Construction of Marker-Free Genetically Modified Maize Using a Heat-Inducible Auto-Excision Vector

National Key Laboratory of Crop Genetic Improvement and College of Plant Science and Technology, Huazhong Agricultural University, Wuhan 430070, China
*
Author to whom correspondence should be addressed.
Genes 2019, 10(5), 374; https://doi.org/10.3390/genes10050374
Submission received: 22 February 2019
/
Revised: 10 May 2019
/
Accepted: 13 May 2019
/
Published: 17 May 2019
(This article belongs to the Section Technologies and Resources for Genetics)
Abstract
:Gene modification is a promising tool for plant breeding, and gradual application from the laboratory to the field. Selectable marker genes (SMG) are required in the transformation process to simplify the identification of transgenic plants; however, it is more desirable to obtain transgenic plants without selection markers. Transgene integration mediated by site-specific recombination (SSR) systems into the dedicated genomic sites has been demonstrated in a few different plant species. Here, we present an auto-elimination vector system that uses a heat-inducible Cre to eliminate the selectable marker from transgenic maize, without the need for repeated transformation or sexual crossing. The vector combines an inducible site-specific recombinase (hsp70::Cre) that allows for the precise elimination of the selectable marker gene egfp upon heating. This marker gene is used for the initial positive selection of transgenic tissue. The egfp also functions as a visual marker to demonstrate the effectiveness of the heat-inducible Cre. A second marker gene for anthocyanin pigmentation (Rsc) is located outside of the region eliminated by Cre and is used for the identification of transgenic offspring in future generations. Using the heat-inducible auto-excision vector, marker-free transgenic maize plants were obtained in a precisely controlled genetic modification process. Genetic and molecular analyses indicated that the inducible auto-excision system was tightly controlled, with highly efficient DNA excision, and provided a highly reliable method to generate marker-free transgenic maize.
1. Introduction
Genetic engineering techniques generally involve the addition of gene (single or multiple) integrated into a plant leading to the modification of its genome [1]. Transgene technique can be used to alter the expression patterns of individual gene in a more precise and predictable manner than conventional breeding [2], and has progressed from being a basic laboratory research technique to become the sole provider of new plant varieties worldwide over the last two decades [3]. There is much controversy over genetically modified (GM) crop: the uncertainty of their safety is the main concern of governments, non-governmental organizations, and the general public [4]. A common concern is the marker genes used for selecting transformed plants, asserting that the food produced from GM plants could be harmful to human health due to potential toxicity or allergenicity [5].
Selectable marker genes are essential to plant genetic engineering and used in nearly all transformation procedures to simplify the identification of transgenic plants. Selectable markers, such as hygromycin, kanamycin, or glyphosate, differentiate transformed cells pass resistance toward antibiotics or herbicides to the transformed cells, whereas untransformed cells and tissues are killed by treatment with lethal compounds [6]. These systems not only allow relatively straightforward identification and selection of stably incorporated plants [7], but also are used to follow the inheritance of foreign gene in the segregating generation [8]. Selectable marker genes are required for obtaining transgenic plants, but serve undesirable to be retained in transgenic plants because of their possible toxicity or allergenicity to humans and their unpredictable hazards to the environment [9]. Moreover, the continued presence can pose technological problems because it precludes additional transformation of constructs with the same marker system [10]. Compared with the existing selection marker genes, the selection of novel selection marker genes is an effective method to eliminate their potential risks. According to the mechanism of action of the safety marker developed and the type of agent selected, it can be divided into amino acid metabolism related genes [11,12], hormone related genes [13,14], sugar metabolism related genes [15], stress resistance related genes [16], and fluorescent protein genes [17,18,19]. The use of these selection markers reduces the potential risks of transgenic plants.
The removal of marker genes would likely hasten public acceptance of transgenic crops in a sense [20,21,22], because the production of marker-free transgenic crops is crucial to avoid these potential risks and promote commercial deployment of GM plants [23,24,25]. Several approaches have been developed for the removal of selectable marker genes from transgenic plants [26,27,28,29]. In the first approach, the selectable marker genes and the gene of interest are introduced at different loci of the plant genome by co-transformation (the mechanism is shown in Figure 1(a1–a4), after which the selectable marker gene is segregated out by crossing sexually [30]. The second method entails the elimination of selectable markers by transposition [31,32]. To eliminate the selectable marker using transposon-mediated transgene reintegration is an advantageous strategy for marker gene removal (the mechanism is shown in Figure 1(b1,b2), because it allows intact transgene insertion with defined boundaries and requires only a few primary transformants [33]. The third approach used for the removal of selectable marker genes is by intrachromosomal site-specific recombination (the mechanism is shown in Figure 1c) [34,35,36]. Site-specific recombination (SSR) systems carryout recombination between specific DNA sequences without addition or deletion of nucleotides [37,38,39,40]. There are well-known recombination systems, including Cre/lox from bacteriophage P1 [3,41], Flp/frt from Saccharomyces cerevisiae [42,43], R/RS from Zygosaccharomyces rouxii [44], and Gin/gix from bacteriophage [45] to remove the marker gene [46].These precisions are highly desirable in plant genetic engineering as they minimize unintended mutations introduced by the transformation process. Marker excision combined with site-specific gene integration requires the use of two SSR systems [13,47], and for a long time, only Cre/lox system was popularly used [48].
These simple SSR systems consist of two basic components: a recombination enzyme and small DNA recognition sites, enabling a variety of applications such as site-specific integration, copy number reduction, and marker gene removal [49,50,51,52]. A number of strategies can be used with Cre/lox to remove marker genes from transgenic plants and each strategy differs in how Cre function is delivered [41,53]. In all strategies, directly repeated lox sites flank the marker gene and excision occurs when Cre is active. In one strategy, similar to the co-transformation of the transposon system, transgenic plants that carry a marker gene flanked by lox sites can be retransformed with a Cre-expressing plasmid, and the Cre gene segregates in the next generation [47,54,55,56]. Alternatively, marker gene flanked by lox sites and Cre-expressing construction were on two plasmids, whereby lox plants can be crossed with plants that express Cre (herein referred to as the “crossing strategy”); in this case, marker excision occurs in the F1 progeny followed by the loss of the Cre gene by genetic segregation in the F2 generation [57,58,59,60].
The third strategy is referred to as the “induction-excision strategy”; in this strategy, Cre and selective marker genes are built between two lox sites directly, through induction type promoter or driven by cell-specific promoter for Cre recombinant enzyme gene expression, to remove selection marker gene to solve the simple delete operation [38,47]. Gleave et al. [61] successfully deleted the marker genes by expressing the Cre recombinase in transgenic tobacco, thus avoiding the process of retransformation and hybridization. Based on this principle, Hoff et al. [62] constructed a pCrox vector system: the thermal promoter GSP81-1Cre induced the Cre recombinase gene expression, and eliminated the kanamycin resistance gene which was inserted between the two loxP sites in transgenic Arabidopsis thaliana. An important improvement of this early technique was reported by Zuo et al. [63] using a chemically inducible artificial transcription factor for indirect transcriptional regulation of Cre-recombinase gene expression. Thus, the recombinase gene and the lox recombination sites could coexist without leading to premature recombination. Following the selection of transgenic tissue, chemical induction of the recombinase gene produced the desired excision events. Heat-inducible promoters are the widely used and studied promoters for driving site-specific recombinases [42,56,64,65,66,67].
In maize, co-transformation strategies [68,69,70,71] and transposon-induced recombination [72] have been reported to remove the selection marker genes. The Cre/lox system has also attracted the attention of maize researchers. In 2003, Zhang et al. [3] first reported the use of site-specific recombination to eliminate the selection marker gene in the transgenic maize. The researchers used the strategy to transfer the Cre-recombinase gene and the selection marker gene with lox sequence into two maize transformed lines. The Cre/lox system activated in the F1 hybridization and marker-free transgenic maize were separated in backcross material. Vega et al. [73] constructed a BIBAC vector system to adapt for maize transformation with a chemically induced marker elimination system, and avoided the complex operation of separating hybrid offspring. Based on the previous studies, site-specific recombinant system Cre/lox that was used to eliminate the selection markers from transgenic maize should solve two drawbacks: the difficulty in establishing a control elimination system and the formation of genetic chimeras due to incomplete DNA excision. For this purpose, the following works were carried out in this study: (i) We constructed a heat-inducible auto-excision vector using a heat-shock-responsive HSP70 promoter [74,75,76,77]; compared to previously reported systems, this new system is tightly controlled and the induced DNA excision is highly efficient. (ii) We used green fluorescent protein gene (egfp) as selection marker to screen transgenic callus. Compared with the existing herbicide resistance markers, this marker has the advantages of convenient detection and no harm to living cells in the screening process. In addition, the visible fluorescence phenotype can effectively identify the elimination of selection marker genes [18]. In the present study, marker-free transgenic maize plants were obtained via recombination-programmed auto-excision without any extra handling borrowed from the traditional transformation process with a few modifications by the vector.
2. Materials and Methods
2.1. Plant Material
Hi-II hybrid maize (A188 × B73 origin) was used as the plant material and was cultivated on an experimental farm at Huazhong Agricultural University in Wuhan, China. Following self-pollination at approximately 10–14 days, the ears were harvested when the immature embryos were 1.0–2.0 mm in length. To sterilize their surface, the ears were incubated in 70% ethanol for 5 min and then in 2.4% sodium hypochlorite supplemented with 0.1% Tween for 20 min, before being washed five times in sterile distilled water for 5 min every time. Immature embryos of normal and healthy morphology were isolated with a lancet and collected in a 2 mL tube containing 1 mL of sterile distilled water. A type II callus, which is clumps of nonpolar cells with bright color, loose structure, granular shape, dry surface, and rapid growth, was generated after 60–90 days in the induction medium.
2.2. Construction of Automatic Elimination Vector
The goal of this strategy is to obtain transformation and marker auto-excision in a single process using an induced expression site-specific recombination system. We constructed a vector using the skeleton of pEGAD, which is schematically represented in Figure 2. The egfp gene driven by the CaMV35S promoter functioned as a transformation selectable marker, constructed to assemble the DNA fragments of the selection element [78,79]. The egfp carried by the vector was used for two purposes. First, green fluorescent protein expression functioned as a visual selection signal that allows the direct visualization of transformed cells. Second, the loss of green fluorescence in the heat-shock-treated calli served as a counter-selectable marker that effectively eliminates the recombination element and selection markers. To remove the selection markers from the resulting transfer DNA (T-DNA), we employed the Cre/lox system [53]. The bacteriophage Cre recombinase specifically recognizes both lox sites in transformation cells. The recombinase Cre gene carrying a nuclear localization signal and the heat-shock promoter Hsp70 from maize [75,76,77], which is used to control its expression, was positioned between a lox site and the selection element. The whole element was flanked by two lox sites. Further, the heat-inducible cre gene located within the two lox sites can be removed by inducing Cre activity at any time after selection of the transgenic clones [80]. To test the efficacy of the vector system described above, we cloned the polyubiquitin gene (ubi) promoter [81] and controlled a maize anthocyanin pigmentation (Rsc) gene [82] into the construct (pHZM1N-Rsc) between the lox site and the left border (LB).
The vector was compounded by Make Research Easy (Nanjing, China) and was transferred into Agrobacterium tumefaciens strain EHA105 by electroporation. The bacteria were precultivated for 2–3 days on LB solid medium with 100 mg/L kanamycin at 28 °C in the dark [79]. On the day of transformation, Agrobacterium colonies were collected from the plate with a spatula, resuspended in Infection medium with 100 μM acetosyringone (Table 1), and incubated for 2–3 h at 28 °C and 200 rpm [79]. Cell density, measured as OD600, was adjusted prior to callus infection (see below).
2.3. Media
2.4. Plant Transformation by Agrobacterium Transgenic System on Calli
Plant transformation followed an established standard protocol with some modifications [79]. Embryonic calli were collected in 10 mL microcentrifuge tubes containing liquid infection medium with 100 μM acetosyringone and were allowed to sit for 20 min. The solution was drawn off, the calli were then infected with 5 mL of Agrobacterium suspension containing the pHZM1N-Rsc binary vector, followed by brief vortexing for 5 min at 28 °C. The Agrobacterium liquid was drawn off and the infected calli were transferred onto the solidified co-cultivation medium, and were tiled on the medium (Table 1). The plate was sealed with Parafilm and incubated in the dark at 19 °C for 3 days. After co-cultivation, the calli were rinsed 4 times with sterile distilled water followed by 2 times (5 min) with sterile water containing 100 mg L−1 carbenicillin (ICN, Costa Mesa, CA, USA) and were blotted dry on a sterile filter paper. They were then transferred onto the resting medium (Table 1) containing 100 mg L−1 carbenicillin (ICN, Costa Mesa, CA, USA) and incubated at a temperature of 28 ± 2 °C for 7 days in the dark.
2.5. GFP Fluorescence Assay
After 7 days restoration of culture, the instantaneous conversion efficiency was calculated by observing the green fluorescence performance on a DR-46B Dark Reader transilluminator (Clare Chemical Research, Dolores, CO, USA) and a digital camera (a550; Sony, Tokyo, Japan) with a GFP filter. Then, the calli were transferred to selection medium (Table 1) to selection culture in the dark at 28 ± 2 °C. Two weeks after the 1st round of selection, tissues were transferred to fresh selection medium and sub-cultured at 2–3 weeks intervals. After the third round of selection, the conversion efficiency was calculated by observing the green fluorescence performance on the DR-46B Dark Reader transilluminator; the statistical method showed the same instantaneous conversion efficiency as calculated by the experiment [32,79].
2.6. Heat-Shock Induction
The selected GFP-positive calli were broken into small pieces (1 × 1 mm) and transformed onto selection media in glass plates after three rounds of GFP fluorescence performance selection. The glass plates were transferred to a 42 °C incubator for 2 h in the dark to induce Cre activity as described [66,83]. The calli were then transferred onto new resting medium containing 100 mg/L carbenicillin (ICN, Costa Mesa, CA, USA) and incubated at 28 ± 2 °C in the dark for 7 days. After 3 weeks of subculture on selection medium at 28 ± 2 °C in the dark, the loss of GFP expression was used to examine the auto-excision efficiency of the cre recombinase; GFP-negative callus blocks from calli were subcultured from each independent experiment, and the calli with green fluorescence expression were transferred to a 42 °C incubator and kept for 2 h in the dark for heat-shock treatment for the second time. After two rounds of selection, the green fluorescence phenotypic characterization in maize calli were detected with a fluorescent protein macro detector set (Nikon, Tokyo, Japan); the GFP-negative callus blocks from calli were subcultured and redifferentiated. Regenerative green tissues were then transferred to regeneration medium (Table 1) and incubated at 28 ± 2 °C under dim light (10–30 mE m−2 s−1, 16/8 h day and night photoperiod) for 2–3 weeks. The shoots were transferred to rooting medium (Table 1) and exposed to higher light intensity (50–150 mE m−2 s−1, 16/8 h day and night photoperiod) for rooting. The plantlets were then transferred to soil [79].
2.7. Maize Genomic DNA Extraction
Maize calli and/or leaf tissue was frozen in liquid nitrogen and ground to a fine powder. Genomic DNA was extracted by the CTAB (hexadecyltrimethylammonium bromide) method from the frozen tissue. Callus or seedling leaves broken by physical means was incubated in a water bath at 65 °C for 40 min in a tube containing 500 µL CTAB buffer, and then centrifuged at 12,000 rpm for 10 min. The supernatant was transferred into a new tube, and an equal volume of isopropanol was added [79]. The solution was gently mixed and then centrifuged at 12,000 rpm for 5 min. After washing with 75% (v/v) ethanol and drying in air, the DNA pellet was dissolved in 2 mL of distilled water [79].
2.8. PCR Amplification of egfp, Rsc and Residual Sequence (RS) in Transgenic Maize Plants
The PCR monitoring area and probe location are shown in Figure 2b. The selection marker gene egfp, the Rsc gene, and the residual sequence (RS) after the marker was eliminated were amplified by PCR using the primer pairs P1 (5′-GGACTGGGTGCTCAGGTAGTGG-3′; 5′-CTGGACGGCGACGTAAACGG-3′), P2 (5′-GGGTTTAGGGTTAATGGT-3′; 5′-CACTGGCAAGTTAGCAAT-3′), and P3 (5′-AAACGGAGCATAGAGGATA-3′; 5′-CACTGGCAAGTTAGCAAT-3′), respectively. The expected size of the egfp-PCR product was 879 bp, and the expected size of the Rsc-PCR product was 465 bp, which contained part of the sequence of the ubi promoter and part of the Rsc gene. The size of the selection construct was initially 6836 bp, and it was 1729 bp after marker elimination. The cycling parameter was 94 °C for 5 min; 33 cycles of 94 °C for 30 s, X °C for 30 s, and 72 °C for 2 min; and 72 °C for 5 min. The annealing temperatures (X marks in the parameter) were 55 °C for egfp-PCR, 58 °C for Rsc-PCR and 50 °C for residual sequence (RS), separately.
2.9. Southern Blot Analysis
Southern-blot hybridization was conducted using the PCR DIG Probe Synthesis Kit and the DIG High Prime DNA Labeling and Detection Starter Kit II (Roche, Basel, Switzerland) according to the manufacturer’s instructions. Genomic DNA of the transgenic plants was extracted by the CTAB method [84] from GFP-positive callus. For Southern blot analysis, 25 μm of maize genomic DNA from each sample was digested to completion with 10 units of AscI, fractionated on a 1% (w/v) agarose gel, and transferred onto an Amersham Hybond N++ membrane (GE Healthcare, Chicago, IL, USA) according to the instructions [85]. Probe 1 (Rsc: 465 bp, 5′-GGGTTTAGGGTTAATGGT-3′ and 5′-CACTGGCAAGTTAGCAAT-3′) was synthesized using the PCR DIG Probe Synthesis Kit according to the manufacturer’s instructions. DNA membranes were hybridized with the probes in DIG Easy Hyb (Roche) solution at 42 °C. After hybridization, the membranes were washed at 65 °C two times with 2 × SSC and 0.1% (w/v) SDS for 10 min and two times with 0.1 × SSC and 0.1% (w/v) SDS for 15 min [36]. Probe hybridization signal was examined by digoxigenin chemiluminescence detection. Membranes were stripped afterward by washing at 37 °C two times with 0.2 N NaOH and 0.1% (w/v) SDS for 30 min and one time with 2 × SSC for 5 min. Further hybridization was performed using the AscI marker (New England Biolabs, Ipswich, MA, USA) that was labeled using the DIG High Prime DNA Labeling Kit.
3. Results
3.1. Determination of Transformed Callus
High quality embryogenic calli of Hi-ΙΙ maize were chosen for highly efficient maize transformation achieved using the Agrobacterium–mediated methods shown by Du et al. [79]. Three weeks after an infection, we obtained 200 transgenic calli when GFP fluorescence phenotypic (Figure 3a) assayed on the DR-46B Dark Reader transilluminator. The GFP phenotype was because of the transformation-positive calli, while the non-fluorescing parts were due to the callus being untransformed. The transformation efficiency, defined as the weight ratio of the calli with GFP activity to the total weight of co-cultivated calli that produced secondary calli, was calculated to be approximately 16.7% at three weeks of culture selection from the co-cultivated calli.
Six transgenic events, based on the fluorescence assays, were chosen to analysis by PCR and Southern blotting. The PCR monitoring area and probe location are shown in Figure 3b and the results are shown in Figure 3c: P1 and P2 produced the expected product sizes; the PCR bands were 879 bp and 465 bp, respectively, for all six events, but no band was amplified from any of the seven calli by P3 because the product is too large for the given PCR conditions. An additional independent confirmation was obtained by Southern blot analysis. Genomic DNA from each event was digested and hybridized with probes specific to the polyubiquitin gene (ubi) promoter and the maize reporter gene (Rsc) linked sequence, as revealed for all six events in Figure 3d. Most transgenic plants contained a single band of the transgene. No. 2, No. 5, and No. 6 had a single fragment; however, the sizes of the copies were different. No. 2 and No. 5 had larger fragments compared to the positive control, while No. 6 with had a smaller band. No. 1 had two bands, and No. 3 had three bands. No. 4 perhaps had one smaller band at the similar size with one band in No. 3.
3.2. Induction of Marker Excision and Molecular Analysis of Calli
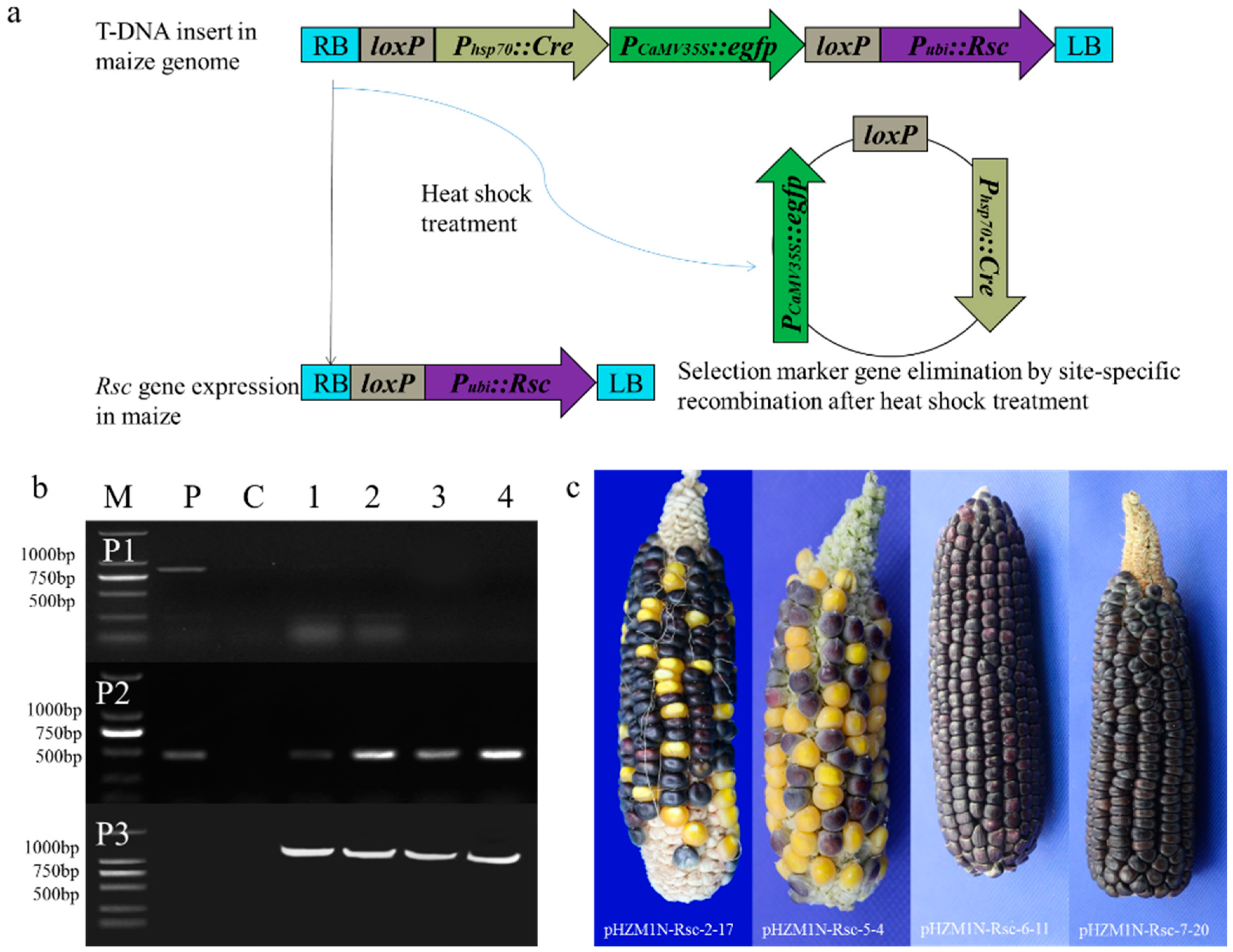
The transformed calli containing a single copy were chosen for marker excision by heat-shock treatment; the loss of GFP expression was used to examine the auto-excision efficiency of the cre recombinase (schematic diagram shown in Figure 4a). After the first heat-shock treatment, the calli were transferred onto the resting medium for 7 days and then to subcultured on selection medium at 28 ± 2 °C in the dark for 3 weeks. The frequency of elimination condition was 21.95%, 6.38%, 19.05%, and 16.28%, by the mean percentage of GFP-negative callus blocks to calli subcultured from each independent experiment (Table 2). The selection marker gene had not been successfully removed in the calli in which green fluorescence had not completely disappeared, and those calli were treated for the second round of heat-shock-induction treatment. After two rounds of heat-shock treatment, the calli were transferred onto the resting medium at 28 ± 2 °C in the dark for 7 days. After 3 weeks, subcultured on selection medium at 28 ± 2 °C in the dark, callus blocks without the green fluorescence phenotype were characterized with a fluorescent protein macro detector set (Nikon, Tokyo, Japan) as the GFP-negative callus blocks. In addition, PCR analysis also obtained the product of the anthocyanin pigmentation gene (Rsc) with the 465 bp banding, marker gene detection product disappeared at the same time, and primer P3 PCR analysis obtained expectant 1729 bp band after marker elimination (Figure 4b). These results illustrate that the Rsc gene remains stable in the callus genome after heat-shock treatment but that the selection marker was eliminated, which is consistent with the effect on GFP expression. Conversely, due to the elimination of the selection marker, the fragment between the LB and the Rsc gene greatly shortened. A 1729-bp PCR product was detected with the P3 primers, as expected for excision of the reporter gene. At the same time, the positive control was too large for the given PCR conditions. In summary, PCR analysis demonstrated the efficient excision of both fragments flanking the selection marker and recombinant enzyme construct after heat treatment of the GFP-positive calli derived from four different transformations. Next, the inheritance of the marker-free locus was studied by analyzing the progeny of these plant lines [86].
3.3. Regenerated, Molecular Analyzed, and Phenotypes Authenticate of Transgenic Plants
The transformed plants (T0) were regenerated from four independent events. Plants were transplanted into soil and transferred to greenhouse, where they grew for 2–3 weeks before harvesting leaves for the isolation of genomic DNA. Selection marker excision was detected in 100% (17/17) of the tested plants, while the transformed anthocyanin pigmentation gene (Rsc) remained integrated in the transgenic plants (Table 3). The maize seeds of each T0 line were harvested to examine the expression of the regulator gene to distinguish transformed from non-transformed seeds (Figure 4c).
Transformed seeds were randomly selected from each of the cultivar combinations for sowing. A total of 113 regulator-positive T1 plants were used for DNA analysis using regulator-specific primers (P2) and individual plants were further analyzed using PCR to amplify the gfp (P1) and the RS of the marker-eliminated sequence (P3). The parents of all the selected marker-free T1 plants showed the rsc+gfp−RS+, as shown by PCR results. The segregation ratio of purple seeds to yellow seeds in each T1 line was investigated using the χ2 test by the percentage count of purple and yellow grains on the ear (Table 4). Fifteen (88%) lines segregated with a 3:1 Mendelian pattern of inheritance, and the other two (12%) lines segregated with non-Mendelian ratios of distortion toward non-regulator seeds. The progeny of each T0 plant showed a complete marker excision, and a high percentage of T1 progeny from each T0 line is expected to contain a marker-free locus, which was confirmed by PCR on the excision footprint (RS).
4. Discussion
The transformation of most crop species has been rapidly achieved using various techniques, but it’s always relatively inefficient no matter which is used [87]. Successful genetic transformation requires not only efficient gene delivery but also an efficient selection system [88] that actively separates the transformed cells that are capable of regenerating plants from non-transformed cells. Currently, a gene with resistance to antibiotics or herbicides is usually selected in maize as the screening marker of transgenic genes. Selectable markers allow the transformed cells survive to tolerate an otherwise lethal exposure and grow into plants, and when the untransformed tissues are killed [89]. Without them, the creation of transgenic crops is not feasible on purely economic and practical terms [90]. There is concern that antibiotic resistance genes in genetic modification crops has chance to flow of into the genomes of the microorganisms living in the soil, might lead to the development of antibiotic-resistant pathogens [23,91]. A similar concern arises when genes that confer herbicide resistance are present; some people fear that super weed will be created by crosspollination between a genetic modification and wild (weedy) [92].
To avoid the above-mentioned risks, the screening of new relatively safe selection marker genes is an effective method to eliminate the potential risk of screening marker genes [6,9,93]. The use of fluorescent protein genes reduces the potential risks, to produce transgenic crops without any other sequences that are not desired in the final product [94]. Since its introduction in 1994 [95], green fluorescent protein has been utilized successfully as a marker in many species [17,19,74], and provided a more efficient direct visual screening method. This study constructed egfp gene instead of the bar gene as a transformation selectable marker. Transgenic positive callus was obtained by green fluorescence phenotype as a visual selection signal that allows the direct visualization of transformed cells. A total of 16.7% of the conversion efficiency was basically obtained from transformed callus. Results show that GFP would be useful for identifying transformed tissue, as we have shown that when GFP is synthesized and resides in the callus it accumulates or fluoresces and the transformed cells can be isolated easily and accurately in living callus. In addition, we found no interference with the subsequent growth of transformed maize calli in the selection stage. Second, the loss of green fluorescence in the heat-shock-treated calli served as a counter-selectable marker that effectively eliminates the recombination element and selection markers.
These new marker genes are regarded as less risky [40,96], the complete elimination of selectable marker gene has always been considered the most elegant way to overcome all the concerns [97]. Site-specific recombinant system is an important system to obtain marker elimination of transgenic plants [45], and Cre/lox recombinant system is the most widely used site-specific recombinant system [33]. Retransformation, crossing, and auto-excision are three successful application strategies for using the Cre/lox system to remove marker genes from transgenic plants [3]. The goal of our strategy is to obtain transformation and marker auto-excision in a single process using an induced expression of site-specific recombination system. By employing the well-known heat-inducible Cre/lox system (based on the use of the maize heat-shock hsp70 gene promoter described), it was easy to generate marker-free first-generation (T0) transgenic plants because sexual crosses are not required for selectable marker gene elimination. These plants obtain marker-free transformation before regeneration, through complete control and detection, and can transmit the target gene expression to the next generation stably, which improves the efficiency of safe transgenic plants and optimizes the operation process [65].
Although this is not the first reported use of Cre/lox system for marker-free transgenic maize operations, our strategy and vector system have their own advantages [3]. In Zhang’s reports [3], the researchers transformed the recombinase gene cre and selection marker gene with lox sites in maize, separately. Then the Cre/lox system activated in the F1 hybridization and marker-free transgenic maize were separated in backcross material. We constructed a heat-inducible auto-excision vector using a heat-shock-responsive Hsp70 promoter [74,75,76,77]; compared to previously reported systems, this new system is tightly controlled and the induced DNA excision is highly efficient. It is well known that the preparation and tracking of samples is time-consuming when screening large numbers of transformed molecules, and even worse, the false positive cells can hardly be identified from the tissue with the elimination of markers [21]. Compared with Vega’s work [73], we used green fluorescent protein gene (egfp) as selection marker to screen transgenic callus. Compared with the existing herbicide resistance markers, this marker has the advantages of convenient detection and no harm to living cells in the screening process. In addition, the visible fluorescence phenotype can effectively identify the elimination of selection marker genes [18]. In the present study, marker-free transgenic maize plants were obtained via recombination-programmed auto-excision without any extra handling borrowed from the traditional transformation process with a few modifications by the vector. The same mechanism has been successfully used for making marker-free hybrid aspen [48], potato [64], and rice [65].
5. Conclusions
In conclusion, an efficient method to combine the advantages of precise genetic engineering with marker gene excision into a single platform is proposed. The introduction of the visual selection markers egfp significantly improves the filtering accuracy and simplifies the operation.
Author Contributions
Conceptualization, D.D. and F.Z.; formal analysis, D.D.; funding acquisition, F.Z.; methodology, R.J.; resources, D.D. and J.G.; writing—review and editing, D.D. and F.Z.
Funding
This research was funded by National Science and Technology Major Project, grant number 2016ZX08003003 and The National Natural Science Foundation of China, grant number 31271737.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bruce, B.R.; Petke, J.; Harman, M. Reducing energy consumption using genetic improvement. In Proceedings of the 2015 Annual Conference on Genetic and Evolutionary Computation ACM, Madrid, Spain, 11–15 July 2015; pp. 1327–1334. [Google Scholar]
- Zhang, Q.; Walawage, S.L.; Tricoli, D.M.; Dandekar, A.M.; Leslie, C.A. A red fluorescent protein (DsRED) from Discosoma sp. as a reporter for gene expression in walnut somatic embryos. Plant Cell Rep. 2015, 34, 861–869. [Google Scholar] [CrossRef]
- Zhang, W.; Subbarao, S.; Addae, P.; Shen, A.; Armstrong, C.; Peschke, V.; Gilbertson, L. Cre/lox-mediated marker gene excision in transgenic maize (Zea mays L.) plants. Theor. Appl. Genet. 2003, 107, 1157–1168. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.K.; Perales, M.; Gruel, J.; Ohno, C.; Heisler, M.; Girke, T.; Jonsson, H.; Reddy, C.V. Plant stem cell maintenance involves direct transcriptional repression of differentiation program. Mol. Syst. Biol. 2013, 9, 1–13. [Google Scholar] [CrossRef]
- Qi, W.; Tinnenbroek-Capel, I.E.; Salentijn, E.M.; Schaart, J.G.; Cheng, J.; Denneboom, C.; Zhang, Z.; Zhang, X.; Zhao, H.; Visser, R.G.; et al. Screening for recombinants of Crambe Abyssinica after transformation by the pMF1 marker-free vector based on chemical selection and meristematic regeneration. Sci. Rep. 2015, 5, 14033. [Google Scholar] [CrossRef]
- Sundar, I.K.; Sakthivel, N. Advances in selectable marker genes for plant transformation. Plant Physiol. 2008, 165, 1698–1716. [Google Scholar] [CrossRef] [PubMed]
- Rosellini, D. Selectable markers and reporter genes: A well furnished toolbox for plant science and genetic engineering. Crit. Rev. Plant Sci. 2012, 31, 401–453. [Google Scholar] [CrossRef]
- Altpeter, F.; Springer, N.M.; Bartley, L.E.; Blechl, A.E.; Brutnell, T.P.; Citovsky, V.; Conrad, L.J.; Gelvin, S.B.; Jackson, D.P.; Kausch, A.P.; et al. Advancing crop transformation in the era of genome editing. Plant Cell 2016, 28, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Miki, B.; McHugh, S. Selectable marker genes in transgenic plants: Applications, alternatives and biosafety. J. Biotechnol. 2004, 107, 193–232. [Google Scholar] [CrossRef]
- Yau, Y.Y.; Stewart, C.N., Jr. Less is more: Strategies to remove marker genes from transgenic plants. BMC Biotechnol. 2013, 13, 36. [Google Scholar] [CrossRef]
- Perl, A.; Galili, S.; Shaul, O.; Ben-Tzvi, I.; Galili, G. Bacterial dihydrodipicolinate synthase and desensitized aspartate kinase: Two novel selectable markers for plant transformation. Bio Technol. 1993, 11, 715. [Google Scholar] [CrossRef]
- Ebmeier, A.; Allison, L.; Cerutti, H.; Clemente, T. Evaluation of the Escherichia coli threonine deaminase gene as a selectable marker for plant transformation. Planta 2004, 218, 751–758. [Google Scholar] [Green Version]
- Endo, S.; Kasahara, T.; Sugita, K.; Matsunaga, E.; Ebinuma, H. The isopentenyl transferase gene is effective as a selectable marker gene for plant transformation in tobacco (Nicotiana tabacum cv. Petite Havana SRI). Plant Cell Rep. 2001, 20, 60–66. [Google Scholar] [CrossRef]
- Roef, L.; Van Onckelen, H. Cytokinin regulation of the cell division cycle. In Plant Hormones; Springer: Dordrecht, The Netherlands, 2010; pp. 241–261. [Google Scholar]
- Wang, T.; Liu, L.Y.; Tang, Y.Y.; Zhang, X.B.; Zhang, M.D.; Zheng, Y.L.; Zhang, F.D. Using the Phosphomannose Isomerase (PMI) gene from saccharomyces cerevisiae for selection in rice transformation. J. Integr. Agric. 2012, 11, 1391–1398. [Google Scholar] [CrossRef]
- Chen, J.Q.; Meng, X.P.; Zhang, Y.; Xia, M.; Wang, X.P. Over-expression of OsDREB genes lead to enhanced drought tolerance in rice. Biotechnol. Lett. 2008, 30, 2191–2198. [Google Scholar] [CrossRef] [PubMed]
- Jordan, M.C. Green fluorescent protein as a visual marker for wheat transformation. Plant Cell Rep. 2000, 19, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Vain, P.; Worland, B.; Kohli, A.; Snape, J.W.; Christou, P. The green fluorescent protein (GFP) as a vital screenable marker in rice transformation. Theor. Appl. Genet. 2000, 96, 164–169. [Google Scholar] [CrossRef]
- Carlson, A.; Letarte, J.; Chen, J.; Kasha, K. Visual screening of microspore derived transgenic barley (Horde-cum vulgare L.) with green fluorescent protein. Plant Cell Rep. 2001, 20, 331–337. [Google Scholar] [CrossRef]
- Zuo, J.; Niu, Q.W.; Møller, S.G.; Chua, N.H. Chemical-regulated, site-specific DNA excision in transgenic plants. Nat. Biotechnol. 2001, 19, 157–161. [Google Scholar] [CrossRef]
- Lowe, K.; Wu, E.; Wang, N.; Hoerster, G.; Hastings, C.; Cho, M.J.; Scelonge, C.; Lenderts, B.; Chamberlin, M.; Cushatt, J.; et al. Morphogenic regulators Baby boom and Wuschel improve monocot transformation. Plant Cell 2016, 28, 1998–2015. [Google Scholar] [CrossRef]
- Darbani, B.; Eimanifar, A.; Stewart, C.N., Jr.; Camargo, W.N. Methods to produce marker-free transgenic plants. Biotechnol. J. 2007, 2, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goossens, H.; Ferech, M.; Vander, S.R.; Elseviers, M.; ESAC Project Group. Outpatient antibiotic use in Europe and association with resistance: A cross-national database study. Lancet 2005, 365, 579–587. [Google Scholar] [CrossRef]
- Depicker, A.; Herman, L.; Jacobs, A.; Schell, J.; Van, M.M. Frequencies of simultaneous transformation with different T-DNAs and their relevance to the Agrobacterium/plant cell interaction. Mol. Gen. Genet. 1985, 201, 477–484. [Google Scholar] [CrossRef]
- Puchta, H. Marker-free transgenic plants. Plant Cell Tissue Organ Culture 2003, 74, 123–134. [Google Scholar] [CrossRef]
- Wang, Y.; Yau, Y.Y.; Perkins-Balding, D.; Thomson, J.G. Recombinase technology: Applications and possibilities. Plant Cell Rep. 2011, 30, 267–285. [Google Scholar] [CrossRef]
- Kapusi, E.; Hensel, G.; Coronado, M.J.; Broeders, S.; Marthe, C.; Otto, I.; Kumlehn, J. The elimination of a selectable marker gene in the doubled haploid progeny of co-transformed barley plants. Plant. Mol. Biol. 2013, 81, 149–160. [Google Scholar] [CrossRef]
- Dalla Costa, L.; Piazza, S.; Campa, M.; Flachowsky, H.; Hanke, M.V.; Malnoy, M. Efficient heat-shock removal of the selectable marker gene in genetically modified grapevine. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 124, 471–481. [Google Scholar] [CrossRef]
- Oliva, N.; Chadha-Mohanty, P.; Poletti, S.; Abrigo, E.; Atienza, G.; Torrizo, L.; Garcia, R.; Dueñas, C., Jr.; Poncio, M.A.; Balindong, J.; et al. Large-scale production and evaluation of marker-free indica rice IR64 expressing phytoferritin genes. Mol. Breed. 2014, 33, 23–37. [Google Scholar] [CrossRef]
- Hare, P.D.; Chua, N.H. Excision of selectable marker genes from transgenic plants. Nat. Biotechnol. 2002, 20, 575–580. [Google Scholar] [CrossRef]
- Maeser, S.; Kahmann, R. The gin recombinase of phage Mu can catalyse site-specific recombination in plant protoplasts. Mol. Gen. Genet. 1991, 230, 170–176. [Google Scholar] [CrossRef]
- Gao, X.; Zhou, J.; Li, J.; Zou, X.; Zhao, J.; Li, Q.; Xia, R.; Yang, R.F.; Wang, D.K.; Zuo, Z.X.; et al. Efficient generation of marker-free transgenic rice plants using an improved transposon-mediated transgene reintegration strategy. Plant. Physiol. 2015, 167, 11–24. [Google Scholar] [CrossRef]
- Cotsaftis, O.; Sallaud, C.; Breitler, J.C.; Meynard, D.; Greco, R.; Pereira, A.; Guiderdoni, E. Transposon-mediated generation of T-DNA- and marker-free rice plants expressing a Bt endotoxin gene. Mol. Breed. 2002, 10, 165–180. [Google Scholar] [CrossRef]
- Lichtenstein, C.P.J.H.B.; Pazkowski, J.; Hohn, B. Intrachromosomal recombination between genomic repeats. In Homologous Recombination and Gene Silencing in Plants; Paszkowski, J., Ed.; Kluwer Publishing House: Dordrecht, The Netherlands, 1994; pp. 95–122. [Google Scholar]
- Puchta, H. Removing selectable marker genes: Taking the shortcut. Trends Plant Sci. 2000, 5, 273–274. [Google Scholar] [CrossRef]
- Zubko, E.; Scutt, C.; Meyer, P. Intrachromosomal recombination between attP regions as a tool to remove selectable marker genes from tobacco transgenes. Nat. Biotechnol. 2000, 18, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Dale, E.C.; Ow, D.W. Intra- and intermolecular site-specific recombination in plant cells mediated by bacteriophage P1 recombinase. Gene 1990, 91, 79–85. [Google Scholar] [CrossRef]
- Van, D.G.D. A structural view of cre-loxP site-specific recombination. Annu. Rev. Biophys. Biomol. Struct. 2001, 30, 87–104. [Google Scholar]
- Gidoni, D.; Srivastava, V.; Carmi, N. Site-specific excisional recombination strategies for elimination of undesirable transgenes from crop plants. In Vitro Cell. Dev. Biol. Plant 2008, 44, 457–467. [Google Scholar] [CrossRef]
- Tuteja, N.; Verma, S.; Sahoo, R.K.; Raveendar, S.; Reddy, I.N. Recent advances in development of marker-free transgenic plants: Regulation and biosafety concern. J. Biosci. 2012, 37, 167–197. [Google Scholar] [CrossRef] [PubMed]
- Louwerse, J.D.; Van, L.M.C.; van der, S.D.M.; de Vlaam, C.M.; Hooykaas, P.J.; Vergunst, A.C. Stable recombinase-mediated cassette exchange in Arabidopsis using Agrobacterium tumefaciens. Plant Physiol. 2007, 145, 1282–1293. [Google Scholar] [CrossRef]
- Chong-Pérez, B.; Kosky, R.G.; Reyes, M.; Rojas, L.; Ocaña, B.; Tejeda, M. Heat shock induced excision of selectable marker genes in transgenic banana by the Cre-lox site-specific recombination system. J. Biotechnol. 2012, 159, 265–273. [Google Scholar] [CrossRef]
- Lyznik, L.A.; Hirayama, L.; Rao, K.V.; Abad, A.; Hodges, T.K. Heat-inducible expression of FLP gene in maize cells. Plant J. 1995, 8, 177–186. [Google Scholar] [CrossRef]
- Nandy, S.; Srivastava, V. Site-specific gene integration in rice genome mediated by the FLP-FRT recombination system. Plant Biotechnol. J. 2011, 9, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Onouchi, H.; Yokoi, K.; Machida, C.; Matsuzaki, H.; Oshima, Y.; Matsuoka, K.; Nakamura, K.; Machida, Y. Operation of an efficient site-specific recombination system of Zygosaccharomyces rouxii in tobacco cells. Nucl. Acids Res. 1991, 19, 6373–6378. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.S.; Abercrombie, L.L.; Eda, S.; Blanvillain, R.; Thomson, J.G.; Ow, D.W.; Stewart, C.N., Jr. Transgene excision in pollen using a codon optimized serine resolvase CinH-RS2 site-specific recombination system. Plant Mol. Biol. 2001, 75, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Odell, J.; Caimi, P.; Sauer, B.; Russell, S. Site-directed recombination in the genome of transgenic tobacco. Mol. Gen. Genet. 1990, 223, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Fladung, M.; Becker, D. Targeted integration and removal of transgenes in hybrid aspen (Populus tremula L. × P. tremuloides Michx.) using site-specific recombination systems. Plant. Biol. 2010, 12, 334–340. [Google Scholar] [CrossRef]
- Gilbertson, L. Cre-lox recombination: Cre-ative tools for plant biotechnology. Trends Biotechnol. 2003, 21, 550–555. [Google Scholar] [CrossRef]
- Fladung, M.; Schenk, T.M.H.; Polak, O.; Becker, D. Elimination of marker genes and targeted integration via FLP ⁄ FRT recombination system from yeast in hybrid aspen (Populus tremula L. × P. tremuloides Michx.). Tree Genet. Gen. 2010, 6, 205–217. [Google Scholar] [CrossRef]
- Thomson, J.G.; Chan, R.; Smith, J.; Thilmony, R.; Yau, Y.Y.; Wang, Y.; Ow, D.W. The Bxb1 recombination system demonstrates heritable transmission of site-specific excision in Arabidopsis. BMC Biotechnol. 2012, 12, 9. [Google Scholar] [CrossRef]
- Zhou, Y.; Yau, Y.Y.; Ow, D.W.; Wang, Y. Site-specific deletions in the tomato genome by the CinH-RS2 and ParA-MRS recombination systems. Plant Biotechnol. Rep. 2012, 6, 225–232. [Google Scholar] [CrossRef]
- Verweire, D.; Verleyen, K.; De, B.S.; Claeys, M.; Angenon, G. Marker-free transgenic plants through genetically programmed auto-excision. Plant Physiol. 2007, 145, 1220–1231. [Google Scholar] [CrossRef]
- Russell, S.H.; Hoopes, J.L.; Odell, J.T. Directed excision of a transgene from the plant genome. Mol. Gen. Genet. 1992, 234, 49–59. [Google Scholar]
- Mészáros, K.; Éva, C.; Kiss, T.; Bányai, J.; Kiss, E.; Téglás, F.; LángIldikó, L.; Tamás, K.L. Generating marker-free transgenic wheat using minimal gene cassette and cold-inducible Cre/Lox system. Plant Mol. Biol. Rep. 2015, 33, 1221–1231. [Google Scholar] [CrossRef]
- Hoff, T.; Schnorr, K.M.; Mundy, J. A recombinase-mediated transcriptional induction system in transgenic plants. Plant. Mol. Biol. 2001, 45, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Bayley, C.C.; Morgan, M.; Dale, E.C.; Ow, D.W. Exchange of gene activity in transgenic plants catalyzed by the Cre-lox site-specific recombination system. Plant Mol. Biol. 1992, 18, 353–361. [Google Scholar] [CrossRef]
- Hoa, T.T.C.; Bong, B.B.; Huq, E.; Hodges, T.K. Cre/lox site-specific recombination controls the excision of a transgene from the rice genome. Theor. Appl. Genet. 2002, 104, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Dale, E.C.; Ow, D.W. Gene transfer with subsequent removal of the selection gene from the host genome. Proc. Natl. Acad. Sci. USA 1991, 88, 10558–10562. [Google Scholar] [CrossRef]
- Zhao, Z.; Gu, W.; Cai, T.; Tagliani, L.; Hondred, D.; Bond, D.; Schroeder, S.; Rudert, M.; Pierce, D. High throughput genetic transformation mediated by Agrobacterium tumefaciens in maize. Mol. Breed. 2002, 8, 323–333. [Google Scholar] [CrossRef]
- Gleave, A.P.; Mitra, D.S.; Mudge, S.R.; Morris, B.A. Selectable marker-free transgenic plants without sexual crossing: Transient expression of cre recombinase and use of a conditional lethal dominant gene. Plant. Mol. Biol. 1999, 40, 223–235. [Google Scholar] [CrossRef]
- Hoff, P.M.; Ansari, R.; Batist, G.; Cox, J.; Kocha, W.; Kuperminc, M.; Burger, H.U. Comparison of oral capecitabine versus intravenous fluorouracil plus leucovorin as first-line treatment in 605 patients with metastatic colorectal cancer: Results of a randomized phase III study. J. Clin. Oncol. 2001, 19, 2282–2292. [Google Scholar] [CrossRef]
- Zuo, J.; Niu, Q.W.; Ikeda, Y.; Chua, N.H. Marker-free transformation: Increasing transformation frequency by the use of regeneration-promoting genes. Plant. Biotechnol. 2002, 13, 121–124. [Google Scholar] [CrossRef]
- Cuellar, W.; Gaudin, A.; Solórzano, D.; Casas, A.; Ñopo, L.; Chudalayandi, P.; Giuliana, M.; Jan, K.; Ghislain, M. Self-excision of the antibiotic resistance gene nptII using a heat inducible Cre-loxP system from transgenic potato. Plant Mol. Biol. 2006, 62, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Khattri, A.; Nandy, S.; Srivastava, V. Heat-inducible Cre-lox system for marker excision in transgenic rice. J. Biosci. 2011, 36, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Herzog, K.; Flachowsky, H.; Deising, H.B.; Hanke, M.V. Heat-shock-mediated elimination of the nptII marker gene in transgenic apple (Malus × domestica Borkh.). Gene 2012, 498, 41–49. [Google Scholar] [CrossRef]
- Nishihama, R.; Ishida, S.; Urawa, H.; Kamei, Y.; Kohchi, T. Conditional gene expression/deletion systems for Marchantia polymorpha using its own heat-shock promoter and Cre/lox P-mediated site-specific recombination. Plant Cell Physiol. 2015, 57, 271–280. [Google Scholar] [CrossRef]
- McKnight, T.D.; Lillis, M.T.; Simpson, R.B. Segregation of genes transferred to one plant cell from two separate Agrobacterium strains. Plant Mol. Biol. 1987, 8, 439–445. [Google Scholar] [CrossRef]
- Komari, T.; Hiei, Y.; Saito, Y.; Murai, N.; Kumashiro, T. Vectors carrying two separate T-DNAs for co-transformation of higher plants mediated by Agrobacterium tumefaciens and segregation of transformants free from selection markers. Plant J. 1996, 10, 165–174. [Google Scholar] [CrossRef]
- Daley, M.; Knauf, V.C.; Summerfelt, K.R.; Turner, J.C. Co-transformation with one Agrobacterium tumefaciens strain containing two binary plasmids as a method for producing marker-free transgenic plants. Plant Cell Rep. 1998, 17, 489–496. [Google Scholar] [CrossRef]
- Miller, M.; Tagliani, L.; Wang, N.; Berka, B.; Bidney, D.; Zhao, Z.Y. High efficiency transgene segregation in co-transformed maize plants using an Agrobacterium tumefaciens 2 T-DNA binary system. Transgen. Res. 2002, 11, 381–396. [Google Scholar] [CrossRef]
- Xiao, Y.L.; Li, X.; Peterson, T. Ac insertion site affects the frequency of transposon-induced homologous recombination at the maize p1 locus. Genetics 2000, 156, 2007–2017. [Google Scholar]
- Vega, J.M.; Yu, W.; Han, F.; Kato, A.; Peters, E.M.; Zhang, Z.J.; Birchler, J.A. Agrobacterium-mediated transformation of maize (Zea mays L.) with Cre-lox site specific recombination cassettes in BIBAC vectors. Plant Mol. Biol. 2008, 66, 587–598. [Google Scholar] [CrossRef]
- Sung, D.Y.; Vierling, E.; Guy, C.L. Comprehensive expression profile analysis of the Arabidopsis Hsp70 gene family. Plant Physiol. 2001, 126, 789–800. [Google Scholar] [CrossRef]
- Spena, A.; Haun, R.; Ziervogel, U.; Saedle, H.; Schell, J. Construction of a heat-inducible gene for plants: Demostration of heat-inducible activity of the Drosophila hsp70 promoter in plants. EMBO J. 1985, 4, 2739–2743. [Google Scholar] [CrossRef]
- Rochester, D.E.; Winer, J.A.; Shah, D.M. The structure and expression of maize genes encoding the major heat shock protein, hsp70. EMBO J. 1986, 5, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.T.; Li, B.; Zhou, G.M.; Tang, W.Q.; Bai, J.; Sun, D.Y.; Zhou, R.G. Binding of the maize cytosolic Hsp70 to calmodulin, and identification of calmodulin-binding site in Hsp70. Plant Cell Physiol. 2000, 41, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Hirasawa, T.; Ohsawa, K.; Imai, Y.; Ondo, Y.; Akazawa, C.; Uchino, S.; Kohsaka, S. Visualization of microglia in living tissues using Iba1-EGFP transgenic mice. J. Neurosci. Res. 2005, 81, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Du, D.; Jin, R.C.; Guo, J.J.; Zhang, F.D. Infection of embryonic callus with Agrobacterium enables high-speed transformation of maize. Int. J. Mol. Sci. 2019, 20, 279. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, Y.; Zhao, J.; Dong, M.; Zhang, J. Heat-shock-induced removal of transgenes using the gene-deletor system in hybrid aspen (Populus tremula × P. tremuloides). Genes 2018, 9, 484. [Google Scholar] [CrossRef]
- Christensen, A.H.; Quail, P.H. Ubiquitin promoter-based vectors for high-level expression of selectable and/or screenable marker genes in monocotyledonous plants. Transgen. Res. 1996, 5, 213–218. [Google Scholar] [CrossRef]
- Tonelli, C.; Dolfini, S.; Ronchi, A.; Consonni, G.; Gavazzi, G. Light inducibility and tissue specificity of the R gene family in maize. Genetica 1994, 94, 225–234. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, B.; Hu, Y.; Li, J.; Lin, Z. Inducible excision of selectable marker gene from transgenic plants by the Cre⁄lox site-specific recombination system. Transgen. Res. 2005, 14, 605–614. [Google Scholar] [CrossRef]
- Du, D.X.; Geng, C.; Zhang, X.; Zhang, Z.; Zheng, Y.; Zhang, F.; Lin, Y.; Qiu, F. Transgenic maize lines expressing a cry1C* gene are resistant to insect pests. Plant. Mol. Biol. Rep. 2014, 32, 549–557. [Google Scholar] [CrossRef]
- Southern, E. Southern blotting. Nat. Protoc. 2006, 1, 518. [Google Scholar] [CrossRef]
- Nandy, S.; Srivastava, V. Marker-free site-specific gene integration in rice based on the use of two recombination systems. Plant Biotechnol. J. 2012, 10, 904–912. [Google Scholar] [CrossRef]
- Rakoczy-Trojanowska, M. Alternative methods of plant transformation—A short review. Cell Mol. Biol. Lett. 2002, 7, 849–858. [Google Scholar]
- Angenon, G.; Dillen, W.; Van, M.M. Antibiotic resistance markers for plant transformation. In Plant Molecular Biology Manual, 2nd ed.; Gelvin, S.B., Schilperoort, R.A., Eds.; Springer Science+Business Media: Dordrecht, The Netherlands, 1994; pp. 125–137. [Google Scholar]
- Bevan, M.W.; Flavell, R.B.; Chilton, M.D. A chimeric antibiotic resistance gene as a selectable marker for plant cell transformation. Biotechnology 1983, 24, 367–370. [Google Scholar]
- Frame, B.R.; McMurray, J.M.; Fonger, T.M.; Main, M.L.; Taylor, K.W.; Torney, F.J.; Paz, M.M.; Wang, K. Improved Agrobacterium-mediated transformation of three maize inbred lines using MS salts. Plant Cell Rep. 2006, 25, 1024–1034. [Google Scholar] [CrossRef]
- Allen, H.K.; Donato, J.; Wang, H.H.; Cloud-Hansen, K.A.; Davies, J.; Handelsman, J. Call of the wild: Antibiotic resistance genes in natural environments. Nat. Rev. Microbiol. 2010, 8, 251–259. [Google Scholar] [CrossRef]
- Dale, P.J.; Clarke, B.; Fontes, E.M. Potential for the environmental impact of transgenic crops. Nat. Biotechnol. 2002, 20, 567–574. [Google Scholar] [CrossRef]
- Wright, T.R.; Shan, G.; Walsh, T.A.; Lira, J.M.; Cui, C.; Song, P.; Russell, S.M. Robust crop resistance to broadleaf and grass herbicides provided by aryloxyalkanoate dioxygenase transgenes. Proc. Natl. Acad. Sci. USA 2010, 107, 20240–20245. [Google Scholar] [CrossRef]
- Stewart, C.N. Monitoring transgenic plants using in vivo markers. Nat. Biotechnol. 1996, 14, 682. [Google Scholar] [CrossRef]
- Chalfie, M.; Tu, Y.; Euskirchen, G.; Ward, W.W.; Prasher, D.C. Green fluorescent protein as a marker for gene expression. Science 1994, 263, 802–805. [Google Scholar] [CrossRef] [PubMed]
- Sreekala, C.; Wu, L.; Gu, K.; Wang, D.; Tian, D.; Yin, Z. Excision of a selectable marker in transgenic rice (Oryza sativa L.) using a chemically regulated Cre/loxP system. Plant Cell Rep. 2005, 24, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Afolabi, A.S.; Worl, B.; Snape, J.; Vain, P. Novel pGreen/pSoup dual-binary vector system in multiple T-DNA co-cultivation as a method of producing marker-free (clean gene) transgenic rice (Oriza sativa L) plant. Afr. J. Biotechnol. 2005, 4, 531–540. [Google Scholar]
Figure 1.
Mechanism of the three approaches for the removal of selectable marker genes from transgenic plants [10]. (a) The mechanism of co-transformation to removal of selectable marker genes from transgenic plants. a1, Mixture method of Agrobacterium co-transformation. a2, One-strain method of Agrobacterium co-transformation. a3, Super binary vector for co-transformation. a4, Two pairs of right borders and left borders used for co-transformation; (b) The mechanism of transposon-mediated transgene reintegration to removal of selectable marker genes from transgenic plants. b1, T-DNA contains both the GOI (flanked by transposase-recognizing sites) and the transposase is transformed into plants. b2, T-DNA containing both the SMG (flanked by transposase recognizing inverted repeat sequences) and the GOI is transformed into plants; (c) The mechanism of site-specific recombination (SSR) to removal of selectable marker genes from transgenic plants.
Figure 1.
Mechanism of the three approaches for the removal of selectable marker genes from transgenic plants [10]. (a) The mechanism of co-transformation to removal of selectable marker genes from transgenic plants. a1, Mixture method of Agrobacterium co-transformation. a2, One-strain method of Agrobacterium co-transformation. a3, Super binary vector for co-transformation. a4, Two pairs of right borders and left borders used for co-transformation; (b) The mechanism of transposon-mediated transgene reintegration to removal of selectable marker genes from transgenic plants. b1, T-DNA contains both the GOI (flanked by transposase-recognizing sites) and the transposase is transformed into plants. b2, T-DNA containing both the SMG (flanked by transposase recognizing inverted repeat sequences) and the GOI is transformed into plants; (c) The mechanism of site-specific recombination (SSR) to removal of selectable marker genes from transgenic plants.

Figure 2.
Transfer DNA (T-DNA) organization of the binary plasmid. LB is left border; RB is right border. loxP is the recombination-site (lox); Phsp70::Cre is the Cre gene carrying a nuclear localization signal with the heat-shock-responsive promoter Hsp70; PCaMV35S::egfp is egfp gene as visual marker for selection driven by the CaMV35S (35S) promoter; Pubi::Rsc is a reporter gene anthocyanin pigmentation (Rsc), controlled by the maize polyubiquitin gene (ubi) promoter.
Figure 2.
Transfer DNA (T-DNA) organization of the binary plasmid. LB is left border; RB is right border. loxP is the recombination-site (lox); Phsp70::Cre is the Cre gene carrying a nuclear localization signal with the heat-shock-responsive promoter Hsp70; PCaMV35S::egfp is egfp gene as visual marker for selection driven by the CaMV35S (35S) promoter; Pubi::Rsc is a reporter gene anthocyanin pigmentation (Rsc), controlled by the maize polyubiquitin gene (ubi) promoter.

Figure 3.
Determination of transformed callus. (a) The GFP phenotype of the transformation-positive calli on the DR-46B Dark Reader transilluminator at 3 weeks after infection, the non-fluorescing parts due to the callus being untransformed; (b) The PCR monitoring area and probe location in T-DNA organization of pHZM1-Rsc. Specific primer sites frame the T-DNA. P1 detected the selection marker egfp, P2 detected the regulator gene (Rsc), and P3 was used to detect the RS after the markers were eliminated. Probe 1 is on the gene Rsc, beside the restriction of endonuclease site Asc I, which is used as a marker for the southern blotting probe. (c) PCR analysis of the transformed maize calli. DNA was extracted from six independent transgenic GFP+ maize calli; specific primer pairs P1, P2, and P3 were used to detect the selection marker gene egfp, the regulator gene (Rsc), and the RS, respectively. M, BM2000 DNA marker; P, positive control P was used as the plasmid DNA; C, negative control; lanes no. 1–6, DNA from independent transgenic calli. (d) Southern blot analysis of the transferred gene in independent transgenic experiments. M, HindIII DNA Marker; P, positive control P was used as the plasmid DNA; lanes no 1–6, DNA from independent transgenic calli.
Figure 3.
Determination of transformed callus. (a) The GFP phenotype of the transformation-positive calli on the DR-46B Dark Reader transilluminator at 3 weeks after infection, the non-fluorescing parts due to the callus being untransformed; (b) The PCR monitoring area and probe location in T-DNA organization of pHZM1-Rsc. Specific primer sites frame the T-DNA. P1 detected the selection marker egfp, P2 detected the regulator gene (Rsc), and P3 was used to detect the RS after the markers were eliminated. Probe 1 is on the gene Rsc, beside the restriction of endonuclease site Asc I, which is used as a marker for the southern blotting probe. (c) PCR analysis of the transformed maize calli. DNA was extracted from six independent transgenic GFP+ maize calli; specific primer pairs P1, P2, and P3 were used to detect the selection marker gene egfp, the regulator gene (Rsc), and the RS, respectively. M, BM2000 DNA marker; P, positive control P was used as the plasmid DNA; C, negative control; lanes no. 1–6, DNA from independent transgenic calli. (d) Southern blot analysis of the transferred gene in independent transgenic experiments. M, HindIII DNA Marker; P, positive control P was used as the plasmid DNA; lanes no 1–6, DNA from independent transgenic calli.

Figure 4.
Induction of marker excision and molecular analysis of regenerated plants. (a) Design of DNA constructs and molecular strategy of marker-free site-specific gene integration. (b) PCR analysis of the transformed maize lines. Specific primer pairs P1, P2, and P3 to detect the selection marker gene egfp, the regulator gene (Rsc), and the residual sequence (RS), respectively. M, BM2000 DNA marker; P, positive control P was used as the plasmid DNA; C, negative control; lanes no. 1–4, DNA from independent transgenic transformants plants; (c) Rsc gene expression in genetically modified materials in T1 generation. Wild type ears appeared with yellow and white kernels, while the transformation ears with the Rsc gene expression phenotype showed a lot of purple kernels.
Figure 4.
Induction of marker excision and molecular analysis of regenerated plants. (a) Design of DNA constructs and molecular strategy of marker-free site-specific gene integration. (b) PCR analysis of the transformed maize lines. Specific primer pairs P1, P2, and P3 to detect the selection marker gene egfp, the regulator gene (Rsc), and the residual sequence (RS), respectively. M, BM2000 DNA marker; P, positive control P was used as the plasmid DNA; C, negative control; lanes no. 1–4, DNA from independent transgenic transformants plants; (c) Rsc gene expression in genetically modified materials in T1 generation. Wild type ears appeared with yellow and white kernels, while the transformation ears with the Rsc gene expression phenotype showed a lot of purple kernels.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Composition of media used in Agrobacterium-mediated transformations.
| Medium | Composition |
|---|---|
| LB (solid) | Yeast extract 5 g/L, NaCl 10 g/L, peptone 10 g/L, agar 15 g/L, pH 6.8 |
| LB (liquid) | Yeast extract 5 g/L, NaCl 10 g/L, peptone 10 g/L, pH 6.8 |
| Infection | N6 1 2 g/L, 2,4-D 1 2.0 mg/L, L-proline 0.7 g/L, sucrose 68.4 g/L, D-glucose 2 36 g/L, MES 1 0.5 g/L, myo-inositol 0.1 g/L, As 1,2 200 μM, pH 5.2 |
| Co-cultivation | N6 4 g/L, 2,4-D 2.0 mg/L, L-proline 0.7 g/L, sucrose 30 g/L, MES 0.5 g/L, myo-inositol 0.1 g/L, CuSO4 1,2 0.05 µM, DTT 1,2 1 M, L-cysteine 0.4 g/L, As 100 μM, agar 8 g/L, pH 5.8 |
| Resting | N6 4 g/L, 2,4-D 2.0 mg/L, L-proline 0.7 g/L, sucrose 30 g/L, MES 0.5 g/L, myo-inositol 0.1 g/L, AgNO3 1,2 0.85 mg/L, carbenicillin 1,2 0.1 g/L, gelrite 2.5 g/L, pH 5.8 |
| Selection | Resting medium without carbenicillin, pH 5.8 |
| Regeneration | MS 1 4.3 g/L, sucrose 30 g/L, myo-inositol 0.1 g/L, 6-BA 3.5 mg/L, gelrite 3.0 g/L, pH 5.8 |
| Rooting | MS 4.3 g/L, sucrose 25 g/L, NAA 0.5 mg/L, gelrite 2 g/L, pH 5.8 |
1 6-BA, 6-Benzylaminopurine; N6, Chu medium salt with N6 vitamins; 2,4-D, 2,4-dichlorophenoxyacetic acid; MES, 2-(N-morpholino) ethanesulfonic acid; As, acetosyringone; CuSO4, copper sulfate; DTT, dithiothreitol; AgNO3, silver nitrate; MS, MS basal salt and vitamins; NAA, 1-Naphthaleneacetic acid. 2 Components were filter sterilized.
Table 2.
GFP fluorescence assays and PCR analysis of the auto-excision marker in calli.
| Cultivar | First Heat Shock | |||
| GFP Fluorescence Assays | PCR Analysis of Non-Fluorescent Calli | |||
| Positive | Negative | gfp Detected (+/−) | rsc Detected (+/−) | |
| pHZM1N-Rsc-2 | 9 | 41 | 9/41 | 50/50 |
| pHZM1N-Rsc-5 | 3 | 47 | 3/47 | 50/50 |
| pHZM1N-Rsc-6 | 8 | 42 | 8/42 | 50/50 |
| pHZM1N-Rsc-7 | 7 | 43 | 7/43 | 50/50 |
| Cultivar | Second Heat Shock | |||
| GFP Fluorescence Assays | PCR Analysis of Non-Fluorescent Calli | |||
| Positive | Negative | gfp Detected (+/−) | rsc Detected (+/−) | |
| pHZM1N-Rsc-2 | 0 | 50 | 0/50 | 50/50 |
| pHZM1N-Rsc-5 | 0 | 50 | 0/50 | 50/50 |
| pHZM1N-Rsc-6 | 0 | 50 | 0/50 | 50/50 |
| pHZM1N-Rsc-7 | 0 | 50 | 0/50 | 50/50 |
Table 3.
PCR analysis of T1 transgenic maize plants.
| Cultivar | Plants 1 | PCR Analysis of T1 Transgenic Plants | |||
|---|---|---|---|---|---|
| P1 | P2 | P3 | rsc+gfp−RS+ | ||
| pHZM1N-Rsc-2 | 9 | 0 | 9 | 9 | 9 |
| pHZM1N-Rsc-5 | 5 | 0 | 5 | 5 | 5 |
| pHZM1N-Rsc-6 | 1 | 0 | 1 | 1 | 1 |
| pHZM1N-Rsc-7 | 2 | 0 | 2 | 2 | 2 |
1 Transformed seeds were randomly selected from each of the cultivar combinations for sowing, and their leaf samples were bulked for DNA purification. The bulked maize DNA was then amplified using primers to select gfp (P1), regulator-specific sequences (P2), and the RS resulting from marker-eliminated sequences (P3).
Table 4.
Segregation ratio analysis of T1 kernel.
| Cultivar | Plants | Purple Seeds | Yellow Seeds | Segregation Ratio | Consistency with 3:1 |
|---|---|---|---|---|---|
| pHZM1N-Rsc-2 | pHZM1N-Rsc-2-1 | 307 | 96 | 3.20:1 | Yes |
| pHZM1N-Rsc-2-2 | 218 | 75 | 2.91:1 | Yes | |
| pHZM1N-Rsc-2-3 | 114 | 36 | 3.17:1 | Yes | |
| pHZM1N-Rsc-2-5 | 222 | 68 | 3.26:1 | Yes | |
| pHZM1N-Rsc-2-7 | 256 | 80 | 3.20:1 | Yes | |
| pHZM1N-Rsc-2-8 | 187 | 71 | 2.63:1 | No | |
| pHZM1N-Rsc-2-5 | 299 | 113 | 2.65:1 | No | |
| pHZM1N-Rsc-2-16 | 196 | 69 | 2.84:1 | Yes | |
| pHZM1N-Rsc-2-17 | 185 | 51 | 3.63:1 | No | |
| pHZM1N-Rsc-5 | pHZM1N-Rsc-5-1 | 209 | 71 | 2.94:1 | Yes |
| pHZM1N-Rsc-5-2 | 179 | 55 | 3.25:1 | Yes | |
| pHZM1N-Rsc-5-3 | 315 | 107 | 2.94:1 | Yes | |
| pHZM1N-Rsc-5-4 | 122 | 41 | 2.96:1 | Yes | |
| pHZM1N-Rsc-5-5 | 237 | 64 | 3.70:1 | Yes | |
| pHZM1N-Rsc-6 | pHZM1N-Rsc-6-11 | 374 | 0 | No | |
| pHZM1N-Rsc-7 | pHZM1N-Rsc-7-3 | 191 | 67 | 2.85:1 | Yes |
| pHZM1N-Rsc-7-20 | 409 | 0 | No |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Du, D.; Jin, R.; Guo, J.; Zhang, F. Construction of Marker-Free Genetically Modified Maize Using a Heat-Inducible Auto-Excision Vector. Genes 2019, 10, 374. https://doi.org/10.3390/genes10050374
AMA Style
Du D, Jin R, Guo J, Zhang F. Construction of Marker-Free Genetically Modified Maize Using a Heat-Inducible Auto-Excision Vector. Genes. 2019; 10(5):374. https://doi.org/10.3390/genes10050374
Chicago/Turabian StyleDu, Dengxiang, Ruchang Jin, Jinjie Guo, and Fangdong Zhang. 2019. "Construction of Marker-Free Genetically Modified Maize Using a Heat-Inducible Auto-Excision Vector" Genes 10, no. 5: 374. https://doi.org/10.3390/genes10050374
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.