Biphenyl/PCB Degrading bph Genes of Ten Bacterial Strains Isolated from Biphenyl-Contaminated Soil in Kitakyushu, Japan: Comparative and Dynamic Features as Integrative Conjugative Elements (ICEs)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Cultivation
2.2. Genome Sequencing and Computational Analysis
2.3. Phylogenetic Analysis and Gene Alignment
2.4. Conjugation Experiments
2.5. DNA Manipulation
3. Results
3.1. KF Strains and their Genomic Features
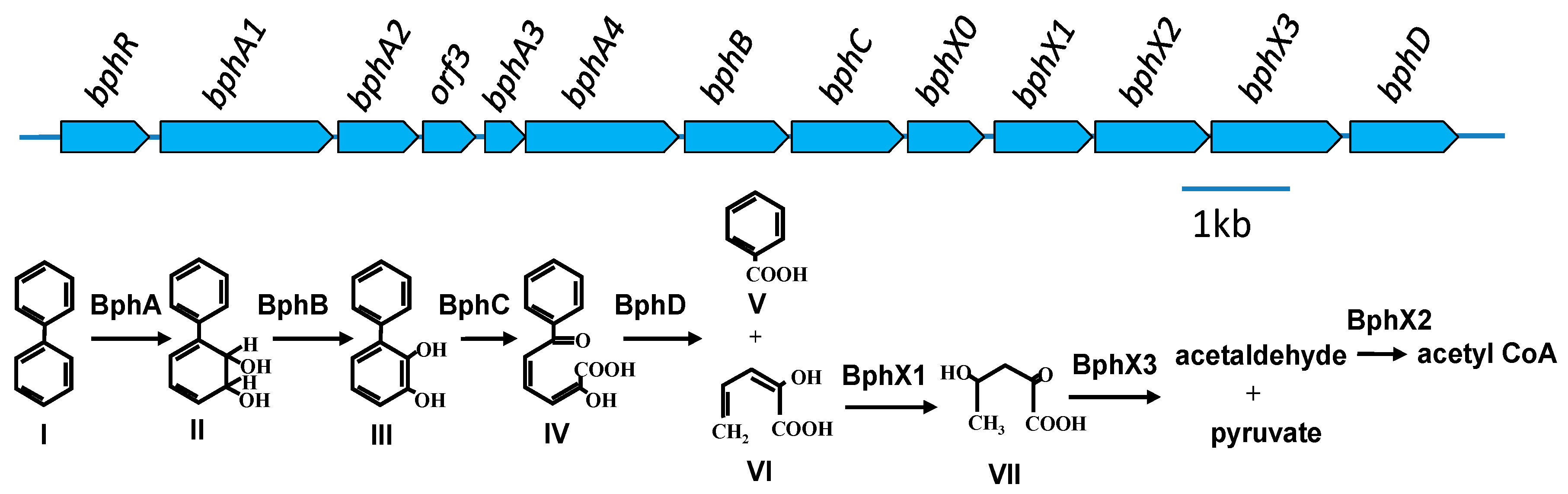
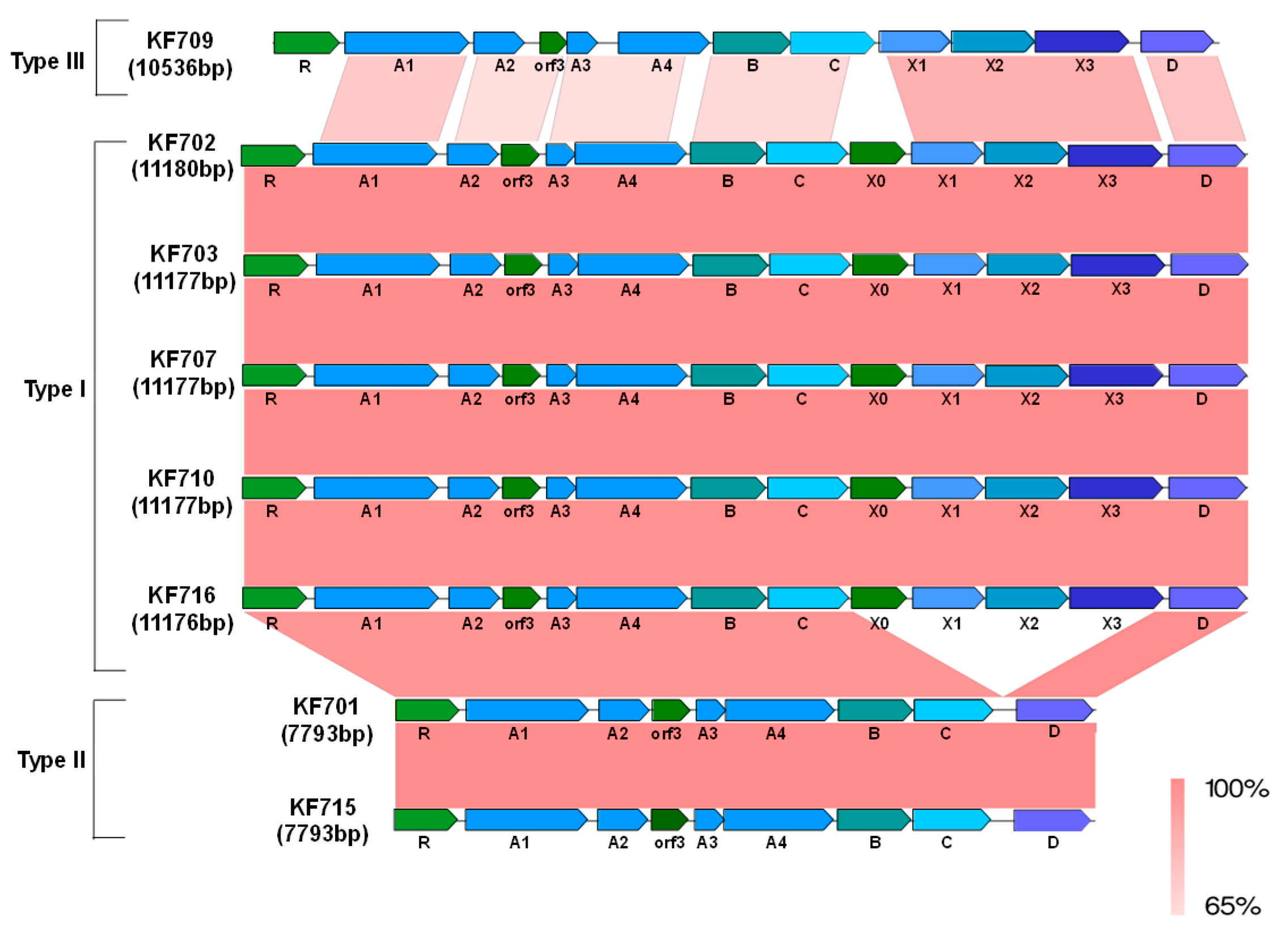
3.2. Comparison of the bph Genes
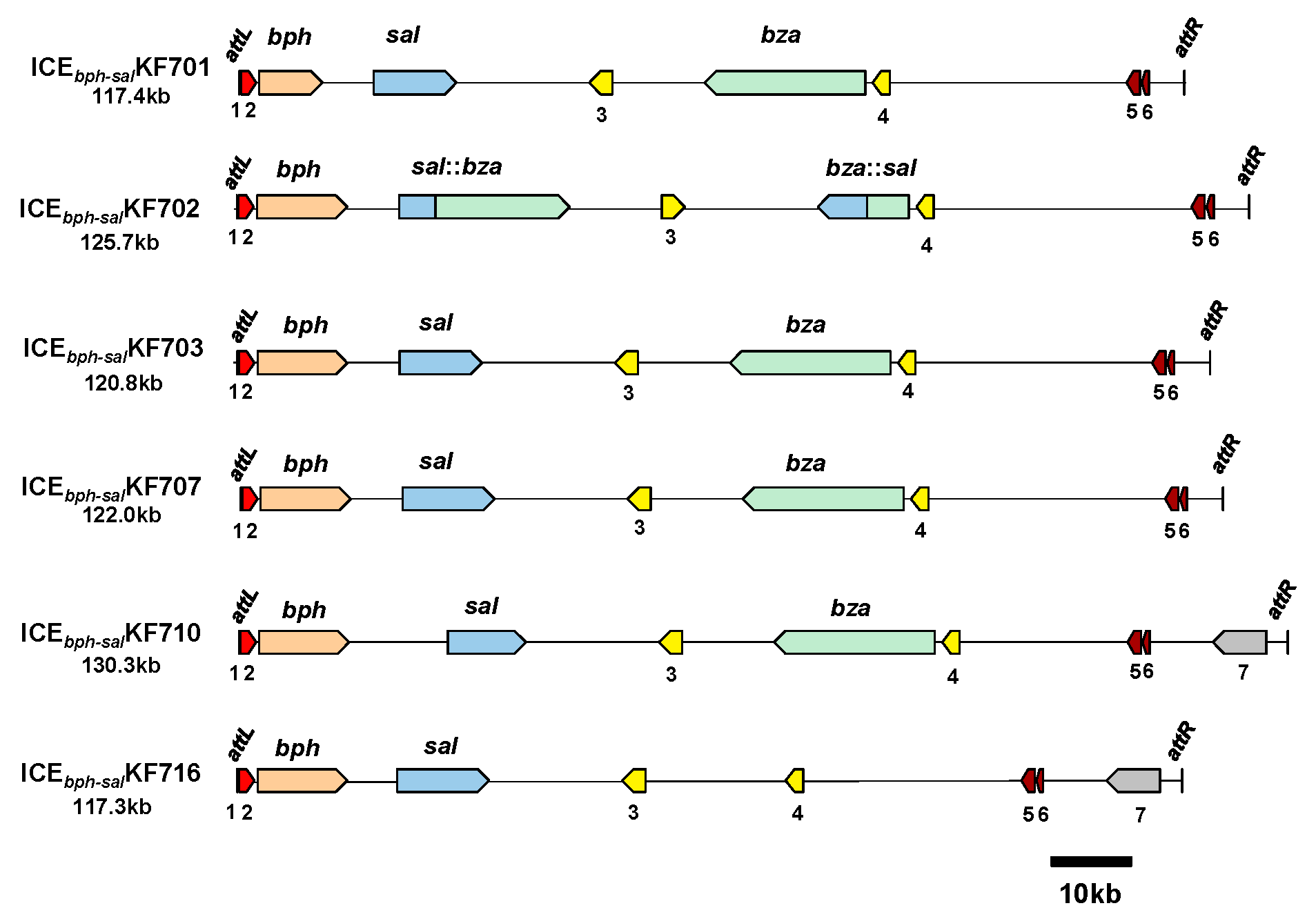
3.3. ICEbph-sal in KF Strains Belonging to Groups I and II
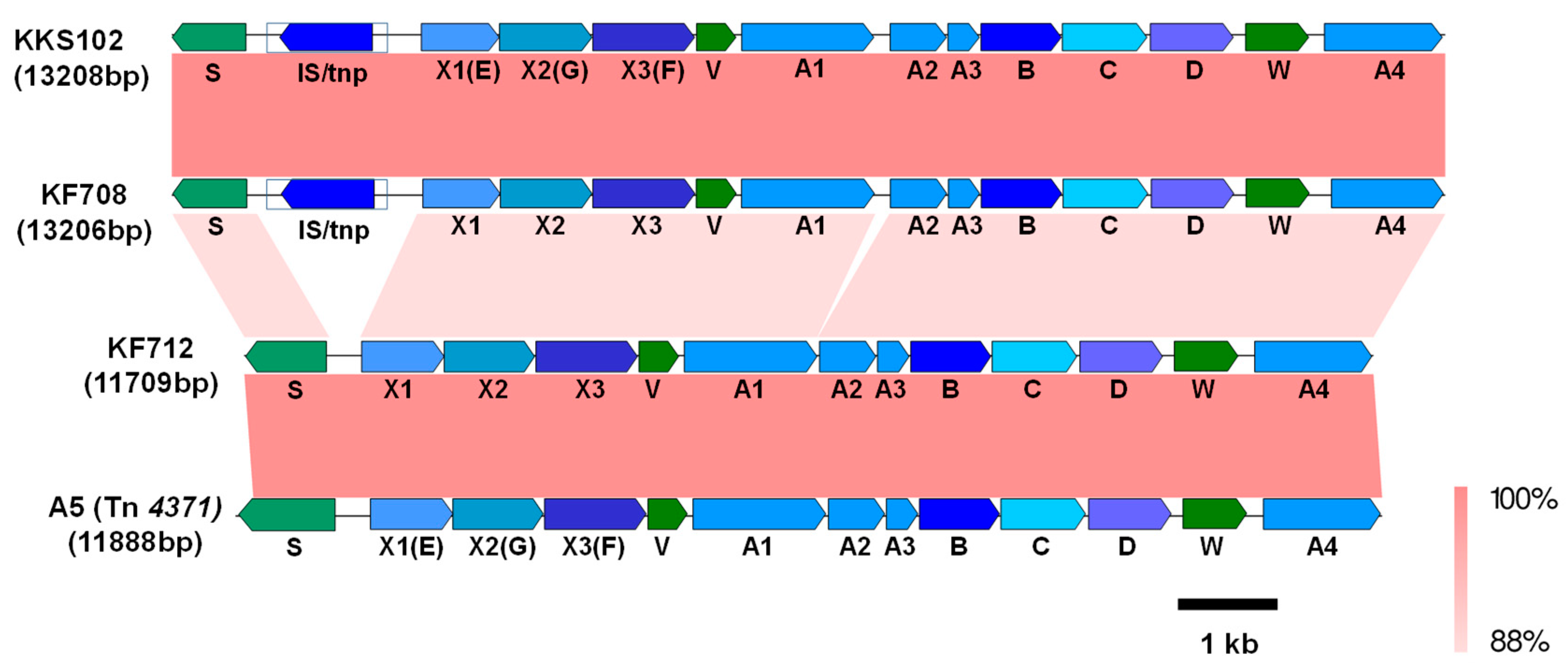
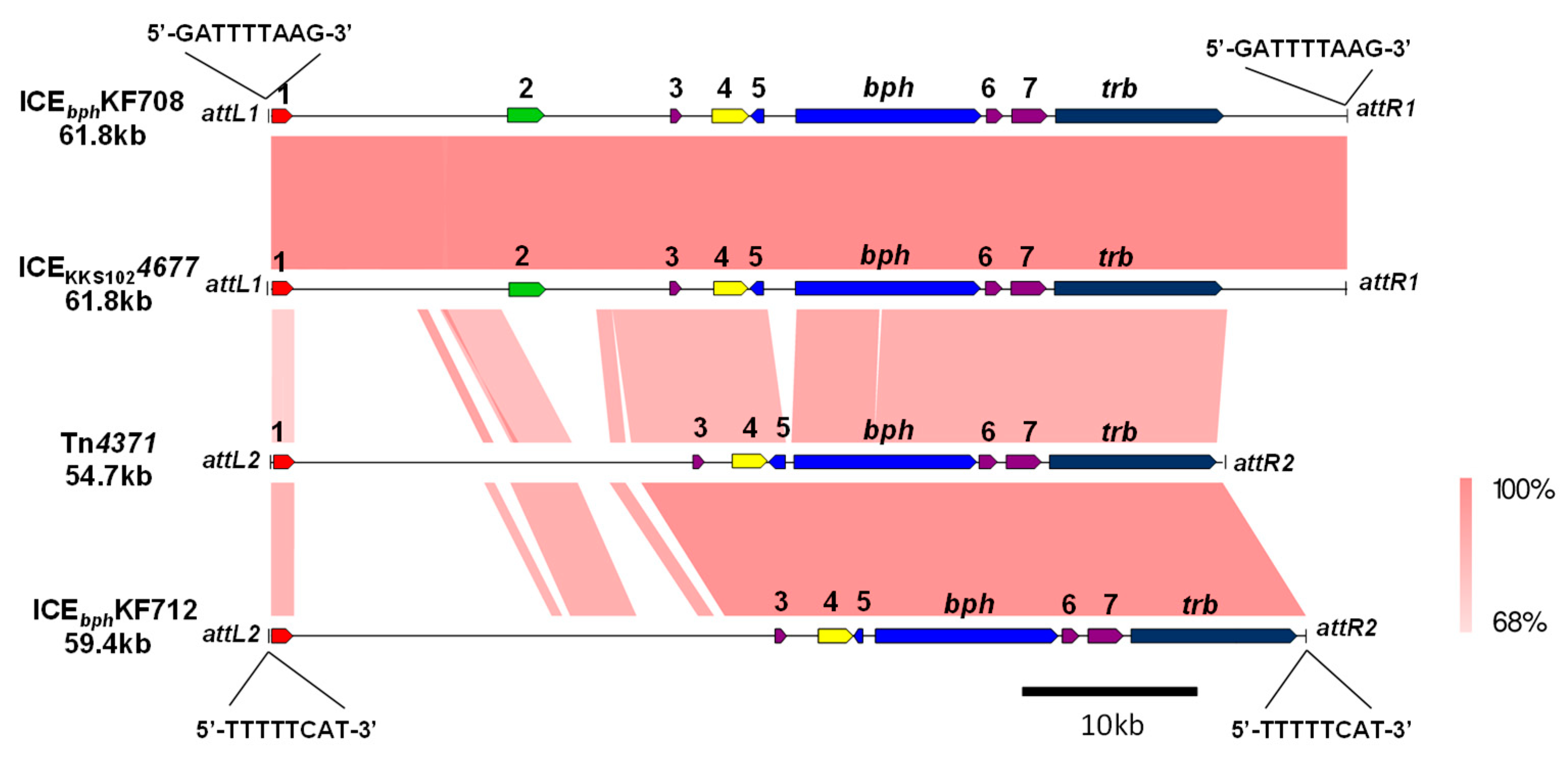
3.4. ICEbph in the Group IV Strains
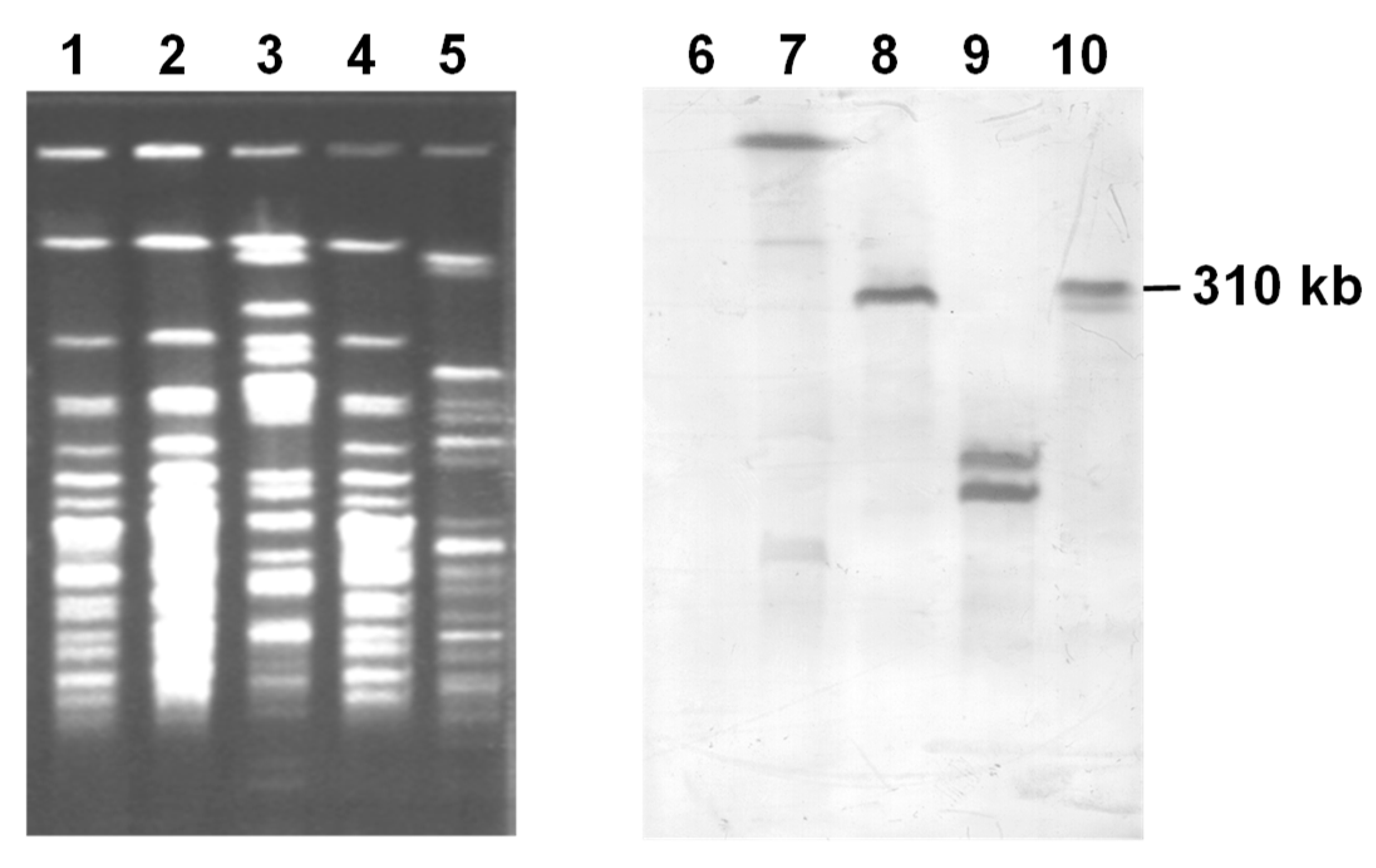
3.5. Conjugal Transfer of Extrachromosomal ICEbph-salKF715 Into P. putida F39/D
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Furukawa, K. Microbial degradation of polychlorinated biphenyls. In Biodegradation and Detoxification of Environmental Pollutants; Chakrabarty, A.M., Ed.; CRC Press: Boca Raton, FL, USA, 1982; pp. 33–57. [Google Scholar]
- Abramowicz, D.A. Aerobic and anaerobic biodegradation of PCBs: A review. Crit. Rev. Biotechnol. 1990, 10, 241–251. [Google Scholar] [CrossRef]
- Furukawa, K.; Fujihara, H. Microbial degradation of polychlorinated biphenyls: Biochemical and molecular features. J. Biosci. Bioeng. 2008, 105, 433–449. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Tonomura, K.; Kamibayashi, A. Effect of chlorine substitution on the biodegradability of polychlorinated biphenyls. Appl. Env. Microbiol. 1978, 35, 223–227. [Google Scholar]
- Furukawa, K.; Miyazaki, T. Cloning of a gene cluster encoding biphenyl and chlorobiphenyl degradation in Pseudomonas pseudoalcaligenes. J. Bacteriol. 1986, 166, 392–398. [Google Scholar] [CrossRef]
- Toussaint, A.; Merlin, C.; Monchy, S.; Benotmane, M.A.; Leplae, R.; Mergeay, M.; Springael, D. The biphenyl- and 4-chlorobiphenyl-catabolic transposon Tn4371, a member of a new family of genomic islands related to IncP and Ti plasmids. Appl. Env. Microbiol. 2003, 69, 4837–4845. [Google Scholar] [CrossRef] [PubMed]
- Ohtsubo, Y.; Ishibashi, Y.; Naganawa, H.; Hirokawa, S.; Atobe, S.; Nagata, Y.; Tsuda, M. Conjugal transfer of polychlorinated biphenyl/biphenyl degradation genes in Acidovorax sp. strain KKS102, which are located on an integrative and conjugative element. J. Bacteriol. 2012, 194, 4237–4248. [Google Scholar] [CrossRef]
- Masai, E.; Sugiyama, K.; Iwashita, N.; Shimizu, S.; Hauschild, J.E.; Hatta, T.; Kimbara, K.; Yano, K.; Fukuda, M. The bphDEF meta-cleavage pathway genes involved in biphenyl/polychlorinated biphenyl degradation are located on a linear plasmid and separated from the initial bphACB genes in Rhodococcus sp. strain RHA1. Gene 1997, 187, 141–149. [Google Scholar] [CrossRef]
- Shimizu, S.; Kobayashi, H.; Masai, E.; Fukuda, M. Characterization of the 450-kb linear plasmid in a polychlorinated biphenyl degrader, Rhodococcus sp. strain RHA1. Appl. Env. Microbiol. 2001, 67, 2021–2028. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Inoue, R.; Kimura, N.; Furukawa, K. Versatile transcription of biphenyl catabolic bph operon in Pseudomonas pseudoalcaligenes KF707. J. Biol. Chem. 2000, 275, 31016–31023. [Google Scholar] [CrossRef]
- Fujihara, H.; Yoshida, H.; Matsunaga, T.; Goto, M.; Furukawa, K. Cross-regulation of biphenyl- and salicylate-catabolic genes by two regulatory systems in Pseudomonas pseudoalcaligenes KF707. J. Bacteriol. 2006, 188, 4690–4697. [Google Scholar] [CrossRef]
- Kumamaru, T.; Suenaga, H.; Mitsuoka, M.; Watanabe, T.; Furukawa, K. Enhanced degradation of polychlorinated biphenyls by directed evolution of biphenyl dioxygenase. Nat. Biotechnol. 1998, 16, 663. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, H.; Goto, M.; Furukawa, K. Emergence of multifunctional oxygenase activities by random priming recombination. J. Biol. Chem. 2001, 276, 22500–22506. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, H.; Mitsuoka, M.; Ura, Y.; Watanabe, T.; Furukawa, K. Directed evolution of biphenyl dioxygenase: Emergence of enhanced degradation capacity for benzene, toluene, and alkylbenzenes. J. Bacteriol. 2001, 183, 5441–5444. [Google Scholar] [CrossRef]
- Furukawa, K.; Hayase, N.; Taira, K.; Tomizuka, N. Molecular relationship of chromosomal genes encoding biphenyl/polychlorinated biphenyl catabolism: Some soil bacteria possess a highly conserved bph operon. J. Bacteriol. 1989, 171, 5467–5472. [Google Scholar] [CrossRef]
- Suenaga, H.; Fujihara, H.; Kimura, N.; Hirose, J.; Watanabe, T.; Futagami, T.; Goto, M.; Shimodaira, J.; Furukawa, K. Insights into the genomic plasticity of Pseudomonas putida KF715, a strain with unique biphenyl-utilizing activity and genome instability properties. Environ. Microbiol. Rep. 2017, 9, 589–598. [Google Scholar] [CrossRef]
- Fujihara, H.; Yamazoe, A.; Hosoyama, A.; Suenaga, H.; Kimura, N.; Hirose, J.; Watanabe, T.; Futagami, T.; Goto, M.; Furukawa, K. Draft genome sequence of Pseudomonas abietaniphila KF701 (NBRC110664), a polychlorinated biphenyl-degrading bacterium isolated from biphenyl-contaminated soil. Genome Announc. 2015, 3, e00473-15. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, H.; Yamazoe, A.; Hosoyama, A.; Kimura, N.; Hirose, J.; Watanabe, T.; Fujihara, H.; Futagami, T.; Goto, M.; Furukawa, K. Draft genome sequence of the polychlorinated biphenyl-degrading bacterium Pseudomonas putida KF703 (NBRC 110666) isolated from biphenyl-contaminated soil. Genome Announc. 2015, 3, e00142-15. [Google Scholar] [CrossRef]
- Kimura, N.; Watanabe, T.; Suenaga, H.; Fujihara, H.; Futagami, T.; Goto, M.; Hanada, S.; Hirose, J. Pseudomonas furukawaii sp. nov., a polychlorinated biphenyl-degrading bacterium isolated from biphenyl-contaminated soil in Japan. Int. J. Syst. Evol. Microbiol. 2018, 68, 1429–1435. [Google Scholar] [CrossRef]
- Suenaga, H.; Yamazoe, A.; Hosoyama, A.; Kimura, N.; Hirose, J.; Watanabe, T.; Fujihara, H.; Futagami, T.; Goto, M.; Furukawa, K. Draft genome sequence of the polychlorinated biphenyl-degrading bacterium Cupriavidus basilensis KF708 (NBRC 110671) isolated from biphenyl-contaminated soil. Genome Announc. 2015, 3, e00143-15. [Google Scholar] [CrossRef]
- Hirose, J.; Yamazoe, A.; Hosoyama, A.; Kimura, N.; Suenaga, H.; Watanabe, T.; Fujihara, H.; Futagami, T.; Goto, M.; Furukawa, K. Draft genome sequence of the polychlorinated biphenyl-degrading bacterium Comamonas testosteroni KF712 (NBRC 110673). Genome Announc. 2015, 3, e01214-15. [Google Scholar] [CrossRef]
- Nishi, A.; Tominaga, K.; Furukawa, K. A 90-kilobase conjugative chromosomal element coding for biphenyl and salicylate catabolism in Pseudomonas putida KF715. J. Bacteriol. 2000, 182, 1949–1955. [Google Scholar] [CrossRef]
- Zylstra, G.J.; Gibson, D.T. Toluene degradation by Pseudomonas putida F1. Nucleotide sequence of the todC1C2BADE genes and their expression in Escherichia coli. J. Biol. Chem. 1989, 264, 14940–14946. [Google Scholar]
- Fujihara, H.; Yamazoe, A.; Hosoyama, A.; Suenaga, H.; Kimura, N.; Hirose, J.; Watanabe, T.; Futagami, T.; Goto, M.; Furukawa, K. Draft genome sequence of Pseudomonas aeruginosa KF702 (NBRC 110665), a polychlorinated biphenyl-degrading bacterium isolated from biphenyl-contaminated soil. Genome Announc. 2015, 3, e00517-15. [Google Scholar] [CrossRef]
- Triscari-Barberi, T.; Simone, D.; Calabrese, F.M.; Attimonelli, M.; Hahn, K.R.; Amoako, K.K.; Turner, R.J.; Fedi, S.; Zannoni, D. Genome sequence of the polychlorinated-biphenyl degrader Pseudomonas pseudoalcaligenes KF707. J. Bacteriol. 2012, 194, 4426–4427. [Google Scholar] [CrossRef]
- Watanabe, T.; Yamazoe, A.; Hosoyama, A.; Fujihara, H.; Suenaga, H.; Hirose, J.; Futagami, T.; Goto, M.; Kimura, N.; Furukawa, K. Draft genome sequence of Cupriavidus pauculus strain KF709, a biphenyl-utilizing bacterium isolated from biphenyl-contaminated soil. Genome Announc. 2015, 3, e00222-15. [Google Scholar] [CrossRef]
- Watanabe, T.; Yamazoe, A.; Hosoyama, A.; Fujihara, H.; Suenaga, H.; Hirose, J.; Futagami, T.; Goto, M.; Kimura, N.; Furukawa, K. Draft genome sequence of Pseudomonas toyotomiensis KF710, a polychlorinated biphenyl-degrading bacterium isolated from biphenyl-contaminated soil. Genome Announc. 2015, 3, e00223-15. [Google Scholar] [CrossRef]
- Suenaga, H.; Yamazoe, A.; Hosoyama, A.; Kimura, N.; Hirose, J.; Watanabe, T.; Fujihara, H.; Futagami, T.; Goto, M.; Furukawa, K. Complete genome sequence of the polychlorinated biphenyl-degrading bacterium Pseudomonas putida KF715 (NBRC 110667) isolated from biphenyl-contaminated soil. Genome Announc. 2017, 5, e01624-16. [Google Scholar] [CrossRef]
- Hirose, J.; Yamazoe, A.; Hosoyama, A.; Kimura, N.; Suenaga, H.; Watanabe, T.; Fujihara, H.; Futagami, T.; Goto, M.; Furukawa, K. Draft genome sequence of the polychlorinated biphenyl-degrading bacterium Pseudomonas stutzeri KF716 (NBRC 110668). Genome Announc. 2015, 3, e01215-15. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Li, K.-B. ClustalW-MPI: ClustalW analysis using distributed and parallel computing. Bioinformatics 2003, 19, 1585–1586. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; CSHL Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Erickson, B.D.; Mondello, F.J. Nucleotide sequencing and transcriptional mapping of the genes encoding biphenyl dioxygenase, a multicomponent polychlorinated-biphenyl-degrading enzyme in Pseudomonas strain LB400. J. Bacteriol. 1992, 174, 2903–2912. [Google Scholar] [CrossRef] [PubMed]
- Shingler, V.; Powlowski, J.; Marklund, U. Nucleotide sequence and functional analysis of the complete phenol/3,4-dimethylphenol catabolic pathway of Pseudomonas sp. strain CF600. J. Bacteriol. 1992, 174, 711–724. [Google Scholar] [CrossRef]
- Brown, N.L.; Misra, T.K.; Winnie, J.N.; Schmidt, A.; Seiff, M.; Silver, S. The nucleotide sequence of the mercuric resistance operons of plasmid R100 and transposon Tn501: further evidence for mer genes which enhance the activity of the mercuric ion detoxification system. Mol. Gen. Genet. 1986, 202, 143–151. [Google Scholar] [CrossRef]
- Intorne, A.C.; de Oliveira, M.V.V.; de M Pereira, L.; de Souza Filho, G.A. Essential role of the czc determinant for cadmium, cobalt and zinc resistance in Gluconacetobacter diazotrophicus PAl 5. Int. Microbiol. Off. J. Span. Soc. Microbiol. 2012, 15, 69–78. [Google Scholar]
- Ohtsubo, Y.; Delawary, M.; Kimbara, K.; Takagi, M.; Ohta, A.; Nagata, Y. BphS, a key transcriptional regulator of bph genes involved in polychlorinated biphenyl/biphenyl degradation in Pseudomonas sp. KKS102. J. Biol. Chem. 2001, 276, 36146–36154. [Google Scholar] [CrossRef]
- Christie, P.J.; Whitaker, N.; González-Rivera, C. Mechanism and structure of the bacterial type IV secretion systems. Biochim. Biophys. Acta 2014, 1843, 1578–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mavrodi, D.V.; Loper, J.E.; Paulsen, I.T.; Thomashow, L.S. Mobile genetic elements in the genome of the beneficial rhizobacterium Pseudomonas fluorescens Pf-5. BMC Microbiol. 2009, 9, 8. [Google Scholar] [CrossRef]
- Johnson, C.M.; Grossman, A.D. Integrative and conjugative elements (ICEs): What they do and how they work. Annu. Rev. Genet. 2015, 49, 577–601. [Google Scholar] [CrossRef]
- Gaillard, M.; Vallaeys, T.; Vorhölter, F.J.; Minoia, M.; Werlen, C.; Sentchilo, V.; Pühler, A.; van der Meer, J.R. The clc element of Pseudomonas sp. strain B13, a genomic island with various catabolic properties. J. Bacteriol. 2006, 188, 1999–2013. [Google Scholar] [CrossRef]
- Bellanger, X.; Payot, S.; Leblond-Bourget, N.; Guédon, G. Conjugative and mobilizable genomic islands in bacteria: Evolution and diversity. FEMS Microbiol. Rev. 2014, 38, 720–760. [Google Scholar] [CrossRef]
- Juhas, M.; van der Meer, J.R.; Gaillard, M.; Harding, R.M.; Hood, D.W.; Crook, D.W. Genomic islands: Tools of bacterial horizontal gene transfer and evolution. FEMS Microbiol. Rev. 2009, 33, 376–393. [Google Scholar] [CrossRef]
- Wozniak, R.A.F.; Waldor, M.K. Integrative and conjugative elements: Mosaic mobile genetic elements enabling dynamic lateral gene flow. Nat. Rev. Microbiol. 2010, 8, 552–563. [Google Scholar] [CrossRef]
- Lee, C.A.; Babic, A.; Grossman, A.D. Autonomous plasmid-like replication of a conjugative transposon. Mol. Microbiol. 2010, 75, 268–279. [Google Scholar] [CrossRef] [Green Version]
- Carraro, N.; Poulin, D.; Burrus, V. Replication and active partition of integrative and conjugative elements (ICEs) of the SXT/R391 family: The line between ICEs and conjugative plasmids is getting thinner. PLoS Genet. 2015, 11, e1005298. [Google Scholar] [CrossRef]
- Ravatn, R.; Studer, S.; Springael, D.; Zehnder, A.J.B.; van der Meer, J.R. Chromosomal integration, tandem amplification, and deamplification in Pseudomonas putida F1 of a 105-kilobase genetic element containing the chlorocatechol degradative genes from Pseudomonas sp. strain B13. J. Bacteriol. 1998, 180, 4360–4369. [Google Scholar]
- Sentchilo, V.; Ravatn, R.; Werlen, C.; Zehnder, A.J.B.; van der Meer, J.R. Unusual integrase gene expression on the clc genomic island in Pseudomonas sp. strain B13. J. Bacteriol. 2003, 185, 4530–4538. [Google Scholar] [CrossRef]
- Sentchilo, V.; Czechowska, K.; Pradervand, N.; Minoia, M.; Miyazaki, R.; van der Meer, J.R. Intracellular excision and reintegration dynamics of the ICEclc genomic island of Pseudomonas knackmussii sp. strain B13. Mol. Microbiol. 2009, 72, 1293–1306. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | NBRC Number | DDBJ/EMBL/GenBank Accession Number | References |
|---|---|---|---|
| Pseudomonas abietaniphila KF701 | 110664 | B BQJ01000001-BBQJ01000140 | [15,17] |
| Pseudomonas aeruginosa KF702 | 110665 | B BQK01000001-BBQK01000091 | [15,24] |
| Pseudomonas putida KF703 | 110666 | BBQL01000001-BBQL01000135 | [15,18] |
| Pseudomonas furukawaii KF707 | 110670 | AP014862 | [15,19,25] |
| Cupriavidus basilensis KF708 | 110671 | B BQM01000001-BBQM01000062 | [15,20] |
| Cupriavidus pauculus KF709 | 110672 | BBQN01000001-BBQN01000227 | [15,26] |
| Pseudomonas toyotomiensis KF710 | 110674 | BBQO01000001-BBQO01000029 | [15,27] |
| Comamonas testosteroni KF712 | 110673 | BBQP01000001-BBQP01000097 | [15,21] |
| Pseudomonas putida KF715 | 110667 | AP015029, AP015030-AP015033 | [15,16,22,28] |
| Pseudomonas stutzeri KF716 | 110668 | BBQQ01000001-BBQQ01000030 | [29] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirose, J.; Fujihara, H.; Watanabe, T.; Kimura, N.; Suenaga, H.; Futagami, T.; Goto, M.; Suyama, A.; Furukawa, K. Biphenyl/PCB Degrading bph Genes of Ten Bacterial Strains Isolated from Biphenyl-Contaminated Soil in Kitakyushu, Japan: Comparative and Dynamic Features as Integrative Conjugative Elements (ICEs). Genes 2019, 10, 404. https://doi.org/10.3390/genes10050404
Hirose J, Fujihara H, Watanabe T, Kimura N, Suenaga H, Futagami T, Goto M, Suyama A, Furukawa K. Biphenyl/PCB Degrading bph Genes of Ten Bacterial Strains Isolated from Biphenyl-Contaminated Soil in Kitakyushu, Japan: Comparative and Dynamic Features as Integrative Conjugative Elements (ICEs). Genes. 2019; 10(5):404. https://doi.org/10.3390/genes10050404
Chicago/Turabian StyleHirose, Jun, Hidehiko Fujihara, Takahito Watanabe, Nobutada Kimura, Hikaru Suenaga, Taiki Futagami, Masatoshi Goto, Akiko Suyama, and Kensuke Furukawa. 2019. "Biphenyl/PCB Degrading bph Genes of Ten Bacterial Strains Isolated from Biphenyl-Contaminated Soil in Kitakyushu, Japan: Comparative and Dynamic Features as Integrative Conjugative Elements (ICEs)" Genes 10, no. 5: 404. https://doi.org/10.3390/genes10050404