanzón, M.P.; Zini, N.; Naaijkens, B.; Perut, F.; Niessen, H.W.M.; Baldini, N.; et al. Human bone marrow- and adipose- mesenchymal stem cells secrete exosomes enriched in distinctive miRNA and tRNA species. Stem Cell Res. Ther. 2015, 6, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Zong, L.; Zhu, Y.; Liang, R.; Zhao, H.-B. Gap junction mediated miRNA intercellular transfer and gene regulation: A novel mechanism for intercellular genetic communication. Sci. Rep. 2016, 6, 19884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.-T.; Chen, H.-T.; Tsou, H.-K.; Tan, T.-W.; Fong, Y.-C.; Chen, P.-C.; Yang, W.-H.; Wang, S.-W.; Chen, J.-C.; Tang, C.-H. CCL5 promotes VEGF-dependent angiogenesis by downregulating miR-200b through PI3K/Akt signaling pathway in human chondrosarcoma cells. Oncotarget 2014, 5, 10718–10731. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.C.; Khanna, S.; Roy, S.; Sen, C.K. miR-200b targets Ets-1 and is down-regulated by hypoxia to induce angiogenic response of endothelial cells. J. Biol. Chem. 2011, 286, 2047–2056. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.-L.; Guo, F.; Liu, F.; Gao, F.-L.; Zhang, P.-Q.; Niu, X.; Guo, S.-C.; Yin, J.-H.; Wang, Y.; Deng, Z.-F. miR-210 activates notch signaling pathway in angiogenesis induced by cerebral ischemia. Mol. Cell Biochem. 2012, 370, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Fasanaro, P.; Greco, S.; Lorenzi, M.; Pescatori, M.; Brioschi, M.; Kulshreshta, R.; Banfi, C.; Stubbs, A.; Calin, G.A.; Ivan, M.; et al. An integrated approach for experimental target identification of hypoxia-induced miR-210. J. Biol. Chem. 2009, 284, 35134–35143. [Google Scholar] [CrossRef] [PubMed]
- Ivan, M.; Huang, X. miR-210: Fine-Tuning the Hypoxic Response. Adv. Exp. Med. Biol. 2014, 772, 205–227. [Google Scholar] [CrossRef]
- Ivan, M.; Harris, A.L.; Martelli, F.; Kulshreshtha, R. Hypoxia response and microRNAs: No longer two separate worlds. J. Cell Mol. Med. 2008, 12, 1426–1431. [Google Scholar] [CrossRef]
- Kuijper, S.; Turner, C.J.; Adams, R.H. Regulation of angiogenesis by Eph-ephrin interactions. Trends Cardiovasc. Med. 2007, 17, 145–151. [Google Scholar] [CrossRef]
- Pandey, A.; Shao, H.; Marks, R.M.; Polverini, P.J.; Dixit, V.M. Role of B61, the ligand for the Eck receptor tyrosine kinase, in TNF-alpha-induced angiogenesis. Science 1995, 268, 567–569. [Google Scholar] [CrossRef]
- Matsui, J.; Wakabayashi, T.; Asada, M.; Yoshimatsu, K. Stem cell factor/c-kit signaling promotes the survival, migration, and capillary tube formation of human umbilical vein endothelial cells. J. Biol. Chem. 2004, 279, 18600–18607. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Kleinheinz, J. Angiogenesis—The Key to Regeneration. In Tissue Engineering and Regenerative Medicine; Andrades, J.A., Ed.; InTechOpen: London, UK, 2013; pp. 453–473. [Google Scholar]
- Kanczler, J.M.; Oreffo, R.O.C. Osteogenesis and angiogenesis: The potential for engineering bone. Eur. Cells Mater. 2008, 15, 100–114. [Google Scholar] [CrossRef]
- Hou, H.; Zhang, X.; Tang, T.; Dai, K.; Ge, R. Enhancement of bone formation by genetically-engineered bone marrow stromal cells expressing BMP-2, VEGF and angiopoietin-1. Biotechnol. Lett. 2009, 31, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Qiu, T.; Wu, X.; Wan, C.; Shi, W.; Wang, Y.; Chen, J.; Wan, M.; Clemens, T.L.; Cao, X. Sustained BMP Signaling in Osteoblasts Stimulates Bone Formation by Promoting Angiogenesis and Osteoblast Differentiation. J. Bone Miner. Res. 2009, 24, 1224–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z.; Wang, K. Effects of recombinant adeno-associated viral vectors on angiopoiesis and osteogenesis in cultured rabbit bone marrow stem cells via co-expressing hVEGF and hBMP genes: A preliminary study in vitro. Tissue Cell 2010, 42, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Hon, L.S.; Zhang, Z. The roles of binding site arrangement and combinatorial targeting in microRNA repression of gene expression. Genome Biol. 2007, 8, R166. [Google Scholar] [CrossRef]
- Grimson, A.; Farh, K.K.; Johnston, W.K.; Garrett-Engele, P.; Lim, L.P.; Bartel, D.P. MicroRNA targeting specificity in mammals: Determinants beyond seed pairing. Mol. Cell 2007, 27, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microRNA.org resource: Targets and expression. Nucleic Acids Res. 2008, 36, D149–D153. [Google Scholar] [CrossRef]
- Krek, A.; Grün, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; MacMenamin, P.; da Piedade, I.; Gunsalus, K.C.; Stoffel, M.; et al. Combinatorial microRNA target predictions. Nat. Genet. 2005, 37, 495–500. [Google Scholar] [CrossRef]
- Brennecke, J.; Stark, A.; Russell, R.; Cohen, S. Principles of microRNA-target recognition. PLoS Biol. 2005, 3, e85. [Google Scholar] [CrossRef]
- Tsang, J.; Zhu, J.; van Oudenaarden, A. MicroRNAmediated feedback and feedforward loops are recurrent network motifs in mammals. Mol. Cell 2007, 26, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.; Burge, C.; Bartel, D. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xie, H.; Liu, W.; Hu, R.; Huang, B.; Tan, Y.; Xu, K.; Sheng, Z.; Zhou, H.; Wu, X.; et al. A novel microRNA targeting HDAC5 regulates osteoblast differentiation in mice and contributes to primary osteoporosis in humans. J. Clin. Invest. 2009, 119, 3666–3677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariner, P.; Johannesen, E.; Anseth, K. Manipulation of miRNA activity accelerates osteogenic differentiation of hMSCs in engineered 3D scaffolds. J. Tissue Eng. Regen Med. 2012, 6, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.Y.; Li, N.; Lin, S.; Wang, B.; Lan, H.Y.; Li, G. miRNA-29b improves bone healing in mouse fracture model. Mol. Cell. Endocrinol. 2016, 430, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, L.; Liu, S.; Liu, W.; Zhang, H.; Zhou, T.; Wu, D.; Yang, P.; Shen, L.; Chen, J.; et al. The promotion of bone regeneration through positive regulation of angiogenic-osteogenic coupling using microRNA-26a. Biomaterials 2013, 34, 5048–5058. [Google Scholar] [CrossRef] [PubMed]
- Yoshizuka, M.; Nakasa, T.; Kawanishi, Y.; Hachisuka, S.; Furuta, T.; Miyaki, S.; Adachi, N.; Ochi, M. Inhibition of microRNA-222 expression accelerates bone healing with enhancement of osteogenesis, chondrogenesis, and angiogenesis in a rat refractory fracture model. J. Orthop. Sci. 2016, 21, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Ørom, U.A.; Kauppinen, S.; Lund, A.H. LNA-modified oligonucleotides mediate specific inhibition of microRNA function. Gene 2006, 372, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Adams, B.D.; Parsons, C.; Walker, L.; Zhang, W.C.; Slack, F.J. Targeting noncoding RNAs in disease. J. Clin. Investig. 2017, 127, 761–771. [Google Scholar] [CrossRef] [Green Version]
- Simonson, B.; Das, S. MicroRNA Therapeutics: The Next Magic Bullet? Mini Rev. Med. Chem. 2016, 15, 467–474. [Google Scholar] [CrossRef]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs-microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef] [PubMed]
- López-Camarillo, C.; Marchat, L.A.; Aréchaga-Ocampo, E.; Azuara-Liceaga, E.; Pérez-Plasencia, C.; Fuentes-Mera, L.; Fonseca-Sánchez, M.A.; Flores-Pérez, A. Functional Roles of microRNAs in Cancer: microRNomes and oncomiRs Connection; Oncogenomi.; In Tech Open Science: London, UK, 2013. [Google Scholar]
- Senanayake, U.; Das, S.; Vesely, P.; Alzoughbi, W.; Fröhlich, L.F.; Chowdhury, P.; Leuschner, I.; Hoefler, G.; Guertl, B. miR-192, miR-194, miR-215, miR-200c and miR-141 are downregulated and their common target ACVR2B is strongly expressed in renal childhood neoplasms. Carcinogenesis 2012, 33, 1014–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
MicroRNAs (miRs/miRNAs) involved in the regulation and coupling of bone angiogenesis (“CouplingmiRs/CPLGmiRs”). Reported miRNAs contributing to the formation of blood vessels during the processes of formation, repair and regeneration of bone were allocated with the individual functions of their target genes during osteogenesis, angiogenesis, or hypoxic regulation of bone angiogenesis. OB, osteoblast; OC, osteoclast; CC, chondrocyte; EC, endothelial cell.
Figure 1.
MicroRNAs (miRs/miRNAs) involved in the regulation and coupling of bone angiogenesis (“CouplingmiRs/CPLGmiRs”). Reported miRNAs contributing to the formation of blood vessels during the processes of formation, repair and regeneration of bone were allocated with the individual functions of their target genes during osteogenesis, angiogenesis, or hypoxic regulation of bone angiogenesis. OB, osteoblast; OC, osteoclast; CC, chondrocyte; EC, endothelial cell.

Figure 2.
Regulating effects of microRNAs (miRs/miRNAs) involved in the regulation and coupling of bone angiogenesis (“CouplingmiRs”) during cell fate determination. MiRNAs were assigned with their individual positive/stimulatory (green colored) or negative/inhibitory (red colored) function and occurrence during the specific differentiation steps of osteogenic and angiogenic cells. Cell images from Servier Medical Art by Servier licensed under a Creative Commons Attribution 3.0.
Figure 2.
Regulating effects of microRNAs (miRs/miRNAs) involved in the regulation and coupling of bone angiogenesis (“CouplingmiRs”) during cell fate determination. MiRNAs were assigned with their individual positive/stimulatory (green colored) or negative/inhibitory (red colored) function and occurrence during the specific differentiation steps of osteogenic and angiogenic cells. Cell images from Servier Medical Art by Servier licensed under a Creative Commons Attribution 3.0.


{kind=link}
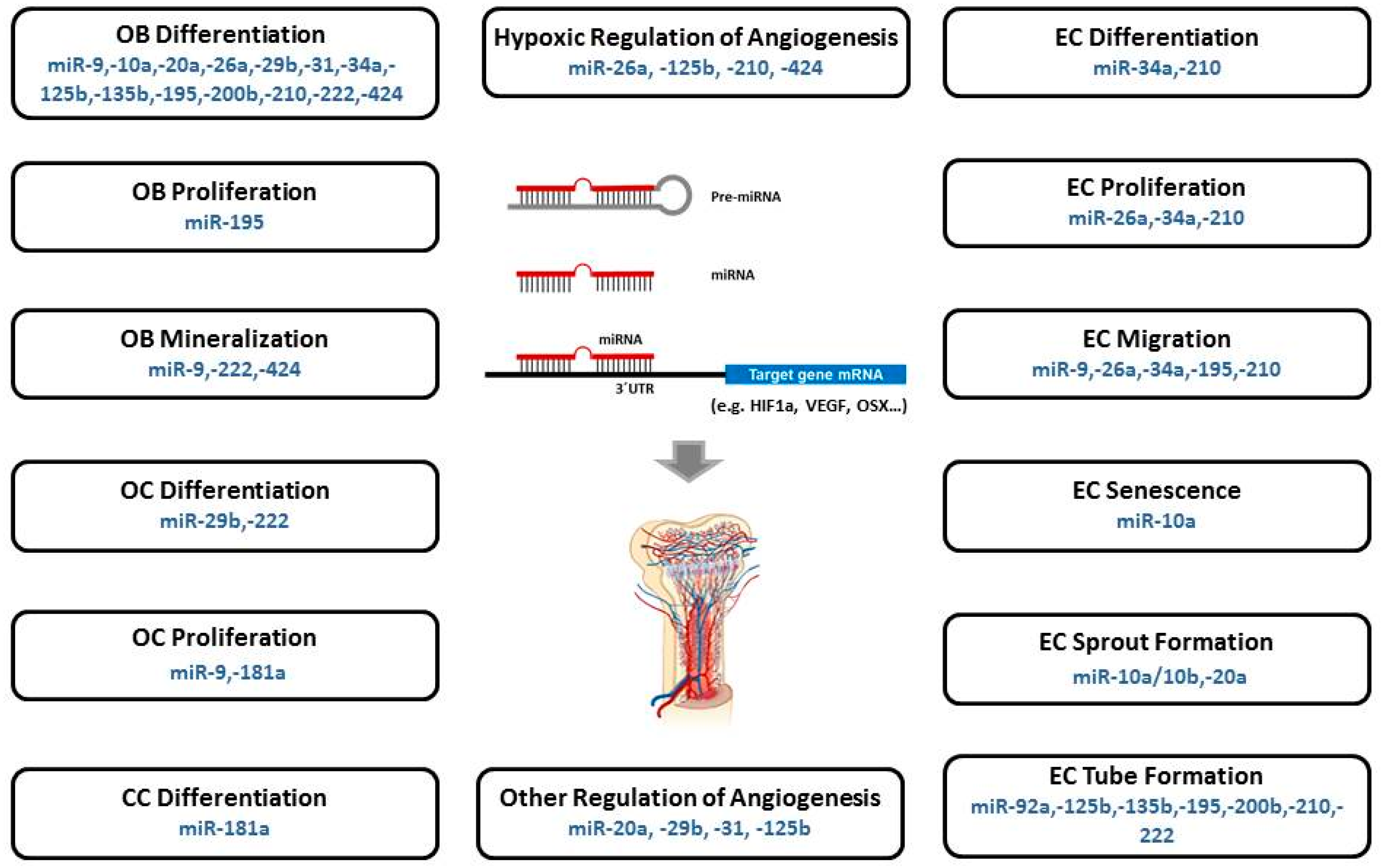{kind=link}
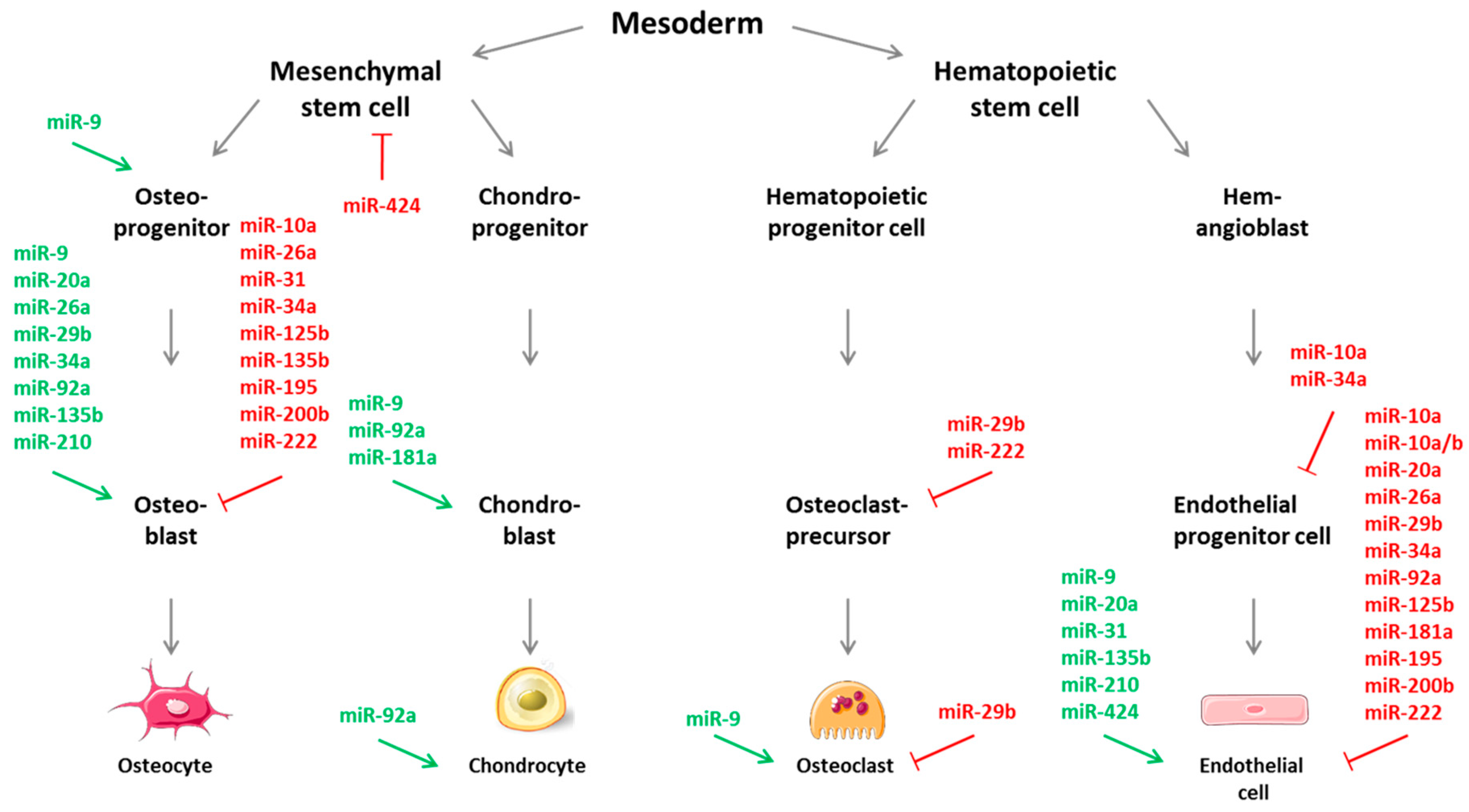{kind=link}
Table 1.
Summary of microRNAs with a presumed role in osteogenic-angiogenic coupling paving the way for bone angiogenesis (“CouplingmiRs/CPLGmiRs”).
Table 1.
Summary of microRNAs with a presumed role in osteogenic-angiogenic coupling paving the way for bone angiogenesis (“CouplingmiRs/CPLGmiRs”).
| MicroRNAs | Targets 1 | Regulatory Role | Effects | Study Models | Ref. |
|---|---|---|---|---|---|
| MiR-9 | VEGF, VE-CAD (CD144) | AMPK signaling pathway | Enhanced osteogenic diff. & mineral.; increased angiogenesis | MC3T3-E1 | [106] |
| DKK1 | COL1, OCN, BSP; ALP activity | OB diff. & mineralization | C2C12 cells | [107] | |
| SOCS5 | JAK-STAT signaling pathway | Promotion of EC migration & angiogenesis | Primary microvascular ECs, HUVECs | [108] | |
| Cbl | Bim ubiquitination, apoptosis | Promotion of OC survival | OC, OC precursor cells (RAW264.7) | [110] | |
| MiR-10a | β-catenin, LEF1; VEGF, VE-CAD (CD144), cyclin D1, MMP2 | Wnt signaling; angiogenesis-related gene expression | Inhibition of osteogenic diff. & blood vessel formation | MC3T3-E1 MUVECS | [111] |
| HMGA2 | β–galactosidase expr; p16Ink4a/p19Arf expression | EPC senescence & angiogenesis; self-renewal potential | lin−BM-MSCs | [113] | |
| MiR-10a/10b | MIB1 | Notch signaling | Regulating blood vessel outgrowth/tip cell behavior | HUVECs | [115] |
| MiR-20a | BMP2, BMP4, RUNX2 | Effects BMP/RUNX2 signaling positively; blocks OB inhibitors & PPARγ | Enhances osteogenic differentiation; suppresses adipogenesis | hBM-MSC | [116] |
| JAK1; p21, S1P receptor EDG | Downregulation of proangiogenic JAK 1 & cell cycle inhibitors | Inhibits EC sprout formation | HUVECs | [117] | |
| TNFSF15 | VEGF-AKT/ERK –miR20a/31 signaling | Stimulation of angiogenesis | HUVECs | [118] | |
| MiR-26a | VEGF, ANG1, RUNX2, BMP2 OCN, ALP; GSK3β | WNT signaling activation | Enhanced angiogenesis & bone regeneration | Primary hBM-MSC, MC3T3-E1 | [76,98] |
| VEGF | PIK3C2α/AKT/HIF-α/VEGFA pathway | Inhibition of angiogenesis; | HUVECs | [141] | |
| SMAD1 | BMP signaling inhibition | OB differentiation | hADSCs | [119] | |
| SMAD1 | BMP signaling | Inhibits EC growth, proliferation, migration; regulates early angiogenesis | HUVECs | [123] | |
| MiR-29b | TGF-β3, HDAC4, ACTVR2A, CTNNBIP1, DUSP2; COL1A1, 5A3, 4A2 | Silences neg. osteogenic regulators suppresses ECM protein synthesis | Promotes osteoblastogenesis at multiple stages | MC3T3 pre-OB | [124] |
| c-FOS | Reduced TRAP expr., lacunae generation, collagen degradation | Neg. regulator of human OC differentiation and activity | OC (CD14 +) | [125] | |
| TNFSF15 | TNFSF15-enhanced JNK-GATA3 signal. & VEGF inhibition | Suppression of VEGF secretion | Mouse EC line bEnd.3 | [126] | |
| AKT3 | Inhibition of tumor vascularization via VEGF & cancer cell activity via c-MYC | Anti-angiogenic and anti-tumorigenic role | HUVECs, Breast cancer cells | [127] | |
| MiR-31 | OSX | Downregulation of OSX | Influences osteogenic differentiation | hMSC; Osteosarcoma cell | [129] |
| Satb2 protein | Inhibition by RUNX2; Upregulation of Satb2 protein & osteogenic TF | Induces BM-MSC osteogenic differentiation | hBM-MSC | [130] | |
| E-selectin | Regulation of E-selectin expression | Inhibition of angiostatin-induced angiogenesis; TNF-mediated induction of endothelial adhesion | HUVECs | [84] | |
| TNFSF15 | VEGF-AKT/ERK –miR20a/31 signaling | Stimulation of angiogenesis | HUVECs | [118] | |
| MiR-34a | Jagged1 | Regulation of cell cycle regulator & proliferation proteins & Jagged1 | Inhibition of osteoblast differentiation | hMSC; mouse heterotopic bone formation model | [132] |
| JAGGED1 | Activation of Notch signaling | Induction of glucocorticoid-mediated osteogenic differentiation | hMSC | [133] | |
| RBP2 | Promotes mineral, ALP activity & RUNX2 expression; downreg. NOTCH1 & Cyclin D1 expr. | Promotion of osteogenic differentiation; enhanced heterotopic bone formation | hADSCs; mouse heterotopic bone formation model | [134] | |
| VEGF | Inhibitory effects of dexamethasone on EC viability & VEGF | Decreased blood vessel development | Rat Glucocorticoid- induced osteonecrosis | [137] | |
| SIRT1 | Increased SIRT1 expr. & FOXO1 acetylation regulating vascular EC homeostasis | Inhibition of EPC-mediated angiogenesis | Rat EPC | [138] | |
| E2F3a, survivin | Interference with VEGF secretion, EC proliferation & migration | Dysregulated tumor angiogenesis | HNSCC tumors & cells | [140] | |
| MiR-92a | ? | ? | Enhanced fracture healing & inhib. of neovascularization | Mice with femoral fracture | [142] |
| HGF, ANGPT1 | ITGA5, MEK4 | Inhibition of tube formation by HUVECs | hADSCs | [143] | |
| ? | integrin a5, sirtuin1, eNOS | Attenuates neointimal lesion by accelerating re-endothelialization | MiR-92a knockout mice | [144] | |
| MiR-125b | OSX | RUNX2, a-SMC, ALP, matrix mineralization | Calcification of vascular smooth muscle cells | HCASMCs | [145] |
| ErbB2 | ? | Inhibits OB diff by downreg. of cell proliferation | ST2 cells (mMSCs) | [146] | |
| VEGF, ERBB2 | Regulation of angiogenesis during wound healing | HUVECS | [147] | ||
| Cbf-beta | ALP, OCN, OPN | Inhibition of osteogenic differentiation | C3H10T1/2 | [148] | |
| SMAD4 | ALP, RUNX2 | Downregulation of osteogenic differentiation | hMSCs | [149] | |
| VE-Cadherin | Inhibition of blood vessel (tube) formation | HUVECs | [150] | ||
| MiR-135b | ? | ? | OB differentiation | hBM-SCs | [151] |
| HIF-1 | ? | Enhanced endothelial tube formation | Human MM cells; HUVECs | [152] | |
| SMAD5 | ? | Impaired osteogenic differentiation | hMSCs | [153] | |
| MiR-181a | ? | CCN1, aggrecan | Maintaining homeostasis of chondrocytes | Human HCS-2/8 cells | [154] |
| COL10A1 | Chondrocyte differentiation | hMSC | [155] | ||
| RGS16 | CXCR4 signaling; VEGF, MMP1 | Angiogenesis & metastasis in chondrosarcoma | Xenograft mice; JJ chondrosarc. cells | [156] | |
| ? | VEGF expression | Chondrosarcoma-associated angiogenesis | JJ chondrosarc. cell line | [157] | |
| Cbl | Bim ubiquitination, apoptosis | promote OC survival | OC, OC precursor cells (RAW264.7) | [110] | |
| MiR-195 | ? | VEGF | Osteogenic diff. & proliferation; control of angiogenesis | hMSC(MC3T3) chick chorio-allantoic membrane | [158] |
| ? | VEGF, VAV2 CDC42 | HCC-associated angiogenesis & metastasis; migration & capillary tube form. of ECs | QGY-7703, MHCC-97H HCC cells; HUVECs | [159] | |
| MiR-200b | ZEB1 | ZEB1-TF target genes | Inhibits proliferation, migration & invasion of osteosarcoma cells | OsteosarcomaU2OS, Saos2, HOS, MG63 | [160] |
| VEGF-A; ZEB2, ETS1, KDR,GATA2 | Decreases VEGF-A expression & TF-target genes | Inhibition of VEGF-A induced osteogenesis; Inhibition of TF-activated angiogenesis | Rat BM-MSC & HUVEC coculture | [161] | |
| VEGF, FLT-1, and KDR | VEGF-induced phosph. of ERK1/2 | Inhibition of angiogenesis; red. capillary formation | A549 cells, HUVECs | [162] | |
| MiR-210 | AcvR1b | Inhib. of TGFb/activin signaling | Promotes OB differentiation | ST2 stromal cells | [163] |
| VEGF | PPARgamma, ALP, OSX | Promoteion of OB diff., inhibition of adipocyte diff. | hBM-SCs, 17β-estradiol (E2)treated OB | [164] | |
| EFNA3 | VEGF-expression mediated angiogenesis | EC survival, diff., migration; stim. of tubulogen. & chemotaxis | HUVECs | [165] | |
| MiR-222 | SMAD 1, 5, 8 protein & phosphoryl. | Decreased SMAD5-RUNX2 signaling & OSX, ALP, and OC levels & mineral. | Neg. regulator of osteogenic differentiation | hBM-SC | [166] |
| c-Src, Dcstamp | RANKL-induced expression of TRAP & cathepsin K | Inhibitory regulator of c-Src-mediated osteoclastogenesis | RAW264.7 pre-OC cells | [167] | |
| c-KIT | Suppression of tube formation, wound healing, cell migration via SCF | Inhibitory regulation of in vitro angiogenesis | HUVEC | [168] | |
| MiR-424 | RUNX, CBFβ, BMP | Osteogenic diff. of hMSCs | Bone formation | hMSCs | [169] |
| MAPK, WNT & insulin signal. | OB differentiation of hMSCs | Bone formation | hMSCs | [170] | |
| FGF-2; via FOXO1 | Decrease of ALP, mineralization & osteog. markers | Enhances proliferation & osteogenic differentiation of hMSCs | Pigs, cellular oxidative stress model | [171] | |
| CUL2; via RUNX-1→ C/EBPα→ PU.1 | Stabilization of HIF-1α | Regulation of Angiogenesis | ECs, ischemic tissues | [101] |
1 Identified target genes or downstream effectors; HUVECs, human umbilical vein endothelial cells; ?, unknown molecular target(s) and/or regulatory role; MUVECs, mouse umbilical vein endothelial cells; EC, endothelial cells; EPC, endothelial progenitor cells; hMSCs, human mesenchymal stem cells; hBM-SC, bone-marrow derived stem cells; hADSCs, human adipose-derived mesenchymal stem cells; TNFSF15; cytokine tumor necrosis factor superfamily 15; OB, osteoblasts; OC, osteoclasts; TF, transcription factor.
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fröhlich, L.F. MicroRNAs at the Interface between Osteogenesis and Angiogenesis as Targets for Bone Regeneration. Cells 2019, 8, 121. https://doi.org/10.3390/cells8020121
AMA Style
Fröhlich LF. MicroRNAs at the Interface between Osteogenesis and Angiogenesis as Targets for Bone Regeneration. Cells. 2019; 8(2):121. https://doi.org/10.3390/cells8020121
Chicago/Turabian StyleFröhlich, Leopold F. 2019. "MicroRNAs at the Interface between Osteogenesis and Angiogenesis as Targets for Bone Regeneration" Cells 8, no. 2: 121. https://doi.org/10.3390/cells8020121
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.