Trans-Species Polymorphism in Mitochondrial Genome of Camarodont Sea Urchins

1
National Scientific Center of Marine Biology, Far Eastern Branch, Russian Academy of Sciences, 17 Palchevsky Street, 690041 Vladivostok, Russia
2
School of Biomedicine, Far Eastern Federal University, 8 Sukhanov Street, 690950 Vladivostok, Russia
Genes 2019, 10(8), 592; https://doi.org/10.3390/genes10080592
Submission received: 24 June 2019
/
Revised: 30 July 2019
/
Accepted: 2 August 2019
/
Published: 5 August 2019
(This article belongs to the Special Issue Mitochondrial Genomes: Genetic and Transcriptomic Studies)
{kind=link}
{kind=link}
Abstract
:Mitochondrial (mt) genomes of the sea urchins Strongylocentrotus intermedius and Mesocentrotus nudus demonstrate the identical patterns of intraspecific length variability of the ND6 gene, consisting of 489 bp (S variant) and 498 bp (L variant), respectively. For both species, the ND6 length difference is due to the 488A>G substitution, which changes the stop codon TAG in S variant for a tryptophan codon TGG in L variant and elongates the corresponding ND6 protein by three additional amino acids, Trp-Leu-Trp. The phylogenetic analysis based on mt genomes of sea urchins and related echinoderm groups from GenBank has shown the S and L ND6 variants as shared among the camarodont sea urchins; the rest of the echinoderms demonstrate the S variant only. The data suggest that the ND6 488A>G substitution can be the first example of the trans-species polymorphism in sea urchins, persisting at least since the time of the Odontophora diversification at the Eocene/Oligocene boundary (approximately 34 million years ago), which was characterized by an abrupt climate change and significant global ocean cooling. Alternative hypotheses, including the convergent RNA editing and/or codon reassignment, are not supported by direct comparisons of the ND6 gene sequences with the corresponding transcripts using the basic local alignment search tool (BLAST) of full sea urchin transcriptomes.
1. Introduction
Strongylocentrotus intermedius (A. Agassiz, 1863) and Mesocentrotus nudus (A. Agassiz, 1863) are sea urchin species common in the Northwest Pacific [1,2]. These species are distributed mostly from the littoral and upper sublittoral zones to a depth of 25 m [3]. The species represent two major clades (Strongylocentrotus and Mesocentrotus) of the strongylocentrotid sea urchins (order Camarodonta, family Strongylocentrotidae) identified by genetic studies [4,5,6]. The estimated time of divergence between the Strongylocentrotus and Mesocentrotus clades is around 8.5–19 million years ago (MYA) [6,7,8]. Interspecies crosses between S. intermedius and M. nudus have been successfully performed in a laboratory [9,10], which is not unusual for sea urchins (review in [11]).
Little is known about the population genetics of S. intermedius and M. nudus; the available data are limited mostly to allozyme [12,13,14,15], mitochondrial, and microsatellite [16] markers. Using allozyme markers, Zaslavskaya et al. [15] studied the population genetic structure of S. intermedius in the northwestern Sea of Japan. The study revealed a significant genetic heterogeneity of sea urchin settlements without clear geographical pattern. The authors suggested that the genetic heterogeneity may be presumably a result of both genetic drift and natural selection [15]. Using microsatellite and mtDNA markers, Nam et al. [17] studied 10 natural settlements of M. nudus around the Japanese islands and the Korean Peninsula. They did not find any genetic heterogeneity and assumed the recent population expansion (about 5000 to 10,000 years ago) to be a cause of the lack of genetic divergence between the present settlements [17].
Previously, we detected two significantly diverged (FST = 0.671; p < 0.01) haplotype groups in the COI gene (lineage 1 and lineage 2) of S. intermedius collected from three geographical localities of the Sea of Japan [18,19]. We have recently expanded the analysis and sequenced two complete S. intermedius mt genomes (GenBank accession numbers KY964299.1 and KY964300.1) [15] and detected a gene arrangement similar to that of other sea urchin mt genomes published previously (see Tale S1 for references and GenBank accession numbers). The two S. intermedius mt genomes were equal in length (15,705 bp) and similar in nucleotide sequences (Dxy = 0.0083 ± 0.0007) with the only length difference found within the ND6 gene. A conceptual translation revealed equal amino acid lengths of 12 mt proteins except ND6, which consisted of 162 (short, S variant) and 165 (long, L variant) amino acids in the two genomes studied. This length difference was due to the 488A>G substitution (numbered from the beginning of the ND6 gene), which changed the termination codon TAG in S variant for a tryptophan codon TGG in L variant and elongated the corresponding ND6 protein by three additional amino acids, Trp-Leu-Trp. The increased sample of the ND6 sequences (GenBank accession numbers KY964290.1–KY964298.1) revealed five S and four L variants, showing a high level of the 488A>G polymorphism in S. intermedius. Consequently, there may be two variants of the ND6 protein. The two putative variants share the amino acid residues 1–162 but have different C-terminal ends depending on the removal or not of a three-amino acid tail, Trp-Leu-Trp.
In the current study, the complete mt genomes and separate ND6 gene sequences of sea urchins and other echinoderm groups, obtained from GenBank, were analyzed to clarify the ND6 L/S polymorphism detected in S. intermedius. The ND6 DNA sequences were compared with the corresponding RNA transcripts using the Short Read Archive (SRA) sequence database of the National Center for Biotechnology Information (NCBI) to distinguish between the alternative hypotheses explaining the ND6 intra- and interspecific patterns of variability among sea urchins.
2. Materials and Methods
The complete mt genomes and separate ND6 sequences of sea urchins (Echinoidea), sea stars (Asteroidea), brittle stars (Ophiuroidea), sea cucumbers (Holothuroidea), and sea lilies (Crinoidea) were downloaded from the GenBank NCBI (https://www.ncbi.nlm.nih.gov/) [20] genetic sequence database (Flat File Release 231.0, 15 April 2019) (see Table S1 for accession numbers). The nucleotide and protein sequences were aligned using the MUSCLE [21] and MAFFT v. 7 [22] software. The DnaSP v. 5 [23] and PROSEQ v. 2.9 [24] programs were used for intra-and interspecific analyses. Phylogeny reconstructions were based on the full mt genome sequence alignment using the neighbor-joining and maximum-likelihood methods in MEGA v. 7 [25]. MEGA7 or jModelTest [26] were used to find the best-fit model of nucleotide substitution under the maximum likelihood criterion. The general time reversible + gamma + invariant sites (GTR + G + I) model showed the lowest Bayesian information criterion (BIC; [27]) score (407,026.784) and Akaike information criterion (AIC; [28]) value (406,267.663) and was chosen for further phylogenetic reconstructions. Maximum likelihood bootstrap analyses [29] consisted of 500 replicates.
The complete dataset was also analyzed by Bayesian inference using MrBayes v. 3.2.7a (released 6 March 2019; [30]) under the GTR + G + I model with the default run length (1,000,000 generations). The proportion of invariable sites was uniformly distributed over the interval (0.00, 1.00). Gamma distribution was approximated using four categories. Analyses were performed as two independent runs, each with four incrementally heated metropolis-coupled Monte Carlo–Markov chains. Output trees and data were sampled for every 500 generations. Likelihood values reached a plateau within 15,000 generations. A total of 4002 trees in two files were read and 3002 of them were sampled. The number of unique site patterns was 8571. The log likelihood values increased from below −360,656.289 to about −234,790.217 in the first 5000 generations and then to about −234,634.892 after one million generations. The likelihood of the best state for “cold” chain of run 1 was −234,617.92 and the likelihood of the best state for a “cold” chain of run 2 was −234,618.40. The average standard deviation of split frequencies was 0.000269 after 1,000,000 generations indicating stationary conditions. A convergence diagnostic, the potential scale reduction factor (PSRF) [31] was between 1.000 and 1.006 for all parameters, thus, indicating a good sample from the posterior probability distribution.
The tree topologies for the camarodont sea urchins obtained by the neighbor-joining and maximum-likelihood methods, as well as by the Bayesian inference, were very similar. The close congruence could be explained by the facts that the dataset was relatively straightforward and included only complete mt genome sequences of sea urchins. The quality of alignment was high, and the length of alignment was long enough (a total of 19.1 kb). As has been shown by many authors, the relative efficiencies of different methods applied for obtaining the correct tree topology are very close under these conditions (e.g., [32,33]). The difference between the methods applied to the camarodont sea urchins (our dataset) resulted in slightly different topologies and bootstrap values and had no changes concerning the distribution of the ND6 L and S variants. To be conservative, we provide the lowest bootstrap values obtained by the maximum-likelihood method.
Physicochemical features of the putative S and L variants of the ND6 protein were computed using the Protein Feature Server (PROFEAT) [34]. Full sea urchin transcriptomes were searched with the BLAST algorithm [35] using the NCBI Short Read Archive sequence database (https://www.ncbi.nlm.nih.gov/sra/) [36].
3. Results and Discussion
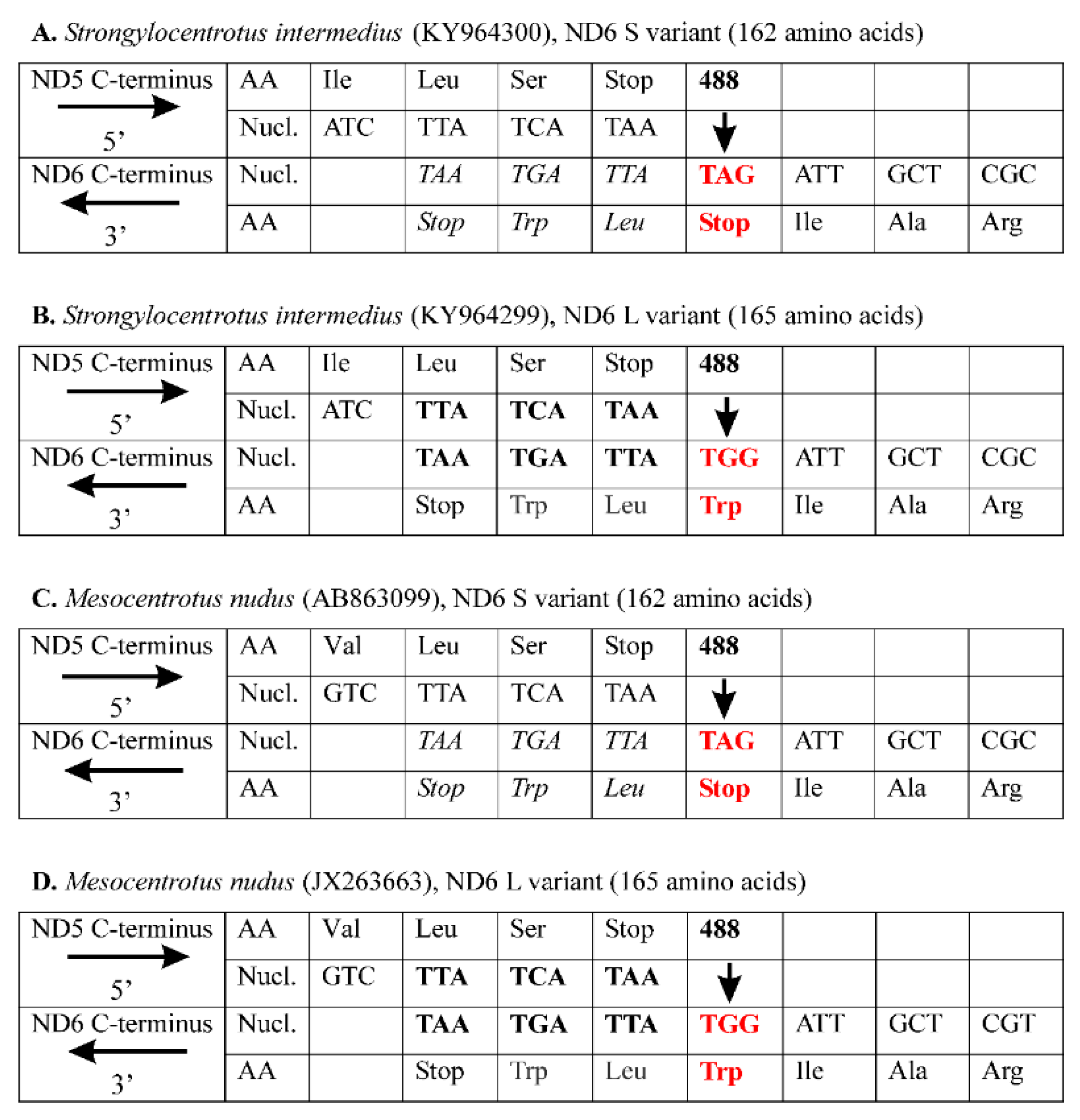
Figure 1 shows the S. intermedius and M. nudus C-terminal amino acid and the corresponding nucleotide sequences for the adjacent ND5 and ND6 genes, which are encoded on the opposite DNA strands with the opposite transcript orientation. To illustrate the pattern of the ND6 variability, we arbitrarily present the last three for S variant and six for L variant of C-terminal amino acids along with a termination codon of the ND6 gene, as well as the last three C-terminal amino acids along with a termination codon of the ND5 gene. For S. intermedius, there is no overlap between the ND5 and ND6 genes for the S variant (Figure 1A), however there is a 9-bp overlap for the L variant (Figure 1B). The length difference is due to the 488A>G substitution (indicated by vertical bold arrow), which changes the ND6 stop codon TAG in S variant for a tryptophan codon TGG in L variant (highlighted in red bold type), adding a 9-bp downstream elongation TTATGATAA (including a termination codon TAA). The conceptually translated proteins consist of 162 and 165 amino acids for the S and L variants, respectively. The L variant is by three amino acids longer than the S variant due to the C-terminal three-amino acid tail, Trp-Leu-Trp. Consequently, there may be two variants of the ND6 protein. The two putative variants share the amino acid residues 1–162 but have different C-termini depending on the removed or not three-amino acid tail, Trp-Leu-Trp (Figure 1A,B).
The same pattern of ND6 intraspecific variability has been found in the other sea urchin, Mesocentrotus nudus, whose multiple ND6 sequences are available in GenBank (accession numbers AB863097.1–AB863112.1; [37]). For this species, there are two main ND6 variants consisting of 162 and 165 amino acids, and the length difference results from the same polymorphic site (488A>G), which determines a termination codon TAG in S variant (Figure 1C) but a tryptophan codon TGG and subsequent downstream elongation TTATGATAA in L variant (Figure 1D). The nucleotide elongation encodes an identical Trp-Leu-Trp C-terminal tail of the ND6 L variant for both sea urchin species (Figure 1). Thus, S. intermedius and M. nudus demonstrate the identical patterns of intraspecific ND6 L/S length polymorphism. Five S and four L variants have been recorded from S. intermedius (GenBank accession numbers KY964290.1–KY964298.1); two S and 14 L variants, from M. nudus (GenBank accession numbers AB863097.1–AB863112.1; [37]). Another length variant of the ND6 sequence has been detected in M. nudus (AB863103.1), which is longer than other sequences belonging to the L variant due to two additional amino acids, Asn–Asp [37].
To clarify the phylogenetic distribution of the observed ND6 variability, the complete mt genomes and separate ND6 gene sequences of sea urchins and other echinoderm groups available in GenBank were analyzed. Figure 2 shows a maximum likelihood phylogenetic tree based on the complete mt genomes of 30 sea urchin species. The topology of the tree is in congruence with the recent most comprehensive morphological and phylogenetic analyses (e.g., [38,39,40,41,42,43,44] and references therein). The ND6 L and S variants (indicated by the corresponding letters, Figure 2, right) are shared among the sea urchins of the order Camarodonta, including mostly the members of the superfamily Odontophora [8,38,43]. Outside Odontophora, the L variant is found in Mespilia globulus (infraorder Temnopleuridea) only (Figure 2). The members of the sea urchin orders Spatangoida (Echinocardium cordatum and Nacospatangus alta), Stomopneustoida (Glyptocidaris crenularis), Arbacioida (Arbacia lixula), and Diadematoida (Diadema setosum and Echinotrix diadema) (Figure 2), as well as other echinoderm classes including Asteroidea, Ophiuroidea, Holothuroidea, and Crinoidea (see Table S1 for species names and accession numbers), demonstrate the ND6 S variant only, suggesting its ancestral state. The ND6 gene of the sea urchin Eucidaris tribuloides (order Cidaroida) is truncated in such a way that the corresponding ND6 protein lacks seven amino acids of the S-terminus (Figure S1); the species was excluded from further analysis.
The ND6 terminus is highly conserved in all the sea urchins studied (Figure S1, Table S2). The elongation of the ND6 L variant includes the same three amino acid residues, Trp-Leu-Trp, in most species except Heterocentrotus mammillatus and Mespilia globulus having Leu–Leu–Trp and Leu–Leu–Cys, respectively (Table S2). The high functional constraint of the ND6 C-terminal region could be explained by the binding site for the transcription factor mtDBP located at the junction between the ND5 and ND6 genes [45,46]. The transcription factor regulates the interplay between mtDNA transcription and replication in mitochondria. The binding sites of the mtDBP are conserved between the sea urchins Paracentrotus lividus and Strongylocentrotus purpuratus (review in [46]). The regulatory network architecture of the developmental genes is highly conserved in sea urchins (e.g., [41,42,47,48]).
Thus, the data have revealed a shared intra- and interspecific variability at the ND6 gene among the camarodont sea urchins. It is unlikely that the ND6 length polymorphism can be explained by a sequencing problem associated with sea urchins, because identical results were obtained by different authors investigating different sea urchin species (see Table S1). The pattern can represent an example of trans-species polymorphism (TSP), persisting at least since the time of the Odontophora sea urchin diversification at the Eocene/Oligocene boundary (approximately 34 MYA) [8,38,43]. TSP refers to the occurrence of identical or similar alleles maintained by the balancing selection in related species, except for the instances where similarity arose through convergence or introgression [49,50,51]. TSP arises when multiple allelic lineages that have originated in an ancestral species are maintained in descendant species. Assuming that multiple ancestral allelic variants are adaptive, their polymorphism, shared by descendant species, may be maintained by balancing selection for longer periods of time beyond neutral expectations [52]. TSP was frequently reported for the immune genes (the major histocompatibility complex, MHC), where balancing selection can maintain ancestral allelic variants for millions of years after the divergence (review in [53,54,55]). Until present, no TSP phenomenon has been reported for sea urchins.
The ND6 protein is crucial for the assembly and function of complex I (NADH: quinone oxidoreductase) of the mitochondrial electron transport chain that produces approximately 40% of the proton-motive force required to synthesize adenosine triphosphate essential for vital functions reviews in [56,57,58,59]. Respiratory complex I performs the reduction of quinone to quinol and pumps protons across the inner membrane of mitochondria. The ND6 protein is an indispensable part of the quinone binding site and even a single or a few amino acid mutations in ND6 may affect quinone binding and dynamics causing severe pathologies [60,61,62,63,64,65,66,67,68]. The residue substitutions that occur in this complex are assumed to interfere with the efficiency of the proton-pumping process, which then influences metabolic performance.
In spite of the strong functional constraint, the complex I genes have been considered important in the adaptive evolution of mitochondrial genome (e.g., [69,70]). The ND6 protein evolution was found to be under positive selection in cephalopod molluscs [71], Alvinocaridid shrimps [72], mosquitoes [73], grasshoppers [74], cichlid fishes [75], Antarctic nototheniid fishes [76], toad-headed lizards [77], songbirds [78], alpine pheasants [79], African elephants [80], Chinese domestic and Tibetan horses [81,82,83], and Chinese snub-nosed monkeys [84]. A significant departure from neutrality in the ND6 gene was also reported for humans [85]. The data suggest that the ND6 modifications may improve protein conformation and, therefore, the entire complex I subunit interactions. Consequently, the ND6 amino acid length difference, detected in the present study, can be an important source for adaptive variation in sea urchins.
The two putative ND6 protein variants, L and S, have the same amino acid sequences, except for the Trp-Leu-Trp C-terminus tail found in the L variant. The tail contains two tryptophan residues, which could be a radical change because the ND6 protein is relatively short in sea urchins (162–165 amino acids), and, except the Trp-Leu-Trp tail, there is only a single tryptophan residue in the ND6 protein alignment (position 95, Figure S1). Indeed, the two additional tryptophan residues lead to a significant difference in amino acid composition between the two putative ND6 variants (Wilcoxon test, p = 0.0181), which, in turn, explain the marginally significant difference in hydrophobicity (Wilcoxon test, p = 0.0580) and the highly significant differences in physicochemical features estimated by the Grantham [86] chemical distance (Wilcoxon test, p < 0.001) and the Schneider and Wrede [87] physicochemical distance (Wilcoxon test, p < 0.001).
As an alternative, we can suggest that the ND6 gene length difference could however not be translated to the protein level if mt transcripts undergo to convergent RNA editing independently in different sea urchin lineages. The term “RNA editing” is defined as targeted modifications to RNAs that result in sequence differences, via nucleotide insertions/deletions or substitutions, between the transcriptome and the corresponding genomic sequences [88]. RNA editing is found in evolutionarily distant groups including viruses, bacteria, protists, fungi, plants, and animals including humans (e.g., review in [89,90,91,92]). It is a mechanism generating an adaptive protein sequence diversity and, thus, providing organisms with the opportunity to express diverse, functionally distinct, protein isoforms (e.g., [93,94]). Despite RNA editing has not yet been described from echinoderms’ mitochondria, it is suggested as a potential source of sequence diversification of the nuclear Transformer gene family involved in the immune function of the sea urchins S. purpuratus [95,96,97] and Heliocidaris erythrogramma [98].
As another alternative hypothesis, we can suggest that the TAG codon (Figure 1A,C) has been reassigned as an amino acid-encoding codon in sea urchins and does not signal translation termination anymore. This is not without precedent, as the stop-to-leucine and stop-to-alanine TAG reassignments were previously reported from mt genomes of several green algae of the order Sphaeropleales [99,100] and the chytrid fungus Spizellomyces punctatus and its relatives [101]. As in the case of the “RNA editing” alternative hypothesis (see above), the reassignment of a stop codon would result in translation of the 3′-UTR, which can produce potentially toxic extended proteins [102,103,104].
Finally, the shared phylogenetic distribution of the L and S ND6 variants might reflect historical hybridization event(s) between sea urchins, which might have resulted in interspecific recombination of their mt DNA (as it has been found for many other organisms; e.g., [105,106] and references therein). Therefore, we analyzed the ND6 alignments for evidence of recombination using various recombination detection methods implemented in the program RDP4 [107]. All methods failed to reveal any signal of recombination between the ND6 sequences (p > 0.05), thus rejecting hybridization as a possible explanation of the shared ND6 polymorphism among sea urchins.
Recently, large arrays of RNA sequence data have been generated and stored in public repositories such as the Short Read Archive sequence database (www.ncbi.nlm.nih.gov/sra/). Many full sea urchin mt genomes (see Table S1) and transcriptomes are there available [108,109,110,111,112,113,114,115,116]. To assess directly the alternative hypotheses outlined above, we compared the ND6 genomic sequences with the corresponding ND6 transcripts using the BLAST algorithm [35]. In case a RNA-DNA difference is found, it would indicate a possible role for the RNA editing and/or codon reassignment in generating the shared ND6 S/L variability; otherwise, the alternative hypotheses cannot be accepted.
We performed a BLAST search of full transcriptomes (for accession numbers see Table S1) for all sea urchin species except Eucidaris tribuloides (see above). The BLAST search revealed the ND6 RNA sequences to completely match that of DNA, which excluded convergent RNA editing and/or codon reassignment as alternative hypotheses, explaining the shared ND6 polymorphism among sea urchins. Moreover, both S and L ND6 RNA transcripts were detected in S. intermedius and M. nudus (Figure S2), which agrees with the intraspecific ND6 gene variability reported for these species (see above). Thus, the alternative hypotheses to explain the shared intra- and interspecific ND6 polymorphism among sea urchins, including convergent RNA editing and/or codon reassignment, are not supported by direct comparisons of the ND6 DNA sequences with the corresponding ND6 RNA transcripts.
The data suggest that the ND6 488A>G substitution can be the first example of TSP in sea urchins, persisting since the time of the Odontophora diversification at the Eocene/Oligocene boundary (approximately 34 MYA; [8,38,43]). The Eocene/Oligocene boundary was characterized by a significant global ocean cooling [117] associated with the largest extinctions of marine invertebrates in the Cenozoic era [118,119]. At the same time, the major global climate changes caused an extensive floral and faunal turnover (e.g., [120]) and species diversification inferred, for instance, for sea urchins [8], ostracods [121], diatoms [122], and whales [123]. Thus, the two variants of the ND6 gene probably represent a genetic adaptation to the abrupt climate change and significant global ocean cooling and were maintained by the long-lasting balancing selection driven by the climate transition at the Eocene/Oligocene boundary. It has been suggested that the thermal environment is likely to impose particularly strong selection on the mt DNA sequences [124,125], and there is experimental evidence that the thermal selection indeed shapes the mt genome evolution of the fruit fly Drosophila melanogaster [126], which is consistent with the “mitochondrial climatic adaptation” hypothesis (review in [127]).
The L ND6 variant found in Mespilia globulus (infraorder Temnopleuridea; Figure 2, Figure S1, Table S2) suggests that the ND6 L/S TSP could be even more ancient than the Odontophora diversification time because the infraorders Echinidea and Temnopleuridea might share a common ancestor more than 100 MYA [38]. The indispensable function of the ND6 protein and the likely signature of positive selection in other organisms (see above) suggest potential functional consequences for the ND6 S and L variants of sea urchins. Anyway, taking into account that the putative ND6 L/S TSP has been maintained over at least 34 million years of echinoid evolution, it is likely that the ND6 variants could have significant consequences for the sea urchin thermal adaptation and, therefore, they deserve further investigations.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4425/10/8/592/s1: Figure S1. (DOC format: doc). Sea urchin ND6 protein alignment. Figure S2. (DOC format: doc). Short Read Archive sequence database search of the Strongylocentrotus intermedius and Mesocentrotus nudus transcriptomes. Table S1. (DOC format: doc). GenBank accession numbers. Table S2. (DOC format: doc). Sea urchin ND6 C-terminal sequences. DOI: 10.5281/zenodo.3352428.
Funding
The present study was conducted at the National Scientific Center of Marine Biology (Vladivostok, Russia) and received Budgetary support of the Research Work no. 14 “Biodiversity of the World Ocean: Composition and distribution of biota” (State registry no. 115081110047, FASO Russia no. 0268-2014-0014).
Acknowledgments
The author greatly appreciates encouragement and help from Francisco J. Ayala and Elena Balakireva. The author also wishes to thank two anonymous reviewers for helpful comments and suggestions on previous versions of the manuscript.
Conflicts of Interest
The author acknowledges no financial interest or benefit from any direct applications of this research. The author declares no conflicts of interest.
Disclosure Statement
The funders had no role in the study’s design, data collection and analysis, decision to publish, or preparation of the manuscript. The author alone is responsible for the content and writing of the paper.
References
- Jensen, M. The Strongylocentrotidae (Echinoidea), a morphologic and systematic study. Sarsia 1974, 57, 113–148. [Google Scholar] [CrossRef]
- Bazhin, A.G.; Stepanov, V.G. Sea Urchins Fam. Strongylocentrotidae of Seas of Russia; KamchatNIRO: Petropavlovsk-Kamchatsky, Russia, 2012; p. 196. [Google Scholar]
- Kafanov, A.I.; Pavlyuchkov, V.A. Ecology of the commercial sea urchins (genus Strongylocentrotus) of continental Japan Sea. Proc. TINRO Center (Vladivostok) 2001, 128, 349–373. [Google Scholar]
- Tatarenko, D.E.; Poltaraus, A.B. Assignment of the sea urchins Pseudocentrotus depressus to the family Strongylocentrotidae and description of a new genus Mesocentrotus belonging to this group based on the data of DNA-DNA hybridization and comparative morphology. Zool. Zhurn. 1993, 72, 61–72. [Google Scholar]
- Biermann, C.H.; Kessing, B.D.; Palumbi, S.R. Phylogeny and development of marine model species: Strongylocentrotid sea urchins. Evol. Dev. 2003, 5, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H. Molecular phylogenies and divergence times of sea urchin species of Strongylocentrotidae, Echinoida. Mol. Biol. Evol. 2003, 20, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Kober, K.M.; Bernardi, G. Phylogenomics of strongylocentrotid sea urchins. BMC Evol. Biol. 2013, 13, 88. [Google Scholar] [CrossRef] [PubMed]
- Láruson, Á.J. Rates and relations of mitochondrial genome evolution across the Echinoidea, with special focus on the superfamily Odontophora. Ecol. Evol. 2017, 7, 4543–4551. [Google Scholar] [CrossRef]
- Motavkin, P.A.; Evdokimov, V.V. A hybrid kind of sea urchins Strongylocentrotus. DAN SSSR 1978, 241, 1451–1453. [Google Scholar]
- Wang, L.; Han, J.; Xu, W.; Wang, X.; Dong, Y.; Zhou, Z. The species hybridization between sea urchin Strongylocentrotus nudus and Strongylocentrotus intermedius and the seeding production. Fish. Sci. 2003, 22, 9–11. [Google Scholar]
- Lessios, H. Reproductive isolation between species of sea urchins. Bull. Mar. Sci. 2007, 81, 191–208. [Google Scholar]
- Pudovkin, A.I.; Serov, O.L.; Glazko, V.I. Allozyme variation in four echinoids from Peter the Great bay of the Sea of Japan. Genetika 1984, 20, 1139–1147. [Google Scholar]
- Manchenko, G.P. Genetic variability of proteins in sea urchin Strongylocentrotus intermedius. Genetika 1985, 21, 763–769. [Google Scholar]
- Manchenko, G.P.; Yakovlev, S.N. Genetic divergence between three sea urchin species of the genus Strongylocentrotus from the Sea of Japan. Biochem. Syst. Ecol. 2001, 29, 31–44. [Google Scholar] [CrossRef]
- Zaslavskaya, N.I.; Vashchenko, M.A.; Zhadan, P.M. The genetic structure of populations of the sea urchin Strongylocentrotus intermedius from the northwestern Sea of Japan in connection with a shift in spawning time. Russ. J. Mar. Biol. 2012, 38, 325–338. [Google Scholar]
- Balakirev, E.S.; Pavlyuchkov, V.A.; Ayala, F.J. Complete mitochondrial genome of the phenotypically-diverse sea urchin Strongylocentrotus intermedius (Strongylocentrotidae, Echinoidea). Mitochond. DNA Part B Res. 2017, 2, 613–614. [Google Scholar] [CrossRef]
- Nam, W.; Kanno, M.; Watanabe, M.; Ikeda, M.; Kijima, A. Genetic population structure of Strongylocentrotus nudus inferred from microsatellite DNA and mtDNA analyses. Nippon Suisan Gakkaishi 2014, 80, 726–740. [Google Scholar] [CrossRef] [Green Version]
- Balakirev, E.S.; Pavlyuchkov, V.A.; Ayala, F.J. DNA variation and symbiotic associations in phenotypically-diverse sea urchin Strongylocentrotus intermedius. Proc. Natl. Acad. Sci. USA 2008, 105, 16218–16223. [Google Scholar] [CrossRef]
- Balakirev, E.S.; Pavlyuchkov, V.A.; Anisimova, M.; Ayala, F.J. DNA polymorphism and selection at the bindin locus in three Strongylocentrotus sp. (Echinoidea). BMC Genet. 2016, 17, 66. [Google Scholar] [CrossRef]
- The National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 19 June 2019).
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Filatov, D.A. PROSEQ: A software for preparation and evolutionary analysis of DNA sequence data sets. Mol. Ecol. Notes 2002, 2, 621–624. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, G.E. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 1–4. [Google Scholar] [CrossRef]
- Gelman, A.; Rubin, D.B. Inference from iterative simulation using multiple sequences. Stat. Sci. 1992, 7, 457–511. [Google Scholar] [CrossRef]
- Russo, C.A.M.; Takezaki, N.; Nei, M. Efficiencies of different genes and different tree-building methods in recovering a known vertebrate phylogeny. Mol. Biol. Evol. 1996, 13, 525–536. [Google Scholar] [CrossRef]
- Douady, C.J.; Delsuc, F.; Boucher, Y.; Doolittle, W.F.; Douzery, E.J. Comparison of Bayesian and maximum likelihood bootstrap measures of phylogenetic reliability. Mol. Biol. Evol. 2003, 20, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.R.; Lin, H.H.; Han, L.Y.; Jiang, L.; Chen, X.; Chen, Y.Z. PROFEAT: A web server for computing structural and physicochemical features of proteins and peptides from amino acid sequence. Nucleic Acids Res. 2006, 34 (Suppl. S2), W32–W37. [Google Scholar] [CrossRef]
- Altschul, S.; Gish, W.; Miller, W.; Myers, E.; Lipman, D. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- The Sequence Read Archive (SRA). The NCBI Database. Available online: https://www.ncbi.nlm.nih.gov/sra/ (accessed on 19 June 2019).
- Nam, W.S.; Kanno, M.; Kijima, A. Strongylocentrotus nudus NADH:ubiquinone oxidoreductase 6 (ND6) gene, partial cds; mitochondrial gene for mitochondrial product. Unpublished. 2019. [Google Scholar]
- Kroh, A.; Smith, A.B. The phylogeny and classification of post-Palaeozoic echinoids. J. Syst. Palaeontol. 2010, 8, 147–212. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.B.; Kroh, A. Phylogeny of sea urchins. In Sea Urchins: Biology and Ecology, 3rd ed.; Lawrence, J.M., Ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 1–14. [Google Scholar]
- Hopkins, M.J.; Smith, A.B. Dynamic evolutionary change in post-Paleozoic echinoids and the importance of scale when interpreting changes in rates of evolution. Proc. Natl. Acad. Sci. USA 2015, 112, 3758–3763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.R.; Petsios, E.; Davidson, E.H.; Erkenbrack, E.M.; Gao, F.; Bottjer, D.J. Reorganization of sea urchin gene regulatory networks at least 268 million years ago as revealed by oldest fossil cidaroid echinoid. Sci. Rep. 2015, 5, 15541. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.R.; Erkenbrack, E.M.; Hinman, V.F.; McCauley, B.S.; Petsios, E.; Bottjer, D.J. Paleogenomics of echinoids reveals an ancient origin for the double-negative specification of micromeres in sea urchins. Proc. Natl. Acad. Sci. USA 2017, 14, 5870–5877. [Google Scholar] [CrossRef]
- Bronstein, O.; Kroh, A. The first mitochondrial genome of the model echinoid Lytechinus variegatus and insights into Odontophoran phylogenetics. Genomics 2019, 111, 710–718. [Google Scholar] [CrossRef]
- Koch, N.M.; Coppard, S.E.; Lessios, H.A.; Briggs, D.E.; Mooi, R.; Rouse, G.W. A phylogenomic resolution of the sea urchin tree of life. BMC Evol. Biol. 2018, 18, 189. [Google Scholar] [CrossRef]
- Roberti, M.; Polosa, P.L.; Musicco, C.; Milella, F.; Qureshi, S.A.; Gadaleta, M.N.; Jacobs, H.T.; Cantatore, P. In vivo mitochondrial DNA-protein interactions in sea urchin eggs and embryos. Curr. Genet. 1999, 34, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Roberti, M.; Polosa, P.L.; Bruni, F.; Deceglie, S.; Gadaleta, M.N.; Cantatore, P. MTERF factors: A multifunction protein family. Biomol. Concepts 2010, 1, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Hinman, V.F.; Nguyen, A.T.; Cameron, R.A.; Davidson, E.H. Developmental gene regulatory network architecture across 500 million years of echinoderm evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 13356–13361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gildor, T.; Ben-Tabou de-Leon, S. Comparative study of regulatory circuits in two sea urchin species reveals tight control of timing and high conservation of expression dynamics. PLoS Genet. 2015, 11, e1005435. [Google Scholar] [CrossRef] [PubMed]
- Klein, J. Origin of major histocompatibility complex polymorphism: The trans-species hypothesis. Hum. Immunol. 1987, 19, 155–162. [Google Scholar] [CrossRef]
- Klein, J.; Sato, A.; Nagl, S.; O’hUigín, C. Molecular trans-species polymorphism. Annu. Rev. Ecol. Syst. 1998, 29, 21. [Google Scholar] [CrossRef]
- Klein, J.; Sato, A.; Nikolaidis, N. MHC, TSP, and the origin of species: From immunogenetics to evolutionary genetics. Annu. Rev. Genet. 2007, 41, 281–304. [Google Scholar] [CrossRef]
- Takahata, N. A simple genealogical structure of strongly balanced allelic lines and trans-species evolution of polymorphism. Proc. Natl. Acad. Sci. USA 1990, 87, 2419–2423. [Google Scholar] [CrossRef]
- Azevedo, L.; Serrano, C.; Amorim, A.; Cooper, D.N. Trans-species polymorphism in humans and the great apes is generally maintained by balancing selection that modulates the host immune response. Hum. Genomics 2015, 9, 21. [Google Scholar] [CrossRef]
- Těšický, M.; Vinkler, M. Trans-species polymorphism in immune genes: general pattern or MHC-restricted phenomenon? J. Immunol. Res. 2015, 838035. [Google Scholar] [CrossRef]
- Wu, Q.; Han, T.S.; Chen, X.; Chen, J.F.; Zou, Y.P.; Li, Z.W.; Xu, Y.C.; Guo, Y.L. Long-term balancing selection contributes to adaptation in Arabidopsis and its relatives. Genome Biol. 2017, 18, 217. [Google Scholar] [CrossRef]
- Efremov, R.G.; Baradaran, R.; Sazanov, L.A. The architecture of respiratory complex I. Nature 2010, 465, 441–445. [Google Scholar] [CrossRef]
- Baradaran, R.; Berrisford, J.M.; Minhas, G.S.; Sazanov, L.A. Crystal structure of the entire respiratory complex I. Nature 2013, 494, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Sazanov, L.A. A giant molecular proton pump: Structure and mechanism of respiratory complex I. Nat. Rev. Mol. Cell Biol. 2015, 16, 375–388. [Google Scholar] [CrossRef]
- Fiedorczuk, K.; Letts, J.A.; Degliesposti, G.; Kaszuba, K.; Skehel, M.; Sazanov, L.A. Atomic structure of the entire mammalian mitochondrial complex I. Nature 2016, 538, 406–410. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Attardi, G. The mtDNA-encoded ND6 subunit of mitochondrial NADH dehydrogenase is essential for the assembly of the membrane arm and the respiratory function of the enzyme. EMBO J. 1998, 17, 4848–4858. [Google Scholar] [CrossRef]
- Cardol, P.; Matagne, R.F.; Remacle, C. Impact of mutations affecting ND mitochondria-encoded subunits on the activity and assembly of complex I in Chlamydomonas. Implication for the structural organization of the enzyme. J. Mol. Biol. 2002, 319, 1211–1221. [Google Scholar] [CrossRef]
- Kirby, D.M.; Kahler, S.G.; Freckmann, M.L.; Reddihough, D.; Thorburn, D.R. Leigh disease caused by the mitochondrial DNA G14459A mutation in unrelated families. Ann. Neurol. 2000, 48, 102–104. [Google Scholar] [CrossRef]
- Chinnery, P.F.; Brown, D.T.; Andrews, R.M.; Singh-Kler, R.; Riordan-Eva, P.; Lindley, J.; Applegarth, D.A.; Turnbull, D.M.; Howell, N. The mitochondrial ND6 gene is a hot spot for mutations that cause Leber’s hereditary optic neuropathy. Brain 2001, 124, 209–218. [Google Scholar] [CrossRef]
- Ravn, K.; Wibrand, F.; Hansen, F.J.; Horn, N.; Rosenberg, T.; Schwartz, M. An mtDNA mutation, 14453G–>A, in the NADH dehydrogenase subunit 6 associated with severe MELAS syndrome. Eur. J. Hum. Genet. 2001, 9, 805–809. [Google Scholar] [CrossRef]
- Ugalde, C.; Triepels, R.H.; Coenen, M.J.; Van Den Heuvel, L.P.; Smeets, R.; Uusimaa, J.; Briones, P.; Campistol, J.; Majamaa, K.; Nijtmans, L.G.; et al. Impaired complex I assembly in a Leigh syndrome patient with a novel missense mutation in the ND6 gene. Ann. Neurol. 2003, 54, 665–669. [Google Scholar] [CrossRef]
- DeHaan, C.; Habibi-Nazhad, B.; Yan, E.; Salloum, N.; Parliament, M.; Allalunis-Turner, J. Mutation in mitochondrial complex I ND6 subunit is associated with defective response to hypoxia in human glioma cells. BMC Mol. Cancer 2004, 3, 19. [Google Scholar]
- Uehara, N.; Mori, M.; Tokuzawa, Y.; Mizuno, Y.; Tamaru, S.; Kohda, M.; Moriyama, Y.; Nakachi, Y.; Matoba, N.; Yamazaki, T.; et al. New MT-ND6 and NDUFA1 mutations in mitochondrial respiratory chain disorders. Ann. Clin. Transl. Neurol. 2014, 5, 361–369. [Google Scholar] [CrossRef]
- Fiedorczuk, K.; Sazanov, L.A. Mammalian mitochondrial complex I structure and disease-causing mutations. Trends Cell Biol. 2018, 28, 835–867. [Google Scholar] [CrossRef]
- Da Fonseca, R.R.; Johnson, W.E.; O’Brien, S.J.; Ramos, M.J.; Antunes, A. The adaptive evolution of the mammalian mitochondrial genome. BMC Genom. 2008, 9, 119. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, S.; Xu, J.; Guo, Y.; Yang, G. Adaptive Evolution of Mitochondrial Energy Metabolism Genes Associated with Increased Energy Demand in Flying Insects. PLoS ONE 2014, 9, e99120. [Google Scholar] [CrossRef]
- Almeida, D.; Maldonado, E.; Vasconcelos, V.; Antunes, A. Adaptation of the mitochondrial genome in Cephalopods: Enhancing proton translocation channels and the subunit interactions. PLoS ONE 2015, 10, e0135405. [Google Scholar] [CrossRef]
- Wang, Z.; Shi, X.; Sun, L.; Bai, Y.; Zhang, D.; Tang, B. Evolution of mitochondrial energy metabolism genes associated with hydrothermal vent adaption of Alvinocaridid shrimps. Genes Genom. 2017, 39, 1367–1376. [Google Scholar] [CrossRef]
- Hao, Y.J.; Zou, Y.L.; Ding, Y.R.; Xu, W.Y.; Yan, Z.T.; Li, X.D.; Fu, W.B.; Li, T.J.; Chen, B. Complete mitochondrial genomes of Anopheles stephensi and An. dirus and comparative evolutionary mitochondriomics of 50 mosquitoes. Sci. Rep. 2017, 7, 7666. [Google Scholar] [CrossRef]
- Li, X.D.; Jiang, G.F.; Yan, L.Y.; Li, R.; Mu, Y.; Deng, W.A. Positive Selection Drove the Adaptation of Mitochondrial Genes to the Demands of Flight and High-Altitude Environments in Grasshoppers. Front. Genet. 2018, 9, 605. [Google Scholar] [CrossRef] [Green Version]
- Bbole, I.; Zhao, J.L.; Tang, S.J.; Katongo, C. Mitochondrial genome annotation and phylogenetic placement of Oreochromis andersonii and O. macrochir among the cichlids of southern Africa. PLoS ONE 2018, 13, e0203095. [Google Scholar] [CrossRef]
- Zhuang, X.; Cheng, C.H. ND6 gene “lost” and found: Evolution of mitochondrial gene rearrangement in Antarctic notothenioids. Mol. Biol. Evol. 2010, 27, 1391–1403. [Google Scholar] [CrossRef]
- Jin, Y.; Wo, Y.; Tong, H.; Song, S.; Zhang, L.; Brown, R.P. Evolutionary analysis of mitochondrially encoded proteins of toad-headed lizards, Phrynocephalus, along an altitudinal gradient. BMC Genom. 2018, 19, 185. [Google Scholar] [CrossRef]
- Lamb, A.M.; Gan, H.M.; Greening, C.; Joseph, L.; Lee, Y.P.; Morán-Ordóñez, A.; Sunnucks, P.; Pavlova, A. Climate-driven mitochondrial selection: A test in Australian songbirds. Mol. Ecol. 2018, 27, 898–918. [Google Scholar] [CrossRef]
- Gu, P.; Liu, W.; Yao, Y.F.; Ni, Q.Y.; Zhang, M.W.; Li, D.Y.; Xu, H.L. Evidence of adaptive evolution of alpine pheasants to high-altitude environment from mitogenomic perspective. DNA Mapp. Seq. Anal. 2016, 27, 455–462. [Google Scholar] [CrossRef]
- Finch, T.M.; Zhao, N.; Korkin, D.; Frederick, K.H.; Eggert, L.S. Evidence of positive selection in mitochondrial complexes I and V of the african elephant. PLoS ONE 2014, 9, e92587. [Google Scholar] [CrossRef]
- Xu, S.; Luosang, J.; Hua, S.; He, J.; Ciren, A.; Wang, W.; Tong, X.; Liang, Y.; Wang, J.; Zheng, X. High altitude adaptation and phylogenetic analysis of Tibetan horse based on the mitochondrial genome. J. Genet. Genom. 2007, 34, 720–729. [Google Scholar] [CrossRef]
- Ning, T.; Xiao, H.; Li, J.; Hua, S.; Zhang, Y.P. Adaptive evolution of the mitochondrial ND6 gene in the domestic horse. Genet. Mol. Res. 2010, 9, 144–150. [Google Scholar] [CrossRef]
- Ning, T.; Li, J.; Lin, K.; Xiao, H.; Wylie, S.; Hua, S.; Li, H.; Zhang, Y.-P. Complex Evolutionary Patterns Revealed by Mitochondrial Genomes of the Domestic Horse. Curr. Mol. Med. 2014, 14, 1286–1298. [Google Scholar] [CrossRef]
- Yu, L.; Wang, X.; Ting, N.; Zhang, Y. Mitogenomic analysis of Chinese snub-nosed monkeys: Evidence of positive selection in NADH dehydrogenase genes in high-altitude adaptation. Mitochondrion 2011, 11, 497–503. [Google Scholar] [CrossRef]
- Elson, J.L.; Turnbull, D.M.; Howell, N. Comparative genomics and the evolution of human mitochondrial DNA: Assessing the effects of selection. Am. J. Hum. Genet. 2004, 74, 229–238. [Google Scholar] [CrossRef]
- Grantham, R. Amino acid difference formula to help explain protein evolution. Science 1974, 185, 862–864. [Google Scholar] [CrossRef]
- Schneider, G.; Wrede, P. The rational design of amino acid sequences by artificial neural networks and simulated molecular evolution: De novo design of an idealized leader peptidase cleavage site. Biophys. J. 1994, 66, 335–344. [Google Scholar] [CrossRef]
- Gott, J.M.; Emeson, R.B. Functions and mechanisms of RNA editing. Annu. Rev. Genet. 2000, 34, 499–531. [Google Scholar] [CrossRef]
- Knoop, V. When you can’t trust the DNA: RNA editing changes transcript sequences. Cell. Mol. Life Sci. 2011, 68, 567–586. [Google Scholar] [CrossRef]
- Bazak, L.; Haviv, A.; Barak, M.; Jacob-Hirsch, J.; Deng, P.; Zhang, R.; Isaacs, F.J.; Rechavi, G.; Li, J.B.; Levanon, E.Y.; et al. A-to-I RNA editing occurs at over a hundred million genomic sites, located in a majority of human genes. Genome Res. 2014, 24, 365–376. [Google Scholar] [CrossRef]
- Lavrov, D.V.; Pett, W. Animal Mitochondrial DNA as We Do Not Know It: Mt-Genome Organization and Evolution in Nonbilaterian Lineages. Genome Biol. Evol. 2016, 8, 2896–2913. [Google Scholar] [CrossRef]
- Liscovitch-Brauer, N.; Alon, S.; Porath, H.T.; Elstein, B.; Unger, R.; Ziv, T.; Admon, A.; Levanon, E.Y.; Rosenthal, J.J.C. Eisenberg, E.; et al. Trade-off between Transcriptome Plasticity and Genome Evolution in Cephalopods. Cell 2017, 169, 191–202. [Google Scholar] [CrossRef]
- Gommans, W.M.; Mullen, S.P.; Maas, S. RNA editing: A driving force for adaptive evolution? Bioessays 2009, 31, 1137–1145. [Google Scholar] [CrossRef]
- Duan, Y.; Dou, S.; Luo, S.; Zhang, H.; Lu, J. Adaptation of A-to-I RNA editing in Drosophila. PLoS Genet. 2017, 13, e1006648. [Google Scholar] [CrossRef]
- Buckley, K.M.; Terwilliger, D.P.; Smith, L.C. Sequence variations in 185/333 Messages from the purple Sea Urchin suggest posttranscriptional modifications to increase immune diversity. J. Immunol. 2008, 181, 8585–8594. [Google Scholar] [CrossRef]
- Smith, L.C. Innate immune complexity in the purple sea urchin: Diversity of the sp185/333 system. Front. Immunol. 2012, 3, 70. [Google Scholar] [CrossRef]
- Smith, L.C.; Lun, C.M. The SpTransformer gene family (formerly Sp185/333) in the purple Sea Urchin and the functional diversity of the anti-pathogen rSpTransformer-E1 protein. Front. Immunol. 2017, 8, 725. [Google Scholar] [CrossRef]
- Roth, M.O.; Wilkins, A.G.; Cooke, G.M.; Raftos, D.A.; Nair, S.V. Characterization of the highly variable immune response gene family, He185/333, in the sea urchin, Heliocidaris erythrogramma. PLoS ONE 2014, 9, e62079. [Google Scholar] [CrossRef]
- Fučíková, K.; Lewis, P.O.; González-Halphen, D.; Lewis, L.A. Gene arrangement convergence, diverse intron content, and genetic code modifications in mitochondrial genomes of Sphaeropleales (Chlorophyta). Genome Biol. Evol. 2014, 6, 2170–2180. [Google Scholar] [CrossRef]
- Hayashi-Ishimaru, Y.; Ohama, T.; Kawatsu, Y.; Nakamura, K.; Osawa, S. UAG is a sense codon in several chlorophycean mitochondria. Curr. Genet. 1996, 30, 29–33. [Google Scholar] [CrossRef]
- Laforest, M.J.; Roewer, I.; Lang, B.F. Mitochondrial tRNAs in the lower fungus Spizellomyces punctatus: tRNA editing and UAG ‘Stop’ codons recognized as leucine. Nucleic Acids Res. 1997, 25, 626–632. [Google Scholar] [CrossRef]
- Clegg, J.B.; Weatherall, D.J.; Milner, P.F. Haemoglobin Constant Spring—A chain termination mutant? Nature 1971, 234, 337–340. [Google Scholar] [CrossRef]
- Clegg, J.B.; Weatherall, D.J.; Contopolou-Griva, I.; Caroutsos, K.; Poungouras, P.; Tsevrenis, H. Haemoglobin Icaria, a new chain-termination mutant with causes α thalassaemia. Nature 1974, 251, 245–247. [Google Scholar] [CrossRef]
- Hashimoto, S.; Nobuta, R.; Izawa, T.; Inada, T. Translation arrest as a protein quality control system for aberrant translation of the 3’-UTR in mammalian cells. FEBS Lett. 2019, 593, 777–787. [Google Scholar] [CrossRef]
- Balakirev, E.S.; Krupnova, T.N.; Ayala, F.J. DNA variation in the phenotypically-diverse brown alga Saccharina japonica. BMC Plant Biol. 2012, 12. [Google Scholar] [CrossRef]
- Balakirev, E.S.; Romanov, N.S.; Mikheev, P.B.; Ayala, F.J. Mitochondrial DNA variation and introgression in Siberian taimen Hucho taimen. PLoS ONE 2013, 8, e71147. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [Green Version]
- Tu, Q.; Cameron, R.A.; Worley, K.C.; Gibbs, R.A.; Davidson, E.H. Gene structure in the sea urchin Strongylocentrotus purpuratus based on transcriptome analysis. Genome Res. 2012, 22, 2079–2087. [Google Scholar] [CrossRef]
- Tu, Q.; Cameron, R.A.; Davidson, E.H. Quantitative developmental transcriptomes of the sea urchin Strongylocentrotus purpuratus. Dev. Biol. 2014, 385, 160–167. [Google Scholar] [CrossRef]
- Chen, Y.; Chang, Y.; Wang, X.; Qiu, X.; Liu, Y. De novo assembly and analysis of tissue-specific transcriptomes revealed the tissue-specific genes and profile of immunity from Strongylocentrotus intermedius. Fish Shellfish. Immunol. 2015, 46, 723–736. [Google Scholar] [CrossRef]
- Jia, Z.; Wang, Q.; Wu, K.; Wei, Z.; Zhou, Z.; Liu, X. De novo transcriptome sequencing and comparative analysis to discover genes involved in ovarian maturity in Strongylocentrotus nudus. Comp. Biochem. Physiol. D Genom. Proteom. 2017, 23, 27–38. [Google Scholar] [CrossRef]
- Ding, J.; Yang, D.; Chang, Y.; Wang, Y.; Zhang, W.; Chen, T. Comparative transcriptome analysis of tube feet of different colors in the sea urchin Strongylocentrotus intermedius. Genes Genom. 2017, 39, 1215–1225. [Google Scholar] [CrossRef]
- Sun, Z.H.; Zhang, J.; Zhang, W.J.; Chang, Y.Q. Gonadal transcriptomic analysis and identification of candidate sex-related genes in Mesocentrotus nudus. Gene 2019, 698, 72–81. [Google Scholar] [CrossRef]
- Zhan, Y.; Li, J.; Sun, J.; Zhang, W.; Li, Y.; Cui, D.; Hu, W.; Chang, Y. The Impact of Chronic Heat Stress on the Growth, Survival, Feeding, and Differential Gene Expression in the Sea Urchin Strongylocentrotus intermedius. Front. Genet. 2019, 10, 301. [Google Scholar] [CrossRef]
- Zhang, W.; Lv, Z.; Li, C.; Sun, Y.; Jiang, H.; Zhao, M.; Zhao, X.; Shao, Y.; Chang, Y. Transcriptome profiling reveals key roles of phagosome and NOD-like receptor pathway in spotting diseased Strongylocentrotus intermedius. Fish Shellfish. Immunol. 2019, 84, 521–531. [Google Scholar] [CrossRef]
- Zhao, C.; Ding, J.; Yang, M.; Shi, D.; Yin, D.; Hu, F.; Sun, J.; Chi, X.; Zhang, L.; Chang, Y. Transcriptomes reveal genes involved in covering and sheltering behaviors of the sea urchin Strongylocentrotus intermedius exposed to UV-B radiation. Ecotoxicol. Environ. Saf. 2019, 167, 236–241. [Google Scholar] [CrossRef]
- Liu, Z.; Pagani, M.; Zinniker, D.; DeConto, R.; Huber, M.; Brinkhuis, H.; Pearson, A. Global cooling during the Eocene-Oligocene climate transition. Science 2009, 323, 1187–1190. [Google Scholar] [CrossRef]
- Raup, D.M.; Sepkoski, J.J. Periodic extinction of families and genera. Science 1986, 231, 833–836. [Google Scholar] [CrossRef]
- Prothero, D.R. The late Eocene—Oligocene extinctions. Annu. Rev. Earth Planet. Sci. 1994, 22, 145–165. [Google Scholar] [CrossRef]
- Sun, J.; Ni, X.; Bi, S.; Wu, W.; Ye, J.; Meng, J.; Windley, B.F. Synchronous turnover of flora, fauna, and climate at the Eocene-Oligocene Boundary in Asia. Sci. Rep. 2014, 4, 7463. [Google Scholar] [CrossRef]
- Ducasse, O.; Lété, C.; Rousselle, L. Polymorphism and speciation medoc Ostracods at the Eocene/Oligocene boundary (Aquitaine, France). Develop. Palaeontol. Stratigr. 1988, 11, 939–947. [Google Scholar]
- Suto, I. The explosive diversification of the diatom genus Chaetoceros across the Eocene/Oligocene and Oligocene/Miocene boundaries in the Norwegian Sea. Mar. Micropaleont. 2006, 58, 259–269. [Google Scholar] [CrossRef]
- Steeman, M.E.; Hebsgaard, M.B.; Fordyce, R.E.; Ho, S.Y.W.; Rabosky, D.L.; Nielsen, R.; Willerslev, E. Radiation of extant cetaceans driven by restructuring of the oceans. Syst. Biol. 2009, 58, 573–585. [Google Scholar] [CrossRef]
- Ballard, J.W.O.; Whitlock, M.C. The incomplete natural history of mitochondria. Mol. Ecol. 2004, 13, 729–744. [Google Scholar] [CrossRef] [Green Version]
- Dowling, D.K.; Friberg, U.; Lindell, J. Evolutionary implications of nonneutral mitochondrial genetic variation. Trends Ecol. Evol. 2008, 23, 546–554. [Google Scholar] [CrossRef]
- Lajbner, Z.; Pnini, R.; Camus, M.F.; Miller, J.; Dowling, D.K. Experimental evidence that thermal selection shapes mitochondrial genome evolution. Sci. Rep. 2018, 8, 9500. [Google Scholar] [CrossRef] [Green Version]
- Camus, M.F.; Wolff, J.N.; Sgrò, C.M.; Dowling, D.K. Experimental evidence that thermal selection has shaped the latitudinal distribution of mitochondrial haplotypes in Australian fruit flies. Mol. Biol. Evol. 2017, 34, 2600–2612. [Google Scholar] [CrossRef]
Figure 1.
The ND5 and ND6 C-terminus sequences of the sea urchins Strongylocentrotus intermedius (A,B) and Mesocentrotus nudus (C,D). Conceptual translation of the ND5 and ND6 genes is shown as three-letter abbreviations (echinoderm mitochondrial genetic code). The ND5 gene is encoded on the H-strand, and the ND6 gene is encoded on the L-strand. The strands and corresponding transcripts are denoted by horizontal arrows → (H strand) and ← (L strand). The 5’ and 3’ ends are shown on the left. For the L variant, there is an overlap (highlighted in bold) between ND5 and ND6 encoded on the opposite strand. Nucl. is nucleotides; AA is amino acid residues. The ND6 polymorphic site 488A>G is indicated by bold vertical arrows under the numeral 488; the corresponding codons (TAG and TGG) are highlighted in bold red type indicating the changes in S and L variants. The readthrough extension for the S variants is highlighted in italics.
Figure 1.
The ND5 and ND6 C-terminus sequences of the sea urchins Strongylocentrotus intermedius (A,B) and Mesocentrotus nudus (C,D). Conceptual translation of the ND5 and ND6 genes is shown as three-letter abbreviations (echinoderm mitochondrial genetic code). The ND5 gene is encoded on the H-strand, and the ND6 gene is encoded on the L-strand. The strands and corresponding transcripts are denoted by horizontal arrows → (H strand) and ← (L strand). The 5’ and 3’ ends are shown on the left. For the L variant, there is an overlap (highlighted in bold) between ND5 and ND6 encoded on the opposite strand. Nucl. is nucleotides; AA is amino acid residues. The ND6 polymorphic site 488A>G is indicated by bold vertical arrows under the numeral 488; the corresponding codons (TAG and TGG) are highlighted in bold red type indicating the changes in S and L variants. The readthrough extension for the S variants is highlighted in italics.

Figure 2.
A maximum likelihood tree constructed using complete mitochondrial genomes of sea urchins (class Echinoidea). The tree is based on the general time reversible + gamma + invariant sites (GTR + G + I) model of nucleotide substitution. The mitochondrial genome of Cucumaria miniata (AY182376; class Holothuroidea) is used as an outgroup. The numbers at the nodes are bootstrap percentage probability values based on 500 replications (values below 75% are omitted). The species above the first (on top) horizontal line belong to the superfamily Odonthophora; above the second horizontal line, to the infraorder Echinidea; above the third horizontal line, to the order Camarodonta including the infraorders Echinidea and Temnopleuridea. The ND6 L and S variants are indicated by the corresponding letters on the right: L, long (165 amino acids) ND6 variant; S, short (162 amino acids) ND6 variant; L, S, both ND6 variants detected for Strongylocentrotus intermedius and Mesocentrotus nudus. The Eucidaris tribuloides ND6 variant (indicated by asterisk) is shorter than the S variant of other sea urchins due to the deletion of seven amino acids at the C-terminus (see Figure S1).
Figure 2.
A maximum likelihood tree constructed using complete mitochondrial genomes of sea urchins (class Echinoidea). The tree is based on the general time reversible + gamma + invariant sites (GTR + G + I) model of nucleotide substitution. The mitochondrial genome of Cucumaria miniata (AY182376; class Holothuroidea) is used as an outgroup. The numbers at the nodes are bootstrap percentage probability values based on 500 replications (values below 75% are omitted). The species above the first (on top) horizontal line belong to the superfamily Odonthophora; above the second horizontal line, to the infraorder Echinidea; above the third horizontal line, to the order Camarodonta including the infraorders Echinidea and Temnopleuridea. The ND6 L and S variants are indicated by the corresponding letters on the right: L, long (165 amino acids) ND6 variant; S, short (162 amino acids) ND6 variant; L, S, both ND6 variants detected for Strongylocentrotus intermedius and Mesocentrotus nudus. The Eucidaris tribuloides ND6 variant (indicated by asterisk) is shorter than the S variant of other sea urchins due to the deletion of seven amino acids at the C-terminus (see Figure S1).

© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Balakirev, E.S. Trans-Species Polymorphism in Mitochondrial Genome of Camarodont Sea Urchins. Genes 2019, 10, 592. https://doi.org/10.3390/genes10080592
AMA Style
Balakirev ES. Trans-Species Polymorphism in Mitochondrial Genome of Camarodont Sea Urchins. Genes. 2019; 10(8):592. https://doi.org/10.3390/genes10080592
Chicago/Turabian StyleBalakirev, Evgeniy S. 2019. "Trans-Species Polymorphism in Mitochondrial Genome of Camarodont Sea Urchins" Genes 10, no. 8: 592. https://doi.org/10.3390/genes10080592
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.