Identification and Expression of SAUR Genes in the CAM Plant Agave
by
 , , ,
, , ,
Gang Deng
1,† ,
,
Xing Huang
2,*,†,
Li Xie
3,†,
Shibei Tan
2,
Thomas Gbokie, Jr.
4,
Yaning Bao
5,
Zhouli Xie
6 and
Kexian Yi
2,* 1
School of Agriculture, Yunnan University, Kunming 650504, China
2
Environment and Plant Protection Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou 571101, China
3
College of Forestry, Hainan University, Haikou 570228, China
4
College of Plant Protection, Nanjing Agricultural University, Nanjing 210095, China
5
College of Plant Science and Technology, Huazhong Agricultural University, Wuhan 430070, China
6
Department of Genetics, Development and Cell Biology, Iowa State University, Ames, IA 50011, USA
*
Authors to whom correspondence should be addressed.
†
These authors have contributed equally to this work.
Genes 2019, 10(7), 555; https://doi.org/10.3390/genes10070555
Submission received: 27 May 2019
/
Revised: 13 July 2019
/
Accepted: 18 July 2019
/
Published: 23 July 2019
(This article belongs to the Special Issue Genetics, genomics, and evolution of CAM photosynthesis)
Abstract
:Agave species are important crassulacean acid metabolism (CAM) plants and widely cultivated in tropical areas for producing tequila spirit and fiber. The hybrid H11648 of Agave ((A. amaniensis × A. angustifolia) × A. amaniensis) is the main cultivar for fiber production in Brazil, China, and African countries. Small Auxin Up-regulated RNA (SAUR) genes have broad effect on auxin signaling-regulated plant growth and development, while only few SAUR genes have been reported in Agave species. In this study, we identified 43, 60, 24, and 21 SAUR genes with full-length coding regions in A. deserti, A. tequilana, A. H11648, and A. americana, respectively. Although phylogenetic analysis revealed that rice contained a species-specific expansion pattern of SAUR gene, no similar phenomena were observed in Agave species. The in silico expression indicated that SAUR genes had a distinct expression pattern in A. H11648 compared with other Agave species; and four SAUR genes were differentially expressed during CAM diel cycle in A. americana. Additionally, an expression analysis was conducted to estimate SAUR gene expression during different leaf developmental stages, abiotic and biotic stresses in A. H11648. Together, we first characterized the SAUR genes of Agave based on previously published transcriptome datasets and emphasized the potential functions of SAUR genes in Agave’s leaf development and stress responses. The identification of which further expands our understanding on auxin signaling-regulated plant growth and development in Agave species.
1. Introduction
Small Auxin Up-regulated RNA (SAUR) family is one of the important gene families that are involved in auxin signaling-regulated plant growth and development [1]. Genes in this family have been reported as a marker gene in soybean, Arabidopsis, and tobacco during early auxin responses [2,3,4]. Nowadays, the auxin signaling-related function of SAUR genes has also been reported in several other species, including tomato, mung, apple, radish, maize, pepper, rice, cotton, litchi, potato, peach, citrus, ramie, and sorghum [5]. A series of molecular studies in Arabidopsis indicate that these genes participate in plant developmental processes, including in cell elongation [6], cell expansion [7,8,9], light signaling [10,11], branch angle formation [12], pollen tube growth [13] and interactions with brassinosteroid [14], gibberellin [15], and ethylene [16]. In other species, the SAUR genes are associated with fruitlet abscission in citrus [17], auxin-dependent hypocotyl elongation in tomato [18], auxin synthesis, and transport in rice [19], and starch accumulation in cassava [20] as well. Recently, the rapid development of next-generation sequencing (NGS) allows researchers to obtain and explore more information [21]. For example, genome-wide identification of SAUR genes has been performed in rice [22], Arabidopsis, maize, sorghum [23], tomato, potato [24], citrus [17], moso bamboo [25], watermelon [26], cotton [27] and poplar [28]. Moreover, most of those contain the species-specific expansion pattern, which probably contributes to the evolution of special traits among different species [23,26]. Consider SAUR genes are crucial effectors of hormonal and environmental signals, functional characterization of SAUR genes will broaden our understanding in plant growth and development [29,30].
Up till now, only few SAUR genes are reported in Agave species, despite these species are largely applied in alcoholic beverages, fiber, and food production [31]. It reasons that the genomes of Agave are too large to sequence, while the most recent publications on Agave transcriptomes provides a great opportunity for their genetic researches [32]. Furthermore, NGS tools are utilized in Agave species for further functional gene mining, such as stress-related genes in Agave deserti, fructan-related genes in A. tequilana [33], fiber-related genes in Agave hybrid H11648 ((A. amaniensis × A. angustifolia) × A. amaniensis) [31] and CAM photosynthesis-related genes in A. americana [34]. These transcriptome datasets make the identification of SAUR genes and evaluation of their phylogenetic relations in Agave species to be available. In this study, we select the main cultivar in China, A. H11648 to perform further gene expression analysis of SAUR genes at different leaf developmental stages and under abiotic/biotic stresses. Therefore, our findings enhance the understanding of the SAUR genes on auxin signaling-regulated plant growth, development and stress responses in Agave species.
2. Materials and Methods
2.1. Sequence Retrieval and Subcellular Localization
Fifty-six rice SAUR genes were downloaded from public databases [22] and employed as queries to search against Agave transcriptomes by TBlastx method [35]. The transcriptomes of A. deserti, A. tequilana, A. H11648 and A. americana were selected for sequence retrieval [31,33,34,36]. Target sequences from the four Agave transcriptomes were analyzed for coding sequence with ORF-FINDER [37]. SAUR genes of Agave with full coding sequences were used for subcellular localization prediction using CELLO software [38].
2.2. Phylogenetic Analysis
The proteins of SAUR in Arabidopsis, rice and the four Agave species were utilized for phylogenetic analysis. A maximum likelihood (ML) tree was constructed using MEGA 5.0 software [39]. Bootstrap values were tested for 1000 trails to construct the most parsimonious tree. DNAMAN 7 software was used to predict the conserved domains of SAUR [40].
2.3. Plant Materials and RNA Extraction
The plants of A. H11648 were grown in pots at Environment and Plant Protection Institute, Chinese Academy of Tropical Agricultural Sciences (19.99° N, 110.33° E). Shoot, unexpanded leaf and, expanded leaf were separately collected from 2-year-old plants at different developmental stages. Abiotic and biotic stress treatments were conducted using 1-year-old plants. It has been reported that A. H11648 has a high tolerance to heavy metal stress, such as copper and lead [41,42]. Thus, CuSO4 and Pb(NO3)2 solutions were utilized as abiotic stresses for watering plants at the concentrations of 1 g/Kg and 1.3 g/Kg (heavy metal salt/soil), respectively [41,42]. About 2 weeks later, the leaves of plants with treatment were starting curling and collected as samples. Moreover, Zebra disease is the most serious problem of sisal production in China and the pathogen has been identified as Phytophthora nicotianae Breda [36,43]. A Phytophthora nicotianae Breda strain was inoculated on A. H11648 leaves as biotic stress, and the leaves were sampled after 5 days as previously reported [43]. Untreated leaves were also sampled as control. Each treatment was repeated in three individual plants as biological replicates. The collected leaves were immediately placed into liquid nitrogen. A Tiangen RNA prep Pure Plant Kit (Tiangen Biomart, Beijing, China) was used for RNA extraction according to the manufacturer’s protocol. Total RNAs were stored at −80 °C.
2.4. Expression Analysis
SAUR genes in the four Agave species were selected for in silico expression analysis and Reads Per Kilobase per Million mapped reads (RPKM) values in leaves were obtained from previous studies [31,33,34,36]. For qRT-PCR analysis, total RNA of A. H11648 were reverse transcribed with GoScript Reverse Transcription System (Promega, Madison, WI, USA). Each qRT-PCR reaction with a final volume of 20 μL contained 0.5 μL gene-specific primers (10 μM), 1 μL cDNA template, 10 μL TransStart Tip Green qPCR Supermix (Transgen Biotech, Beijing, China), 0.4 μL Passive Reference Dye (50×) (Transgen Biotech, Beijing, China) and 7.6 μL ddH2O. qRT-PCR reaction was carried out in a QuantStudio 6 Flex Real-Time PCR System (Thermo Fisher Scientific, Waltham, MA, USA) with thermal cycles as follows: 94 °C, 30 s; 94 °C, 5 s and 60 °C, 30 s for 40 cycles; dissociation stage. Each sample was repeated three times as technical repeat. Specific primers for eight SAUR genes of A. H11648 were designed with Primer 3, together with the protein phosphatase 2A (PP2A) gene as endogenous control according to a previous study (Table S1) [31,44]. The ΔΔCt method was used for calculating relative expression levels [45].
3. Results
3.1. Identification and Subcellular Localization of Agave SAUR Genes
After sequence retrieval, we found 43, 60, 24 and 21 SAUR genes with full-length coding regions in A. deserti, A. tequilana, A. H11648 and A. americana, respectively (Table S2). These genes ranged from 234–537 base pairs in the coding region with predicted proteins of 77–178 amino acids. About 148 genes were further analyzed for their subcellular localization (Table S2). As a result, most Agave SAUR genes were located in the nucleus or mitochondria (Table 1). And more genes in A. deserti and A. tequilanas were located in the nucleus or mitochondria than those in A. H11648 and A. americana. Only a few genes were located in cytoplasm, chloroplast or plasma membrane. The similar numbers of Agave genes located in chloroplast and plasma membranes, while more genes in A. deserti were located in the cytoplasm than other Agave species. Interestingly, two SAUR genes were located extracellularly in A. tequilana (Table 1).
3.2. Phylogenetic Analysis of Agave SAUR Genes
All SAUR proteins in Arabidopsis (79), rice (56) and Agave species (148) were utilized in the phylogenetic analysis, by which these genes were clustered into eight groups (Figure 1). Typically, Agave sequences were grouped together, and eight subbranches (tetrads) contained sequences from the four Agave species. A. H11648 and A. americana shared similar numbers of SAUR genes in all groups, while the number of which were much smaller than those in Arabidopsis, rice, A. deserti, and A. tequilana (Table 2). Furthermore, more Agave sequences exist in group I, II, and VIII compared with more rice sequences were found in group III and VII and more Arabidopsis sequences were observed in group IV and V. Interestingly, 17 rice sequences and 21 Arabidopsis sequences were clustered together in group III and IV, which also formed a larger amount than in Agave species (Figure 1). About 14 highly conserved amino acid residues of SAUR protein in Agave species were identified based on the alignment (Figure S1).
3.3. In Silico Expression of SAUR Genes in Agave
Based on transcriptomic data, the in silico expression dynamics of SAUR genes in Agave leaves were obtained (Table S2). We further compared the expression patterns of SAUR genes in the eight tetrads, from which two expression modes were characterized (Figure 2A). In mode I, four SAUR genes of A. tequilana showed higher expression levels than other three species, while four SAUR genes of A. deserti were highly expressed than others in mode II. Remarkably, SAUR genes in A. H11648 showed a more distinct expression pattern than other species. Furthermore, four SAUR genes were differentially expressed across the diel cycle of CAM photosynthesis in A. americana (Figure 2B). In addition, GBHM01008063.1 and GBHM01016483.1 tended to perform opposite expression patterns, compared with GBHM01026142.1 and GBHM01043948.1.
3.4. Expression of SAUR Genes in Agave during Leaf Development
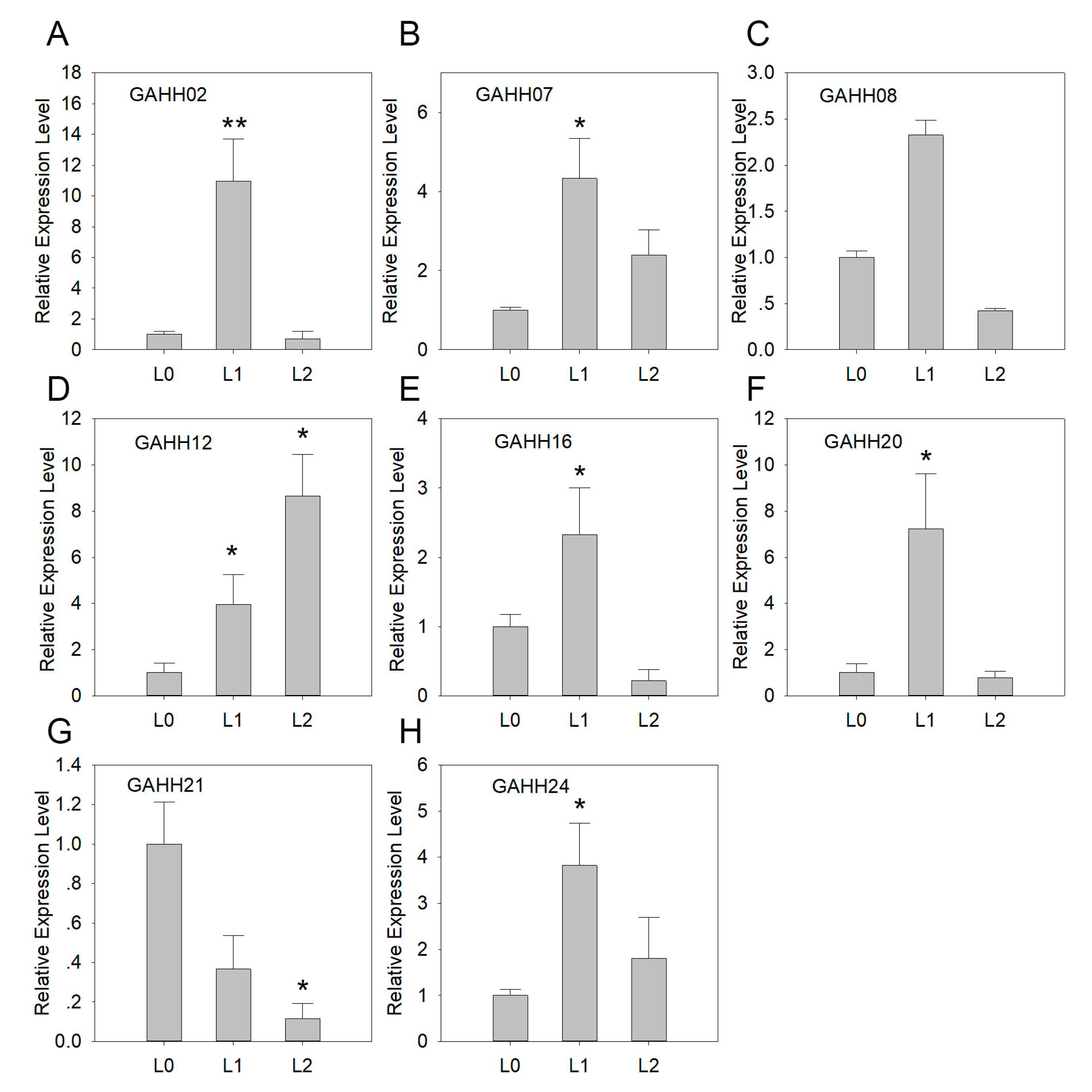
A. H11648 was selected for further qRT-PCR analysis, and we firstly estimated SAUR expression patterns at different leaf developmental stages (Figure 3). Compared with shoot, the expression of six of SAUR genes were increased in unexpanded leaf and then decreased in expanded leaf. Among these, four were significantly increased in unexpanded leaves and GAHH16 was significantly decreased in expanded leaf. Besides, the expression of GAHH12 was significantly increased during the process, while GAHH21 was significantly decreased in both unexpanded and expanded leaf. Furthermore, only four genes had significantly decreased expressions in expanded leaf compared with unexpanded leaf.
3.5. Expression of Agave SAUR Genes under Abiotic and Biotic Stresses
A. H11648 has a high tolerance to Cu and Pb stresses and Phytophthora nicotianae Breda was its main pathogen in cultivation. Thus, the two abiotic stresses and one biotic stress were carried out to evaluate SAUR expressions in A. H11684 leaves, respectively. Five genes were differentially expressed under one of these stresses, i.e. GAHH16 and GAHH20 under Cu stress, GAHH07 under Pb stress and GAHH02 and GAHH12 under biotic stress (Figure 4). The other three genes were highly expressed under the biotic stress and CuSO4/Pb(NO3)2 treatment.
4. Discussion
4.1. Identification and Evolution of Agave SAUR Genes
In this study, we successfully identified 148 SAUR genes with full-length coding regions in four Agave species, indicating the high efficiency of RNA-Seq for genome mining [31]. Different amounts of SAUR genes were obtained in the four Agave species. Especially in A. tequilana, it had relatively more SAUR genes than rice. It was predictable that the large Agave genomes could have more SAUR genes than rice, which might be caused by the whole genome duplications [5,32]. The phylogenetic analysis depicted a species-specific expansion pattern of SAUR gene family in Arabidopsis and rice (Figure 1), which was consistent with previous study [22]. In contrast, no similar expansion pattern was observed in the Agave species, implying that the transcriptome data might not cover the whole SAUR gene family in Agave genomes, and the tissue-specific expression of Agave SAUR genes as well. Several kinds of tissues were sequenced in A. tequilana (4) than in A. deserti (3), while in A. H11648 and A. americana, only leaves were sequenced with the results that were positively correlated with the numbers of SAUR gene identified in the four Agave species [31,33,34]. However, these results were limited to explain the evolution of Agave SAUR genes. Although the availability of Agave genome information could partially explain the evolutionary story, it is very difficult to assemble such large Agave genomes [32]. The recently published walnut genomes have provided a new clue for the assembly of large and heterozygotic genomes [46]. Taken together, too many challenges limit the understanding of the mechanism of SAUR evolution, which needs further investigation.
4.2. Candidate SAUR Genes Involved in Agave Leaf Development and Stress Response
It has been reported that SAUR family is involved in plant growth and developmental processes [29,30], while few SAUR genes have been reported in Agave species. In fact, SAUR genes have crucial roles in plant growth and development throughout the Agave lifespan. The in silico expression of Agave SAUR tetrads revealed a distinct expression pattern in A. H11648 (Figure 2A), irrespective of the SAUR gene not being positively selected during Agave domestication [36]. Interestingly, Agave SAUR genes were differentially expressed across the diel cycle of CAM photosynthesis (Figure 2B), suggesting the existence of a potential relation between auxin signaling and CAM photosynthesis in Agave. It is possible that the diel expressions are related to the opening and closings of stomatal cells [34,47], implying that SAUR involved auxin signaling might participate in this process. Moreover, the potential functions of SAUR genes in starch accumulation might contribute to the expression pattern [20,47].
We further examined their expression during leaf development of A. H11648 and found that all the eight SAUR genes were differentially expressed at least at one developmental stage (Figure 3). This finding indicates the SAUR genes have potential functions during leaf development. As a kind of leaf fiber crop, leaf development covers the process of fiber development in A. H11648. The differentially expressed SAUR genes are most likely associated with cell elongation and expansion [6,7], which therefore introduces a new view for further studies on fiber development in A. H11648.
As effectors of environmental signals in plant growth and developmental processes, SAUR genes are also involved in salt stress responses in rice [48]. And the histidine-rich AtSAUR30 has a metal-binding capacity, which suggests SAUR genes are associated with heavy metal stress as well [49]. Therefore, we performed the Cu and Pb treatments, and found that each stress caused the significant up-regulation of three SAUR genes in A. H11648 (Figure 4). Surprisingly, none of the six genes were differentially expressed under both stresses and they didn’t contain histidine-rich region. This may be due to the occurrence of different regulations between Cu and Pb stress responses in A. H11648. In addition, the main pathogen of A. H11648, Phytophthora nicotianae Breda was inoculated on leaves to estimate SAUR expression patterns. Five genes were differentially expressed during this process implying that these genes might be related to auxin homeostasis–regulated cell wall integrity, and cell wall-mediated immunity [50]. Altogether, three differentially expressed SAUR genes under both abiotic and biotic stresses also indicate an interaction between these stresses. It has been reviewed that heavy metal stresses directly affect plant responses by modulating auxin homeostasis [51]. Therefore, these three genes might be involved in heavy metal responses and plant cell wall-mediated immunity. In the future, further functional characterization of these candidate SAUR genes could potentially enrich our understanding of their functional diversity.
5. Conclusions
In our study, we presented the first identification and expression analysis of SAUR genes in Agave based on previous transcriptome datasets. About 43, 60, 24, and 21 SAUR genes with full-length coding regions were characterized in A. deserti, A. tequilana, A. H11648, and A. americana, respectively. The difference observed in tissue-specific transcriptome datasets might be reasoned to the distinct amounts of SAUR genes in the four Agave species and the tissue-specific expression of SAUR genes. Phylogenetic analysis revealed a species-specific expansion pattern of SAUR gene family in rice, while no similar phenomenon was observed in Agave species. Genome information is still needed to further investigate the duplication and the evolution of Agave SAUR genes. The in silico expression shows a distinct expression pattern of SAUR genes in A. H11648 compared with other Agave species. According to the expression analysis, the differentially expressed SAUR genes during leaf development might contribute to leaf fiber development of A. H11648. Besides, the stress-induced expression patterns of SAUR genes demonstrate their potential functions under abiotic and biotic stresses, which also indicates the potential interactions among these stresses. Therefore, further functional characterization of these candidate SAUR genes could contribute meaningfully our understanding of their functional diversity.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4425/10/7/555/s1, Figure S1: Alignment of SAUR proteins in the five plant species, Table S1: Primers for qRT-PCR analysis. Table S2: Details of SAUR genes in Agave species.
Author Contributions
Conceptualization: X.H. and K.Y.; Formal analysis: G.D., X.H. and L.X.; Funding acquisition: X.H. and K.Y.; Investigation: G.D., X.H., L.X., S.T., T.G.J., and Y.B.; Supervision: K.Y.; Writing—original draft: G.D. and X.H.; Writing—review and revise: T.G.J. and Z.X.
Funding
This research was funded by National Key R & D Program of China (2018YFD0201100), the earmarked fund for China Agriculture Research System (CARS-16-E16), Central Public-interest Scientific Institution Basal Research Fund for Chinese Academy of Tropical Agricultural Sciences (1630042019012, 1630042019041) and Hainan Provincial Natural Science Foundation of China (317260, 319QN275).
Acknowledgments
We would like to thank Xiaohan Yang from Oak Ridge National Laboratory (Oak Ridge, TN 37831, USA) for his suggestions on experimental design.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Weijers, D.; Friml, J. SnapShot: Auxin signaling and transport. Cell 2009, 136, 1172. [Google Scholar] [CrossRef] [PubMed]
- McClure, B.; Guilfoyle, T. Characterization of a class of small auxin-inducible soybean polyadenylated RNAs. Plant Mol. Biol. 1987, 9, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Gil, P.; Liu, Y.; Orbović, V.; Verkamp, E.; Poff, K.L.; Green, P.J. Characterization of the auxin-inducible SAUR-AC1 gene for use as a molecular genetic tool in Arabidopsis. Plant Physiol. 1994, 104, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Roux, C.; Bilang, J.; Theunissen, B.H.; Perrot-Rechenmann, C. Identification of new early auxin markers in tobacco by mRNA differential display. Plant Mol. Biol. 1998, 37, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Bao, Y.; Wang, B.; Liu, L.; Chen, J.; Dai, L.; Baloch, S.U.; Peng, D. Identification of small auxin-up RNA (SAUR) genes in Urticales plants: Mulberry (Morus notabilis), hemp (Cannabis sativa) and ramie (Boehmeria nivea). J. Genet. 2016, 95, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Chae, K.; Isaacs, C.G.; Reeves, P.H.; Maloney, G.S.; Muday, G.K.; Nagpal, P.; Reed, J.W. Arabidopsis SMALL AUXIN UP RNA63 promotes hypocotyl and stamen filament elongation. Plant J. 2012, 71, 684–697. [Google Scholar] [CrossRef] [PubMed]
- Spartz, A.K.; Lee, S.H.; Wenger, J.P.; Gonzalez, N.; Itoh, H.; Inzé, D.; Peer, W.A.; Murphy, A.S.; Overvoorde, P.J.; Gray, W.M. The SAUR19 subfamily of SMALL AUXIN UP RNA genes promote cell expansion. Plant J. 2012, 70, 978–990. [Google Scholar] [CrossRef] [PubMed]
- Spartz, A.K.; Ren, H.; Park, M.Y.; Grandt, K.N.; Lee, S.H.; Murphy, A.S.; Sussman, M.R.; Overvoorde, P.J.; Gray, W.M. SAUR inhibition of PP2C-D phosphatases activates plasma membrane H+-ATPases to promote cell expansion in Arabidopsis. Plant Cell 2014, 26, 2129–2142. [Google Scholar] [CrossRef]
- Ren, H.; Park, M.Y.; Spartz, A.K.; Wong, J.H.; Gray, W.M. A subset of plasma membrane-localized PP2C.D phosphatases negatively regulate SAUR-mediated cell expansion in Arabidopsis. PLoS Genet. 2018, 14, e1007455. [Google Scholar] [CrossRef]
- Roig-Villanova, I.; Bou-Torrent, J.; Galstyan, A.; Carretero-Paulet, L.; Portolés, S.; Rodríguez-Concepción, M.; Martínez-García, J.F. Interaction of shade avoidance and auxin responses: A role for two novel atypical bHLH proteins. EMBO J. 2007, 26, 4756–4767. [Google Scholar] [CrossRef]
- Sato, A.; Sasaki, S.; Matsuzaki, J.; Yamamoto, K.T. Light-dependent gravitropism and negative phototropism of inflorescence stems in a dominant Aux/IAA mutant of Arabidopsis thaliana, axr2. J. Plant Res. 2014, 127, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Bemer, M.; van Mourik, H.; Muiño, J.M.; Ferrándiz, C.; Kaufmann, K.; Angenent, G.C. FRUITFULL controls SAUR10 expression and regulates Arabidopsis growth and architecture. J. Exp. Bot. 2017, 68, 3391–3403. [Google Scholar] [CrossRef] [PubMed]
- He, S.L.; Hsieh, H.L.; Jauh, G.Y. SMALL AUXIN UP RNA62/75 are required for the translation of transcripts essential for pollen tube growth. Plant Physiol. 2018, 178, 626–640. [Google Scholar] [CrossRef] [PubMed]
- Favero, D.S.; Le, K.N.; Neff, M.M. Brassinosteroid signaling converges with SUPPRESSOR OF PHYTOCHROME B4-#3 to influence the expression of SMALL AUXIN UP RNA genes and hypocotyl growth. Plant J. 2017, 89, 1133–1145. [Google Scholar] [PubMed]
- Stamm, P.; Kumar, P.P. Auxin and gibberellin responsive Arabidopsis SMALL AUXIN UP RNA36 regulates hypocotyl elongation in the light. Plant Cell Rep. 2013, 32, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.G.; Chen, H.W.; Li, Q.T.; Tao, J.J.; Bian, X.H.; Ma, B.; Zhang, W.K.; Chen, S.Y.; Zhang, J.S. Three SAUR proteins SAUR76, SAUR77 and SAUR78 promote plant growth in Arabidopsis. Sci. Rep. 2015, 5, 12477. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Dong, C.; Ma, Y.; Deng, L.; He, S.; Yi, S.; Lv, Q.; Zheng, Y. Comprehensive analysis of SAUR gene family in citrus and its transcriptional correlation with fruitlet drop from abscission zone A. Funct. Integr. Genomics 2015, 15, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Spartz, A.K.; Lor, V.S.; Ren, H.; Olszewski, N.E.; Miller, N.D.; Wu, G.; Spalding, E.P.; Gray, W.M. Constitutive expression of Arabidopsis SMALL AUXIN UP RNA19 (SAUR19) in tomato confers auxin-independent hypocotyl elongation. Plant Physiol. 2017, 173, 1453–1462. [Google Scholar] [CrossRef]
- Xu, Y.X.; Xiao, M.Z.; Liu, Y.; Fu, J.L.; He, Y.; Jiang, D.A. The small auxin-up RNA OsSAUR45 affects auxin synthesis and transport in rice. Plant Mol. Biol. 2017, 94, 97–107. [Google Scholar] [CrossRef]
- Ma, P.; Chen, X.; Liu, C.; Meng, Y.; Xia, Z.; Zeng, C.; Lu, C.; Wang, W. MeSAUR1, encoded by a Small Auxin-Up RNA gene, acts as a transcription regulator to positively regulate ADP-Glucose Pyrophosphorylase Small Subunit1a gene in cassava. Front. Plant Sci. 2017, 8, 1315. [Google Scholar] [CrossRef]
- Jiao, W.B.; Schneeberger, K. The impact of third generation genomic technologies on plant genome assembly. Curr. Opin. Plant Biol. 2017, 36, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Tyagi, A.K.; Khurana, J.P. Genome-wide analysis, evolutionary expansion, and expression of early auxin-responsive SAUR gene family in rice (Oryza sativa). Genomics 2006, 88, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hao, X.; Cao, J. Small auxin upregulated RNA (SAUR) gene family in maize: Identification, evolution, and its phylogenetic comparison with Arabidopsis, rice, and sorghum. J. Integr. Plant Biol. 2014, 56, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, S.; He, Y.; Guan, X.; Zhu, X.; Cheng, L.; Wang, J.; Lu, G. Genome-wide analysis of SAUR gene family in Solanaceae species. Gene 2012, 509, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Bai, Q.; Hou, D.; Li, L.; Cheng, Z.; Ge, W.; Liu, J.; Li, X.; Mu, S.; Gao, J. Genome-wide analysis and expression characteristics of small auxin-up RNA (SAUR) genes in moso bamboo (Phyllostachys edulis). Genome 2017, 60, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Huang, X.; Bao, Y.; Wang, B.; Zeng, H.; Cheng, W.; Tang, M.; Li, Y.; Ren, J.; Sun, Y. Genome-wide identification of SAUR genes in watermelon (Citrullus lanatus). Physiol. Mol. Biol. Plants 2017, 23, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, G.; Geng, Y.; Wu, M.; Pei, W.; Zhai, H.; Zang, X.; Li, X.; Zhang, J.; Yu, S.; et al. A genome-wide analysis of the small auxin-up RNA (SAUR) gene family in cotton. BMC Genom. 2017, 18, 815. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Yan, H.; Luo, S.; Pan, F.; Wang, Y.; Xiang, Y. Genome-wide analysis of poplar SAUR gene family and expression profiles under cold, polyethylene glycol and indole-3-acetic acid treatments. Plant Physiol. Biochem. 2018, 128, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Gray, W.M. SAUR proteins as effectors of hormonal and environmental signals in plant growth. Mol. Plant 2015, 8, 1153–1164. [Google Scholar] [CrossRef]
- Stortenbeker, N.; Bemer, M. The SAUR gene family: The plant’s toolbox for adaptation of growth and development. J. Exp. Bot. 2019, 70, 17–27. [Google Scholar] [CrossRef]
- Huang, X.; Xiao, M.; Xi, J.; He, C.; Zheng, J.; Chen, H.; Gao, J.; Zhang, S.; Wu, W.; Liang, Y.; et al. De novo transcriptome assembly of Agave H11648 by Illumina sequencing and identification of cellulose synthase genes in Agave species. Genes 2019, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.L.; Lim, K.Y.; Hanson, L.; Sanchez-Teyer, F.; Bennett, M.D.; Leitch, A.R.; Leitch, I.J. Wild and agronomically important Agave species (Asparagaceae) show proportional increases in chromosome number, genome size, and genetic markers with increasing ploidy. Bot. J. Linn. Soc. 2010, 158, 215–222. [Google Scholar] [CrossRef]
- Gross, S.M.; Martin, J.A.; Simpson, J.; Abraham-Juarez, M.J.; Wang, Z.; Visel, A. De novo transcriptome assembly of drought tolerant CAM plants, Agave deserti and Agave tequilana. BMC Genomics 2013, 14, 563. [Google Scholar] [CrossRef] [PubMed]
- Abraham, P.E.; Yin, H.; Borland, A.M.; Weighill, D.; Lim, S.D.; De Paoli, H.C.; Engle, N.; Jones, P.C.; Agh, R.; Weston, D.J.; et al. Transcript, protein and metabolite temporal dynamics in the CAM plant Agave. Nat. Plants 2016, 2, 16178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Huang, X.; Wang, B.; Xi, J.; Zhang, Y.; He, C.; Zheng, J.; Gao, J.; Chen, H.; Zhang, S.; Wu, W.; et al. Transcriptome comparison reveals distinct selection patterns in domesticated and wild Agave species, the important CAM plants. Int. J. Genom. 2018, 2018, 5716518. [Google Scholar] [CrossRef]
- Rombel, I.T.; Sykes, K.F.; Rayner, S.; Johnston, S.A. ORF-FINDER: A vector for high-throughput gene identification. Gene 2002, 282, 33–41. [Google Scholar] [CrossRef]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins 2006, 64, 643–651. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- DNAMAN-Bioinformatics Solutions. Available online: www.lynnon.com (accessed on 30 March 2019).
- Li, F.; Zhang, L.; Li, X.; Guo, B.; Chen, L.; Qi, Z. Sisal tolerance of cupreous and its accumulation preliminary explore. Chin. Agric. Sci. Bull. 2006, 22, 417–420. [Google Scholar]
- Chen, L.; Zhang, L.; L, F.; Guo, B.; Li, X.; Liao, X.; Qi, Z. A primary research on sisal’s uptake property and the accumulation rule to Pb ions. J. Agro Environ. Sci. 2007, 26, 1879–1883. [Google Scholar]
- Wang, P.; Gao, J.; Yang, F.; Zheng, J.; Liu, Q.; Chen, H.; Yi, K. Transcriptome of sisal leaf pretreated with Phytophthora nicotianae Breda. Chin. J. Trop. Crops 2014, 35, 576–582. [Google Scholar]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Wang, L.; You, F.M.; Rodriguez, J.C.; Deal, K.R.; Chen, L.; Li, J.; Chakraborty, S.; Balan, B.; Jiang, C.Z.; et al. Sequencing a Juglans regia × J. microcarpa hybrid yields high-quality genome assemblies of parental species. Hortic. Res. 2019, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Aubry, S.; Aresheva, O.; Reyna-Llorens, I.; Smith-Unna, R.D.; Hibberd, J.M.; Genty, B. A Specific Transcriptome Signature for Guard Cells from the C4 Plant Gynandropsis gynandra. Plant Physiol. 2016, 170, 1345–1357. [Google Scholar] [CrossRef]
- Jain, M.; Khurana, J.P. Transcript profiling reveals diverse roles of auxin-responsive genes during reproductive development and abiotic stress in rice. FEBS J. 2009, 276, 3148–3162. [Google Scholar] [CrossRef]
- Hara, M.; Kashima, D.; Horiike, T.; Kuboi, T. Metal-binding characteristics of the protein which shows the highest histidine content in the Arabidopsis genome. Plant Biotechnol. 2010, 27, 475–480. [Google Scholar] [CrossRef]
- Bacete, L.; Mélida, H.; Miedes, E.; Molina, A. Plant cell wall-mediated immunity: Cell wall changes trigger disease resistance responses. Plant J. 2018, 93, 614–636. [Google Scholar] [CrossRef]
- Jalmi, S.K.; Bhagat, P.K.; Verma, D.; Noryang, S.; Tayyeba, S.; Singh, K.; Sharma, D.; Sinha, A.K. Traversing the Links between Heavy Metal Stress and Plant Signaling. Front. Plant Sci. 2018, 9, 12. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree of SAUR proteins from Arabidopsis (red), rice (pink), A. deserti (dark green), A. tequilana (green), A. H11648 (blue) and A. americana (brown). Agave homolog tetrads were highlighted in red. The species-specific expansion of SAUR genes was highlighted in rice (a) and Arabidopsis (b), respectively.
Figure 1.
Phylogenetic tree of SAUR proteins from Arabidopsis (red), rice (pink), A. deserti (dark green), A. tequilana (green), A. H11648 (blue) and A. americana (brown). Agave homolog tetrads were highlighted in red. The species-specific expansion of SAUR genes was highlighted in rice (a) and Arabidopsis (b), respectively.

Figure 2.
(A) The in silico expression of SAUR tetrad homologs in the leaves of A. deserti (Ad), A. tequilana (At), A. H11648 (Ah) and A. americana (Am) according to previous studies [31,33,34]. Blanked squares represent no expression data. (B) The in silico expression of SAUR genes across the diel cycle of CAM photosynthesis in A. americana according to a previous study [34]. The numbers of x-axis represent diel times 3, 6, 9, 12, 15, 18, 21 and 24 h from the beginning of the light period. Differentially expressed SAUR genes were highlighted with **. Error bars represent standard deviations.
Figure 2.
(A) The in silico expression of SAUR tetrad homologs in the leaves of A. deserti (Ad), A. tequilana (At), A. H11648 (Ah) and A. americana (Am) according to previous studies [31,33,34]. Blanked squares represent no expression data. (B) The in silico expression of SAUR genes across the diel cycle of CAM photosynthesis in A. americana according to a previous study [34]. The numbers of x-axis represent diel times 3, 6, 9, 12, 15, 18, 21 and 24 h from the beginning of the light period. Differentially expressed SAUR genes were highlighted with **. Error bars represent standard deviations.

Figure 3.
Expression patterns of GAHH02 (A), GAHH07 (B), GAHH08 (C), GAHH12 (D), GAHH16 (E), GAHH20 (F), GAHH21 (G), and GAHH24 (H) at different leaf developmental stages in A. H11648 by qRT-PCR. Y-axis represents relative expression level. L0, L1 and L2 of x-axis represent shoot, unexpanded leaf and expanded leaf, respectively. The error bar represents the standard error. * and ** represent that expression level was increased or decreased by more than 3-fold and 10-fold, respectively (compared with shoot).
Figure 3.
Expression patterns of GAHH02 (A), GAHH07 (B), GAHH08 (C), GAHH12 (D), GAHH16 (E), GAHH20 (F), GAHH21 (G), and GAHH24 (H) at different leaf developmental stages in A. H11648 by qRT-PCR. Y-axis represents relative expression level. L0, L1 and L2 of x-axis represent shoot, unexpanded leaf and expanded leaf, respectively. The error bar represents the standard error. * and ** represent that expression level was increased or decreased by more than 3-fold and 10-fold, respectively (compared with shoot).

Figure 4.
Expression patterns of GAHH02 (A), GAHH07 (B), GAHH08 (C), GAHH12 (D), GAHH16 (E), GAHH20 (F), GAHH21 (G). and GAHH24 (H) under abiotic (Cu and Pb) and biotic (Phytophthora nicotianae Breda) stresses in A. H11648 by qRT-PCR. Y-axis represents relative expression level. CK, CU, PB and PN of x-axis represent control, CuSO4 treatment, Pb(NO3)2 treatment and Phytophthora nicotianae Breda inoculation, respectively. The error bar represents the standard error. * and ** represent that expression level was higher or lower expressed by more than 3-fold and 10-fold, respectively (compared with control).
Figure 4.
Expression patterns of GAHH02 (A), GAHH07 (B), GAHH08 (C), GAHH12 (D), GAHH16 (E), GAHH20 (F), GAHH21 (G). and GAHH24 (H) under abiotic (Cu and Pb) and biotic (Phytophthora nicotianae Breda) stresses in A. H11648 by qRT-PCR. Y-axis represents relative expression level. CK, CU, PB and PN of x-axis represent control, CuSO4 treatment, Pb(NO3)2 treatment and Phytophthora nicotianae Breda inoculation, respectively. The error bar represents the standard error. * and ** represent that expression level was higher or lower expressed by more than 3-fold and 10-fold, respectively (compared with control).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Numbers of Agave Small Auxin Up-regulated RNA (SAUR) genes located at different subcellular positions.
Table 1.
Numbers of Agave Small Auxin Up-regulated RNA (SAUR) genes located at different subcellular positions.
| Subcellular Position | A. deserti | A. tequilana | A. H11648 | A. americana |
|---|---|---|---|---|
| Chloroplast | 3 | 3 | 2 | 1 |
| Cytoplasm | 7 | 3 | 4 | 2 |
| Extracellular | 0 | 2 | 0 | 0 |
| Mitochondria | 12 | 16 | 7 | 7 |
| Nucleus | 20 | 34 | 9 | 10 |
| Plasma Membrane | 1 | 2 | 2 | 1 |
| Total | 43 | 60 | 24 | 21 |
Table 2.
Numbers of rice and Agave SAUR genes in groups I-VII.
| Species | I | II | III | IV | V | VI | VII | VIII | Total |
|---|---|---|---|---|---|---|---|---|---|
| A. thaliana | 8 | 5 | 11 | 36 | 3 | 5 | 6 | 5 | 79 |
| O. sativa | 5 | 8 | 18 | 3 | 1 | 2 | 16 | 3 | 56 |
| A. deserti | 4 | 5 | 3 | 8 | 0 | 1 | 15 | 7 | 43 |
| A. tequilana | 9 | 10 | 8 | 6 | 0 | 5 | 12 | 10 | 60 |
| A. H11648 | 3 | 2 | 4 | 5 | 0 | 1 | 5 | 4 | 24 |
| A. americana | 2 | 1 | 5 | 3 | 0 | 2 | 4 | 4 | 21 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Deng, G.; Huang, X.; Xie, L.; Tan, S.; Gbokie, T., Jr.; Bao, Y.; Xie, Z.; Yi, K. Identification and Expression of SAUR Genes in the CAM Plant Agave. Genes 2019, 10, 555. https://doi.org/10.3390/genes10070555
AMA Style
Deng G, Huang X, Xie L, Tan S, Gbokie T Jr., Bao Y, Xie Z, Yi K. Identification and Expression of SAUR Genes in the CAM Plant Agave. Genes. 2019; 10(7):555. https://doi.org/10.3390/genes10070555
Chicago/Turabian StyleDeng, Gang, Xing Huang, Li Xie, Shibei Tan, Thomas Gbokie, Jr., Yaning Bao, Zhouli Xie, and Kexian Yi. 2019. "Identification and Expression of SAUR Genes in the CAM Plant Agave" Genes 10, no. 7: 555. https://doi.org/10.3390/genes10070555
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.