Nuclear Hormone Receptors and Their Ligands: Metabolites in Control of Transcription
by
Lian Jing Tao
1, 
Dong Eun Seo
1,
Benjamin Jackson
1,
Natalia B. Ivanova
1,* and
Fabio Rinaldo Santori
2,* 1
Department of Genetics, Center for Molecular Medicine, University of Georgia, Athens, GA 30602, USA
2
Department of Immunobiology, Yale University, New Haven, CT 06520, USA
*
Authors to whom correspondence should be addressed.
Cells 2020, 9(12), 2606; https://doi.org/10.3390/cells9122606
Submission received: 17 November 2020
/
Revised: 30 November 2020
/
Accepted: 1 December 2020
/
Published: 4 December 2020
(This article belongs to the Collection Functions of Nuclear Receptors)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Nuclear hormone receptors are a family of transcription factors regulated by small molecules derived from the endogenous metabolism or diet. There are forty-eight nuclear hormone receptors in the human genome, twenty of which are still orphans. In this review, we make a brief historical journey from the first observations by Berthold in 1849 to the era of orphan receptors that began with the sequencing of the Caenorhabditis elegans genome in 1998. We discuss the evolution of nuclear hormone receptors and the putative ancestral ligands as well as how the ligand universe has expanded over time. This leads us to define four classes of metabolites—fatty acids, terpenoids, porphyrins and amino acid derivatives—that generate all known ligands for nuclear hormone receptors. We conclude by discussing the ongoing efforts to identify new classes of ligands for orphan receptors.
Keywords:
nuclear hormone receptors; ligands; terpenoid; fatty acid; thyroxine; porphyrins; history; evolution1. Introduction
As a family, nuclear hormone receptors (NHRs) represent some of the most biologically important transcription factors that integrate cellular metabolism and function. NHR activities are controlled by binding to small molecules or ligands derived from endogenous metabolism, hormones or vitamins obtained from the diet [1]. A typical NHR contains a DNA binding domain, which recognizes a specific DNA motif, and a ligand binding domain, which regulates the NHR activity. In the absence of the ligand, the ligand binding domain may adopt either an inactive or repressive conformation [2]. Upon ligand binding, the resulting conformational changes allow recruitment of co-activators and induction of target gene expression on promoters containing a positive hormone receptor element or target gene repression on promoters containing a negative hormone receptor element [3]. The ability to be regulated by small molecules makes these receptors ideal targets for drug discovery: 16% of all drugs target NHRs [4].
Metabolites are widely used for intercellular communication in both prokaryotes and eukaryotes [5]. For example, fatty acids are dedicated to “quorum sensing” in bacteria, whereas peptides, cyclic AMP, and lysophosphatidic acid mediate a variety of responses in eukaryotes [5]. Two common metabolite signaling systems, G-protein coupled receptors (GPCRs) and cAMP receptors, are abundant throughout the animal and plant kingdoms [5]. In contrast, NHRs are observed exclusively in animals and were first detected in sponges [6].
Identification of ligands for NHRs is fundamental for understanding how these transcription factors function. In this review, we focus on metabolic pathways that produce ligands for NHRs and the mechanisms by which NHR ligands mediate intercellular communication.
2. A Brief History of NHRs and Their Ligands
2.1. Phenomenology (1849–1914)
It has been known since antiquity that eunuchs are passive, non-aggressive and sexually uninterested in females. The mechanisms underlying this phenotype were not well understood until 1849, when Berthold conducted an experiment that marked the beginning of endocrinology. He removed the testicles of roosters and observed that these animals, like eunuchs, became passive and non-aggressive. The roosters also had underdeveloped vocalization, combs and neck lobes [7]. Most importantly, when Berthold transplanted testicles back to the castrated animals, secondary sexual characteristics were restored, even though the transplanted testicles were in ectopic sites and had their normal innervation removed [7]. Forty years later, Brown-Sequard demonstrated that injection of testicular extract could increase aggression and stamina in male animals, including man [8]. It was at that time proposed that testicular extract could be used as rejuvenation therapy [8]. Brown-Sequard’s experiments clearly suggested that it was a substance in the testicles that could promote this effect, and there was no need for the organ or cells themselves. Less dramatic, but equally significant, were related observations on the thyroid gland. Physicians had observed that surgical removal of the thyroid gland resulted in myxedema and that injection of thyroid extract was sufficient to restore thyroid function of myxedema patients [9]. In 1896, Bauman demonstrated that a peptidic compound rich in iodine, “thyroiodin”, could be isolated from the thyroid extract and used to treat myxedema in animals [10]. Thus, whole organs could be replaced by extracts, suggesting that a messenger substance mediates the function of these glands.
To unify these observations, Starling created the concept of “hormones” that are released by one organ but affect other distant organs. The term “hormone” was derived from the Greek for “I excite” or “arouse” [11]. Starling extended the list of endocrine organs suspected of producing hormonal substances to include the adrenal glands, pancreas and ovary, in addition to the thyroid and the testes [11,12]. The best examples included the role of the supra-renal gland in the control of blood pressure [11], the relationship of the pancreas and diabetes [11], the role of the thyroid in the development of the nervous system and metabolism [12], the control of secondary sexual characteristics by the testes and ovaries [12] and the influence of ovaries on pregnancy [12]. Thus, the concept of hormones brought multiple phenomenological observations into the sphere of one concept that could be tested experimentally.
2.2. Hunting for Hormones (1915–1984)
Immediately, the hunt for hormones began. Starling had reinforced previous observations that the factor present in the thyroid was stable, since the function of the thyroid could be replenished by providing thyroid extract in the diet [12]. Taking this lead, Kendal isolated the active component of the thyroid gland, thyroxine [13]. Bioactive thyroxine that could replace “natural thyroid extract” was synthesized in 1927 by Harington and Barger [14]. Kendal’s discovery of thyroxine in 1915 marks the second phase in the history of NHRs: the era of orphan ligands. After this discovery, a quest for the identification, characterization and synthesis of hormones resulted in the discovery and synthesis of many hormones including estrogens [15,16], androgens [17,18,19], progesterone [20] and corticoids [21,22,23]. The isolation and purification of hormones was a heroic phase in biochemical research. For example, the isolation of the first androgen required the purification of 15 mg of pure steroid from an estimated 25,000 L of human male urine [24].
2.3. Ligands Meet Receptors (1985–1997)
NHR ligands remained orphans until the advent of recombinant DNA technology. The first NHR to be identified was the glucocorticoid receptor (GCR). Here, the ligand, glucocorticoid, was used to purify the receptor [25,26] which was then used to generate GCR-specific monoclonal antibodies to screen bacterial protein expression libraries to identify a GCR cDNA [27]. Identification and sequencing of GCR enabled the discovery of the retinoic acid receptor (RAR) and other members of the NHR family via homology-based cDNA library screening [28,29]. Thus, many orphan ligands were quickly “adopted” by their receptors [1]. The history of NHRs is summarized in Figure 1.
2.4. Orphan NHRs Meet Their Ligands (1998–Present)
Genome sequencing has provided us with a finite number of NHRs (Figure 2). Humans, for example, have 48 NHRs, only 12 of which are classic hormone and vitamin receptors [1]. Other organisms, like C. elegans, have >200 NHRs in their genome [30]. The majority of NHRs are orphan receptors. Over the years, many orphan NHRs have been deorphanized. Examples include the liver X receptors (LXRs) LXRα and LXRβ, which bind oxysterols [31]; farnesoid X receptor (FXR), a receptor for bile acids [32,33]; the retinoic acid-related orphan receptors (RORs) RORα and RORγ that bind cholesterol biosynthetic intermediates [34]. These success stories also apply to orphan nuclear hormone receptors from other species such as Daf12 of C. elegans, which binds dafachronic acids [35]. Identification of ligands for NHRs in nematodes and insects could be of importance for the treatment of diseases caused by parasitic nematodes in man [36] and pest control in agriculture [37]. However, despite these early discoveries, almost half of human NHRs and most NHRs from other species are still orphans.
3. Metabolome as a Source of NHR Ligands
The mammalian metabolome is composed of endogenous and exogenous metabolites. The endogenous metabolome contains all metabolic products produced by the organism itself. For example, cholesterol is an endogenous metabolite synthesized by mammalian cells from the two-carbon acetate group of acetyl-CoA [41]. The endogenous metabolome in mammals is estimated to contain approximately 1500 backbones/compounds, which are further modified to generate an enormous diversity of isomers [42]. The exogenous metabolome contains diet-derived as well as synthetic compounds that are modified by the enzymes of the organism. All degraded drugs are part of the exogenous metabolome, which is estimated to be at least ten to one hundred times larger than its endogenous counterpart [42].
NHRs are found in the most primitive animals, such as the demosponge Amphimedon queenslandica [43]. Sponges have two NHR-like proteins: AqNR1 and AqNR2. AqNR2 is the ortholog of the mammalian HNF4a/NR2A family [6], while AqNR1 is the ancestral receptor for all other NHRs. HNF4a/NR2A1 binds fatty acyl-CoA [44], and indeed fatty acids induce AqNR1 and AqNR2 transactivation [6], suggesting that one of the first ligands of ancestral NHRs was a fatty acid derivative. After the sponges, there was an expansion of the NHR superfamily, and the common ancestor that gave origin to vertebrates had receptors representative of the NHR families NR2C, NR5A1 (SF-1), NR6A1(GCNF), RXRs, ERRs and steroid receptors [6]. Here, we already see a branching of potential ligands: NR5A1 (SF-1) binds phosphatidylinositol [45], a fatty acid conjugated to inositol; RXRs bind 9-cis-retinoic acid [46,47], a retinoid; the steroid hormone receptors and estrogen-related receptors (ERRs) bind steroid hormones (reviewed in [1]). The ligands now include modified fatty acids and terpenes (Figure 2 and Figure 3). A second expansion of ligands recognized by NHRs occurred in the common ancestor of the bilaterians, before the branching of the protostomes and deuterostomes [6]. We now see the inclusion of additional ligand families represented by the thyroid hormone receptor that binds thyroxine [48,49], and REV-ERBα (NR1D1) and REV-ERBβ (NR1D2), which bind heme, a porphyrin [50]. All these ligands are derived from four main classes of metabolites: fatty acids, terpenoids, porphyrins and modified amino acids (Figure 2 and Figure 3).
A common thread between these ligands is the presence of a hydrophobic backbone with a head group that is either charged or capable of forming van der Walls or hydrogen bonds with the receptor. After the bilaterians, the expansion of the NHR superfamily was not followed by an expansion of recognized ligand classes. Rather, receptors recognize other derivatives of the four main classes of metabolites described above. For example, PPARγ binds to modified fatty acids, hydroxylated polyunsaturated fatty acids and prostaglandins [51,52,53], whereas FXR binds bile acids [32,33], which are modified terpenes.
3.1. Fatty Acid Family Ligands
Fatty acids are the ancestral family of ligands for NHRs, and they include components derived from the endogenous metabolism as well as diet-derived essential fatty acids such as linoleic, linolenic and arachidonic acids. Fatty acids are a class of diverse ligands. According to the lipidmaps database [54], there are currently 9985 fatty acids. Fatty acids can be modified at the hydrophobic tail (saturation, desaturation) and/or head group (glycerol, choline, ethanolamine, amino acid or carbohydrates). This greatly increases the diversity of this class of ligands. So far, there are 22,471 known glycerolipid, glycerophospholipid and sphingolipid derivatives of fatty acids (https://www.lipidmaps.org/resources/databases/index.php?tab=lmsd).
3.2. Terpenoid Family Ligands
Most ligands for nuclear hormone receptors come from the terpenoid family (Figure 2). Terpenoids are a class of natural compounds that includes over 80,000 known molecular species [48] and are easily modified by monooxygenation reactions catalyzed by CYP450 enzymes that increase diversity even further [55]. Most terpenoids are derivatives of 5-carbon isoprene units following what is known as Ruzicka’s rule [56]. In mammalian cells, terpenoids are synthesized from acetate [41] through the mevalonate [57] pathway. Terpenoids include all sterol lipids [41,56] and retinoids such as vitamin A and its precursor, β-carotene, which is generated in bacteria, fungi and plants [58]. Of particular interest are the sterol lipids. During evolution, there was an “explosion” of receptors that recognized sterol-type structures in the common ancestor of the bilaterians [6]. In humans, 12 NHRs with known ligands are activated by sterol lipids, including the receptors for estrogen [59,60], progesterone [61,62], testosterone [63,64], mineralocorticoids [65], glucocorticoids [27], oxysterols (LXRα and β) [31], bile acids (FXR) [32,33], cholesterol biosynthetic intermediates RORα and RORγ [34] and, for secosteroids, the vitamin D receptor (VDR) [66]. The diversity of sterol lipids is generated via modification of a basic backbone with four rings, A, B, C and D (Figure 3), by addition or removal of double bonds, hydroxyl and keto groups as well as different isomer patterns [67]. This diversity of modifications generates new compounds, distinct enough to be classified as subclasses of steroids. For example, in secosteroids, vitamin D and its related compounds are formed by the opening of the B ring in the sterol backbone [67]. Similarly, estrogens contain three double bonds in ring A [67]. Thus, receptors that initially recognized one sterol lipid could have been co-opted during evolution to act as a receptor for the new class of steroids, leading to the observed expansion in sterol lipid NHRs.
3.3. Porphyrins
The next class of compounds that generate ligands for NHRs is the porphyrins. These are lipophilic metabolites in which the porphyrin backbone is associated with a metal atom, such as Mg for chlorophyll or iron for heme. NHRs such as E75 in Drosophila melanogaster [68] or REV-ERBα (NR1D1) and REV-ERBαβ (NR1D2) in vertebrates bind heme [50,69]. In the case of E75, heme is a structural component of the receptors that is modified to serve as a sensor for diatomic gases like carbon monoxide and nitric oxide [68]. In contrast, in mammalian cells NR1D1 and NR1D2 function as direct heme sensors [50,69]. The finding of insect NHRs that sense gas-modified heme increases the diversity of potential NHR ligands.
3.4. Amino Acid Derivatives
Amino acids were the first metabolites identified as a source of ligands for NHRs. For example, thyroxine (T4) and the active derivative triiodothyronine (T3) are both derived from tyrosine and synthesized from a protein precursor, thyroglobulin, making it a protein/peptide-derived ligand (reviewed in [70]). In these cases, a dedicated organ, the endostyle in protochordates and the thyroid gland in jawed vertebrates, evolved to produce an iodinated amino acid derivative that has no other function in the organism but to act as a hormone [71]. Thyroid hormone-like substances are also found in other invertebrates, but their functions are still poorly understood. Interestingly, thyroid hormone-like substances are produced by marine algae (Diatoms), suggesting that these substances originally acted like vitamins for plankton-feeding organisms [71]. In accord with this hypothesis, the thyroid gland could have evolved exclusively to generate thyroid hormone-like substances in animals that do not feed on plankton [71]. Thus, a vitamin became a hormone. No other NHRs so far have been identified that recognize amino acid derivatives.
4. Orphan Receptors, What Ligands?
It has been suggested that some orphan NHRs are ligand-independent or constitutively active. A good example is NR4A1, an NHR with strong transcriptional activity in most mammalian cells. The main argument for NR4A1′s ligand-independent function is a crystal structure showing that the ligand-binding pocket of NR4A1 is too small to accommodate a ligand [72]. However, such evidence must be taken with a grain of salt. Initially, the crystal structure of REV-ERBβ identified a small ligand-binding pocket filled with bulky hydrophobic amino acid residues [73]. However, further studies showed that REV-ERBβ binds heme [50]. Another argument, given the conservation of the ligand backbones, is that orphan NHRs bind the same classes of ligands as non-orphan receptors. However, this focus on known ligands could be misleading. It is possible that we have not yet discovered all the molecules that affect our physiology.
What is the possibility that new vitamins could be found? Mice can be maintained on a chemically defined low molecular weight diet for several generations [74]. Similarly, Drosophila melanogaster can be reared in chemically defined conditions [75]. These results seemingly argue against the idea that there are unknown vitamins “out there”. On the other hand, many of these diets have undefined components extracted from vegetable sources that could introduce a contaminant into the system. The dramatic effect that such contaminants may have on development is illustrated by the example of C. elegans. Many nematodes are cholesterol auxotrophic [76]. and C. elegans can develop normally in chemically defined agar plates. However, when the sterol content of the agar is removed by extraction with chloroform and methanol, there is severe impairment of worm development [77]. Addition of the proper cholesterol enantiomer can completely restore C. elegans development in agar plates where all lipids are extracted with organic solvent [77]. Later, cholesterol was shown to be a precursor for dafachronic acid, which is a ligand for Daf12 [35], an NHR that is essential for C. elegans development.
One way to address whether there are exogenous components to NHR function is by testing NHR transcriptional activity in different cell lines in chemically defined medium [34]. For example, most cholesterol-sufficient mammalian cells, such as HEK293 or HeLa cells, show strong RORγ transcriptional activity [34]. In contrast, mammalian cell lines with genetic deletion in the cholesterol biosynthetic pathway and cells derived from cholesterol auxotroph, such as insect cells S2 and Kc167, had a reduction or a complete block in RORγ transcriptional activity [34]. Tissue culture medium is prepared from microbial sources, and there is little contamination with eukaryotic metabolites. An in-house, chemically defined medium can be developed with basal media such as DMEM, RPMI or Grace’s medium supplemented with insulin, transferrin and Pluronic F68 as a replacement for bovine serum albumin, warranting that there are no mammalian- or plant-derived molecules in the medium [34]. Detection of NHR activity in specific cell lines in a chemically defined medium is a good indicator that the ligand is endogenously produced or modified by the cells themselves. One can now identify the ligand by genetic and chemical means to certify whether it is a member of a known ligand family or an entirely new class of ligand.
What about new hormones produced by tissues or specific cells in the organism? This question can be addressed in the same manner as we addressed the possibility of new vitamins. Hormones are “messengers” generated by one cell as a communication component to another cell/organ. Such communication systems could act within the cell (autocrine fashion), on an adjacent cell (paracrine fashion) or on a distantly located cell (endocrine fashion). We would expect that hormone-like molecules should be produced by a restricted number of cell lineages.
New hormone-like molecules could come from many sources. One is intermediates in biosynthetic pathways. For example, 4α-carboxy-zymosterol and other biosynthetic intermediates with a double bond at carbon 8 have been shown to be ligands for RORα and RORγ [34]. Desmosterol, another intermediate of cholesterol biosynthesis, can also function as a ligand for RORγ [78] and LXR [79]. Another intermediate, 7-dehydrocholesterol, is a precursor for vitamin D (reviewed in [1]). Interestingly, some cholesterol biosynthetic intermediates accumulate in tissues. For example, FF-MAS accumulates in ovaries and T-MAS accumulates in the testis, where they promote meiosis [80]. Similarly, metabolic products of the conversion of lanosterol into FF-MAS, such as the 3β-lanost-32-aldehydes, can accumulate in cells [81,82]. The mechanism by which these sterols act is still unclear. However, it is tempting to speculate that these metabolites may act through NHRs via autocrine or paracrine mechanisms. Indeed, FF-MAS has been shown to activate LXR and RORγ in reporter assays [31,34], and lanosterol aldehydes have been suggested as candidate RORγ ligands [34]. It remains to be seen whether such intermediates or their derivatives could also have endocrine functions.
Sterol lipids are not the only pathways producing metabolites that could be used as hormone-like substances. Sphingolipids generally have a head group derived from serine. However, the preference for serine is dictated in cells by the availability of serine and alanine; at low serine, high alanine concentrations there is production of sphingolipids with alanine head-groups (1-deoxysphingosines) [83]. 1-deoxysphingosines lack the C1 hydroxyl group of serine-based sphingolipids, and they cannot serve as precursors for the synthesis of phospho- or glycosphingolipids or be degraded by the known sphingolipid catabolic pathways [84]. These are only a few examples of possible hormone-like compounds. Lipidomics studies have identified a large number of new lipids with unknown function [54], and some of these could possess hormone-like activities.
5. Common Properties of NHR Ligand Biosynthetic Pathways
The common feature of NHR ligands is a hydrophobic backbone associated with chemical groups that allow for the formation of hydrogen bonds or van der Waals interactions, for example the carboxy group of fatty acids and bile acids. There has been no report of a totally hydrophobic compound as an NHR ligand. The main sources of endogenous metabolites with hydrophobic backbones attached to chemical groups that allow for hydrogen bonding are the fatty acid and the terpenoid pathways. It is possible that the preference for these pathways may be an evolutionary accident since fatty acid derivatives and sterol lipids were the first ligands for the ancestral receptors [6]. Alternatively, these backbones could have been selected by chance, since the fatty acid and terpenoid pathways are also the most diverse groups of metabolites. Generally, the compounds that serve as the sources of ligands for NHRs are targeted by many processing enzymes. This includes enzymes that process polyunsaturated fatty acids into prostaglandins, resolvins and maresins [85,86], as well as the monooxygenases and dehydrogenases that process sterol lipids into steroid hormones, vitamin D and bile acids.
The majority of known NHR ligands are soluble metabolites that would allow for autocrine, paracrine or endocrine functions. There are a few notable exceptions: the ligand for PPARα is a phosphatidylcholine, a structural component of the membrane [87]. However, the jury is still out on PPARα, as other potential ligands have been suggested such as Coenzyme Q10 [88], oleylethanolamine [89] or 7-hydroxydocosahexaenoic acid [90]. Oleylethanolamine and 7-hydroxydocosahexaenoic acid are the most exciting findings, since they may play hormone-like or vitamin-like roles that fit with the standard properties we suggest for NHR ligands.
Author Contributions
L.J.T., D.E.S. and B.J. prepared figures and edited text. F.R.S. and N.B.I. conceptualized, wrote and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The work was supported by NIH grant R21-AI131015 (F.R.S.), Yale University institutional grant YD000234 (F.R.S.), NIH grant R01 GM107092 (NBI) and Young National Natural Science Fund of China, grant 81500101.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Santori, F.R. Nuclear hormone receptors put immunity on sterols. Eur. J. Immunol. 2015, 45, 2730–2741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graupner, G.; Wills, K.N.; Tzukerman, M.; Zhang, X.-K.; Pfahl, M. Dual regulatory role for thyroid-hormone receptors allows control of retinoic-acid receptor activity. Nat. Cell Biol. 1989, 340, 653–656. [Google Scholar] [CrossRef] [PubMed]
- Sakai, D.D.; Helms, S.; Carlstedt-Duke, J.; A Gustafsson, J.; Rottman, F.M.; Yamamoto, K.R. Hormone-mediated repression: A negative glucocorticoid response element from the bovine prolactin gene. Genes Dev. 1988, 2, 1144–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, R.; Ursu, O.; Gaulton, A.; Bento, A.P.; Donadi, R.S.; Bologa, C.G.; Karlsson, A.; Al-Lazikani, B.; Hersey, A.; Oprea, T.; et al. A comprehensive map of molecular drug targets. Nat. Rev. Drug Discov. 2017, 16, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Shpakov, A.O.; Pertseva, M.N. Chapter 4 Signaling Systems of Lower Eukaryotes and Their Evolution. Int. Rev. Cell Mol. Biol. 2008, 269, 151–282. [Google Scholar] [CrossRef] [PubMed]
- Bridgham, J.T.; Eick, G.N.; Larroux, C.; Deshpande, K.; Harms, M.J.; Gauthier, M.-E.A.; Ortlund, E.A.; Degnan, B.M.; Thornton, J.W. Protein Evolution by Molecular Tinkering: Diversification of the Nuclear Receptor Superfamily from a Ligand-Dependent Ancestor. PLoS Biol. 2010, 8, e1000497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthold, A.A. Transplantation der hoden. Arch. Anat. Physiol. Wissen Med. 1849, 16, 42–46. [Google Scholar]
- Séquard, B. Note on the effects produced on man by subcutaneous injections of a liquid obtained from the testicles of animals. Lancet 1889, 134, 105–107. [Google Scholar] [CrossRef] [Green Version]
- Murray, G.R. Note on the Treatment of Myxœdema by Hypodermic Injections of an Extract of the Thyroid Gland of a Sheep. Br. Med. J. 1891, 2, 796–797. [Google Scholar] [CrossRef]
- Baumann, E. Ueber das Thyrojodin. Munch. Med. Wschr. 1896, 43, 309–312. [Google Scholar]
- Starling, E.H. Croonian Lecture: On the chemical correlation of the functions of the body. I. Lancet 1905, 166, 339–341. [Google Scholar] [CrossRef]
- Starling, E.H. Croonian Lecture: On the chemical correlation of the functions of the body. IV. Lancet 1905, 166, 579–583. [Google Scholar] [CrossRef]
- Kendall, E.C. The isolation in crystalline form of the compound containing iodin, which occurs in the thyroid. Its chemical nature and physiological activity. J. Am. Med Assoc. 1915, 30, 2042–2043. [Google Scholar] [CrossRef] [Green Version]
- Harington, C.R.; Barger, G. Chemistry of Thyroxine: Constitution and Synthesis of Thyroxine. Biochem. J. 1927, 21, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Doisy, E.A.; Veler, C.D.; Thayer, S.A. Folliculin from Urine of Pregnant Women; American Phsyiological Society: Rockville, MD, USA, 1929. [Google Scholar]
- Butenandt, A. Über“ Progynon ”ein krystallisiertes weibliches Sexualhormon. NW 1929, 17, 879. [Google Scholar]
- Butenandt, A.; Hanisch, G. Über die Umwandlung des Dehydro-androsterons in Δ4-Androsten-ol-(17)-0n-(3) (Testosteron); ein Weg zur Darstellung des Testosterons aus Cholesterin (Vorläuf. Mitteil.). Ber. Dtsch. Chem. Ges. (A B Ser.) 1935, 68, 1859–1862. [Google Scholar] [CrossRef]
- Ruzicka, L.; Wettstein, A. Sexualhormone VII. Über die künstliche Herstellung des Testikelhormons Testosteron (Androsten-3-on-17-ol). Helvetica Chim. Acta 1935, 18, 1264–1275. [Google Scholar] [CrossRef]
- David, K.; Dingemanse, E.; Freud, J.; Laqueur, E. Über krystallinisches männliches Hormon aus Hoden (Testosteron), wirksamer als aus Harn oder aus Cholesterin bereitetes Androsteron. Biol. Chem. 1935, 233, 281. [Google Scholar] [CrossRef]
- Butenandt, A.; Westphal, U. Zur Isolierung und Charakterisierung des Corpus-luteum-Hormons. Ber. Dtsch. Chem. Gesellschaf 1934, 67, 1440–1442. [Google Scholar] [CrossRef]
- Mason, H.L.; Hoehn, W.M.; Kendall, E.C. Chemical studies of the suprarenal cortex: IV. Structures of compounds C,D,E,F, and G. J. Biol. Chem. 1938, 124, 459–474. [Google Scholar]
- Mason, H.L.; Myers, C.S.; Kendall, E.C. Chemical studies of the suprarenal cortex: II. The identification of a substance which possesses the qualitative action of cortin; its conversion into a diketone closely related to androstenedione. J. Biol. Chem. 1936, 116, 267–276. [Google Scholar]
- Reichstein, T. Über Bestandteile der Nebennieren-Rinde VI. Trennungsmethoden, sowie Isolierung der Substanzen F. a. H und J. Helv. Chem. Acta 1936, 19, 1107–1126. [Google Scholar] [CrossRef]
- Butenandt, A. Über die chemische Untersuchung der Sexualhormone. Angew. Chem. 1931, 44, 905–908. [Google Scholar] [CrossRef]
- Simons, S.S., Jr.; Thompson, E.B. Dexamethasone 21-mesylate: An affinity label of glucocorticoid receptors from rat hepatoma tissue culture cells. Proc. Natl. Acad. Sci. USA 1981, 78, 3541–3545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehring, U.; Hotz, A. Photoaffinity labeling and partial proteolysis of wild-type and variant glucocorticoid receptors. Biochemistry 1983, 22, 4013–4018. [Google Scholar] [CrossRef] [PubMed]
- Hollenberg, S.M.; Weinberger, C.; Ong, E.S.; Cerelli, G.; Oro, A.; Lebo, R.; Thompson, E.B.; Rosenfeld, M.G.; Evans, R.M. Primary structure and expression of a functional human glucocorticoid receptor cDNA. Nat. Cell Biol. 1985, 318, 635–641. [Google Scholar] [CrossRef]
- Giguere, V.; Ong, E.S.; Segui, P.; Evans, R.M. Identification of a receptor for the morphogen retinoic acid. Nat. Cell Biol. 1987, 330, 624–629. [Google Scholar] [CrossRef]
- Petkovich, M.; Brand, N.J.; Krust, A.; Chambon, P. A human retinoic acid receptor which belongs to the family of nuclear receptors. Nature 1987, 330, 444–450. [Google Scholar] [CrossRef]
- Clarke, N.D. Zinc Fingers in Caenorhabditis elegans: Finding Families and Probing Pathways. Science 1998, 282, 2018–2022. [Google Scholar] [CrossRef] [Green Version]
- Arenas, E. Faculty Opinions recommendation of An oxysterol signalling pathway mediated by the nuclear receptor LXR alpha. Nature 2015, 383, 728–731. [Google Scholar] [CrossRef]
- Makishima, M.; Okamoto, A.Y.; Repa, J.J.; Tu, H.; Learned, R.M.; Luk, A.; Hull, M.V.; Lustig, K.D.; Mangelsdorf, D.J.; Shan, B. Identification of a nuclear receptor for bile acids. Science 1999, 284, 1362–1365. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.J.; Blanchard, S.G.; Bledsoe, R.K.; Chandra, G.; Consler, T.G.; Kliewer, S.A.; Stimmel, J.B.; Willson, T.M.; Zavacki, A.M.; Moore, D.D.; et al. Bile acids: Natural ligands for an orphan nuclear receptor. Science 1999, 284, 1365–1368. [Google Scholar] [CrossRef] [PubMed]
- Santori, F.R.; Huang, P.; van de Pavert, S.A.; Douglass, E.F., Jr.; Leaver, D.J.; Haubrich, B.A.; Keber, R.; Lorbek, G.; Konijn, T.; Rosales, B.N.; et al. Identification of Natural RORgamma Ligands that Regulate the Development of Lymphoid Cells. Cell Metab. 2015, 21, 286–297. [Google Scholar] [CrossRef] [Green Version]
- Motola, D.L.; Cummins, C.L.; Rottiers, V.; Sharma, K.K.; Li, T.; Li, Y.; Suino-Powell, K.; Xu, H.E.; Auchus, R.J.; Antebi, A.; et al. Identification of ligands for DAF-12 that govern dauer formation and reproduction in C. elegans. Cell 2006, 124, 1209–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Zhou, X.E.; Motola, D.L.; Gao, X.; Suino-Powell, K.; Conneely, A.; Ogata, C.; Sharma, K.K.; Auchus, R.J.; Lok, J.B.; et al. Identification of the nuclear receptor DAF-12 as a therapeutic target in parasitic nematodes. Proc. Natl. Acad. Sci. USA 2009, 106, 9138–9143. [Google Scholar] [CrossRef] [Green Version]
- Retnakaran, A.; Krell, P.; Feng, Q.; Arif, B. Ecdysone agonists: Mechanism and importance in controlling insect pests of agriculture and forestry. Arch. Insect. Biochem. Physiol. 2003, 54, 187–199. [Google Scholar] [CrossRef]
- Nuclear Receptors Nomenclature Committee. A unified nomenclature system for the nuclear receptor superfamily. Cell 1999, 97, 161–163. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, B.E.; Hogenesch, J.B.; Bradfield, C.A. Mammalian Per-Arnt-Sim proteins in environmental adaptation. Ann. Rev. Phys. 2010, 72, 625–645. [Google Scholar] [CrossRef]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Bloch, K. The biological synthesis of cholesterol. Science 1965, 150, 19–28. [Google Scholar] [CrossRef]
- Villas-Boas, S.G. Metabolome Analysis: An Introduction; Wiley-Interscience: Hoboken, NJ, USA, 2007. [Google Scholar]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E.A.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon queenslandica genome and the evolution of animal complexity. Nat. Cell Biol. 2010, 466, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Hertz, R.; Magenheim, J.; Berman, I.; Bar-Tana, J. Fatty acyl-CoA thioesters are ligands of hepatic nuclear factor-4alpha. Nature 1998, 392, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Krylova, I.N.; Sablin, E.P.; Moore, J.; Xu, R.X.; Waitt, G.M.; Mackay, J.A.; Juzumiene, D.; Bynum, J.M.; Madauss, K.; Montana, V.; et al. Structural Analyses Reveal Phosphatidyl Inositols as Ligands for the NR5 Orphan Receptors SF-1 and LRH-1. Cell 2005, 120, 343–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyman, R.A.; Mangelsdorf, D.J.; Dyck, J.A.; Stein, R.B.; Eichele, G.; Evans, R.M.; Thaller, C. 9-cis retinoic acid is a high affinity ligand for the retinoid X receptor. Cell 1992, 68, 397–406. [Google Scholar] [CrossRef]
- Levin, A.A.; Sturzenbecker, L.J.; Kazmer, S.; Bosakowski, T.; Huselton, C.; Allenby, G.; Speck, J.; Kratzeisen, C.; Rosenberger, M.; Lovey, A.; et al. 9-cis retinoic acid stereoisomer binds and activates the nuclear receptor RXR alpha. Nature 1992, 355, 359–361. [Google Scholar] [CrossRef]
- Sap, J.; Muñoz, A.; Damm, K.; Goldberg, Y.; Ghysdael, J.; Leutz, A.; Beug, H.; Vennström, B. The c-erb-A protein is a high-affinity receptor for thyroid hormone. Nature 1986, 324, 635–640. [Google Scholar] [CrossRef]
- Weinberger, C.; Thompson, C.C.; Ong, E.S.; Lebo, R.; Gruol, D.J.; Evans, R.M. The c-erb-A gene encodes a thyroid hormone receptor. Nature 1986, 324, 641–646. [Google Scholar] [CrossRef]
- Raghuram, S.; Stayrook, K.R.; Huang, P.; Rogers, P.M.; Nosie, A.K.; McClure, D.B.; Burris, L.L.; Khorasanizadeh, S.; Burris, T.P.; Rastinejad, F. Identification of heme as the ligand for the orphan nuclear receptors REV-ERBalpha and REV-ERBbeta. Nat. Struct. Mol. Biol. 2007, 14, 1207–1213. [Google Scholar] [CrossRef] [Green Version]
- Umeno, A.; Sakashita, M.; Sugino, S.; Murotomi, K.; Okuzawa, T.; Morita, N.; Tomii, K.; Tsuchiya, Y.; Yamasaki, K.; Horie, M.; et al. Comprehensive analysis of PPARgamma agonist activities of stereo-, regio-, and enantio-isomers of hydroxyoctadecadienoic acids. Biosci. Rep. 2020, 40, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Kliewer, S.A.; Lenhard, J.M.; Willson, T.M.; Patel, I.; Morris, D.C.; Lehmann, J.M. A prostaglandin J2 metabolite binds peroxisome proliferator-activated receptor gamma and promotes adipocyte differentiation. Cell 1995, 83, 813–819. [Google Scholar] [CrossRef] [Green Version]
- Forman, B.M.; Tontonoz, P.; Chen, J.; Brun, R.P.; Spiegelman, B.M.; Evans, R.M. 15-Deoxy-delta 12, 14-prostaglandin J2 is a ligand for the adipocyte determination factor PPAR gamma. Cell 1995, 83, 803–812. [Google Scholar] [CrossRef] [Green Version]
- Fahy, E.; Subramaniam, S.; Murphy, R.C.; Nishijima, M.; Raetz, C.R.H.; Shimizu, T.; Spener, F.; Van Meer, G.; Wakelam, M.J.O.; Dennis, E.A. Update of the LIPID MAPS comprehensive classification system for lipids. J. Lipid Res. 2008, 50, S9–S14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christianson, D.W. Structural and Chemical Biology of Terpenoid Cyclases. Chem. Rev. 2017, 117, 11570–11648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruzicka, L. The isoprene rule and the biogenesis of terpenic compounds. Cell. Mol. Life Sci. 1953, 9, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.L.; Brown, M.S. Regulation of the mevalonate pathway. Nat. Cell Biol. 1990, 343, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.-H.; Zhu, J.; Jiang, J.-G. Carotenoids biosynthesis and cleavage related genes from bacteria to plants. Crit. Rev. Food Sci. Nutr. 2017, 58, 2314–2333. [Google Scholar] [CrossRef]
- Green, S.; Walter, P.; Kumar, V.; Krust, A.; Bornert, J.-M.; Argos, P.; Chambon, P. Human oestrogen receptor cDNA: Sequence, expression and homology to v-erb-A. Nat. Cell Biol. 1986, 320, 134–139. [Google Scholar] [CrossRef]
- Greene, G.L.; Gilna, P.; Waterfield, M.; Baker, A.; Hort, Y.; Shine, J. Sequence and expression of human estrogen receptor complementary DNA. Science 1986, 231, 1150–1154. [Google Scholar] [CrossRef]
- Jeltsch, J.M.; Krozowski, Z.; Quirin-Stricker, C.; Gronemeyer, H.; Simpson, R.J.; Garnier, J.M.; Krust, A.; Jacob, F.; Chambon, P. Cloning of the chicken progesterone receptor. Proc. Natl. Acad. Sci. USA 1986, 83, 5424–5428. [Google Scholar] [CrossRef] [Green Version]
- Conneely, O.M.; Sullivan, W.P.; O Toft, D.; Birnbaumer, M.; Cook, R.G.; Maxwell, B.L.; Zarucki-Schulz, T.; Greene, G.L.; Schrader, W.T.; O’Malley, B.W. Molecular cloning of the chicken progesterone receptor. Science 1986, 233, 767–770. [Google Scholar] [CrossRef]
- Chang, C.S.; Kokontis, J.; Liao, S.T. Molecular cloning of human and rat complementary DNA encoding androgen receptors. Science 1988, 240, 324–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubahn, D.B.; Joseph, D.R.; Sullivan, P.M.; Willard, H.F.; French, F.S.; Wilson, E.M. Cloning of human androgen receptor complementary DNA and localization to the X chromosome. Science 1988, 240, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Arriza, J.L.; Weinberger, C.; Cerelli, G.; Glaser, T.M.; Handelin, B.L.; E Housman, D.; Evans, R.M. Cloning of human mineralocorticoid receptor complementary DNA: Structural and functional kinship with the glucocorticoid receptor. Science 1987, 237, 268–275. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, D.P.; Mangelsdorf, D.J.; Pike, J.W.; Haussler, M.R.; O’Malley, B.W. Molecular cloning of complementary DNA encoding the avian receptor for vitamin D. Science 1987, 235, 1214–1217. [Google Scholar] [CrossRef] [PubMed]
- IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN). The Nomenclature of Steroids. JBIC J. Biol. Inorg. Chem. 2005, 10, 1–19. [Google Scholar]
- Reinking, J.; Lam, M.M.; Pardee, K.; Sampson, H.M.; Liu, S.; Yang, P.; Williams, S.P.; White, W.; Lajoie, G.; Edwards, A.M.; et al. The Drosophila Nuclear Receptor E75 Contains Heme and Is Gas Responsive. Cell 2005, 122, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Wu, N.; Curtin, J.C.; Qatanani, M.; Szwergold, N.R.; Reid, R.A.; Waitt, G.M.; Parks, D.J.; Pearce, K.H.; Wisely, G.B.; et al. Rev-erb, a Heme Sensor That Coordinates Metabolic and Circadian Pathways. Science 2007, 318, 1786–1789. [Google Scholar] [CrossRef]
- Brent, G.A. Mechanisms of thyroid hormone action. J. Clin. Investig. 2012, 122, 3035–3043. [Google Scholar] [CrossRef] [Green Version]
- Eales, J.G. Iodine metabolism and thyroid-related functions in organisms lacking thyroid follicles: Are thyroid hormones also vitamins? Proc. Soc. Exp. Boil. Med. 1997, 214, 302–331. [Google Scholar] [CrossRef]
- Wang, Z.; Benoit, G.; Liu, J.; Prasad, S.; Aarnisalo, P.; Liu, X.; Xu, H.; Walker, N.P.C.; Perlmann, T. Structure and function of Nurr1 identifies a class of ligand-independent nuclear receptors. Nat. Cell Biol. 2003, 423, 555–560. [Google Scholar] [CrossRef]
- Woo, E.J.; Jeong, D.G.; Lim, M.Y.; Jun Kim, S.; Kim, K.J.; Yoon, S.M.; Park, B.C.; Ryu, S.E. Structural insight into the constitutive repression function of the nuclear receptor Rev-erbbeta. J. Mol. Biol. 2007, 373, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Pleasants, J.R.; Johnson, M.H.; Wostmann, B.S. Adequacy of Chemically Defined, Water-Soluble Diet for Germfree BALB/c Mice Through Successive Generations and Litters. J. Nutr. 1986, 116, 1949–1964. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.C.; Micchelli, C.A. Development and characterization of a chemically defined food for Drosophila. PLoS ONE 2013, 8, e67308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hieb, W.F.; Rothstein, M. Sterol Requirement for Reproduction of a Free-Living Nematode. Science 1968, 160, 778–780. [Google Scholar] [CrossRef]
- Crowder, C.M.; Westover, E.J.; Kumar, A.S.; Ostlund, R.E., Jr.; Covey, D.F. Enantiospecificity of cholesterol function in vivo. J. Biol. Chem. 2001, 276, 44369–44372. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Wang, Y.; Hao, L.Y.; Liu, X.; Lesch, C.A.; Sanchez, B.M.; Wendling, J.M.; Morgan, R.W.; Aicher, T.D.; Carter, L.L.; et al. Sterol metabolism controls TH17 differentiation by generating endogenous RORgamma agonists. Nat. Chem. Biol. 2015, 11, 141–147. [Google Scholar] [CrossRef]
- Spann, N.J.; Garmire, L.X.; McDonald, J.G.; Myers, D.S.; Milne, S.B.; Shibata, N.; Reichart, D.; Fox, J.N.; Shaked, I.; Heudobler, D.; et al. Regulated Accumulation of Desmosterol Integrates Macrophage Lipid Metabolism and Inflammatory Responses. Cell 2012, 151, 138–152. [Google Scholar] [CrossRef] [Green Version]
- Byskov, A.G.; Andersen, C.Y.; Nordholm, L.; Thogersen, H.; Guoliang, X.; Wassmann, O.; Andersen, J.V.; Guddal, E.; Roed, T. Chemical structure of sterols that activate oocyte meiosis. Nature 1995, 374, 559–562. [Google Scholar] [CrossRef]
- Tabacik, C.; Aliau, S.; Serrou, B.; Crastes de Paulet, A. Post-HMG CoA reductase regulation of cholesterol biosynthesis in normal human lymphocytes: Lanosten-3 betal-ol-32-al, a natural inhibitor. Biochem. Biophys. Res. Commun. 1981, 101, 1087–1095. [Google Scholar] [CrossRef]
- Trzaskos, J.M.; Favata, M.F.; Fischer, R.T.; Stam, S.H. In situ accumulation of 3 beta-hydroxylanost-8-en-32-aldehyde in hepatocyte cultures. A putative regulator of 3-hydroxy-3-methylglutaryl-coenzyme A reductase activity. J. Biol. Chem. 1987, 262, 12261–12268. [Google Scholar]
- Esaki, K.; Sayano, T.; Sonoda, C.; Akagi, T.; Suzuki, T.; Ogawa, T.; Okamoto, M.; Yoshikawa, T.; Hirabayashi, Y.; Furuya, S. l-Serine Deficiency Elicits Intracellular Accumulation of Cytotoxic Deoxysphingolipids and Lipid Body Formation. J. Biol. Chem. 2015, 290, 14595–14609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lone, M.A.; Santos, T.; Alecu, I.; Silva, L.C.; Hornemann, T. 1-Deoxysphingolipids. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Pro-resolving lipid mediators are leads for resolution physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Petasis, N.A. Resolvins and Protectins in Inflammation Resolution. Chem. Rev. 2011, 111, 5922–5943. [Google Scholar] [CrossRef] [Green Version]
- Chakravarthy, M.V.; Lodhi, I.J.; Yin, L.; Malapaka, R.R.; Xu, H.E.; Turk, J.; Semenkovich, C.F. Identification of a physiologically relevant endogenous ligand for PPARalpha in liver. Cell 2009, 138, 476–488. [Google Scholar] [CrossRef] [Green Version]
- Tiefenbach, J.; Magomedova, L.; Liu, J.; Reunov, A.A.; Tsai, R.; Eappen, N.S.; Jockusch, R.A.; Nislow, C.; Cummins, C.L.; Krause, H.M. Idebenone and coenzyme Q10 are novel PPARalpha/gamma ligands, with potential for treatment of fatty liver diseases. Dis. Model. Mech. 2018, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Gaetani, S.; Oveisi, F.; Verme, J.L.; Serrano, A.; De Fonseca, F.R.; Rosengarth, A.; Luecke, H.; Di Giacomo, B.; Tarzia, G.; et al. Oleylethanolamide regulates feeding and body weight through activation of the nuclear receptor PPAR-α. Nature 2003, 425, 90–93. [Google Scholar] [CrossRef]
- Zhang, M.; Sayyad, A.; Dhesi, A.; Orellana, A. (Enantioselective Synthesis of 7(S)-Hydroxydocosahexaenoic Acid, a Brain-Specific Endogenous Ligand for PPARalpha. J. Org. Chem. 2020. [Google Scholar] [CrossRef]
Figure 1.
A brief history of nuclear hormone receptors (NHRs). Shown are the landmark moments in the history of NHRs.
Figure 1.
A brief history of nuclear hormone receptors (NHRs). Shown are the landmark moments in the history of NHRs.

Figure 2.
The 48 human NHRs and their ligands. Nuclear hormone receptors are defined by their homology to the steroid receptors [38]. This feature distinguishes the NHRs from other nuclear receptors regulated by small molecules such as the Per-Arnt-Sim (PAS) superfamily of transcription factors [39]. A dendrogram of the human nuclear hormone receptor family was generated using phylogeny.fr [40]. Color-coded balloons are used to label the ligand class for each individual NHR.
Figure 2.
The 48 human NHRs and their ligands. Nuclear hormone receptors are defined by their homology to the steroid receptors [38]. This feature distinguishes the NHRs from other nuclear receptors regulated by small molecules such as the Per-Arnt-Sim (PAS) superfamily of transcription factors [39]. A dendrogram of the human nuclear hormone receptor family was generated using phylogeny.fr [40]. Color-coded balloons are used to label the ligand class for each individual NHR.

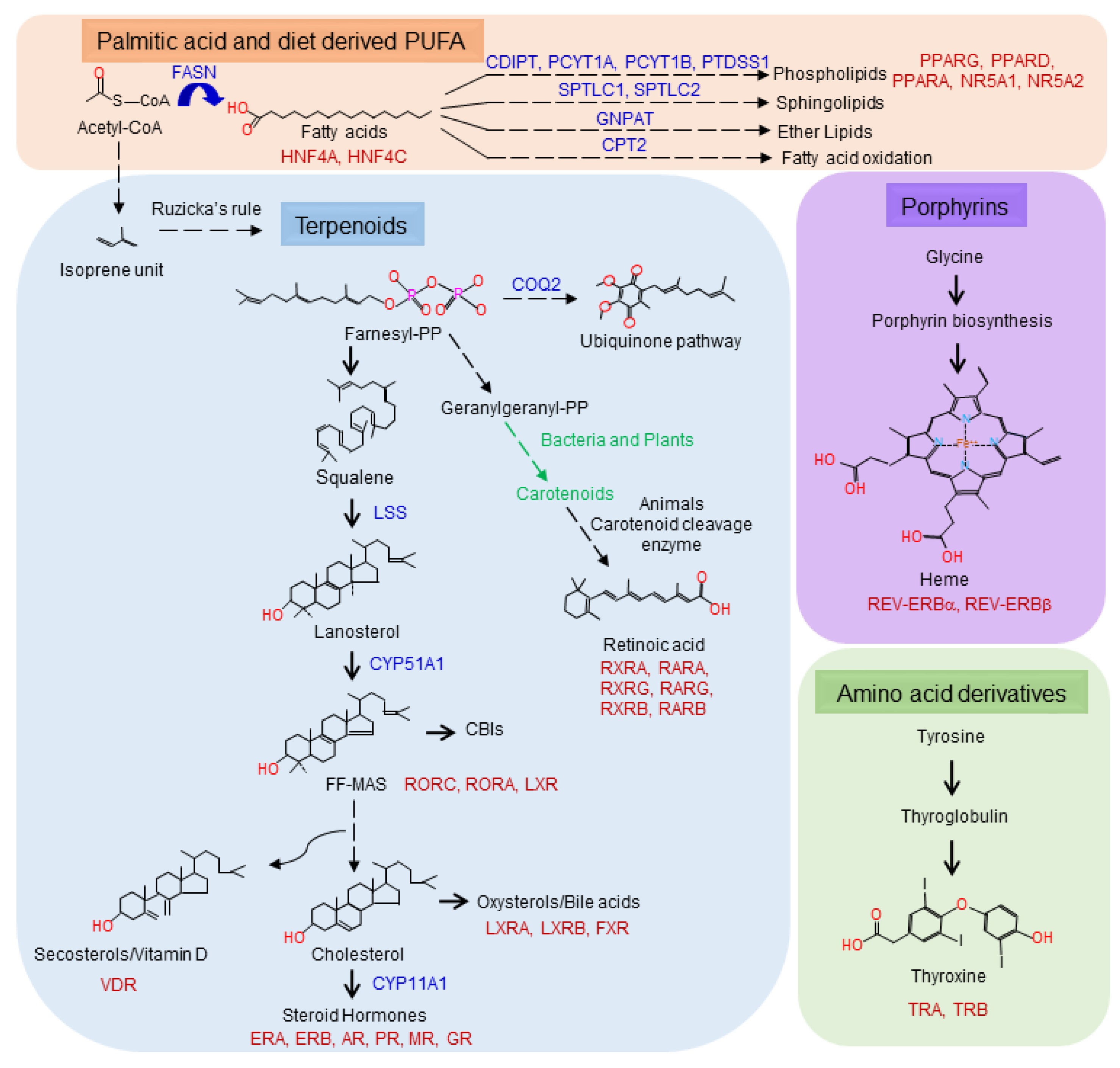
Figure 3.
The biosynthetic routes for the known sources of human NHR ligands. Fatty acids (orange) and terpenoids (blue) are generated from acetyl-CoA. Fatty acids can be further modified into phospholipids, glycerolypids, sphingolipids, ether lipids and fatty acid oxidation products. Acetyl-CoA is used to generate the isoprene units for terpenoid biosynthesis. Mammals will generate farnesyl and geranylgeranyl pyrophosphate, dolichol (not shown), ubiquinone and cholesterol. Bacteria and plants utilize geranylgeranyl pyrophosphate to generate carotenoids that can be used by animals to generate retinoic acid. Porphyrins are synthesized from glycine and thyroid hormones from tyrosine.
Figure 3.
The biosynthetic routes for the known sources of human NHR ligands. Fatty acids (orange) and terpenoids (blue) are generated from acetyl-CoA. Fatty acids can be further modified into phospholipids, glycerolypids, sphingolipids, ether lipids and fatty acid oxidation products. Acetyl-CoA is used to generate the isoprene units for terpenoid biosynthesis. Mammals will generate farnesyl and geranylgeranyl pyrophosphate, dolichol (not shown), ubiquinone and cholesterol. Bacteria and plants utilize geranylgeranyl pyrophosphate to generate carotenoids that can be used by animals to generate retinoic acid. Porphyrins are synthesized from glycine and thyroid hormones from tyrosine.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tao, L.J.; Seo, D.E.; Jackson, B.; Ivanova, N.B.; Santori, F.R. Nuclear Hormone Receptors and Their Ligands: Metabolites in Control of Transcription. Cells 2020, 9, 2606. https://doi.org/10.3390/cells9122606
AMA Style
Tao LJ, Seo DE, Jackson B, Ivanova NB, Santori FR. Nuclear Hormone Receptors and Their Ligands: Metabolites in Control of Transcription. Cells. 2020; 9(12):2606. https://doi.org/10.3390/cells9122606
Chicago/Turabian StyleTao, Lian Jing, Dong Eun Seo, Benjamin Jackson, Natalia B. Ivanova, and Fabio Rinaldo Santori. 2020. "Nuclear Hormone Receptors and Their Ligands: Metabolites in Control of Transcription" Cells 9, no. 12: 2606. https://doi.org/10.3390/cells9122606
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.