
Overexpression of NtGPX8a Improved Cadmium Accumulation and Tolerance in Tobacco (Nicotiana tabacum L.)

Abstract
: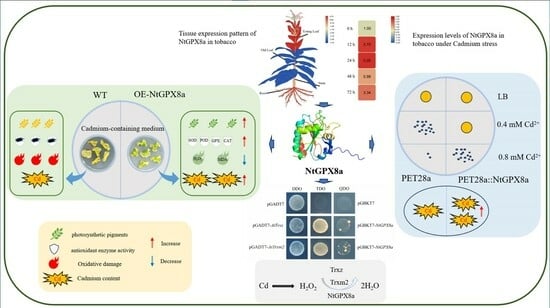
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Genome-Wide Analysis and Identification of the NtGPX Family Genes
2.2. Plant Growth Conditions and Cd2+ Stress
2.3. RNA Extraction, Reverse Transcription, and qRT-PCR
2.4. Heterologous Expression of NtGPX in E. coli and Cadmium Tolerance Assay
2.5. Yeast Two-Hybrid
2.6. Tobacco Genetic Transformation
2.7. Determination of Cd2+ Content in E. coli and Transgenic Tobacco
2.8. Determination of Chlorophylls, Proline, TPC, and TFL Content
2.9. Measurement of MDA and H2O2 Content
2.10. Measurement of GPX, SOD, CAT, POD, and GR Enzyme Activities
2.11. Statistical Analysis
3. Results
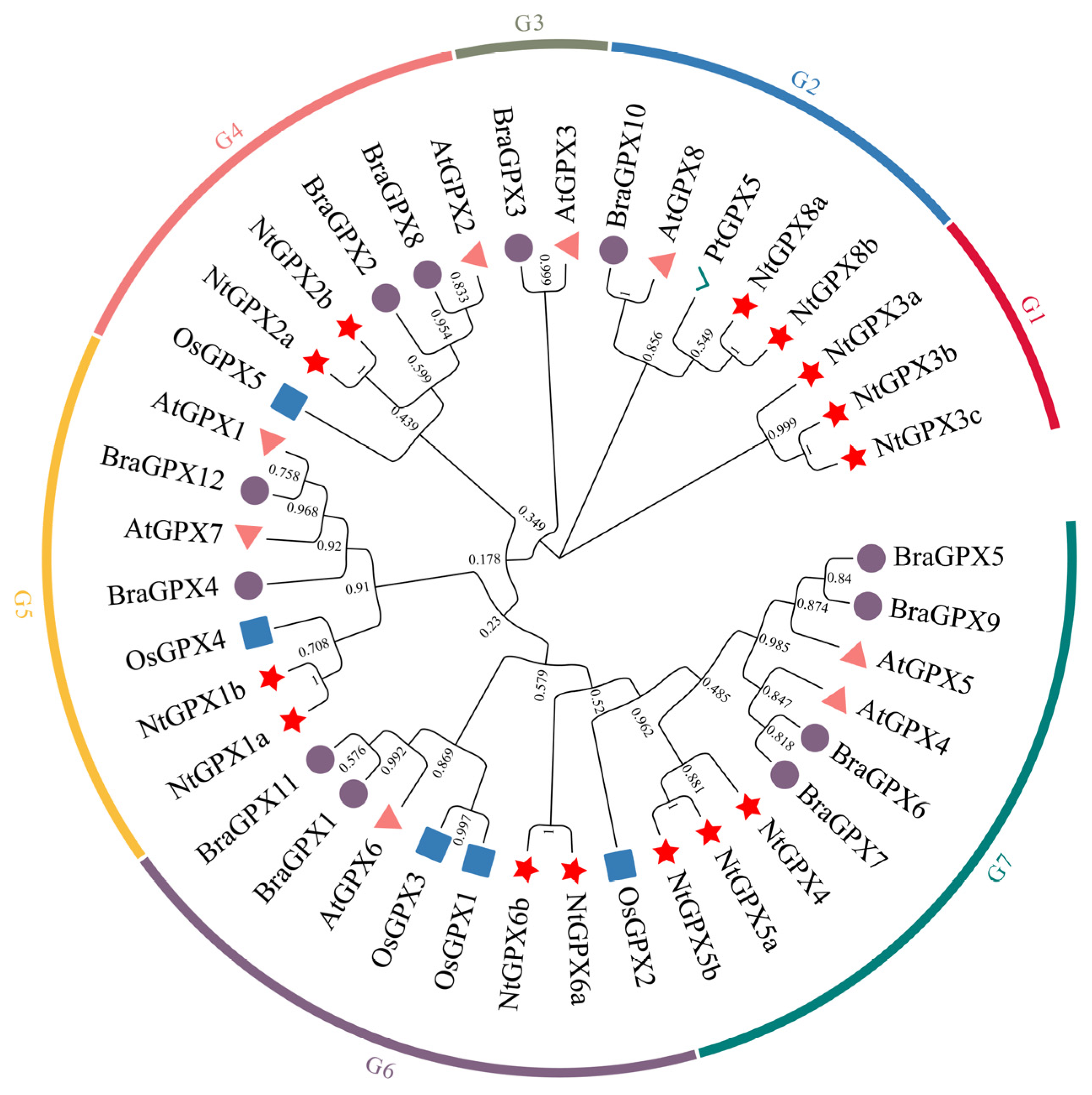
3.1. Genome-Wide Identification and Characterization of NtGPX Gene Family
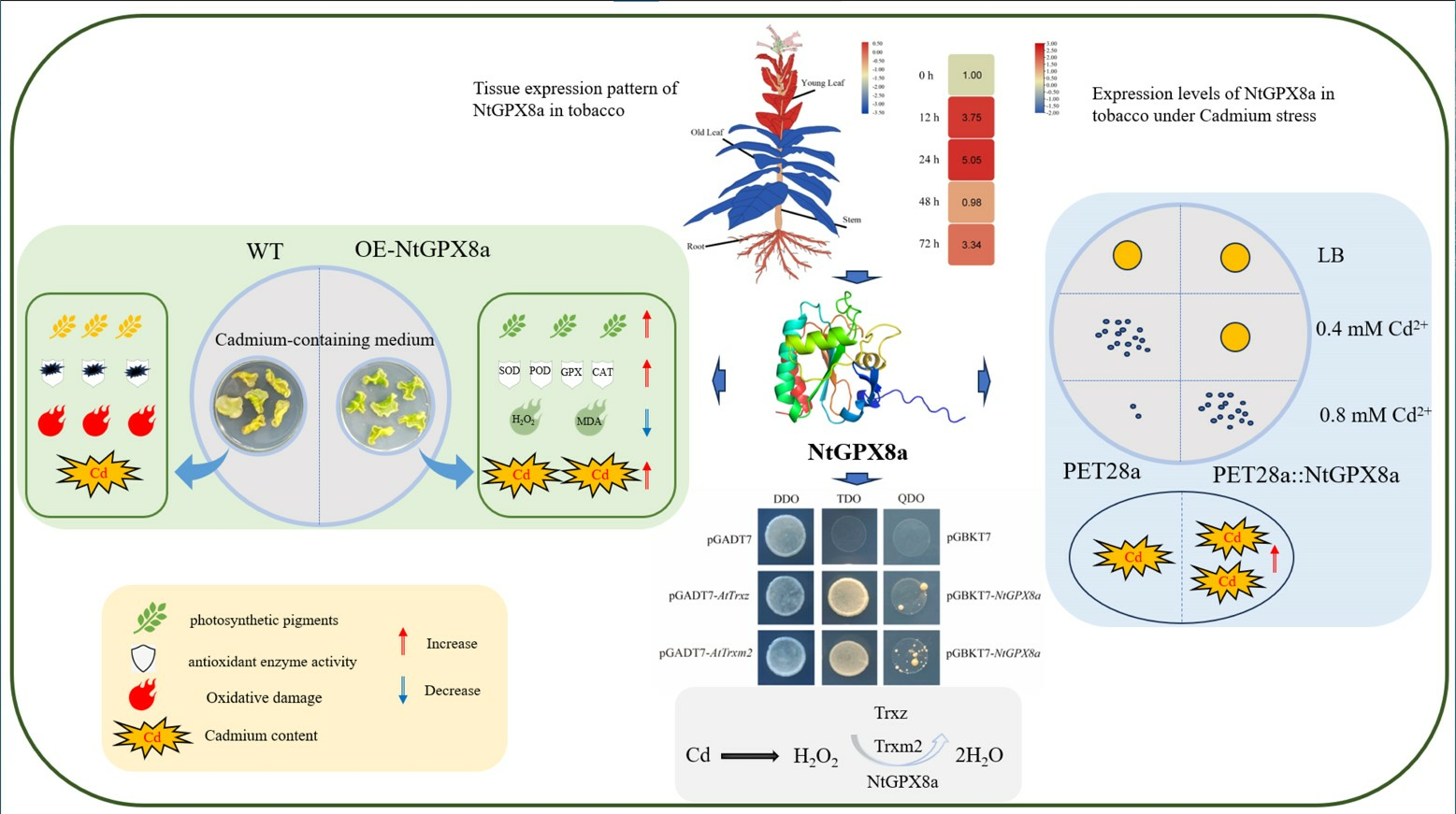
3.2. Tissue Expression Patterns and Cadmium-Induced Expression Patterns of NtGPX Family Genes
3.3. Heterologous Expression of NtGPX8a Confers Cadmium Tolerance in E. coli and Enhances Cadmium Accumulation
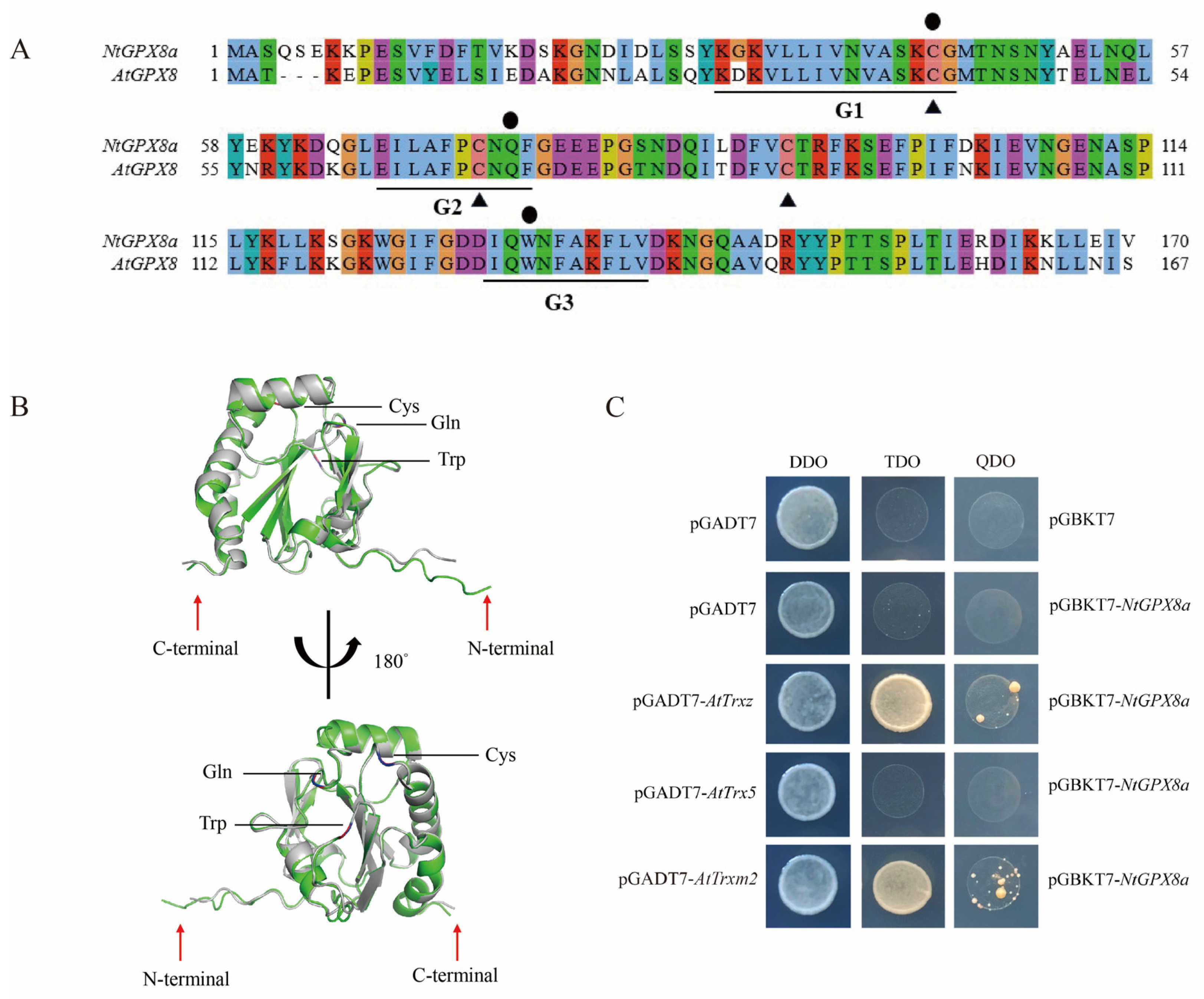
3.4. Structure of NtGPX8a and Selection of Electron Donors
3.5. Overexpression of NtGPX8a Alleviates Cd-Induced Decrease in Photosynthetic Pigments and Enhances Cd Accumulation
3.6. Overexpression of NtGPX8a Mitigates Oxidative Damage Caused by Cd2+ Stress
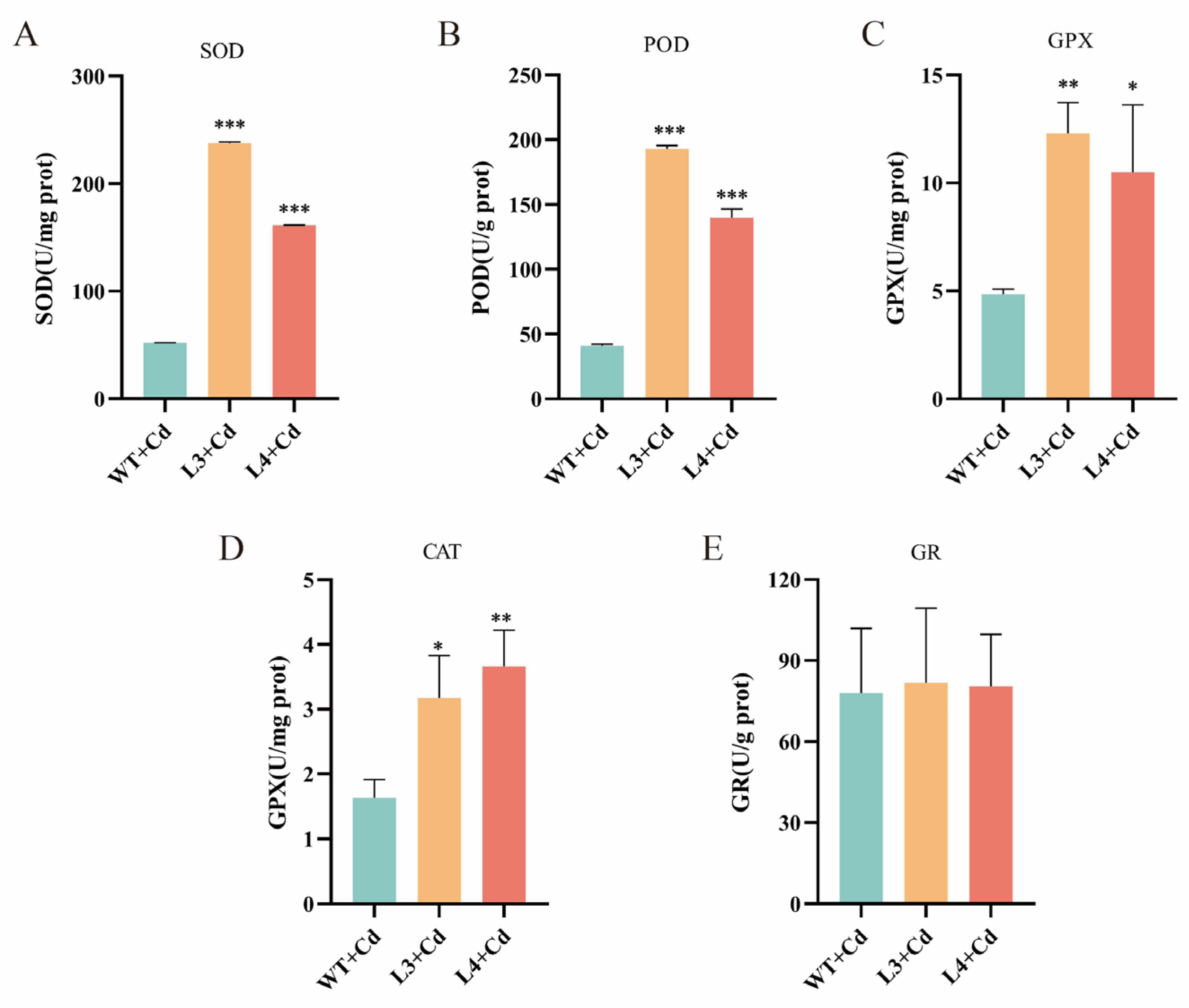
3.7. Impact of NtGPX8a Overexpression on Antioxidant Enzyme Activities
4. Discussion
4.1. NtGPX8a Is the Primary Member of the NtGPX Family in Response to Cd2+ Stress
4.2. NtGPX8a Selects Special Thioredoxins as the Electron Donor
4.3. Overexpression of NtGPX8a Alleviates Cd-Induced Oxidative Stress in Transgenic Tobacco
4.4. NtGPX8a Could Serve as a Candidate Gene for Engineering a High-Tolerance Cd Hyperaccumulator
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carvalho, M.E.A.; Castro, P.R.C.; Azevedo, R.A. Hormesis in Plants under Cd Exposure: From Toxic to Beneficial Element? J. Hazard. Mater. 2020, 384, 121434. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hou, H.; Zhao, L.; Sun, Z.; Lu, Y.; Li, H. Mitigation of Cd Accumulation in Rice from Cd-Contaminated Paddy Soil by Foliar Dressing of S and P. Sci. Total Environ. 2019, 690, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Flores-Cáceres, M.L.; Ortega-Villasante, C.; Carril, P.; Sobrino-Plata, J.; Hernández, L.E. The Early Oxidative Stress Induced by Mercury and Cadmium Is Modulated by Ethylene in Medicago sativa Seedlings. Antioxidants 2023, 12, 551. [Google Scholar] [CrossRef] [PubMed]
- Żabka, A.; Winnicki, K.; Polit, J.T.; Wróblewski, M.; Maszewski, J. Cadmium (II)-Induced Oxidative Stress Results in Replication Stress and Epigenetic Modifications in Root Meristem Cell Nuclei of Vicia Faba. Cells 2021, 10, 640. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, A.; Plusquin, M.; Remans, T.; Jozefczak, M.; Keunen, E.; Gielen, H.; Opdenakker, K.; Nair, A.R.; Munters, E.; Artois, T.J.; et al. Cadmium Stress: An Oxidative Challenge. Biometals 2010, 23, 927–940. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fang, J.; Leonard, S.S.; Krishna Rao, K.M. Cadmium Inhibits the Electron Transfer Chain and Induces Reactive Oxygen Species. Free Radic. Biol. Med. 2004, 36, 1434–1443. [Google Scholar] [CrossRef]
- Garnier, L.; Simon-Plas, F.; Thuleau, P.; Agnel, J.; Blein, J.; Ranjeva, R.; Montillet, J. Cadmium Affects Tobacco Cells by a Series of Three Waves of Reactive Oxygen Species That Contribute to Cytotoxicity. Plant Cell Environ. 2006, 29, 1956–1969. [Google Scholar] [CrossRef]
- Gupta, D.K.; Pena, L.B.; Romero-Puertas, M.C.; Hernández, A.; Inouhe, M.; Sandalio, L.M. NADPH Oxidases Differentially Regulate ROS Metabolism and Nutrient Uptake under Cadmium Toxicity. Plant Cell Environ. 2017, 40, 509–526. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, P.; Salacinski, H.J. Evidence That the Reactions of Cadmium in the Presence of Metallothionein Can Produce Hydroxyl Radicals. Arch. Toxicol. 1998, 72, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Henkler, F.; Brinkmann, J.; Luch, A. The Role of Oxidative Stress in Carcinogenesis Induced by Metals and Xenobiotics. Cancers 2010, 2, 376–396. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Kafle, A.; Timilsina, A.; Gautam, A.; Adhikari, K.; Bhattarai, A.; Aryal, N. Phytoremediation: Mechanisms, Plant Selection and Enhancement by Natural and Synthetic Agents. Environ. Adv. 2022, 8, 100203. [Google Scholar] [CrossRef]
- Jacob, J.M.; Karthik, C.; Saratale, R.G.; Kumar, S.S.; Prabakar, D.; Kadirvelu, K.; Pugazhendhi, A. Biological Approaches to Tackle Heavy Metal Pollution: A Survey of Literature. J. Environ. Manag. 2018, 217, 56–70. [Google Scholar] [CrossRef]
- Li, G.; Yan, L.; Chen, X.; Lam, S.S.; Rinklebe, J.; Yu, Q.; Yang, Y.; Peng, W.; Sonne, C. Phytoremediation of Cadmium from Soil, Air and Water. Chemosphere 2023, 320, 138058. [Google Scholar] [CrossRef]
- Soubasakou, G.; Cavoura, O.; Damikouka, I. Phytoremediation of Cadmium-Contaminated Soils: A Review of New Cadmium Hyperaccumulators and Factors Affecting Their Efficiency. Bull. Environ. Contam. Toxicol. 2022, 109, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First Line Defence Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role in the Entire Antioxidant Defence Grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent Developments in Enzymatic Antioxidant Defence Mechanism in Plants with Special Reference to Abiotic Stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef]
- Pan, C.; Lu, H.; Yu, J.; Liu, J.; Liu, Y.; Yan, C. Identification of Cadmium-Responsive Kandelia obovata SOD Family Genes and Response to Cd Toxicity. Environ. Exp. Bot. 2019, 162, 230–238. [Google Scholar] [CrossRef]
- Sarcheshmeh, M.K.; Abedi, A.; Aalami, A. Genome-Wide Survey of Catalase Genes in Brassica rapa, Brassica oleracea, and Brassica napus: Identification, Characterization, Molecular Evolution, and Expression Profiling of BnCATs in Response to Salt and Cadmium Stress. Protoplasma 2023, 260, 899–917. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Tang, Y.; Liu, C.; Du, X.; Miao, C.; Shi, G. Comparative Transcriptomic Analysis Reveals the Roles of ROS Scavenging Genes in Response to Cadmium in Two Pak Choi Cultivars. Sci. Rep. 2017, 7, 9217. [Google Scholar] [CrossRef] [PubMed]
- Huo, C.; He, L.; Yu, T.; Ji, X.; Li, R.; Zhu, S.; Zhang, F.; Xie, H.; Liu, W. The Superoxide Dismutase Gene Family in Nicotiana tabacum: Genome-Wide Identification, Characterization, Expression Profiling and Functional Analysis in Response to Heavy Metal Stress. Front. Plant Sci. 2022, 13, 904105. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Shih, C.; Huang, C.; Lin, C.; Chou, C.; Tsai, M.; Liu, T.P.; Chiu, J.; Chen, C. Effects of Cadmium on Structure and Enzymatic Activity of Cu,Zn-SOD and Oxidative Status in Neural Cells. J. Cell. Biochem. 2006, 98, 577–589. [Google Scholar] [CrossRef]
- Casalino, E.; Calzaretti, G.; Sblano, C.; Landriscina, C. Molecular Inhibitory Mechanisms of Antioxidant Enzymes in Rat Liver and Kidney by Cadmium. Toxicology 2002, 179, 37–50. [Google Scholar] [CrossRef]
- Ingold, I.; Berndt, C.; Schmitt, S.; Doll, S.; Poschmann, G.; Buday, K.; Roveri, A.; Peng, X.; Porto Freitas, F.; Seibt, T.; et al. Selenium Utilization by GPX4 Is Required to Prevent Hydroperoxide-Induced Ferroptosis. Cell 2018, 172, 409–422.e21. [Google Scholar] [CrossRef] [PubMed]
- Gaschler, M.M.; Andia, A.A.; Liu, H.; Csuka, J.M.; Hurlocker, B.; Vaiana, C.A.; Heindel, D.W.; Zuckerman, D.S.; Bos, P.H.; Reznik, E.; et al. FINO2 Initiates Ferroptosis through GPX4 Inactivation and Iron Oxidation. Nat. Chem. Biol. 2018, 14, 507–515. [Google Scholar] [CrossRef]
- Jung, B.G.; Lee, K.O.; Lee, S.S.; Chi, Y.H.; Jang, H.H.; Kang, S.S.; Lee, K.; Lim, D.; Yoon, S.C.; Yun, D.-J.; et al. A Chinese Cabbage cDNA with High Sequence Identity to Phospholipid Hydroperoxide Glutathione Peroxidases Encodes a Novel Isoform of Thioredoxin-Dependent Peroxidase. J. Biol. Chem. 2002, 277, 12572–12578. [Google Scholar] [CrossRef]
- Navrot, N.; Collin, V.; Gualberto, J.; Gelhaye, E.; Hirasawa, M.; Rey, P.; Knaff, D.B.; Issakidis, E.; Jacquot, J.-P.; Rouhier, N. Plant Glutathione Peroxidases Are Functional Peroxiredoxins Distributed in Several Subcellular Compartments and Regulated during Biotic and Abiotic Stresses. Plant Physiol. 2006, 142, 1364–1379. [Google Scholar] [CrossRef] [PubMed]
- Koh, C.S.; Didierjean, C.; Navrot, N.; Panjikar, S.; Mulliert, G.; Rouhier, N.; Jacquot, J.-P.; Aubry, A.; Shawkataly, O.; Corbier, C. Crystal Structures of a Poplar Thioredoxin Peroxidase That Exhibits the Structure of Glutathione Peroxidases: Insights into Redox-Driven Conformational Changes. J. Mol. Biol. 2007, 370, 512–529. [Google Scholar] [CrossRef] [PubMed]
- Islam, T.; Manna, M.; Reddy, M.K. Glutathione Peroxidase of Pennisetum glaucum (PgGPx) Is a Functional Cd2+ Dependent Peroxiredoxin That Enhances Tolerance against Salinity and Drought Stress. PLoS ONE 2015, 10, e0143344. [Google Scholar] [CrossRef]
- Li, W.; Huai, X.; Li, P.; Raza, A.; Mubarik, M.S.; Habib, M.; Fiaz, S.; Zhang, B.; Pan, J.; Khan, R.S.A. Genome-Wide Characterization of Glutathione Peroxidase (GPX) Gene Family in Rapeseed (Brassica napus L.) Revealed Their Role in Multiple Abiotic Stress Response and Hormone Signaling. Antioxidants 2021, 10, 1481. [Google Scholar] [CrossRef]
- Zhou, Y.; Hu, L.; Ye, S.; Jiang, L.; Liu, S. Genome-Wide Identification of Glutathione Peroxidase (GPX) Gene Family and Their Response to Abiotic Stress in Cucumber. 3 Biotech 2018, 8, 159. [Google Scholar] [CrossRef]
- Gao, F.; Chen, J.; Ma, T.; Li, H.; Wang, N.; Li, Z.; Zhang, Z.; Zhou, Y. The Glutathione Peroxidase Gene Family in Thellungiella salsuginea: Genome-Wide Identification, Classification, and Gene and Protein Expression Analysis under Stress Conditions. Int. J. Mol. Sci. 2014, 15, 3319–3335. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, S.; Sui, X.; Wang, J.; Geng, Y.; Gao, F.; Zhou, Y. Genome-Wide Characterization, Evolution, and Expression Analysis of the Ascorbate Peroxidase and Glutathione Peroxidase Gene Families in Response to Cold and Osmotic Stress in Ammopiptanthus nanus. J. Plant Growth Regul. 2023, 42, 502–522. [Google Scholar] [CrossRef]
- Liu, H.; Wang, H.; Ma, Y.; Wang, H.; Shi, Y. Role of Transpiration and Metabolism in Translocation and Accumulation of Cadmium in Tobacco Plants (Nicotiana tabacum L.). Chemosphere 2016, 144, 1960–1965. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.Z.U.; Rizwan, M.; Sohail, M.I.; Ali, S.; Waris, A.A.; Khalid, H.; Naeem, A.; Ahmad, H.R.; Rauf, A. Opportunities and Challenges in the Remediation of Metal-Contaminated Soils by Using Tobacco (Nicotiana tabacum L.): A Critical Review. Environ. Sci. Pollut. Res. 2019, 26, 18053–18070. [Google Scholar] [CrossRef]
- He, L.; Peng, X.; Cao, H.; Yang, K.; Xiang, L.; Li, R.; Zhang, F.; Liu, W. The NtSPL Gene Family in Nicotiana tabacum: Genome-Wide Investigation and Expression Analysis in Response to Cadmium Stress. Genes 2023, 14, 183. [Google Scholar] [CrossRef] [PubMed]
- Milla, M.A.R.; Maurer, A.; Huete, A.R.; Gustafson, J.P. Glutathione Peroxidase Genes in Arabidopsis Are Ubiquitous and Regulated by Abiotic Stresses through Diverse Signaling Pathways. Plant J. 2003, 36, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Islam, T.; Manna, M.; Kaul, T.; Pandey, S.; Reddy, C.S.; Reddy, M.K. Genome-Wide Dissection of Arabidopsis and Rice for the Identification and Expression Analysis of Glutathione Peroxidases Reveals Their Stress-Specific and Overlapping Response Patterns. Plant Mol. Biol. Rep. 2015, 33, 1413–1427. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, M.; Yu, D.; Teng, Y.; Wei, T.; Chen, C.; Song, W. Identification of Glutathione Peroxidase (GPX) Gene Family in Rhodiola crenulata and Gene Expression Analysis under Stress Conditions. Int. J. Mol. Sci. 2018, 19, 3329. [Google Scholar] [CrossRef]
- Liu, W.; Huo, C.; He, L.; Ji, X.; Yu, T.; Yuan, J.; Zhou, Z.; Song, L.; Yu, Q.; Chen, J.; et al. The NtNRAMP1 Transporter Is Involved in Cadmium and Iron Transport in Tobacco (Nicotiana tabacum). Plant Physiol. Biochem. 2022, 173, 59–67. [Google Scholar] [CrossRef]
- López-Hidalgo, C.; Meijón, M.; Lamelas, L.; Valledor, L. The Rainbow Protocol: A Sequential Method for Quantifying Pigments, Sugars, Free Amino Acids, Phenolics, Flavonoids and MDA from a Small Amount of Sample. Plant Cell Environ. 2021, 44, 1977–1986. [Google Scholar] [CrossRef]
- Margis, R.; Dunand, C.; Teixeira, F.K.; Margis-Pinheiro, M. Glutathione Peroxidase Family—An Evolutionary Overview. FEBS J. 2008, 275, 3959–3970. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive Oxygen Species (ROS) and Response of Antioxidants as ROS-Scavengers during Environmental Stress in Plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Xue, Y.; Chen, L.; Li, B.; Xiao, J.; Wang, H.; Dong, C.; Li, X.; Xu, P. Genome-Wide Mining of GPX Gene Family Provides New Insights into Cadmium Stress Responses in Common Carp (Cyprinus carpio). Gene 2022, 821, 146291. [Google Scholar] [CrossRef]
- Song, S.; Han, Y.; Zhang, Y.; Ma, H.; Zhang, L.; Huo, J.; Wang, P.; Liang, M.; Gao, M. Protective Role of Citric Acid against Oxidative Stress Induced by Heavy Metals in Caenorhabditis elegans. Environ. Sci. Pollut. Res. 2019, 26, 36820–36831. [Google Scholar] [CrossRef]
- Bela, K.; Riyazuddin, R.; Horváth, E.; Hurton, Á.; Gallé, Á.; Takács, Z.; Zsigmond, L.; Szabados, L.; Tari, I.; Csiszár, J. Comprehensive Analysis of Antioxidant Mechanisms in Arabidopsis Glutathione Peroxidase-like Mutants under Salt- and Osmotic Stress Reveals Organ-Specific Significance of the AtGPXL’s Activities. Environ. Exp. Bot. 2018, 150, 127–140. [Google Scholar] [CrossRef]
- Wang, X.; Fang, G.; Yang, J.; Li, Y. A Thioredoxin-Dependent Glutathione Peroxidase (OsGPX5) Is Required for Rice Normal Development and Salt Stress Tolerance. Plant Mol. Biol. Rep. 2017, 35, 333–342. [Google Scholar] [CrossRef]
- Casatejada, A.; Puerto-Galán, L.; Pérez-Ruiz, J.M.; Cejudo, F.J. The Contribution of Glutathione Peroxidases to Chloroplast Redox Homeostasis in Arabidopsis. Redox Biol. 2023, 63, 102731. [Google Scholar] [CrossRef]
- Björnstedt, M.; Xue, J.; Huang, W.; Akesson, B.; Holmgren, A. The Thioredoxin and Glutaredoxin Systems Are Efficient Electron Donors to Human Plasma Glutathione Peroxidase. J. Biol. Chem. 1994, 269, 29382–29384. [Google Scholar] [CrossRef] [PubMed]
- Arnér, E.S.J.; Holmgren, A. Physiological Functions of Thioredoxin and Thioredoxin Reductase. Eur. J. Biochem. 2000, 267, 6102–6109. [Google Scholar] [CrossRef] [PubMed]
- Mata-Pérez, C.; Spoel, S.H. Thioredoxin-Mediated Redox Signalling in Plant Immunity. Plant Sci. 2019, 279, 27–33. [Google Scholar] [CrossRef]
- Chibani, K.; Pucker, B.; Dietz, K.; Cavanagh, A. Genome-wide Analysis and Transcriptional Regulation of the Typical and Atypical Thioredoxins in Arabidopsis thaliana. FEBS Lett. 2021, 595, 2715–2730. [Google Scholar] [CrossRef]
- Fernandez-Calvino, L.; Faulkner, C.; Walshaw, J.; Saalbach, G.; Bayer, E.; Benitez-Alfonso, Y.; Maule, A. Arabidopsis Plasmodesmal Proteome. PLoS ONE 2011, 6, e18880. [Google Scholar] [CrossRef]
- Meng, L.; Wong, J.H.; Feldman, L.J.; Lemaux, P.G.; Buchanan, B.B. A Membrane-Associated Thioredoxin Required for Plant Growth Moves from Cell to Cell, Suggestive of a Role in Intercellular Communication. Proc. Natl. Acad. Sci. USA 2010, 107, 3900–3905. [Google Scholar] [CrossRef]
- Liu, W.; Yu, T.; Cao, H.; Peng, X.; Yang, Y.; Li, R. Effects of EDTA and Organic Acids on Physiological Processes, Gene Expression Levels, and Cadmium Accumulation in Solanum nigrum Under Cadmium Stress. J. Soil Sci. Plant Nutr. 2023, 23, 3823–3833. [Google Scholar] [CrossRef]
- Chen, S.; Wang, Q.; Lu, H.; Li, J.; Yang, D.; Liu, J.; Yan, C. Phenolic Metabolism and Related Heavy Metal Tolerance Mechanism in Kandelia obovata under Cd and Zn Stress. Ecotoxicol. Environ. Saf. 2019, 169, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Kaur, S.; Bhardwaj, R. Physiological and Biochemical Changes in Brassica juncea Plants under Cd-Induced Stress. Biomed Res. Int. 2014, 2014, 726070. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.J.; Shao, G.; Zhang, G. Influence of Cadmium Toxicity on Growth and Antioxidant Enzyme Activity in Rice Cultivars with Different Grain Cadmium Accumulation. J. Plant Nutr. 2005, 28, 1259–1270. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Satheesh, N.; Kherawat, B.S.; Kumar, A.; Kim, H.-U.; Chung, S.-M.; Kumar, M. Regulation of Reactive Oxygen Species during Salt Stress in Plants and Their Crosstalk with Other Signaling Molecules—Current Perspectives and Future Directions. Plants 2023, 12, 864. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Li, H. Cost-Effectiveness Analysis for Soil Heavy Metal Contamination Treatments. Water Air Soil Pollut. 2018, 229, 126. [Google Scholar] [CrossRef]
- Ghuge, S.A.; Nikalje, G.C.; Kadam, U.S.; Suprasanna, P.; Hong, J.C. Comprehensive Mechanisms of Heavy Metal Toxicity in Plants, Detoxification, and Remediation. J. Hazard. Mater. 2023, 450, 131039. [Google Scholar] [CrossRef] [PubMed]
- Unsal, V.; Dalkiran, T.; Çiçek, M.; Kölükçü, E. The Role of Natural Antioxidants Against Reactive Oxygen Species Produced by Cadmium Toxicity: A Review. Adv. Pharm. Bull. 2020, 10, 184–202. [Google Scholar] [CrossRef]
- Đukić-Ćosić, D.; Baralić, K.; Javorac, D.; Djordjevic, A.B.; Bulat, Z. An Overview of Molecular Mechanisms in Cadmium Toxicity. Curr. Opin. Toxicol. 2020, 19, 56–62. [Google Scholar] [CrossRef]
- Nouairi, I.; Ben Ammar, W.; Ben Youssef, N.; Ben Miled, D.D.; Ghorbal, M.H.; Zarrouk, M. Antioxidant Defense System in Leaves of Indian Mustard (Brassica juncea) and Rape (Brassica napus) under Cadmium Stress. Acta Physiol. Plant. 2009, 31, 237–247. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, X.; Ma, T.; Song, K.; Ji, X.; Xiang, L.; Chen, N.; Zu, R.; Xu, W.; Zhu, S.; Liu, W. Overexpression of NtGPX8a Improved Cadmium Accumulation and Tolerance in Tobacco (Nicotiana tabacum L.). Genes 2024, 15, 366. https://doi.org/10.3390/genes15030366
Peng X, Ma T, Song K, Ji X, Xiang L, Chen N, Zu R, Xu W, Zhu S, Liu W. Overexpression of NtGPX8a Improved Cadmium Accumulation and Tolerance in Tobacco (Nicotiana tabacum L.). Genes. 2024; 15(3):366. https://doi.org/10.3390/genes15030366
Chicago/Turabian StylePeng, Xiang, Tengfei Ma, Kejin Song, Xue Ji, Lien Xiang, Nan Chen, Ronglei Zu, Wenyi Xu, Shunqin Zhu, and Wanhong Liu. 2024. "Overexpression of NtGPX8a Improved Cadmium Accumulation and Tolerance in Tobacco (Nicotiana tabacum L.)" Genes 15, no. 3: 366. https://doi.org/10.3390/genes15030366